
5
Biological Dosimetry at RERF
Unlike DS86, which provides a calculated estimate of a person’s organ dose, biological dosimetry estimates dose by evaluating a marker left in the tissue by radiation exposure. The marker can be any of a number of signatures or can be evidence of radiation damage itself. These markers are, in general, a function of the magnitude of an exposure, and they have for some time offered the promise of specific dose determination for each person exposed to ionizing radiation.
Efforts to determine biological doses of survivors in Hiroshima and Nagasaki began at RERF in 1968, shortly after cytogenetic dosimetry was first shown to be useful in radiation-dose reconstruction. However, not all markers of radiation exposure persist; the body repairs or deletes damage that is used as an indicator of exposure. For instance, the most reliable and best documented bioindicator of radiation exposure—the number of dicentrics in peripheral lymphocytes—persists for only about 3 years or less; although dicentrics are excellent for recent accidents, they are not useful for dose reconstruction in the Japanese survivors more than 50 years later. Therefore, biological dosimetry at RERF has had to turn to stable aberrations or other markers that persist for a lifetime. Until very recently, the assay of such persistent markers has been cumbersome and time-consuming, and this has resulted in biodoses for only a small fraction of the people in the Life Span Study (LSS) cohort.
Efforts to find faster assays and more reliable biomarkers of dose have led to RERF attempts to do work by conventional staining analysis and G-banding for cytogenetic aberrations, glycophorin-A assays, electron-spin resonance (ESR) of calcium-tissue samples, and fluorescence in situ hybridization (FISH) for the detection of reciprocal translocations in human chromosomes (See Table 5–1).
Table 5–1 summarizes a number of markers that have been used in biological dosimetry and compares their attributes. Most of these methods have been explored
TABLE 5–1 Comparison of Biomarker Attributes (Straume and Lucas 1995)
|
Biomarker |
Human in vivo |
in vitro |
Animal Model |
Inter-Person Variation |
Persistence Post-Exposure |
|
Translocationsa |
Yes |
Yes |
Yes |
Low |
0-lifetime |
|
Dicentrics |
Yes |
Yes |
Yes |
Low |
0–6 mos. |
|
Micronuclei |
Yes |
Yes |
Yes |
High |
0–6 mos. |
|
HPRTb |
Yes |
Yes |
Yes |
Medium |
1 mo.−1 yr. |
|
GPAc |
50% |
No |
No |
High |
6 mo.-lifetime |
|
TCRd |
Yes |
No |
No |
High |
1 mo.−2yrs. |
|
HLAe |
50% |
Yes |
No |
? |
1 mo.−1 yr. |
|
SCEsf |
Yes |
Yes |
Yes |
? |
0–6 mos. |
|
DNA Adducts |
Yes |
Yes |
Yes |
? |
0–6 mos. |
|
Protein Adducts |
Yes |
Yes |
Yes |
? |
0–6 mos. |
|
aReciprocal chromosome translocations. bHypoxanthine phosphoribosyltransferase assay. cGlycophorin-A somatic mutation assay. dT-cell antigen receptor mutation assay. eHuman leukocyte antigen mutation assay. fSister chromatid exchanges. |
|||||
at RERF and for various reasons, some obvious, many have not been suitable for the type of dose determination required.
Despite the promise of FISH and other new assays to produce the assay speed that would allow biodose determination for large numbers of survivors in the RERF study cohort, fewer than 4000 of the over 86,000 in the LSS population currently have biologically-determined doses. Most of these individual doses are derived from the very labor-intensive work with G-banding or from ESR. In addition to the biomarkers compared in Table 5–1, individual exposures to x or gamma radiation can be determined by using electron spin resonance to detect the physical signature left in tooth enamel by irradiation. The ESR signal is proportional to dose between 0.1 and 10 Gy and is independent of photon energy above 200 KeV. As such, ESR signals obtained from the teeth of survivors are good indicators of the doses from the high gamma ray energies released by the A-bombs. This technique has been used successfully to reconstruct dose in some radiation accidents (Pass and others 1997) and at RERF for a limited number of survivors (about 60). The comparison in Figure 5–1 of the dose derived from ESR and chromosome translocations for the same Japanese survivors shows excellent agreement within the uncertainties of these types of assay.
Given the small numbers of biodoses available and the labor currently involved in producing such individual doses, biological dosimetry is not currently a viable alternative to an overall dosimetry system that produces a dose estimate for all of the members of the LSS. Although only a small number of biologically-derived doses for people in the LSS cohort have been determined, these estimates
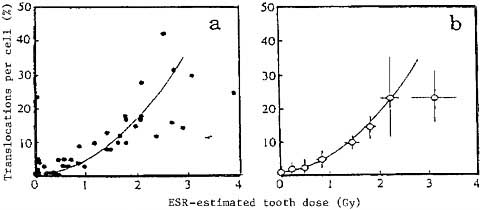
FIGURE 5–1 Translocation frequency of lymphocytes from 41 tooth donors measured by conventional Giemsa staining plotted against ESR-estimated gamma-ray dose for lingual portions of molars, (a) Individual data; (b) grouped data (each point consists of five individuals) (Nakamura and others 1998).
are important in finding out whether there are problems with DS86. As an independent individual dose assessment, biological dosimetry can help to characterize problems and suggest work that must be done to make any successor dosimetry system an accepted source of dose information. For example, areas where biological indicators of radiation dose and the doses derived from DS86 disagree can be looked for. Comparisons of specifically selected subsets of the LSS cohort can yield important evidence to support or refute discrepancies between DS86 and in situ physical activation measurements. Analysis of the trends in such comparisons can be vital clues as to the nature and direction of discrepancies. Two issues of DS86 accuracy in predicting dose are addressed with biological data: the possible underestimation of neutron doses in Hiroshima and the possible overestimation of gamma doses to workers in the Nagasaki torpedo factories.
Differences Between Hiroshima and Nagasaki
The first source of biological data, which has been interpreted by some as evidence of underestimation of neutron dose in Hiroshima, comes first from epidemiology, and second from biological dosimetry. The most recent analysis of cancer risk data from the LSS cohort suggests that excess cancer rates are higher in Hiroshima than Nagasaki (Pierce and others 1996). Interpretation of that observation is a contentious issue, and it can be attributed to various factors. Some have pointed to it as evidence that the more biologically injurious neutrons are the reason for a higher excess rate in Hiroshima than in Nagasaki, where the neutron doses are generally conceded to have been lower (see Chapter 7 for a discussion of this issue). The epidemiological observation is consistent with the biological dosimetry for the
two cities. Stram and others (1993) reported a greater number of stable chromosomal aberrations in the lymphocytes of members of the LSS cohort in Hiroshima than for the same DS86-estimated dose in Nagasaki. Analyses and explanations of these findings have looked to differences in the populations of the cities and to overall underestimation of the Hiroshima dose as possible explanations, but they have not been able to rule out the possibility that the difference is attributable to an underestimation of the neutrons in Hiroshima DS86 doses.
Recent analysis of stable chromosome aberration data obtained from Geimsastained cultures from approximately 3000 survivors in Hiroshima and Nagasaki confirms there is a statistically significant difference in the number of chromosome aberrations seen in the two cities at any given dose (Nakamura and Preston 2000). Survivors in Hiroshima had an average of 6.6% aberrant cells per sievert of exposure; whereas Nagasaki survivors had 3.7% aberrant cells per sievert (see Figure 5–2).
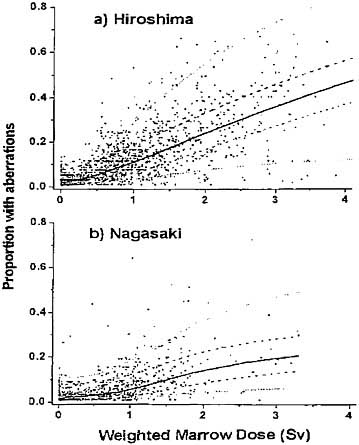
FIGURE 5–2 Scoring efficiency-adjusted proportion of cells with aberrations versus dose. The solid curve is a nonparametric estimate of the dose-response function (Kodama and others 2001).
The Nagasaki Factory Workers
A comparison of aberrations reveals inconsistencies between the number of aberrations and the shielding calculated for survivors in DS86. This is especially true for people located in the large metal torpedo factories in Nagasaki. These survivors had lower numbers of aberrations than individuals exposed to the same dose calculated by DS86 in Nagasaki houses, suggesting that DS86 over-estimates the doses to these people by approximately 40%. This observation agrees with previous ESR work based on the examination of tooth enamel.
When gamma-ray doses for the workers in the Nagasaki torpedo factories are determined with ESR (Nakamura 1999, 2000), they are 40–60% less than those estimated by DS86 (Preston 1999). The fundamental difference between this cluster of large sheet-metal factories filled with heavy machinery and the typical DS86 radiation-shielding case—a wood frame residence—has led to concern that DS86 does not adequately account for the shielding of the factory workers. If that is the case, it causes the current dosimetry system to overestimate the gamma-ray dose received by the roughly 800 workers in the factories. This does not seem like a large percentage of the 86,000 people in the LSS cohort, but such an overestimation becomes important when one recognizes that the 800 are about 40% of the survivors who received 0.5–2.0 Sv in Nagasaki. They are a large percentage of the moderate to high-dose survivors in Nagasaki and are therefore crucial to any accurate risk analysis that involves the Nagasaki data. Thus, it seems vital that any future work to upgrade the dosimetry system for RERF contain a state-of-the-art method of handling the shielding in complex structures. This matter is presently under investigation. Obviously, without biological dosimetry studies on these workers as a source of independent dose determination, the magnitude and direction of the possible miscalculation of the doses to the factory workers would not have been seen.
The limited amount of biological dosimetry that is available for the survivors in the RERF cohort has proven to be extremely useful. Biodoses have served to confirm a potentially significant difference between the two cities, which if substantiated could provide valuable information about the effect of the different qualities of radiation in the two cities (see Chapter 7). ESR and stable chromosome aberrations have highlighted important potential inadequacies in the shielding models currently used in DS86. Given the vital role that biological dosimetry played in this specific case, it is prudent to expand the number of cases in which it is used. Techniques like FISH and ESR that are in place at RERF could be extremely useful in providing an independent dose assessment for complex and uncertain shielding configurations. With careful controls for background aberration rates, FISH could potentially resolve the persistent differences in aberrations and effects observed for the two cities. ESR is equally valuable in cases where the gamma dose needs independent corroboration. Biodosimetry cannot currently replace the dose calculated for the entire LSS; but it does provide an invaluable source of dose evaluation for testing the integrity of DS86 or any subsequent dosimetry system.





