
CHAPTER TWO
Adult Stem Cells
HEMATOPOIETIC STEM CELLS
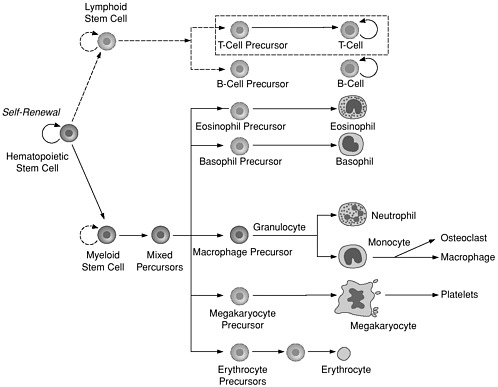
The hope that many diseases can someday be treated with stem cell therapy is inspired by the historical success of bone marrow transplants in increasing the survival of patients with leukemia and other cancers, inherited blood disorders, and diseases of the immune system (Thomas and Blume, 1999). Nearly 40 years ago, the cell type responsible for those successes was identified as the hematopoietic stem cell (Till and McCullough, 1961). The ability of hematopoietic stem cells (HSCs) to self-renew continuously in the marrow and to differentiate into the full complement of cell types found in blood qualifies them as the premier adult stem cells (Figure 3).
HSCs are among the few stem cells to be isolated in adult humans. They reside in the bone marrow and under some conditions migrate to other tissues through the blood. HSCs are also normally found in the fetal liver and spleen and in umbilical cord and placenta blood.
There is a growing body of evidence that HSCs are plastic—that, at least under some circumstances, they are able to participate in the generation of tissues other than those of the blood system. A few studies have shown that HSCs can give rise to liver cells (Lagasse et al., 2000; Taniguchi et al., 1996; Thiese et al., 2001). Those findings have scientists speculating about the biological response of HSCs to disease or tissue damage and about the early differentiation of the
FIGURE 3
Blood Cell Differentiation from Hematopoietic Stem Cells (HSCs). HSCs normally divide to generate either more HSCs (self-renewal) or progenitor cells, which are precursors to various blood cell types. HSCs are found mainly in bone marrow, although T cells develop in thymus, and some other cell types develop from blood monocytes. Once HSCs partly differentiate into progenitor cells, further differentiation into one or a few types of blood cell is irreversible. Solid lines indicate known pathways; dash lines indicate pathways about which there is uncertainty.
embryonic tissues into discrete layers. It was unexpected that a component of blood could cross over a developmental separation to form a tissue type that ordinarily has a completely different embryonic origin (Lagasse et al., 2000). The findings noted above and other reports of cardiac and muscle tissue formation after bone marrow transplantation in mice (Bittner et al., 1999; Orlic et al., 2001) and of the development of neuron-like cells from bone marrow (Brazelton et al., 2000; Mezey, et al., 2000) have raised expectations that HSCs will eventually be shown to be able to give rise to multiple cell types from all three germ layers. One study has, in fact, demonstrated that a single HSC transplanted into an irradiated mouse generated not only blood components (from the mesoderm layer of the embryo), but also epithelial cells in the lungs, gut, (endoderm layer) and skin (ectoderm layer) (Krause et al., 2001). If HSCs are truly multipotent, their potential for life-saving regenerative therapies may be considerably expanded in the future.
The full potential of bone marrow transplantation to restore a healthy blood system in every needy patient is currently limited by the unavailability of HSCs in the quantity and purity that are crucial for successful transplantation. Because of their relative rarity (one in every 10,000 bone marrow cells) and the difficulty of separating them from other components of the blood, so-called bone marrow stem cell transplants are generally impure (NIH, 2001). The significance of such impurity is great. All cells of the body express on their surface a set of molecules called histocompatibility (i.e. tissue compatibility) antigens. If a patient receives a transplant of HSC cells from a donor that has histocompatibility antigens different from his own, the patient’s body will recognize and react to the cells as foreign. To increase the likelihood that histocompatibility antigens will match, it is preferred that donors be a related sibling of the transplant recipient. Even if their histocompatibility antigens do match, however, HSC transplants can be contaminated by T cells from the donor’s immune system.
That contamination can cause the recipient’s body to reject the material or can produce an immune reaction in which the T cells of the transplant attack the tissues of the recipient’s body, leading to a poten-
tially lethal condition known as graft versus host disease. Although autologous transplants, in which material from a person is implanted into the same person (for example, when a cancer patient stockpiles his own blood in advance of chemotherapy or irradiation) solve the problem of immune system rejection, the inability to purify the material leads to the risk that diseased or cancerous cells in the transplant will later be reintroduced to the patient along with the stem cells.
In contrast, transplants of highly purified and concentrated populations of HSCs in mice have been shown to greatly reduce the incidence of graft versus host disease (Shizuru et al., 1996; Uchida et al., 1998). Purified and concentrated populations of autologous HSCs transplanted in breast cancer patients after chemotherapy have been shown to engraft more swiftly and with fewer complications (Negrin et al., 2000). Transplants of concentrated HSCs also have been shown to repopulate the blood more readily, reducing the period during which an individual is vulnerable to infection.
There is also evidence that transplants derived from umbilical cord blood are less likely to provoke graft versus host disease, possibly because the cells in cord blood are immature and less reactive immunologically (Laughlin, 2001). The quantity of HSCs present in cord blood and its attached placenta is small, and transplants from cord blood take longer to graft, but for children, whose smaller bodies require fewer HSCs, cord blood transplants are valuable, especially when there is no related sibling to donate HSCs (Gluckman et al, 2001). Banks of frozen umbilical cord and placenta blood (drawn out of the umbilical vein of the cord) are an important source of HSCs because the histocompatibility markers on the cells in these tissues can be identified and catalogued in advance of the need for a transplant.
Irving Weissman, who presented research findings on HSC transplantations at the workshop, has explored ways to improve the identification and purification of HSCs by looking for proteins on the surface of the stem cells that can be closely associated only with HSCs. Finding the specific profile of proteins that identifies HSCs, particularly those called long-term HSCs, is important, because these cells are believed to hold
the key to future HSC therapies. Obtaining purified HSCs is a major challenge, and purification in a clinical setting is expensive and difficult.
Another major barrier to progress in HSC research and transplantation therapy is that it has not been possible to culture HSCs in vitro (outside the body), although recent studies of mouse HSCs grown in combination with components of the bone marrow have offered some preliminary promise (Ema et al., 2000; Moore et al. 1997). This stubborn and not insignificant obstacle is faced by researchers with all types of adult stem cells. If it were possible to expand the numbers of stem cells by growing them in culture or to stimulate their expansion in vivo (in the living body), the prospects for patients in need of stem cell transplants would be significantly improved. However, as Ernest Beutler pointed out at the workshop, finding a way to get HSCs to proliferate is not enough. In the long run, it is necessary to understand not only what activates HSCs to self-renew, but also what controls their decisions to differentiate into the various components of the blood and prevents them from developing into leukemic cells (Saito et al., 2000).
OTHER ADULT STEM CELLS
During the past 2 years, scientific reports of stem cells in other organs of adult mice—including brain, muscle, skin, digestive system, cornea, retina, liver, and pancreas—have cast a new light on the body’s own capability to replenish its tissues (NIH, 2001). Their discovery has also fostered speculation that these cells exist in the adult human, that they have the characteristic of plasticity that enables them to change into precursors of cell types of other tissues, and that they will someday be used to produce the tissues for therapeutic use. The finding of stem cells in adult tissues, not all of which have been confirmed, offers a first glimpse at potential solutions to long-standing puzzles about why some human organs have a greater capacity for self-repair than others.
The idea of employing adult stem cells in certain therapeutic applications is appealing for several reasons. First, adult stem cells are naturally poised to generate a particular tissue, which might consist of several cell
types, so they should be able to give rise to all the components of that tissue when transplanted into a patient. Second, some stem cells are able to migrate to injured tissue or other discrete sites in the body; for example, neural stem cells will migrate to tumor sites in the brain of a rodent (Aboody et al., 2000). This might provide more flexibility in choosing where to transplant them and more predictability in where they will localize after transplantation. Third, some adult stem cells are known to secrete growth factors that mobilize or protect other cells residing in the tissue that could increase the salutary effects of the transplant (Noble, 2000). It also might be possible to genetically engineer adult stem cells to produce higher levels of compounds normally produced in the body, to compensate for some deficiency in a patient’s own tissues. Similarly, the cells could be engineered to secrete a therapeutic agent, such as a drug.
In other situations, the use of adult stem cells would be inappropriate—for example, the isolation and autologous transplantation of a person’s stem cells suffering from a genetic disorder—in that case, the stem cells would carry the same incorrect genetic information. Transplantation of stem cells from a donor into another person will be subject to the problems of immune rejection, and this could be a substantial obstacle in time-critical situations, for example, spinal cord trauma or stroke, because characterizing the patient’s tissues and finding a match in a short period of time will be difficult.
However, because recent findings of adult stem cells are so new and studies of them raise so many questions, even the most preliminary generalizations and conclusions as to therapeutic potential are tentative. As was noted by James Thomson at the workshop, the hematopoietic stem cell is the most characterized cell in the body, and “The amount of knowledge we have on other adult stem cells goes down dramatically from there.”
First, human adult stem cells are rare and it is difficult to isolate a unique group of stem cells in pure form. So it is not surprising that what at first appears to be plasticity in a single adult stem cell type could be the result of a mixture of cells of different types, including different types of stem cells. At the workshop, Margaret Goodell explained how her
research initially suggested that, given the right environment, stem cells from mouse muscle could be shown to produce not only muscle, but also components of blood. Later it became clear on rigorous testing that her sample contained two entirely different kinds of stem cells: one that formed blood and one that formed muscle. As she noted, the fact that the two types of stem cells were found in muscle might have interesting therapeutic uses, but in any case, it has not been demonstrated that a single type of stem cell in muscle exhibits this degree of plasticity.
An issue raised at the workshop was the need for more experiments that can show an unequivocal relationship between a stem cell and the tissues that are claimed to have arisen from it. In such an experiment, (an example of which is Krause’s work on HSCs mentioned earlier in this chapter) a single, isolated stem cell would be chemically treated so that it incorporates a chemical “label” that will be passed on to all the cells that arise from it. If the labeled stem cell is injected into a mouse, any cell or tissue that is eventually found to have the label can be assumed to have come from the original single stem cell, and this is the kind of evidence for a definitive relationship that stem cell researchers are seeking.
A second factor that complicates adult stem cell research is that the environment in which stem cells grow or are placed to grow has an important but poorly understood effect on their fate—a theme that was echoed by many speakers at the workshop. For example, Iqbal Ahmad discussed prospects for retinal regeneration, which occurs naturally in goldfish but not in humans. Ahmad has isolated precursor cells in the mammalian eye that can be grown in culture for short periods and will develop into cells that appear to be retinal photoreceptors. If precursor cells from a mouse eye are transplanted into a normal mouse retina, they are not incorporated. In contrast, when transplanted into a diseased retina, the precursors begin to develop into photoreceptor-like cells and integrate into the tissue. Ahmad has not yet determined whether the integrated cells function normally.
What signals does the diseased retina provide that the normal retina does not? The cellular environment has important implications for how cells behave when they grow in a living organism (in vivo) and for what
happens to them in culture (in vitro). For example, Ron McKay described how precursor cells taken from the mouse midbrain can be cultured in vitro to generate cells that appear to be dopaminergic neurons, but only for a very short time. (Dopaminergic neurons produce the chemical mediator L-dopa and are depleted in patients with Parkinson’s disease.) Recalling that it is well known that the spinal cord generates motor neurons in response to signals that come from other tissues, McKay suggested that the capability of the precursors of the midbrain to make dopaminergic neurons might be transient in culture because they require stimulation from signals present only in the brain.
A third problem in understanding the capabilities of adult stem cells is the relationship of the cellular environment to the concept of plasticity in adult stem cells. Markus Grompe showed that HSCs and pancreatic stem cells can give rise to liver cells called hepatocytes that will repopulate a diseased mouse liver, demonstrating the plasticity of adult stem cells. However, in his experiment, HSCs and pancreatic stem cells were very inefficient in repopulating the liver relative to the ability of transplanted hepatocytes themselves. This could mean that the plasticity of adult stem cells is a marginal capacity that can be exploited only with a much greater understanding of the environmental signals that influence adult stem cells. No one knows what steps HSCs or pancreatic cells go through in generating a hepatocyte, or what signals cause them to migrate to the liver in the first place. Moreover, some of the apparent plasticity in adult stem cells is difficult to interpret because it has been accomplished in abnormal environments, for example, in mice that are immunologically impaired (Mezey et al., 2000) or sublethally irradiated (Brazelton et al., 2000).
Fourth, a major weakness of stem cell research asserted by Grompe is that most studies inadequately demonstrate that stem cells have produced a functionally useful cell in the organ. Most studies showing the plasticity of stem cells rely on the detection of proteins in the newly generated tissues that are commonly associated with a particular type of differentiated cell. But there is no consensus in the scientific community that the detection of a particular protein constitutes sufficient evidence that the
cells and tissues formed are, in fact, fully functional and normal. Olle Lindvall, who works with Parkinson’s disease patients, noted that in some experiments in which dopaminergic neurons generated in culture were grafted into the brain of an animal, it was not at all clear that the new neurons were fully functional. The relationship between stem cell type and environmental cues makes problematic the assumption that stem cells cultured in vitro can be expected to perform with predictable results when transplanted in vivo (Morrison, 2001). It might be possible someday to provide cues to reprogram one cell type into another and even to culture these cells in vitro, but evidence of the normal physiological and restorative function of adult stem cells is very limited today.
A fifth limitation relevant to immediate development of therapies based on adult stem cells is the inability to maintain these cells in culture for very long before they differentiate into their mature progeny. One can envision two therapeutic approaches to stem cells. In the first, stem cells themselves are implanted in a diseased or injured organ in the hope that they will give rise to the mature cells needed by that organ. In the second, the stem cells are stimulated to differentiate into the needed mature tissue outside the body, and that tissue is implanted in the organ. That adult stem cells are difficult to isolate, purify, and culture causes problems for either approach, although even the ability to culture stem cells for a limited time, including in the presence of other cells, could have therapeutic potential. An example is the use of autologous skin grafts for burn patients, in which healthy skin (which contains skin stem cells) is removed from the patient, cultured briefly outside the body, and grafted onto the patient’s injured tissue. The grafts are not able to regenerate hair follicles and sweat glands, but are otherwise able to function normally. However, with a few exceptions, the appropriate culture conditions to sustain most adult stem cells indefinitely have yet to be found.
Very few stem cells, strictly defined, have even been isolated from adult human organs, in part because they constitute only a tiny fraction of the cells present and are not likely to be very distinct from the partially differentiated cells they give rise to as they mature and differentiate. For
example, at the workshop, Fred Gage discussed his work with cadavers and brain biopsy material, wherein he found not stem cells, but rather what might be more mature types of neural cell precursors (Palmer et al., 2001). Those cells would differentiate into various neural tissues but would then stop dividing. Unlike stem cells, precursors and other subsequent intermediates generally undergo limited self-renewal in vivo and are committed to a pathway of differentiation into a specific tissue type. Researchers are, however, beginning to understand how a stem cell gives rise to precursor cells and, in at least one case, have used this information to manipulate that process. Using biochemical signals found in the cellular environment, rodent precursor cells in vitro were caused to revert into more primitive, multipotent stem cells (Kondo and Raff, 2000). The ability to “reprogram” a cell may be exploited someday to therapeutic ends; however, the reversal of the normal pathway of differentiation may have biological consequences not yet detected. Rigorous experimentation will be needed to evaluate the implications of this basic research finding for regenerative medicine.
Finally, the implications of what is known about human adult stem cells are often overlooked amid reports of successes with experiments in rodents that simulate heart attack, retinal disease, and diabetes. Confirmed reports of truly multipotent human adult stem cells are scarce. For its recently released report on stem cells, the National Institutes of Health could find few published accounts of the isolation of multipotent adult stem cells from human tissues (NIH, 2001). The much-publicized recent report of stem cells from human fat that produced cartilage, bone, and muscle (Zuk et al., 2001), for example, did not conclusively establish that the cells capable of performing this feat were fat cells. The authors of the paper conceded that the observation might have been due to the presence of another cell type, such as an HSC that had circulated out of blood and into fat. Without conclusive identification, the existence of a multipotent fat cell remains unconfirmed.
That there is little evidence of a wide array of human adult stem cells that can differentiate into multiple tissue types does not mean that they will not eventually be found, nor should it be interpreted to mean that
the results of experiments with stem cells in rodents are not useful. Those results have prompted new theories about the source and significance of regenerative capabilities in all cells, about the process of cellular differentiation, and about the role of the physiological environment in inducing cells of all kinds to express their different characteristics (Blau et al., 2001).
All somatic cells in an organism contain the same genetic information, but it is not yet known what causes parts of the genetic code to be expressed in some cells and different parts to be expressed in others. This raises important and interesting questions about the ability of a cell of one type to become another type. Emphasizing how little is understood about the process that controls a cell’s commitment to one course of action or another, Ihor Lemischka explained his findings that many genes found to be active in stem cells do not correspond to any known gene function ever described. A comparison of mouse and human HSCs shows that only about half of the genes expressed in the mouse HSCs correspond to genes expressed in human HSCs, so there are going to be differences as we move from experiments with mice to regenerative therapies in humans. Even the genetic programs that control the differentiation of human fetal liver stem cells and human HSCs, both of which evolve into the components of the blood, seem to be very different (Phillips et al., 2000). We need to understand much more about the differences between mouse and human stem cells if we are to harness their potential.












