
3
Effects of Trawling and Dredging

“Habitat alteration by the fishing activities themselves is perhaps the least understood of the important environmental effects of fishing” (National Research Council, 1994).
The use of mobile fishing gear has become a source of concern because of the size of the affected fishing grounds, the modification of the substrate, disturbance of benthic communities, and removal of nontarget species. The long-term viability of some fish populations could be threatened if essential fish habitat is degraded. Also, because of declines in many traditional fisheries, efforts to find under-exploited fish populations have increased interest in exploiting less accessible, previously unfished areas. These efforts have been facilitated by the development of new gear and navigational aids. Extensive new regions of the continental shelf, slope, submarine canyons, and seamounts have been exposed to the effects of bottom trawling and dredging. Expansion of fishing into new territory could lead to the loss of habitats that might have provided as refuge for heavily exploited species.
Since the publication of the 1994 National Research Council report, there has been additional research on the effects of fishing gear, especially trawls and dredges, on marine benthic habitats. The magnitude depends on gear configuration, on the subtle modifications various operators make to their gear and on the many and varied habitats fished. Given the inherent difficulty of studying offshore habitats and the problems associated with determining causation under shifting environmental conditions (current, temperature variation, natural migration, storm activity), not all questions regarding the effects of fishing on the seafloor have been answered—nor are they likely to be in the near future. Evaluating the effects of bottom trawling on benthic communities is complicated by the sparseness of data on species abundance and composition before intensive bottom fishing began. This is important because recent analyses of the few existing historical data sets suggest that larger bodied organisms (fish and benthos) were more prevalent before intensive bottom trawling began (Frid and Clark, 2000; Greenstreet and Hall, 1996). Existing studies necessarily indicate changes relative to recent conditions, not changes relative to the less disturbed ecosystem. There has been an increase in the understanding of fishing gear and habitat interactions that can be used for making decisions about habitat management.
Any fishing gear will affect the flora and fauna of a given location to some degree, but the magnitude and duration of the effect depends on several factors, including gear configuration, towing speed, water depth, and the substrate over which the tow occurs. Variations in substrate include differences in sediment type, bed form (sand waves and ripples, flat mud), and biologic structure (shell, macroalgae, vascular plants, sponges, corals, burrows) (Auster and Langton, 1999). What are the ecologic consequences of these fishing effects? What are the short- and long-term effects on populations, community structure, and interspecific dynamics? Is the disturbance caused by fishing less than what occurs naturally? Are some species threatened with local extinction?
In considering the consequences of trawling and dredging it is important to distinguish between the direct and indirect effects of the activity. Direct effects can be summarized as follows:
-
Mortality. Population mortality occurs either as part of the catch (landings plus discards) or incidentally either by killing benthic and demersal species or making them more vulnerable to scavengers and other predators.
-
Increased food availability. Discarded fish, fish offal, and dead benthic organisms become food for scavenging species.
-
Loss of habitat. Some fishing gears cause the disturbance or destruction of seafloor habitat.
Indirect effects are the downstream consequences of a direct effect. Reductions in the total biomass of target fish, along with the direct effects noted above, could be expected to affect predators, prey, competitors of a target species, and overall seafloor community structure. Indirect effects also encompass potential changes in the flow of materials and energy through ecosystems and shifts in the balance among the processes of primary production, primary consumption, and secondary production.
Human activities such as trawling can be considered a disturbance to environments, and their effects are often compared with natural disturbances that occur in the same ecosystems. It is important to ask whether human disturbances represent selective pressures at novel spatial or temporal scales or are just slight changes in the scale of existing natural disturbance. Natural disturbances can occur with different periodicities, spatial effects, and patterns of recovery (e.g., Lake, 1990; Pickett and White, 1995). Periodic disturbances can be considered pulse events, and a population or community assemblage might respond in several ways. If the disturbance is not too intense, and if the interval between disturbances is long relative to the attributes of the community, or if the system is resilient, the community could return to its previous state. Ecological disturbance theory also suggests that, even if each individual pulse disturbance does not have a large acute effect, there could be a threshold of intensity or a cumulative level beyond which persistent changes in the ecosystem occur. Resilience is the degree to which an ecosystem’s long-standing composition, structure, and function can recover from disturbance (Holling, 1973). The disturbance paradigm predicts that short-lived, highly motile or dispersing species with high reproduction rates will recover from disturbance faster than will long-lived, sessile, low-dispersing species (Pickett and White, 1995).
The following sections are based on the results of previous studies and reviews. They summarize commonly observed effects of fishing on the seafloor with respect to gear type, the nature of the seafloor habitat, frequency of disturbance (natural and from fishing gear), and rates of recovery to the pretrawling or predredging state.
DIRECT EFFECTS ON SPECIES AND HABITAT STRUCTURE
Research Approaches
Studies of the effects of mobile fishing gear on benthic habitat have used observation and experiment. Observational studies compare the benthic habitat in trawled areas with the habitat in lightly trawled or untrawled places. One difficulty with this approach is finding habitats that are similar in all respects other than the degree of fishing. In any given region, benthic areas inhabited by commercial concentrations of fish and shellfish, not closed by regulation, will be trawled or dredged at some frequency. Quantifying how much trawling has occurred in lightly trawled areas can be impractical given the limited scale of benthic studies. It is difficult to assess how much trawling actually occurs in an area solely from effort data collected in most fisheries. A full evaluation of the effects of trawling and dredging on habitat will require higher resolution effort data to translate the results of smallscale experimental studies to effects at the ecosystem level.
Experimental studies generally use the before/after control/impact design. In this approach, an experimental area is trawled and compared before and after trawling (before/after comparison) and with a site that has not been trawled recently (control/impact comparison). This design often involves direct sampling of fauna, video observations, and sonar scans of the control and disturbed sites. The primary limitation of this design is that it is based on the assumption that the control and experimental sites are equivalent. A study by Lindegarth et al. (2000) suggests multiple evaluation sites are needed to assess the effect of trawling on benthic habitat. The authors showed that the interpretation of experimental studies varies depending on the control and treatment sites compared. Although the need for multiple control sites and replicate trawling is acknowledged within the scientific community, application is limited by ship time, funding constraints, and existing and shifting management regimes.
Research Summary
The effects of mobile bottomfishing gear on benthic habitats depend on the susceptibility of the habitat and on the type gear used. Table 3.1 provides examples of observed effects of different gear in various habitats as catalogued in recent literature reviews (e.g., Auster and Langton, 1999; Barnette, 1999; Jennings and Kaiser, 1988). The extensive primary literature, many review articles, and a meta-analysis of 57 published studies (Collie et al., 2000b), reveal several generalities about the response of seafloor communities to trawling and dredging. These generalities, highlighted in bold text, are discussed below.
Trawling and dredging reduce habitat complexity. The direct effects of trawling and dredging include loss of erect and sessile epifauna, smoothing of sedimentary bedforms and reduction of bottom roughness, and removal of taxa that produce structure. Trawl gear can crush, bury, or expose marine flora and fauna and reduce structural diversity (Auster and Langton, 1999). On Florida’s Oculina Banks, for example, trawl fisheries for rock shrimp and trawl and dredge fisheries for calico scallops have been implicated in the reduction of 1–2 m diameter Oculina varicosa tree corals to 2–3cm rubble (Koenig et al., 2000). If the interval between trawls is shorter than the recovery time, the original benthic structure and species populations might not have the opportunity to recover to pretrawl conditions (Watling and Norse, 1998). Most research bears out the paradigm of variable environments inhabited by short-lived species recovering more rapidly than stable
TABLE 3.1 Examples of Mobile Fishing Gear Effects on Habitat (Based on Reviews by Auster and Langton, 1999, and Barnette, 1999)
|
Gear |
SAV |
Sand |
Hardbottom/ Biogenic |
Muddy Sand |
Gravel |
|
Scallop dredge |
Increased dredging resulted in significant reductions in biomass and number of shoots (1) |
Smoothed bedforms; reduction of epifaunal coverage; shell aggregate dispersal (2, 3, 4) |
• Single passage can kill 70% of the living maerl in the dredge path. Flora and megafauna to a depth of 10 cm are damaged. • Dredge tracks remain visible for 2.5 years in maerl habitats. • Maerl is a “living sediment” that is slow to recover from disturbance due to extremely low growth rates (5) |
A gradient of increasing large epifaunal cover correlated with decreasing fishing effort (4) |
• Undredged sites had higher numbers of organisms, biomass, species richness, and species diversity than dredged sites. Undredged sites had bushy epifauna, dredged sites were dominated by hard-shelled mollusks, crabs, and echinoderms (6, 7) • Suspended fine sediment and buried gravel below the sediment water interface (3) • Smoothed bedforms; hydrozoan cover removed; reduced densities of shrimp (2) |
|
Oyster dredge |
Gear modified for clam harvest reduction in coverage; loss of rhizomes; extended recovery time; sediment suspension; smothering of SAV (8) |
|
Reduction in height of oyster reefs, increased susceptibility to hypoxia (9) |
|
|
communities composed of sessile, long-lived species, which sustain longer term damage.
Repeated trawling and dredging result in discernable changes in benthic communities. Many studies report that repeated trawling and dredging causes a shift from communities dominated by species with relatively large adult body size toward dominance by high abundances of small-bodied organisms (Auster et al., 1996; Engel and Kvitek, 1998; Jennings et al., 2001; Kaiser et al., 2000; Kaiser and Spencer, 1996b; Watling and Norse, 1998). Intensively fished areas are likely to remain permanently altered, inhabited by fauna that readapted to frequent physical disturbance. In some habitats these differences will be profound, in others they will be rather subtle (Kenchington et al., 2001). Species richness (the number of species per unit area) and evenness (the relative abundance of resident species)—two measures of species diversity— can decline in response to bottom fishing, but not all communities show reduced diversity. For example, if bottom fishing reduces the abundance of a dominant species, the disturbed community might have higher evenness and hence lower species diversity (Collie et al., 1997). Untrawled, silty habitat in the Aegean Sea had lower species diversity than did similar, trawled, silty habitat. Measurements of species diversity is not always a reliable indicator of disturbance because a change in the structure of the benthic community can increase or decrease overall species diversity.
Bottom trawling reduces the productivity of benthic habitats. It has been hypothesized that the shift to communities of smaller, fast-growing species after removal of larger, slow-growing species by trawling could maintain benthic productivity and support predacious fish. However, Jennings et al. (2001) found a 75 percent reduction in total infaunal productivity between unfished and heavily trawled areas. Although productivity per unit biomass was higher in heavily trawled areas because of the shift to smaller organisms, overall productivity was lower because of the loss of biomass.
The effects of mobile fishing gear are cumulative, and depend on trawling frequency. Repeated trawling (or dredging) can exceed a threshold above which a disturbance can result in observable, long-term ecological effects. Even shallow, high-energy areas that often experience natural disturbances can be affected if the frequency and seasonality of the trawling disturbances are different from those of natural events (e.g., Auster and Langton, 1999). Small-scale fishing disturbances can be masked by larger scale natural events (Kaiser and Spencer, 1996b). A three-year study by Tuck et al. (1998) compared the benthic infauna at sites that were trawled regularly and at untrawled control sites. After five months of trawling, they observed changes only in the relative abundances of different species, but after 16 months total species richness began to decline in the trawled sites. Unfortunately, most research has focused on acute effects, quantifying changes to benthic habitat after only a limited number of trawl passes over a short period. These acute studies do not document long-term changes attributable to repeated trawling and dredging. More long-term studies are needed to assess the full range of consequences in areas that are trawled or dredged regularly.
Fauna that live in low natural disturbance regimes are generally more vulnerable to fishing gear disturbance. According to ecologic disturbance theory, initial responses and rates of recovery from trawling should reflect the stability of the substrate in a particular habitat and the character of the benthic community that it supports (Figure 3.1) (Lake, 1990; Pickett and White, 1995). Habitats consisting of unconsolidated sediments that experience high rates of natural disturbance can have more subtle responses to trawling than will habitats characterized by boulders or pebbles (Tuck et al., 2000; Kenchington et al., 2001). Animals that live in unconsolidated sediments in high natural disturbance regimes are adapted to periodic sediment resuspension and smothering like that caused by mobile bottom gear. In contrast, epifaunal communities that stabilize sediments, reef-forming species, or fauna in habitats that experience low rates of natural disturbance have been observed to be particularly vulnerable. Individual studies support the generalizations summarized in Figure 3.1, but a quantitative meta-analysis was less conclusive (Collie et al., 2000a). Responses in sand habitats were usually less negative than in the other habitats, but a consistent ranking of impacts with respect to a priori expectations by habitat did not emerge. However, the outcome of the meta-analysis could be confounded by limitations in the available data and by interactions among the factors (gear type, habitat type).
Fishing gears can be ranked according to effects on benthic organisms. Intertidal dredging (with gear that causes the direct removal of sediments, shells, and
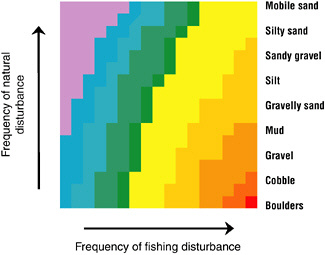
FIGURE 3.1 Conceptual model of fishing disturbance to benthic communities. The response variable is the percent decrease in abundance due to bottom fishing. The response is ranked from lowest (top left) to highest (bottom right). The frequency of natural disturbance corresponds roughly with sediment type, but not directly with particle size. The axes correspond to measurements that should be readily obtainable for most parts of the continental shelf.
rocks) has more marked initial effects than either scallop dredging or intertidal raking, which in turn cause greater damage than beam and otter trawling (Collie et al., 2000a). Otter trawls have been evaluated more often than have other types of gear, because of their widespread use (Barnette, 2001; Collie et al., 2000a). This ranking is consistent with the degree of bottom contact and sediment penetration of the different gears.
Benthic fauna can be ranked according to vulnerability. The most consistent research observation is that vulnerability to mobile gear is predicated on the morphology and behavior of the benthic species. Softbodied, erect, sessile organisms are more vulnerable to mobile gear than are hard-bodied prostrate organisms. Despite limits in the taxonomic resolution of the data, the meta-analysis identified a 68 percent reduction in anemone abundance, as opposed to a 21 percent mean reduction in starfish, after a single trawling event (Collie et al., 2000a). Similarly, chronic exposure (repeated dredging) resulted in a 93 percent reduction for anemones, malacostracan crustaceans, brittle stars, and polychaetes, whereas a single dredge event resulted in a 76 percent reduction. On average, none of these taxa increased in abundance, and the average reductions across taxa amounted to 55 percent (Collie et al., 2000a).
Modeling Mortality in Relation to Fishing Effort
Based on the general principles outlined above, a model can be derived to predict the effects of bottom fishing. The depletion of a nontarget species can be modeled with the exponential equation:
NE = Noe-mE [1]
N0 is initial abundance, and NE is abundance after E passes of a particular kind of fishing gear. The mortality coefficient, m, is analogous to catchability: it
includes the mortality of individuals not captured but still killed by the trawl. The mortality rate depends on factors such as gear, habitat type, and life history. One obvious but important implication of this exponential model is that repeated trawls at the same location kill diminishing numbers of organisms. Hence, if the distribution of the nontarget species is not positively correlated with that of the target species, a more aggregated fishery will inflict a lower mortality rate.
The depletion equation can be normalized to the proportion of animals killed as a function of the number of tows:
[2]
This response variable is bounded between 0 and 1, and larger values correspond to greater hazard or risk. One can envision a family of curves corresponding to different values of each explanatory variable (Figure 3.2). The curves show that species in sandy habitats experience a lower mortality rate than do those in a gravel habitat. In very few cases have the shapes of these mortality curves been systematically measured. Most trawl studies consist of a single disturbance event (1tow) or spatial comparisons of chronically fished and unfished areas (at the asymptote of the curve).
The depletion equation also can be expressed as a linear model of potential explanatory variables:
[3]
Mortality, m, has been expanded as a linear combination of factors (G, gear; H, habitat) or continuous variables (D, depth). This equation provides the basis for estimating the importance of potential explanatory variables and is similar to the response variable used in the meta-analysis of Collie et al. (2000a).
Modifications to Habitat Structure
An important consequence of trawling is the reduction in habitat complexity (architecture) that accompanies the removal of sessile epifauna and the alteration of physical structure, such as rocks and cobble. Emergent epifauna, such as sponges, hydroids, and bryozoans, provide habitat for invertebrates and fishes. Disturbance of emergent epifauna can increase the predation risk for juvenile fish. Decreased prey abundance increases the foraging time for juvenile fish, thus exposing them to higher predation risk (Walters and Juanes, 1993). Laboratory studies (Lindholm et al.,
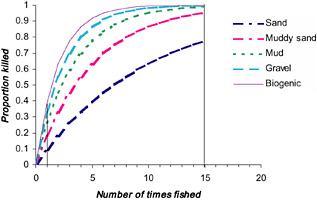
FIGURE 3.2 Hypothetical depletion curves for non-target species in different habitats. The vertical lines indicate that most trawl-impact studies either have been acute (trawled once, vertical line at 1) or compare chronically fished areas (vertical line at 15).
1999) and field studies (Tupper and Boutilier, 1995) have shown that increased epifaunal cover reduces predation risk to juvenile cod.
Information on the linkages between habitat and fish population dynamics is limited; most experimental studies have been conducted in coral reef systems. An extensive literature shows links among larval supply, postsettlement predation, physical attributes of habitat, and adult population size (e.g., Sale, 1991). For example, Sainsbury et al. (1997) provided compelling evidence that loss of structural epibenthos in a tropical system resulted in a shift from a high-value community dominated by lethrinids and lutjanids (emperors and snappers) to a lower value one dominated by saurids and nemipterids (lizard fish and bream). By inference, structurally rich habitats in temperate ecosystems also can support a greater diversity of fish species, but the influence of habitat structure on the productivity of economically important species in temperate and boreal ecosystems has not been determined. Where studies have been conducted, and they have been mostly correlative—results are consistent with the assumption that there are linkages between habitat attributes and fish survivorship (e.g., Auster et al., 1995, 1998; Langton et al., 1995; Stein et al., 1992; Tupper and Boutilier, 1995; Yoklavich et al., 2000).
With repeated trawling, the physical relief of the seafloor could be reduced, with a concomitant decrease in the quality of habitat for some species. Juveniles of many demersal fish species are known to aggregate near seabed structure. In trawled areas of the North Sea, the abundance of larger bodied, long-lived benthic species was depleted more than that of smaller, short-lived species, and there was an overall reduction in benthic production (Jennings et al., 2001). Also, removal of physical structure in a habitat can force some species into less optimal environments. For instance, the dredging of oyster reefs in North Carolina has lowered the reefs’ vertical height relative to the seafloor. Thus, the only suitable substrate for the oysters is closer to the bottom in deeper areas that are more prone to anoxic events that result from nutrient overloading (Lenihan and Peterson, 1998; Lenihan et al., 2001).
The life histories of demersal fishes exhibit a gradient of linkages to habitat attributes, and the degree of habitat affinity varies by life-history stage. Identifying and quantifying linkages is difficult, especially with data collected during routine population surveys. Figure 3.3 illustrates how the proportion of overall mortality mediated by habitat attributes could change based on life stage and movement rate (as a proxy for
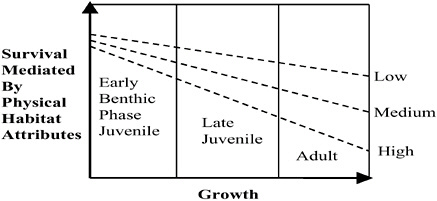
FIGURE 3.3 Conceptual model of the link between habitat attributes and mortality of demersal fishes based on general life history stages. Survival mediated by physical habitat attributes is a direct function of annual movement rates (low, medium, high) that serve as a proxy for habitat affinity. Movement rates are based on movements between habitat patches and are a function of patch size.
habitat affinity). Early benthic-phase juveniles have the highest rates of habitat-mediated mortality. Mortality rates for species that migrate become uncoupled from the physical attributes of habitat because their growth occurs at stages that are earlier than found in species that move less often and have greater affinities for habitat. For example, mortality rates for early benthic-phase Atlantic cod vary in relation to substrate complexity, but adult cod do not seem to exhibit particular small-scale shelter-related behaviors. Mortality of adults can be attributed to disease, senescence, predation from sharks and other large piscivores, and fishing.
INDIRECT EFFECTS
The relatively few studies of the indirect effects of trawling and dredging on marine ecosystems show results that are consistent with the basic principles of marine ecosystem dynamics and predator–prey interactions. Those potential indirect effects, summarized in Box 3.1, should be considered in evaluating the effects of fishing, and they should be used to inform future research and management decisions.
|
Box 3.1 Potential Indirect Effects
|
Sediment Processes
Fishing gear that disturbs the sediment surface can change sediment grain size distribution or characteristics, suspended load, and the magnitude of sediment transport processes (Churchill, 1989; Dyekjaer et al., 1995; Pilskaln et al., 1998; Riemann and Hoffmann, 1991). For example, water jets used in hydraulic dredges to harvest razor clams fluidize substrate for extensive periods (Tuck et al., 2000). Because water content and pore water turnover are important determinants of nutrient regeneration in marine sediments (Hopkinson et al., 1999), hydraulic dredging could alter the nutrient flux.
Bottom trawling and dredging can both resuspend and bury biologically recyclable organic material, changing the flow of nutrients through the food web (Mayer et al., 1991). Studies in relatively shallow depths (30–40 m) show a reduction in primary production by benthic microalgae after a disturbance (Cahoon et al., 1990, 1993; Cahoon and Cooke, 1992). Hence, disturbance in shallow water, including resuspension in the wake of trawls and dredges, could affect nutrient recycling and cause shifts in the abundance or type of microalgae.
The effects of gear-induced disturbance on ecosystem processes are difficult to predict for large marine ecosystems. It could be easier to identify systemwide effects at small spatial scales in semi-enclosed systems—such as bays, estuaries, or fjords—where water exchange with openshelf waters is restricted. However, in open coastal and outer continental shelf systems, the effects of gear disturbances can be small relative to the scale and rate of natural processes. Therefore, the spatial and temporal extent of disturbance by trawl and dredge gear should be evaluated to place these indirect effects within the context of the size and complexity of the ecosystem.
Species Interactions
Direct alterations of habitat can cause species shift and a general decline in the abundance of benthic organisms. Even species that are not directly exploited by a fishery are likely to be affected by the removal or disturbance of benthic and demersal biomass. For example, the early life stages of some pelagic species reside in or depend on benthic communities, for food and shelter. It is difficult to separate the indirect effects of trawling and dredging on benthic and pelagic com-
munities from other sources of variation, such as climate change (Jennings et al., 2001).
In some cases, removal of one species can have cascading effects on the rest of the ecosystem. For example, the combination of disease and high harvest rates over the past 150 years has reduced oyster density in the Chesapeake Bay to less than 1 percent. The loss of the filter-feeding oyster’s capacity to consume algae is hypothesized to be partially responsible for the proliferation of algal blooms. This appears to have shifted the composition of the pelagic community from mesozooplankton and fish to a community dominated by predatory jellyfish and comb jellies (Caddy, 1993; Ulanowicz and Tuttle, 1992).
RATES OF RECOVERY
Recovery is the return of an ecosystem to a state that existed before a disturbance, as measured by ecosystem processes, species composition, and species interactions. Recovery from trawling will depend on the type and extent of the habitat alteration, the frequency of the disturbance compared with natural changes, habitat characteristics, and species and life history characteristics. Recovery times vary according to the intensity and frequency of the disturbance, the spatial scale of the disturbance, and the physical characteristics of the habitat (sediment type, hydrodynamics). Superimposed on these human-related alterations are natural fluctuations, caused by storms or long-term climate changes, for example.
In most circumstances, only a first-order approximation of recovery rate is possible. Experimental evaluations recovery after cessation of trawling are limited and have focused on biotic recovery of small-bodied, short-lived invertebrates. Despite that, we can make some observations about the amount of physical disturbance that is sustainable in some types of habitat. The meta-analysis by Collie et al. (2000a) showed that recovery rate appears to be slowest in the more stable muddy habitats and biogenic (structurally complex) habitats. By comparison, mobile sandy sediment communities could be able to withstand 2–3 trawl passes per year without changing markedly. It is important to bear in mind, however, that although available data allow for prediction of the recovery rate for small-bodied taxa such as polychaetes (which dominate data sets for sandy sediment communities), less abundant, long-lived, and hence more vulnerable species could recover more slowly.
In some biogenic habitats, physical disturbance by dredging and trawling has a long-lasting effect. For example, clam dredging causes severe and persistent changes to seagrass ecosystems (Peterson et al., 1987; Stephan et al., 2000). After a single pass, seagrass biomass fell by about 65 percent below controls, and recovery did not begin for more than two years with seagrass biomass still roughly 35 percent below controls four years later (Peterson et al., 1987). The abundance of fish and shellfish that depend on seagrass for settling locations for protection from predators could be reduced where seagrass is damaged.
Environmental recovery after disturbance depends on the life histories of the organisms that live in or create the habitat. Recovery time is often one to five times the generation time of the organism (Emeis et al., 2001). Therefore recovery times could range from a few months—or less—to several decades (Hutchings, 2000). Many of the larger biogenic structure-forming organisms, such as soft corals and sponges, are slow growing and long-lived (Dayton, 1979; Leys and Lauzon, 1998). Empirical data about recovery times of corals and coral-line algae are sparse, but recovery times of decades to centuries can be inferred from the age of these organisms.
Recovery from trawling also depends on the size of the area disturbed (Thrush et al., 1998), and on the spatial pattern of the disturbance (Auster and Langton, 1999). Each trawl track is a small disturbance, but over a long enough period and with widespread coverage, the small changes can result in a large effect. The consequent habitat loss, and effects on resident species, depends to a large extent on its scale (Deegan and Buchsbaum, 2002). A single small loss might not, by itself, have an observable effect on species that are not directly damaged by trawling. However, the cumulative impact of many small losses may be quite significant at a regional scale (Odum, 1982). In some coastal ecosystems, mosaic-type damage could allow faster recovery than would a large-scale, isolated disturbance (Emeis et al., 2001).
Areas that are trawled with greater frequency could take longer to recover. Almost all studies have examined recovery after a single, acute pass by a trawl rather than after the multiple passes that are typical in frequently trawled, heavily fished areas. There, recovery would be expected to take longer because a larger fraction of the population is removed and immigration rates are lower (Figure 3.2). Results from the meta-analysis (Collie et al., 2000a) indicate that, on average, a single
dredge event results in a 76 percent, whereas repeated dredging results in an average reduction of 93 percent for anemones, malacostracan crustaceans, brittle stars, and polychaetes (Figure 3.3).
Few studies have examined the recovery of ecosystem processes or whole communities. Brylinski et al. (1994) showed that trawling significantly affected benthic diatoms that occurred in the intertidal zone, but that recovery occurred at all stations after about 30 days. The higher light intensity (and spectral composition) in the experimental area than at deeper sites, where trawling normally occurs, might have contributed to the relatively fast recovery.
The limited findings to date concur with theoretical predictions that suggest longer recovery times for more stable and complex habitats (Auster, 1998; Auster and Langton, 1999; Kaiser, 1998). Clearly, habitats with extended recovery periods are strong candidates for protection from disturbance caused by fishing. However, much better data on the geographic distribution and long-term effects of chronic trawling and dredging in these physically more stable habitats are required to estimate recovery rates that will promote strategic, rather than precautionary, management decisions.
UNCERTAINTY
Underlying the concept of the reversibility of the effects of dredging and trawling is the implicit assumption that eventual recovery to the former state will occur if the activity is halted. This assumption derives from an ecological paradigm in which ecosystems and communities are viewed as part of a successional continuum along a disturbance gradient. An alternative approach recognizes multiple-equilibria, non-linearity, and threshold effects (Holling, 1973; Holling et al., 1995; Patten and Constanza, 1997). In an alternative state, ecosystems have different species compositions, functions, and ability to provide ecological services. They might therefore be valued quite differently by society. Resilience, or the counteractive capacity of the ecosystem, is measured by the ability to maintain structure and function in the presence of stress or disturbance. When resilience is exceeded, the system can flip to an alternative state from which it will not return simply by removing the source of disturbance (Holling, 1973; Holling et al., 1995). These regime shifts can affect valuable ecosystem services, including fisheries yield (Collie and Spencer, 1994; Knowlton, 1992).
Human or natural modification of the marine environment might result in the shift of a community from one stable state that provides economically valuable fish to another stable state dominated by fish of higher or lesser value. Ecosystems respond to perturbation in many ways, including changes in species composition (by loss, inclusion, or replacement) and in the relative abundance of biomass (with an increase or decrease) of some species. Overall production and biomass of an ecosystem can remain the same as species respond to natural or human-induced stress (some species increase while others decrease) (Breitburg, 1998; Fogarty and Murawski, 1998). This change to a new stable state might take place abruptly, right after the disturbance occurs, or it could result from small cumulative shifts in natural forcing variables. Examples of species replacements are the apparent increase in cephalopod species in the Gulf of Thailand, which coincided with the increase in trawl fishing and the reduced abundance of demersal fish, and the increase in pelagic species that seems to have occurred in the North Sea and elsewhere. Its duration may vary. The return time to the initial stage has been predictable in some cases. In other cases, with nonlinear interactions and multiple-equilibrium states, the time the ecosystem will remain in a new state is not predictable. For benthic communities with a long history of fishing disturbance, it is unknown whether the community would return to the undisturbed state if the disturbance were stopped.
Human modifications to marine environments compromise the capacity of marine populations to recover from stresses, such as storms, eutrophication, and climate change, whether natural or anthropogenic. Seagrass ecosystems provide an example in which the synergistic effects of habitat loss due to trawling could compromise the ability of the system to withstand or recover from other disturbance. Seagrass ecosystems are important habitats and locations of fisheries for numerous fish and invertebrate species. The natural distribution of seagrass habitat is controlled by light availability that is a function of water quality, including the presence of phytoplankton and suspended sediments. The physical structure of seagrass, including stem density and the size of beds, increases water clarity by filtering water column particulates and depositing them on the bottom (Thayer et al., 1984). This creates a zone of clear water around seagrass beds that allows them to persist and expand. Trawling can fragment the seagrass bed into small pieces that do not effectively trap suspended particles, resulting in light
limitation. Eutrophication also enhances the proliferation of faster growing phytoplankton, epiphytic algae, and macroalgae that compete with seagrass for light and space (Kemp et al., 1983; Phillips et al., 1978; Short et al., 1995; Twilley et al., 1985). Light limitation of seagrasses leads to diminished growth and stature, increased shoot mortality and declines in shoot density (Duarte, 1995; Moore et al., 1996; Short et al., 1995), resulting in declines in seagrass habitat area. Initial habitat fragmentation by trawling and dredging can make seagrass habitats more susceptible to the negative effects of eutrophication.
The maintenance of the ecosystem in an alternative state will depend on interactions with adjacent ecosystems and the intensity of the new biologic links. Additional disturbance generated by natural events or by new trawling and dredging can help maintain the assemblage in this state of equilibrium or transfer it to a new state. In the benthos, disturbances can be physical (hurricanes, suspension of sediment by surf, lateral transport by bottom currents, seasonal hypoxia generated by the input of nutrients, limited export of biogenic carbon) or biological (predation, flux and export of biogenic carbon, deposition of debris, bioturbation, competitive exclusion). Their common action is to remove organisms and to open spaces for colonization by other organisms. If disturbances are frequent, gaps will constantly reset to one of the multiple stable stages. If disturbances are rare, most of the community will remain in a stable state for most of the time. The loss of complexity and biodiversity can threaten important ecologic functions (the cycling of important elements or the control of populations of particular species) or the resilience of ecosystems to change or disturbance.
SUMMARY
For the most part, existing information about the direct responses of benthic communities to trawling and dredging is consistent with the general principles that govern how ecologists expect communities and ecosystems to respond to acute and chronic physical disturbance. Trawling and dredging change the physical habitat and biologic structure of ecosystems and therefore can have potentially wide-ranging consequences. Mobile gear reduces benthic habitat complexity by removing or damaging the actual physical structure of the seafloor, and it causes changes in species composition. The reduction of physical structure in repeatedly trawled areas results in lower overall biodiversity. Of direct concern to commercial and recreational fisheries is the possibility that losses of benthic structural complexity and shifts in community composition will compromise the survival of economically important demersal fishes. Mobile gear also can change surficial sediments and sediment organic matter, thereby affecting the availability of organic matter for microbial food webs.
RESEARCH NEEDS
It is clear that the links between habitat alteration and loss of fisheries production can be subtle and diverse and that they operate on many spatial scales, from site-specific to regional. Most studies have been done in shallow water in small areas. Researchers have examined acute disturbances, rather than chronic, and they have studied short-term response focused in animal communities, as opposed to ecosystem processes such as nutrient regeneration. Although there have been many acute studies, few have examined the effects of short-term multiple passes, and future research should address this type of disturbance.
Perhaps the biggest research gap is on chronic effects and recovery dynamics. More studies on chronic disturbance by fishing gear are needed to determine the dose–response relationship as a function of gear, return time, and habitat type. Research also should address recovery dynamics, with consideration given to estimating the large-scale effects at current fishing intensities (e.g., Collie et al., 1997). This research should include quantitative studies undertaken in deeper water (>100 m) and studies in stable and structurally complex habitats, for which the recovery trajectory will be measured in years to decades. The statistical power to detect fishing effects will be greatest when biologic sampling can be combined with high-resolution spatial data on fishing effort.












