
2
Structure and Functioning of Riparian Areas Across the United States
The interaction of climate with the earth’s surface has created a variety of landscapes drained by networks of streams, rivers, lakes, and wetlands. Riparian areas are found adjacent to essentially all of these waterbodies except where human disturbance has intervened. Although riparian areas differ considerably in their structure and function from site to site, there are patterns in the attributes of riparian areas and how they are distributed across the landscape. While a single characteristic (such as the presence of bedrock) may strongly influence the size, characteristics, and functions of a given riparian area, generally the interaction of many climatic, hydrologic, geomorphic, and biological factors shape riparian environments. For example, differences in climate dictate the seasonality of the hydrologic cycle and determine the timing and intensity of flooding. Watershed features such the slope of the land, size of the watershed, storage capacity of the soil, and supplies of groundwater and sediment interact with climate to modulate or amplify these effects. Within the riparian area itself, further sources of variation can be found in channel morphology, sediment dynamics, and floodplain structure. Ultimately, all these factors influence species composition of riparian biota. This chapter focuses on the structure and functions of riparian areas, with an emphasis on those bordering streams and rivers rather than lakes and estuarine–marine waterbodies. Riverine riparian areas, because of their great collective length, comprise the vast majority of riparian areas in the United States.
FLUVIAL PROCESSES AND SEDIMENT DYNAMICS
Streams and rivers, which flow longitudinally downstream from higher elevations, can be classified by their size and the number of tributaries that flow
into them. As shown in Figure 2-1, headwater streams are classified as first order, with order number increasing in a downstream direction. Headwater networks of very small streams accumulate rainfall, overland flow, snowmelt, or aquifer discharge, sending variable amounts of water downstream to increasingly larger channels.
The water budget of all streams and rivers is determined by climate and by other watershed attributes such as topography, soil type, bedrock substrata, groundwater discharge, and vegetation. Natural flow patterns—unregulated by dams and water diversion—will vary with the dynamics of water delivery and cycling, unless the source is a spring fed by a deep (phreatic) aquifer that has very little surface connection (Gibert et al., 1994; Vervier, 1990). According to Poff et al. (1997), the flow regime of a river can be distinguished by several major components, including magnitude, frequency, duration, timing, and rate of change, as described in Box 2-1. River flows are often described using one or more of these components. Thus, for example the bank-full flow, which defines the bank-
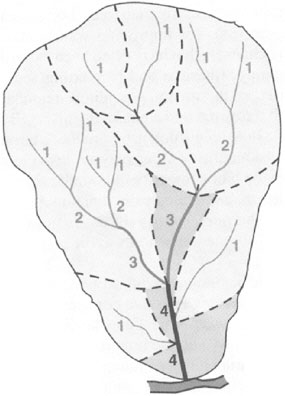
FIGURE 2-1 Stream orders for a watershed that includes first- to fourth-order streams. Ephemeral streams are not shown on this diagram. SOURCE: Reprinted, with permission, Strahler (1952). © 1952 by The Geological Society of America.
|
BOX 2-1 Because streamflow is strongly correlated with critical physical and biological characteristics of rivers, such as water temperature, sediment transport, channel morphology and habitat diversity, it represents a “master variable” that influences the functions of associated riparian areas. Flow magnitude represents the amount of water moving past a given location per unit time. It can influence rates of solute, suspended sediment, and bedload sediment transport, and thus is a critical variable with regard to the creation of alluvial landforms (e.g., point bars, floodplains streambanks, and channel sinuosity). As discussed later in this chapter, high flows are needed for some species to create local zones of erosion/deposition for seedling establishment. Flow frequency refers to how often a flow of a given magnitude is equaled or exceeded over some time interval. Flow frequency, in combination with flow magnitude, indicates the amount of energy a stream has to do work (e.g., sediment transport, channel adjustments, etc.). Flow duration represents the period of time associated with a specific flow magnitude. From the perspective of riparian plant communities and floodplain functions, flow duration represents the length of time that overbank flows occur or that soils remain saturated from high flows. Flow duration is often a crucial variable for many riparian plants that have adapted their physiology to accommodate extended periods of high moisture levels. Flow timing generally refers to the seasonality of a given flow. For example, the timing of most snowmelt runoff for many western streams and rivers occurs in late spring and early summer. Fish and other organisms have adapted their life history strategies to the timing of these flow periods. Superimposed upon the long-term water and sediment budget of the watershed, flow timing determines the relative wetness or dryness of the adjacent riparian area and is therefore a primary structuring process. The rate of change in streamflow or water levels represents how quickly a flow changes from one magnitude to another. Streams and rivers that derive their flow from snowmelt are generally considered less “flashy” than those that respond to large amounts of rainfall. Rate of change can influence water sediment transport rates and riparian plant communities. For example, seedlings of deciduous woody species may need a relatively low rate of change during snowmelt recession flows for them to successfully establish. |
full channel, is the discharge of the 1.5- to 3-year return period storm (Dingman, 1984). Floods are of larger discharge and generally occur less frequently than bank-full events. Floods move and sort sediments and other materials, forming the physical structures that compose the riparian areas of rivers. Big floods, which are relatively rare, often create a physical template that is continually
reworked and modified by lower flows. Hence, diverse alluvial landforms, such as gravel bars, floodplains, islands, terraces, and the channel network, are created by flow-mediated movement of sediments and are in a constant state of change (Ward, 1998; Ward et al., 2000).
The size and character of streamside riparian areas is directly related to water delivery to and flux through the watershed. One pattern is the tendency for riparian areas to be expansive next to big, larger-order rivers, which in part reflects multiple hydrologic sources (e.g., seasonal overbank flows from the river, flood-related flows in secondary channels, and groundwater discharge, all discussed in detail in a later section). Periods of high flow, particularly in unconstrained or relatively wide alluvial valleys, can create a multitude of landforms (e.g., streambanks, floodplains, and terraces) that are common to many riparian systems. However, such basic patterns are often too simplistic to be widely useful in predicting structure and function of riparian areas across many landscapes. For example, the lower Columbia River (9th-order) upstream of Portland, Oregon, is constrained by resistant bedrock, resulting in narrow floodplains and minimal riparian areas (see Figure 2-2 for the difference between constrained and uncon-
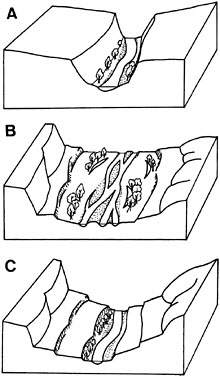
FIGURE 2-2 Geomorphology of a stream corridor in (A) a constrained reach, (B) an unconstrained aggrading reach, and (C) an unconstrained degrading reach. SOURCE: Reprinted, with permission, from Dahm et al. (1998). © 1998 by Blackwell Science Ltd.
strained river reaches). Another generalization is that where fluvial systems encounter relatively wide valleys and low channel gradients, they typically develop a system of meanders and floodplains that represent both sediment and water discharge regimes in dynamic balance with valley and channel gradients, channel morphology, and riparian vegetation. In general, these sinuous channels occur within a definable meander belt, simply defined by a linear boundary that connects the outer margins of the existing channel meanders. More accurate geomorphic delineation of the meander belt would be based on the actual margin of the floodplain, boundaries of historical channel meanders (as shown for the Willamette River in Figure 2-3), or boundaries of past recorded inundation extent.
Because river flow naturally is erosive and water is the universal solvent, particulate materials (sediment, rocks, trees) and dissolved materials (salts, organic compounds) are exported downstream in proportion to stream power and
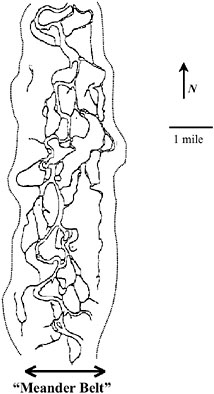
FIGURE 2-3 Meander belt of the Willamette River based on the channel meanders in a reach from Eugene to Harrisburg in 1850. SOURCE: Modified from Williamson et al. (1995).
deposited in a wide array of fluvial landforms within the river channel and floodplain. A generalized three-dimensional view of riparian areas includes portions of the channel and associated features (gravel bars, islands, large wood); a parafluvial zone (which corresponds to the bank-full width) that experiences the seasonal range of flow variation; a vegetated area of varying successional states influenced by floods, sediment deposition, and water availability; and a transitional zone to the uplands (see Figure 1-4). Furthermore, these features are generally underlain by an alluvial aquifer that can have a major influence on riparian processes, particularly where bed sediments are deep (Stanford, 1998). The longitudinal, lateral, and vertical pathways through which water and materials are conveyed through riparian areas are discussed in detail throughout this chapter.
Erosion and Deposition
The processes of erosion, transport, and deposition continually disturb and reshape the riparian environment. Materials from upstream sources such as erosion zones along hillslopes and riparian terraces or landslides are sorted by flowing water and transported downstream until the physics and energetics of the transport process dictate deposition either in the channel or on the floodplain of the river. As shown in Figure 2-4, flow-mediated erosion of sediment occurs in the steep gradients of lower-order segments, deposition of course material (gravel,
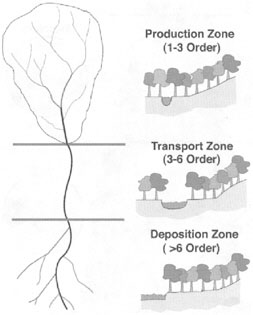
FIGURE 2-4 The geomorphic zones of a fluvial system. SOURCE: Reprinted, with permission, from Schultz et al. (2000). © 2000 by American Society of Agronomy.
cobble) occurs in the middle reaches related to aggradation of the river valley and associated loss of flow velocity (energy dissipation), and deposition of fine materials (sand, silts) occurs in the lowest-velocity environments that characteristically occur in the high-order segments of the Piedmont or coastal plain. Hence, as the size of the stream increases, the size of the floodplain generally increases.
Although these broad patterns in sediment transport explain a trend of downstream fining in the grain size of bed sediment in many river systems (Schumm, 1960), in reality sediments of all sizes are sorted along every channel or floodplain within the river corridor. Flowing water sorts the sediment between different areas of the channel with different capacities for maintaining sediment in suspension. Coarse sediments are suspended and deposited only in the highest-energy environments of the river channel, i.e., areas with relatively high velocity. Fine-grained sediments, in contrast, are generally restricted to the lowest-energy backwaters of the active channel or to the floodplain.
Although bank-full flows maintain channels, floods account for much of the major work in reshaping channels and floodplains. Increased production of sediments from terrestrial sources and acceleration of bank erosion during floods can release large amounts of fine- and coarse-textured sediment into a channel over a short period of time, which are then deposited in downstream channels or on floodplains. Floods also cause substantial realignment of channels because of reoccupation of secondary or abandoned channels by newly released sediment (Beschta et al., 1987a). When significant amounts of coarse sediment become available locally, rapid adjustments to the morphology of the channel can occur. These effects may be transmitted in both the upstream direction (backwater effects, including upstream bed material storage and an altered channel morphology; channel incision and gully head cuts) and the downstream direction (higher levels of sediment transport with the potential for increased channel instabilities).
The net result of fluvial processes over decades to millennia is a slow modification and reworking of the channel and floodplain physical template such that sediment routing must be viewed as a constantly changing feature of all alluvial rivers. Distinct features such as cutbanks, meander scrolls, and point bars migrate over time. Within the meander belt, the deposition of sediments on vegetated floodplains occurs periodically over time and the exact character, dimensions, and location of meanders may incrementally shift as a result of the migration of gravel point bars and the erosion of cutbanks. Secondary channels become plugged, creating backwater sloughs or oxbow lakes. Slugs of sediment derived from an episodic landslide may take many years to move down the river corridor, influencing riparian areas to different extents as they pass through specific segments. Large tree boles eroded from riparian areas substantially increase the variation in sediment transport and deposition, and thus also the variety of habitat types available for biota (Naiman et al., 2001).
Cycling of sediments back and forth between the main channel and the channel’s banks and floodplain is an important component of sediment transport
in rivers (Meade et al., 1990). Floodplains expand laterally and “grow upward” due to the long-term deposition of fine sediment during recurring overbank flows. Seasonal high flows continually disturb new areas. Newly deposited sediments on floodplains undergo biogeochemical changes (i.e., diagenesis) that will over time transform a flood-deposited sediment into a riparian soil. The overall result is the creation of a complex patchwork of riparian areas, each with a slightly different microenvironment of sediment grain size and nutrient and water availability, and each at a different stage of development since the last disturbance (Amoros et al., 1987; Malanson, 1993).
Because of the dynamic flow and sediment transport regimes often associated with riparian areas, their soils reflect a high degree of unevenness in particle sizes, soil depth, and the amount of associated compounds such as organic matter. Highly variable water levels typically result in morphological soil features such as mottling, gleying, oxidation/reduction, and others. However, in instances where floodplains have been slowly built up via the incremental deposition of fine silt layers over many centuries, soil characteristics across extensive areas may be relatively uniform. Floodplain soils have been some of the most productive areas in the nation for agricultural production due to their high levels of nutrients and organic matter.
Flow Modification Within Riparian Areas
Although the energy from water moving down a channel can be used to do work (e.g., scour banks and transport sediment), the vast majority is used to overcome the frictional resistance provided by a channel’s bed and banks and is eventually dissipated as heat. Thus, streamside riparian areas are responsible for the dissipation of energy associated with flowing water. The flow resistance, or roughness, of a stream reach, caused by the physical configuration of its channel, streambanks, and floodplains as well as by the riparian plant communities, can be described by a roughness coefficient, such as Manning’s n (Leopold et al., 1964). Cowan (1956) identified several major channel conditions that affect roughness: bed material, degree of surface irregularity, variations in channel cross section, relative effects of obstructions, degree of meandering, and effects of vegetation. Importantly, vegetation can directly or indirectly affect all these conditions, with the possible exception of bed material, thus indicating it often has a major influence on channel roughness and on how channels dissipate stream energy during periods of high flow. Herbaceous riparian vegetation increases local friction on streambanks by creating flexible and three-dimensional barriers to flow. Riparian graminoids (grasses, sedges, rushes) and shrubs are particularly effective at trapping sediments during high flows and helping to maintain stable streambanks. For forest floodplains, roughness increases directly with the density and size of trees (Li and Shen, 1973; Petryk and Bosmajian, 1975). Large wood provided to streams and rivers from riparian forests can also have a significant effect on
channel roughness via the occurrence of debris jams and other accumulations that alter flow patterns (Abbe and Montgomery, 1996; Montgomery et al., 1996; Piegay and Gurnell, 1997).
At high flow, streambanks, floodplains, and their associated vegetation provide resistance to flowing water, thus locally altering patterns of scour, sediment transport, and deposition (Sedell and Beschta, 1991). For example, low velocity zones have been observed to develop when floods pass through riparian forests, creating sites for the retention of sediment and organic matter and refuges for aquatic organisms (Swanson et al., 1998). Floodplain vegetation is especially effective at providing protection from scour, which is why well-vegetated floodplains typically are areas of long-term sediment accumulation.
During periods of low flow, woody species have a much less significant effect on flow roughness because of the smaller surface area exposed to surface flow, such that flow resistance tends to be controlled more by the morphology of the channel. In contrast, aquatic macrophytes and graminoids can greatly influence the resistance provided during low-flow periods (Kauffman and Krueger, 1984). Finally, the uptake and transpiration of water by riparian and upslope vegetation during low-flow periods can alter discharge, thereby influencing aquatic habitat (Rothacher, 1970; Troendle, 1983; Cheng, 1989; Keppeler and Ziemer, 1990; Hicks et al., 1991).
Lacustrine Riparian Areas
Unlike the riparian areas of stream and river (lotic) environments, riparian areas bordering lakes differ significantly in the energy sources that drive physical mixing (Wetzel, 2001). In the shallow littoral environments of lakeshores, mixing is generally driven by temperature gradients and storm-generated waves. An important contrast with lotic environments is the type and frequency of waterlevel changes at lakeshores. Seiches, for example, can cause substantial changes in water level over periods of days to weeks at the shores of large lakes without the kinds of erosive forces of floods that affect channel floodplains. Lakeshores also tend to have much larger water-level changes over longer-term (interannual) cycles, as determined by interannual variation in climate and the regional water balance. Large reservoirs and other river impoundments used for water storage may exhibit nonseasonal fluctuations in water level, with hydrographs varying erratically under the control of hydropower production or irrigation supply. Consequently, riparian areas around reservoirs are highly variable and often are composed of non-native, invasive species because they have little long-term continuity in water supply and occur in areas of the landscape that have no legacy of native plant colonization (e.g., Nilsson et al., 1997).
Despite major differences in flow velocities and extent of water-level changes, the shallow littoral environment and riparian areas adjacent to lakeshores have much in common with riparian areas bordering streams. As in streams, a
broad range of sediment types and textures is often available, nutrients are often ample and primary productivity is high, water exchange between the surface and subsurface is conducive to high rates of biogeochemical cycling in sediments, and secondary productivity in these environments is typically high (Wetzel, 2001). In the case of large lakes with inlets from rivers, alluvial deltas may develop by sediment deposition in the river–lacustrine confluence. Often, river deltas in lakes and reservoirs facilitate robust riparian areas in a manner similar to the islands and low terraces that occur in alluvial rivers. Deltaic riparian areas can be large landforms up to many square miles in size. Few studies have been done in such environments (e.g., Stanford and Hauer, 1992).
In summary, riparian areas are characterized by a spatial and temporal mosaic of conditions reflecting variability in sediment type and particle size distribution, timing of water sources and water quality, and time since disturbance by floods. Seasonal dynamics in flow and sediment transport constitute the foundation of riparian structure and thus influence the resulting colonization by riparian species and the many functions performed by these areas. Moisture availability and anoxia in riparian soil are additional factors that closely follow the distribution of grain sizes determined by fluvial processes. In many channels, the natural variability of flow has been regulated and sediment inputs have been curtailed downstream of dams and water diversions. As discussed in Chapter 3, the influence of humans in regulating river flow has had overwhelming effects on ecological processes in rivers and riparian areas, because of the disruption of flow seasonality, sediment dynamics, and moisture availability.
HYDROLOGIC AND BIOGEOCHEMICAL PROCESSES
Hydrologic Pathways in Riparian Areas
Riparian areas receive water from three main sources: (1) groundwater discharge, (2) overland and shallow subsurface flow from adjacent uplands with additional input from direct precipitation, and (3) flow from the adjacent surface water body. The major losses of water from riparian areas include groundwater recharge and evapotranspiration. Plate 2-1 illustrates these major water flow paths for a streamside riparian area. Both the quality (in terms of dissolved and particulate constituents) and the timing of water from these sources vary considerably. For example, the discharge of deep groundwater is on the order of centuries, while overbank flows and intense rainstorms can change flows within minutes.
Groundwater Sources
Winter et al. (1998) outlines some of the basic interrelationships between groundwater and surface water in streams and lakes and shows how interactions vary as a result of differences in climate, topography, and surficial geology.
Streams whose downstream flows increase as a result of groundwater discharge are referred to as gaining streams. In contrast, flow in the channel decreases in the downstream direction in losing streams that recharge the groundwater system. Because of variability in water sources and hydrogeologic properties of aquifers, it is typical for streams to simultaneously experience discharge in one reach while experiencing recharge in others. For example, steep mountain streams gain water by groundwater discharge in their upper reaches and then lose water as they flow out of constricted mountain valleys onto alluvial fans.
Lakes and wetlands share some of the same relationships with groundwater as do streams. Lakes and wetlands commonly discharge and recharge simultaneously in different parts of the system and experience flow reversals seasonally (Figure 2-5). As in streams and rivers, movement of water between groundwater and surface water is influenced by the nature of the substrata and the water elevation in the lake compared with water levels and gradients in groundwater of the adjacent aquifer (Sebestyen and Schneider, 2001). Water moves from areas of high elevation to areas of low elevation, sometimes involving streams or rivers at inlets or outlets to the lake.
Because the majority of riparian areas are associated with stream and river channels, this discussion focuses on interactions between groundwater and river channels rather than lakes. From a relatively large-scale perspective (miles or greater), the direction of groundwater flow in the vicinity of rivers is typically associated with patterns of floodplain and channel topography. As a result, flow pathways are seldom entirely parallel or entirely perpendicular to the main channel but instead occur diagonally toward the channel in a downstream direction. The major controls on orientation of groundwater flow paths are hydraulic properties of aquifer materials, regional gradient, and sinuosity of channel (Larkin and Sharp, 1992). Groundwater that tends to flow parallel to a channel is referred to as underflow (Larkin and Sharp, 1992); in contrast, groundwater flow perpendicular to and toward the channel is referred to as baseflow (Hall, 1968) (see Figure 2-6).
At much smaller spatial scales, i.e., feet to tens of feet, interactions between groundwater and riparian areas are influenced primarily by heterogeneities of riparian and channel sediments, which have a critical effect on local direction and flow rate of groundwater. In some settings, baseflow passes directly through riparian sediments, while in others, baseflow may bypass riparian sediments by flowing through coarse material underneath and discharging vertically from directly beneath the stream bed (Phillips et al., 1993). This short-circuiting of the root zone can have important implications for the extent of certain transformation processes that occur in riparian areas. As discussed later, the variation in the specific flow paths characteristic of riparian areas may explain why some buffers are not as effective as others.
An often-overlooked aspect of groundwater–riparian–channel interactions is that groundwater discharge is not equivalent along all parts of a channel. Instead, certain channel subreaches tend to collect a significant proportion of all ground-
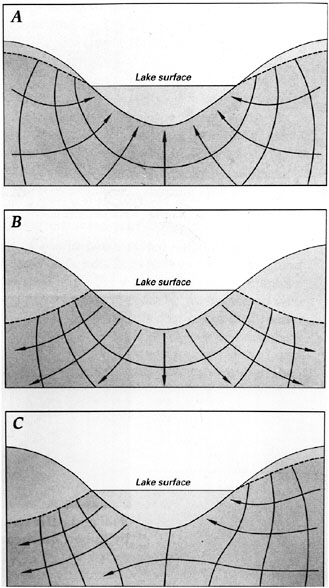
FIGURE 2-5 Lakes, like streams, can receive groundwater inflow (A), lose water as seepage to groundwater (B), or both (C). SOURCE: Winter et al. (1998).
water entering a stream in a given reach. Groundwater discharge points tend to occur at the upstream ends of pools, the upstream side of meanders, anywhere along the channel thalweg (deepest area of central channel), and within side channels or alcoves in streams and rivers (Harvey and Bencala, 1993). An un-
common example of a groundwater discharge point is a visible area of groundwater seepage on the channel bank above the level of the channel.
The importance of groundwater discharge points along channels is twofold. First, areas along channels that collect groundwater discharge tend to favor establishment of rich riparian vegetation, especially in dry climates where water avail-
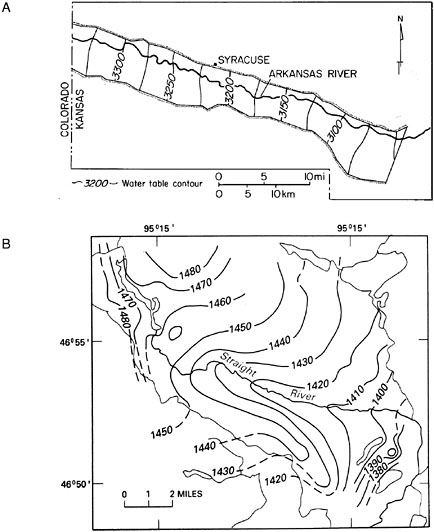
FIGURE 2-6 Examples of (A) underflow-dominated groundwater movement parallel to the channel and (B) baseflow-dominated groundwater movement perpendicular to the channel. Groundwater moves in the direction of decreasing water table contours.
SOURCES: (A) Reprinted, with permission, from Larkin and Sharp (1992). © 1992 by Geological Society of America. (B) Winter et al. (1998).
ability is a major factor limiting the establishment and maintenance of riparian vegetation. Second, groundwater discharge points tend to have cooler water locally in the channel during summer and warmer water in winter compared with other channel areas. As such, they may be important in creating thermal refugia for aquatic organisms at particularly sensitive times in their life cycles.
Hillslope Sources
Hillslope sources of water to riparian areas all begin as precipitation falling on the landscape. There are numerous pathways for water to travel from the hillslope to riparian areas. For example, overland flow begins as precipitation that exceeds the percolation capacity of the soil. Precipitation can also travel downslope via shallow subsurface flow (a portion of which may emerge onto the hillslope surface before reaching the channel). Precipitation that falls directly onto saturated areas of the hillslope further augments these pathways. All of these flow mechanisms are referred to collectively as hillslope runoff. Figure 2-7 contrasts hillslope runoff in three situations differing in climate and soil development. Humid forested landscapes with deep permeable soils have deep percolation and groundwater flow to riparian areas, in addition to shallow flow on the lower hillslope during intense storms (Figure 2-7A). In areas where local geology includes soil layers of low permeability, drainage is often restricted to shallow permeable soil (Figure 2-7B). In arid areas, intense precipitation onto hillslopes with sparse xerophytic vegetation and impervious soils creates a situation where overland flow is often the dominant pathway of drainage (Figure 2-7C).
Topography and hydraulic properties of sediments influence the degree to which riparian areas store hillslope runoff or transmit it to the channel. If comprised of coarse sediments, the riparian area can usually store large quantities of hillslope runoff and release it to the channel by groundwater discharge. Riparian sediments that are relatively fine and are lower in permeability than other soils of the watershed generally cannot store large quantities of water quickly enough, leading to rapid expansion of saturated areas. During intense storms, shallow subsurface flow that cannot move fast enough laterally emerges as return flow onto the surface of the riparian area. This flow is further augmented by rain falling directly on saturated areas. The expanding areas of saturation in the riparian area that result from hillslope runoff are referred to as variable source areas (Hewlett and Hibbert, 1967) or partial contributing areas (Dunne and Black, 1970). Such areas become saturated during the early part of intense storms, expanding further if rainfall continues. Depending on storm intensity and duration, the concave upward areas of valley bottoms and hillslope hollows tend to become saturated because they collect storm water faster than they deliver it to channels.
As is obvious from the preceding discussion, hillslope runoff is a highly variable process in space and time that depends on a variety of factors, particu-
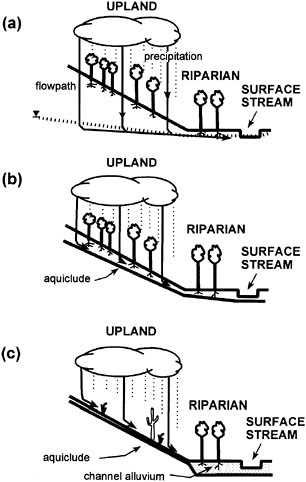
FIGURE 2-7 Hydrologic linkage of upland ecosystems with stream corridor ecosystems in (A) forested landscapes with deep, well-drained soils where water percolates well beyond the rooting zone, (B) forested landscapes with shallow soils where underflow intercepts the rooting zone, and (C) arid and semiarid landscapes where soils of low permeability force overland flow. SOURCE: Reprinted, with permission, from Fisher et al. (1998). © 1998 by Springer-Verlag.
larly topography, sediment hydraulic properties, and antecedent groundwater levels and soil moisture. Weather is also important through its effect on the intensity and duration of precipitation, temperature, and solar radiation patterns. These factors in turn determine the types and amount of vegetation present in riparian areas and thus the extent of evapotranspiration. The interplay between these factors and their effect on hillslope runoff are summarized in Figure 2-8.
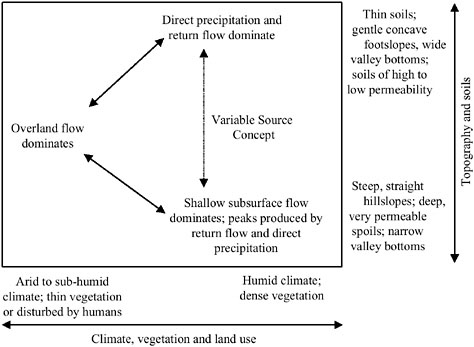
FIGURE 2-8 Dominant processes of hillslope runoff in response to rainfall. SOURCE: Adapted from Dunne (1978).
Channel Sources
A final source of water to riparian areas is from the channel itself via overbank flow, bank storage, or hyporheic flow. The extent and duration of overbank flow, or flooding, of riparian areas is very much dependent on intensity and duration of precipitation, basin area and topography, soil and aquifer type, and morphology of the river channel and floodplain. In general, when channels reach flood stage during storms and floods, riparian areas temporarily store excess water that cannot be quickly conveyed downstream. The overall effect of this water storage is delay and attenuation of the flood peak in downstream areas (Moench et al., 1974; Bhowmilk et al., 1980). Figure 2-9 shows the diversity of flooding environments for a 50-year flood on the Rhône River floodplain system. Floodwaters that overtop channel banks over a period of many decades and centuries and the sediments they carry are essential to the creation and maintenance of floodplain landforms mentioned earlier, such as levees and ridges. Rapid flow across riparian areas can rehydrate abandoned channels that directly flow back to the channel. Floodwaters that become trapped in topographic depressions, such as isolated sloughs, subsequently recharge the groundwater system; deposition of fine sediment, such as fine silt and clay, occurs during these long
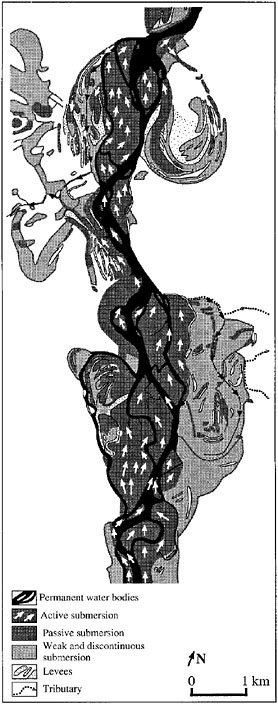
periods of standing water. Hupp (2000) found that deposition rates are relatively high in southeastern coastal plain rivers because of the frequent return intervals of overbank flow and the relatively high sediment loads of the rivers. Riparian plant communities in Oregon have been shown to be associated with floodplain surfaces that receive relatively frequent overbank flows of at least once every five years (Chapin et al., 2000). As shown schematically in Figure 2-10, the relative importance of overbank flow versus hillslope runoff typically increases with increasing stream order.
Subsurface movement of water from the channel into the groundwater aquifer beneath the floodplain is sometimes an important source of water to riparian areas, particularly in ephemeral and intermittent streams. In perennial streams, this usually involves bidirectional exchange back and forth between the surface channel and groundwater beneath the floodplain. Two types of bidirectional interactions deserve special mention. The first is bank storage, which involves channel water moving laterally into subsurface riparian areas when river stage is high, and then gradually moving back to the channel when river stage drops (Pinder and Sauer, 1971) (Figure 2-11A). Bank storage in riparian areas can affect water storage, chemical transformations in streams and rivers, surface water temperature, and the composition and extent of riparian plant communities.
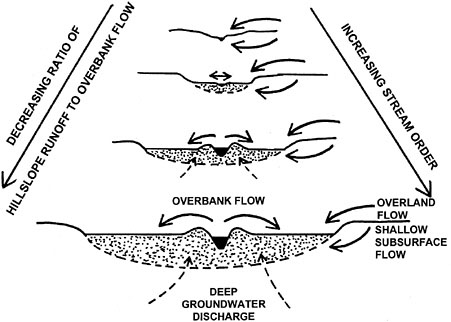
FIGURE 2-10 Relative importance of hillslope runoff versus overbank flow. SOURCE: Reprinted, with permission, from Brinson (1993). © 1993 by Dr. Douglas A. Wilcox, Editor-in-Chief, Wetlands.
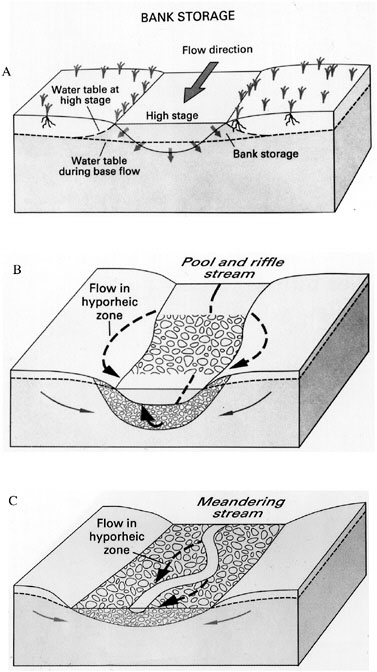
FIGURE 2-11 Water exchange between channel and riparian areas caused by (A) change in stream stage followed by bank storage, (B) streambed topography routing streamflow temporarily through subsurface (hyporheic) flow paths, and (C) hyporheic flows through bends of meandering stream. SOURCE: Winter et al. (1998).
Squillace (1996), for example, demonstrated that bank storage in the Cedar River (Iowa) sequestered (and possibly led to degradation of) pesticides such as atrazine that had previously been transported to the channel by spring runoff from agricultural fields. After the peak spring flows in the Cedar River, pesticides were slowly discharged back to the river over a period of weeks to months, accomplishing a dilution of pesticide reaching the river.
The other type of bidirectional interaction is hyporheic exchange, which is the temporary routing of water through gravel bars and the alluvium surrounding stream channels (i.e., the hyporheic zone—see Figure 1-4). The extent of the hyporheic zone is defined operationally using solute tracers, e.g., as the depth in the sediment where tracer concentrations indicate that 10 percent or more of the water is derived from the channel (Triska et al., 1989). Channel flow enters the hyporheic zone due to uneven pressure gradients over a rough streambed, or due to pooling of water at higher elevations behind flow obstructions such as riffles (Harvey and Bencala, 1993; Wroblickly et al., 1998). As shown in Figure 2-11(B) and (C), channel flow that is routed through hyporheic zones generally returns to the channel within a relatively short distance downstream. Passage of stream water through hyporheic flow paths increases oxygen concentrations in the subsurface, creating specialized habitats for burrowing organisms beneath streamside riparian areas (Jones and Mulholland, 2000) and optimal conditions for bull trout and salmon eggs where hyporheic flow returns to the channel. Certain biogeochemical reactions are also enhanced in hyporheic zones, affecting the transport and transformation of nutrients, metals, and organic compounds (Jones and Mulholland, 2000).
Role of Transpiration
Trees, shrubs, and herbaceous plants contribute significantly to water cycling and material movement in riparian areas. The most direct effect of plants on water flow and storage in riparian areas is transpiration. In smaller channels of headwater basins, riparian evapotranspiration accounts for a percentage of the groundwater that would otherwise be discharged to the channel (Daniel, 1976). Early studies along the Gila River in Arizona estimated that evapotranspiration removes 12.3 percent of the water from the system, while evaporation from the river surface and wet sand bars removed 2.5 percent (Gatewood et al., 1950). In a study of approximately 50 basins in the Appalachian Valley and Piedmont areas of the Mid-Atlantic region, riparian transpiration removed approximately 10 percent of recharged groundwater prior to discharge to streams (Rutledge and Mesko, 1996). Transpiration in relatively small headwater streams is significant enough to cause a diel cycle in streamflow, with decreased streamflows during the day and increased streamflows at night. Harvey et al. (1991) found that the size of the hyporheic zone in riparian areas expanded during the day and contracted at night, in accordance with riparian transpiration, providing circumstantial evidence that
in some areas, streamflow may be a significant source of water for riparian transpiration.
An important need in western riparian areas is to improve our understanding of the water-use requirements of riparian vegetation. This involves quantifying the relative importance of various water sources for transpiration, including recently recharged precipitation, groundwater, and surface water. Modern tools are being applied to this problem, including the use of water-stable isotopes in tree sap as tracers to identify the source of water for transpiration (Flanagan et al., 1992; Dawson and Ehleringer, 1993). Early results indicate that riparian plants use different components of water in the alluvium; sacaton grass, for example, uses recently recharged precipitation from the unsaturated zone (Moran and Heilman, 2000). In contrast, mesquite uses a mixture of groundwater and unsaturated zone water depending on tree size, cottonwood uses mostly groundwater, and willow uses only groundwater. When combined with sap-flow measurements, water-stable isotopes determine how water use by riparian trees changes with forest age, groundwater levels, and climatic fluctuations.
Unlike evapotranspiration, direct evaporation from stream reaches is usually small in comparison with groundwater discharge or flow inputs from upstream. However, evaporation can make a measurable contribution to the water budgets of lakes.
* * *
As evident from the preceding discussion, hydrologic fluxes through riparian areas are highly variable in both space and time. Time scales range from minutes to hours (hyporheic flow and transpiration), days to months (storm and seasonal snowmelt response), and years to decades (climatic effects on recharge and baseflow discharge). As a result of that complexity, it is entirely possible that a single riparian area could function some of the time as a pathway for groundwater discharge, at other times as a hyporheic zone, and at still other times as a zone of bank storage. Any assessment of the hydrology of riparian areas therefore depends not only on physical attributes of the channel, watershed, and climate, but also on spatial and temporal boundaries of the particular problem. For example, determining only the net groundwater exchange in a stream or lake (i.e., the difference between discharge and recharge) is not always adequate for characterizing groundwater interactions that affect riparian areas. In many situations, both groundwater discharge from the watershed and recharge from the surface water body are important, necessitating more thorough investigations of water fluxes using multiple approaches (Krabbenhoft et al., 1990; Harvey and Bencala, 1993; Hunt et al., 1996; Choi and Harvey, 2000). Thus, there is no universally acceptable approach to characterizing the water balance of riparian areas, and many studies employ significant simplifications, assumptions, or other qualifications. Examples of riparian water balances developed for various purposes include
those by Goodrich et al. (2000), MacNish et al. (2000), and Rutledge and Mesko (1996).
Biogeochemical Interactions Between Riparian Areas and the Surrounding Landscape
Along with water flow through riparian areas comes the transport and transformation of chemicals and particulate matter—key factors that affect the ecology of rivers and lakes. These processes have been most intensely studied within stream channels, although the role of riparian areas and groundwater in influencing adjacent aquatic systems is increasingly being explored. Although this section relies primarily on research conducted in riverine settings, broad concepts connecting physical and ecological factors have also been developed for lakes and wetlands (see Labaugh et al., 1996; Kratz et al., 1997; Carpenter et al., 1998; Wetzel and Søndergaard, 1998; and Wetzel, 1999).
Instream Processes
Nutrients are cycled within streams and rivers, moving back and forth between inorganic forms and the living tissue of biota. Dissolved inorganic nutrients and nutrients associated with fine particulate organic or inorganic matter move with the flowing water, while nutrients in biotic compartments, such as microbes, periphyton, aquatic plants, and riparian trees, spend much longer in one place within the stream corridor. The cycling of nutrients between transported and fixed components is the basis of the Nutrient Spiraling Concept, which refers to the sequences of movement and temporary retention that occur during downstream transport (Newbold et al., 1982). The concept has been tested in the field through experimentation using dissolved tracers such as salts to track water flow and isotopic forms of important nutrients such as nitrogen and phosphorus (Figure 2-12). Recent work suggests that variations in hydrologic processes, not in biological and geochemical processes, are the most significant cause of site-to-site variation in the cycling of dissolved and fine particulate materials in streams (Findlay, 1995).
Consistent with the Nutrient Spiraling Concept is the River Continuum Concept (Vannote et al., 1980), which sets forth a classification of ecological processes in streams and rivers that is firmly grounded in the principles of fluvial geomorphology and hydrology. The River Continuum Concept places ecological processes, such as productivity, respiration, and food web structure, in an interdependent upstream–downstream sequence.
A limitation of the Nutrient Spiraling and River Continuum Concepts is that they emphasize longitudinal transport and changes in dominant processes according to channel hydrology. Thus, they refer primarily to the wetted channel and bottom sediment, rather than to riparian areas. The Flood Pulse Concept (Junk et
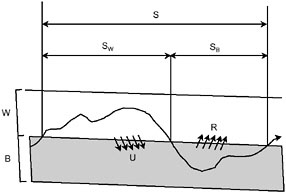
FIGURE 2-12 Two-compartment nutrient spiraling model. The spiraling length S is the average distance a nutrient atom, such as phosphorus, travels downstream during one cycle. A cycle begins with the availability of the nutrient atom in the water column, and includes its distance of transport in the water (SW) until its uptake (U) and assimilation by the biota, and whatever additional distance the atom travels downstream within the biota (SB) until that atom is eventually re-mineralized and released. W = water, B = biota, R = release. SOURCE: Modified from Newbold et al. (1982).
al., 1989; Bayley, 1991), on the other hand, emphasizes the role of lateral exchanges between the channel and floodplain (and thus through riparian areas) as an integral driver of ecosystem processes in river corridors. As shown in Figure 2-13, biogeochemical interactions between riparian areas and channels are prob-
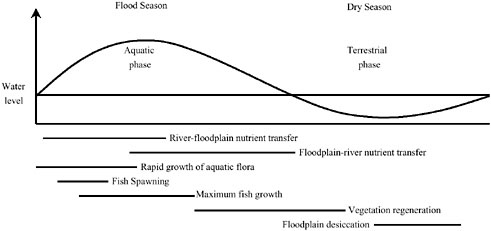
FIGURE 2-13 The influence of the flood pulse within the river–floodplain complex. SOURCE: Reprinted, with permission, from Bard and Wilby (1999). © 1999 by Routledge Publishers.
ably most intense during storms (“river–floodplain nutrient transfer”). Material fluxes of water, sediment, nutrients, and organic matter are maximized at flood stage. For example, floods deposit new sediments with associated nutrients on the riparian surfaces, creating the conditions that favor high primary productivity of riparian vegetation. Flood pulsing is also an efficient means of moving carbon and nutrients out of the forest and into the main channel. The transformation and transport processes that carry out the lateral exchange of material are discussed in detail below.
Fate and Transport Processes in Riparian Areas
Riparian vegetation indirectly influences biogeochemical cycling through transpiration and other effects on water flow. However, it has more direct effects, including uptake or excretion of solutes by roots as well as symbiotic associations with bacteria or fungi that stimulate important biogeochemical reactions. Several characteristics of riparian areas make them important sites for subsurface transformations of nutrients and other chemicals. Riparian soils (defined topographically as valley bottom areas that tend to become saturated during storms) possess greater soil N concentrations, higher-quality particulate organic carbon (as measured by C:N ratio), and greater overall microbial activity than do ridge and slope area soils (Garten et al., 1994). In addition, riparian soil water and dissolved organic carbon are flushed to streams much more quickly than hillslope soil water (Boyer et al., 1997).
Numerous studies have investigated the role of grassed and forested riparian areas in controlling the transport of sediment, nutrients, pesticides, metals, microorganisms, and other pollutants to receiving waters, using buffers both within and upslope of riparian areas. Although the results of these studies are highly variable in terms of “pollutant removal” or “trapping,” they have greatly improved our understanding of the mechanisms controlling transport and fate in riparian areas, especially for sediment and nutrients. The major physical, chemical, and biological fate and transport processes associated with riparian areas include infiltration, deposition, filtration, adsorption, degradation, and assimilation. Figures 2-14(A) and 2-14(B) show the important fate and transport processes for nitrogen (which is generally dissolved) and phosphorus (which is generally bound to sediment), respectively, in riparian areas.
Infiltration (also referred to as percolation) is a primary transport process during which water and dissolved chemicals and particulates enter the subsurface. Infiltration is important because it decreases the volume of overland flow, thus reducing the aboveground transport of chemicals and particulates. Once in the soil profile, pollutants are often removed or degraded by a variety of physical, chemical, and biological processes. Infiltration is one of the more easily quantifiable mechanisms affecting the performance of riparian areas that are to be utilized for removing chemicals. Thus, many constructed riparian buffers are de-
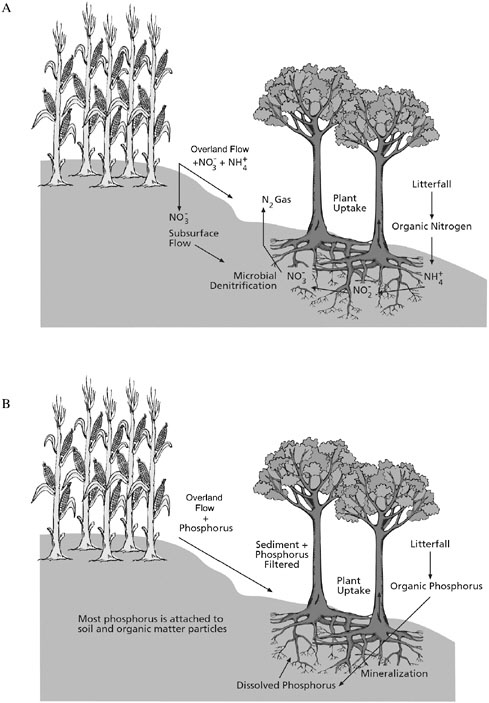
signed to maximize the infiltration of precipitation from a storm (Midwest Plan Service, 1985). (This approach necessitates large land requirements, especially if other removal processes are ignored.)
The dense herbaceous vegetation or litter layers of riparian areas offer high resistance to overland flow and decrease its velocity immediately upslope and within riparian areas. This reduction in velocity can promote deposition of sediments formerly suspended in hillslope runoff. Sediment-bound chemicals may be removed from overland flow during the deposition process. However, it should be noted that some trapped chemicals, such as organic nutrients, may be released into overland flow in dissolved form. Several mathematical models exist to describe deposition of sediment and sediment-bound chemicals in riparian areas (Hayes and Hairston, 1983; Lee et al., 1989; Inamdar et al., 1999).
Filtration of solid particles by vegetation and litter during overland flow and adsorption of dissolved chemicals and microorganisms to soil and plant surfaces are not well understood. Filtration is more significant in trapping larger soil particles, aggregates, and particulate organic matter, while adsorption to clay and organic matter in soils is more effective in trapping dissolved compounds with positive charges, such as orthophosphorus, heavy metals, and some pesticides. Adsorption of chemicals to the soil surface during overland flow is probably not very significant because of the short contact time and because adsorption sites are likely filled with previously adsorbed molecules (Dillaha et al., 1989).
Because the soils of riparian areas are generally enriched with root biomass and organic matter and have diverse soil microbiology, they support a myriad of biological processes that can transform chemicals dissolved in the subsurface. For example, the enhanced biological activity of riparian areas includes the microbial degradation of organic pesticides (USDA, 2000) and petroleum products (Brock and Madigan, 1991, p. 654). Both plants and soil microorganisms can assimilate large amounts of dissolved subsurface chemicals, particularly nutrients. Plant uptake can lead to either short- or long-term nutrient removal, depending on whether nutrients are stored in woody biomass that is retained at the end of the growing season or lost as leaves and twigs that return to the soil surface (Lowrance et al., 1995; Correll, 1997). The recycling of nutrients through plant uptake and release via decomposition contributes to keeping nutrients in the riparian area rather than releasing them to an adjacent waterbody. To maintain active nutrient assimilation, plant biomass must be removed, as is sometimes done in managed riparian areas where fast-growing trees are harvested for lumber and grasses and herbs are harvested for forage and biomass (Schultz et al., 1995). In addition, whether assimilation by vegetation occurs from the unsaturated zone is critical in determining whether there is an effect on flowpath chemistry (the saturated zone). Nonetheless, assimilation by plants is not a widely exploited mechanism of nutrient removal because the process has not been well described analytically.
Denitrification is a transformation process in which soil microorganisms take up dissolved nitrate from subsurface water and convert it to nitrogen gas. It is believed to be highly effective in removing nitrate from subsurface flow where conditions are favorable, that is, where nitrate-laden groundwater flows through areas that are both enriched with organic matter and anaerobic. Denitrification may also occur under aerobic or unsaturated soil conditions in localized soil micro-sites with high concentrations of particulate organic carbon (Parkin, 1987). Given their ample organic matter and diverse microbiology, it is not surprising that riparian areas support denitrification (Groffman et al., 1992; Addy et al., 1999), with riparian forests reported to remove 30–40 kg N ha–1 yr–1 under suitable conditions (Lowrance et al., 1995). Denitrification also occurs at seeps, where groundwater comes to the surface through soil horizons that are enriched with organic matter, as well as in wetlands, shallow groundwater, and other areas where substantial amounts of organic matter exist under saturated conditions. Schade et al. (2001) demonstrated the importance of denitrification over plant assimilation as a mechanism for removing nitrate from shallow groundwater. In this case, organic carbon produced by riparian shrubs acted as an energy source to drive denitrification. Despite the popularity of managing riparian areas to enhance denitrification, there is some doubt as to their importance in removing nitrate in certain agricultural settings. For example, Bohlke and Denver (1995) showed that on the Delmarva Peninsula, contact of groundwater flow paths with geochemically reducing sediments at the base of the shallow aquifer, prior to discharge through riparian areas, was more important in accounting for denitrification.
Sites of Fate and Transport Processes. Because the transformation processes described above require contact between chemical-laden water and either riparian vegetation or microbes in soils and sediment, their extent is obviously limited in instances where groundwater passes below the biologically active riparian area. Such bypassing can occur when riparian soils have low hydraulic conductivity (compared with a sand or loam) because of a relatively large proportion of fine sediment such as clay, silt, or humified organic matter. The extent of bypassing, and thus the ability of riparian areas to support chemical transformation such as denitrification, depends on many factors such as antecedent moisture, soil texture, underlying aquifer or bedrock geology, and human-induced landscape and channel conditions (Gilliam et al., 1997). Transformation mechanisms that occur in the unsaturated zone, such as some plant assimilation of nutrients, will have little effect on groundwater chemistry.
Although the preceding processes have been described as occurring in the subsurface water moving through riparian soils, they also occur in slackwater habitats (i.e., shallow and slowly moving sections of surface water channels) and hyporheic zones. For example, nitrate is removed from flowing water via denitrification within hyporheic zones (Hinkle et al., 2001). A significant proportion of nutrients (both dissolved and particulate) and inorganic and organic components
are associated with sediments of the hyporheic zone as opposed to the active river channel (Pinay et al., 1992, Vervier et al., 1992, 1993). Periphyton, benthic algae, leaf packs, microbes, and some of their consumers are also present in varying degrees within the sediment.
Contact Time. Longer contact times are required for the transformation of some nutrients and other dissolved substances in overland and shallow subsurface flow. Thus, the extent to which these mechanisms occur is dependent on the amount of time that runoff and associated chemicals are retained in the riparian area, which is in turn largely a function of hydrology. Overall pollutant removal occurs to the greatest extent when overland flow and shallow subsurface flow are distributed uniformly across the riparian area. When overland and shallow subsurface flows concentrate and flow through only a portion of the riparian area (which is dependent partly on local topography), the areas with concentrated flow have shorter detention times, and their transformation mechanisms may be overwhelmed. Activities such as agriculture, silviculture, mining, and urbanization tend to concentrate flows (via gullies, channels, and subsurface tiles—see Chapter 3), such that only a small fraction of the riparian area’s chemical and particulate trapping potential is realized (Dillaha et al., 1989).
Role of Stream Order. The importance of biogeochemical transformations, in terms of preventing pollutants from reaching adjacent waters, diminishes as one goes from ephemeral and first- and second-order streams to larger, higher-order streams. A greater portion of the flow passes through riparian areas along low-order streams before reaching the channel network, making their riparian areas more instrumental in removing pollutants from runoff. In contrast, most of the flow in high-order streams comes from low-order stream channels, and only a small portion of the flow in high-order streams actually crosses the riparian areas associated with the high-order stream segment. This suggests that if water-quality protection is a primary objective, priority should be given to restoration of functional riparian areas along ephemeral and first- and second-order streams over larger, higher-order streams (similar to the conclusion reached by Brinson (1993) for wetlands used for water-quality protection). As shown in Figure 2-15, first-and second-order stream channels comprise the vast majority of all stream kilometers in a given watershed (Leopold et al., 1964). It should be noted that the role of stream order is less clear for transformation via hyporheic exchange and the passage of water into and out of the riparian area during longitudinal flow.
Finally, it must be remembered that chemical transformation/ removal is just one function of riparian areas (although frequently the primary target of management actions in agricultural areas—see Chapter 5). Even when specific riparian areas are only marginally effective for pollutant removal, they are still essential for wildlife habitat, flood control, and many other environmental services as described below.
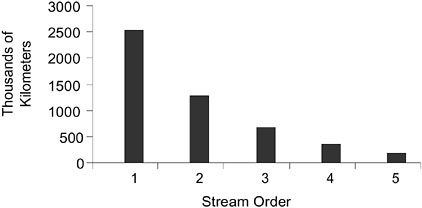
FIGURE 2-15 Relationship between stream order and stream length. Reprinted, with permission, from Leopold et al. (1964). © 1964 by W.H. Freeman and Company.
REGIONAL CLIMATE AND RESULTING RIPARIAN VEGETATION
The streamflow and associated fluvial and sediment processes that shape riparian areas are the products of regional patterns of topography and geomorphology, climate, and runoff. All these factors combine to create the observed distribution of riparian vegetation across the United States.
Climate
Climate has a strong influence on the structure and functioning of riparian areas, mainly through temperature and precipitation. These in turn strongly influence two other factors—evapotranspiration (ET) and runoff. ET refers to the surface water that moves from the liquid phase to water vapor through transpiration by plants and evaporation. (Potential evaporation (PET) is the evapotranspiration that would occur if water were not limiting.) Runoff is water that escapes both evapotranspiration and transport to deeper aquifers, finding its way to streams via overland and shallow subsurface pathways. Regional variation in these parameters contributes to the diversity of riparian vegetation observed across the United States.
Incoming solar radiation from sunlight influences air temperature, precipitation, and the subsequent apportioning of precipitation into evapotranspiration, subsurface recharge, and watershed runoff. Within the contiguous 48 states, solar radiation exhibits significant seasonal variability. Short days, low sun angles, and significant cloud cover result in low solar radiation inputs during winter months. In contrast, long days, high sun angles, and less cloud effects during the summer-
time combine to more than double the solar radiation of the winter months for many areas. Although the seasonal pattern of incoming solar radiation to the atmosphere varies little from year to year, clouds, which are highly variable, reduce the actual amount and timing at any particular point on the earth. Finally, aspect and slope further influence the amount of energy available at a particular location: during clear sky conditions, south-facing slopes receive more incoming solar radiation per unit area than do north-facing slopes (Reifsnyder and Lull, 1965).
Annual precipitation is highest in the eastern United States, particularly along the Gulf of Mexico coast, with up to 155 cm in Tallahassee, Florida (Figure 2-16). In general, precipitation is 90 cm or more to the east of a line extending from the mouth of the Mississippi River to the western shores of the Great Lakes and is distributed evenly throughout the year. (South Florida, with a pronounced winter dry season and summer wet season, is an exception.) The major moisture source is the Gulf of Mexico, even as far north as New England, but east of the Appalachian Mountains, the Atlantic Ocean is also significant. The lowest annual rainfall in the eastern United States usually exceeds 50 percent of the long-term mean; thus, severe droughts are uncommon. Compared to more arid regions, the eastern United States is characterized by gradual gradients in hydrophytic vegetation away from streams.
West of the Mississippi, the Gulf of Mexico is a less significant source of moisture, and north–south mountain ranges reduce the supply of Pacific moisture reaching the region, especially in the valleys. The western half of the Great Plains, the Great Basin, and the Southwest are mostly semiarid or arid. Non-coastal areas of the Southwest have with a strong summer rainfall maximum. The summer precipitation often comes in heavy thundershowers, and runoff generally travels as overland flow to nearby streams with little infiltration into the soil. Thus, little precipitation becomes available to recharge groundwater. The Sonoran Desert has a significant but smaller winter rainfall maximum arising from storms of Pacific origin. Because of the scarcity of rainfall, desert scrub ecosystems are distributed as far north as the Canadian border (49° N) and beyond, even at elevations of 2,000 m. In these climates, the gradient of vegetation with distance from streams is steep, and the well-developed riparian vegetation is usually striking in contrast to the surrounding desert landscape.
Finally, the Pacific Coast has a Mediterranean or modified Mediterranean climate, with a strong precipitation maximum during winter when cool temperatures limit plant growth. This seasonal peculiarity results in a strong moisture deficit for plants during the summer growing season. Along the coast, annual rainfall ranges from about 25 cm in the south to well over 100 cm in the north, and it can exceed twice that amount in coastal mountains and along the western slopes of the Sierra Nevada–Cascade range. The Pacific Coast and the Southwest have much greater variation in annual rainfall than does the eastern United States.
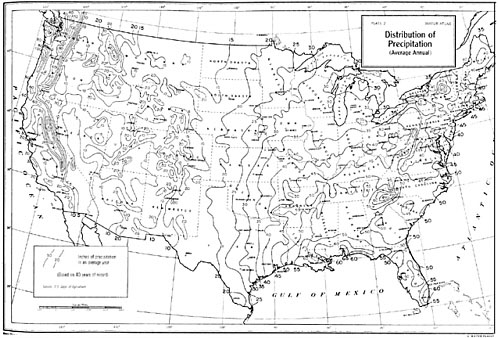
Except in the extreme northern part of the Pacific Coast, the lowest annual rainfall is often considerably less than 50 percent of the long-term average.
Across the United States, mean annual temperature generally decreases from south to north and with increasing elevation. Temperature variability, especially the range between daily and seasonal highs and lows, increases with distance from the ocean, especially the Pacific. Both temperature and precipitation vary with latitude, elevation, and proximity to large waterbodies or mountains.
These regional patterns of solar radiation, precipitation, and temperature, combined with elevation and other local topographic features that affect runoff, result in a highly complex pattern of “life zones” (as illustrated in Plate 2-2). Because of higher variability in precipitation and temperature in the West, well over 75 percent of the possible life zones are found west of the Mississippi River.
Runoff
Runoff describes the flow of water from the terrestrial landscape to surface-water bodies (e.g., stream channels, ponds, and lakes). It comprises all of the water that moves quickly to channels without being evaporated or stored for significant periods in soils and groundwater. Long-term runoff patterns from watersheds, which are reflected in streamflow, play an important role in shaping riparian systems. Streamflow amounts from a given watershed often exhibit general patterns from year to year as a result of topography, soil type, geology, vegetative cover, and the watershed’s climate. Figure 2-17 shows the mean annual runoff in the United States.
The variation in runoff between watersheds can be explained by the intensity of precipitation, season of maximum precipitation relative to evapotranspiration, drainage area, land slope, soil and geologic characteristics, and vegetation type (Gregory and Walling, 1973). These factors determine the extent of infiltration versus overland flow, the proportion of soil water that is evaporated, the flow paths of precipitation across or through soils, and the proportion of soil water that is recharged to groundwater. Together, they affect the timing and rate of water delivery to channels, as well as the total runoff.
To evaluate the relative importance of precipitation and evapotranspiration to runoff amounts, these two variables are sometimes combined into a single factor called excess precipitation, defined as the difference between precipitation and evapotranspiration. Another common way to portray this concept of excess precipitation is the difference between precipitation and potential evaporation, or P–PET (Plate 2-3). In general, the percentage of precipitation that occurs as runoff is highest where excess precipitation is greatest, e.g., in the Northeast, Northwest, and Upper Midwest, and in high mountain areas. In those areas, the proportion of incoming precipitation that becomes runoff is typically greater than 50 percent. Excess precipitation is more moderate in the Southeast and Midwest. In the Great Plains, Great Basin, and Southwest, excess precipitation is at or near
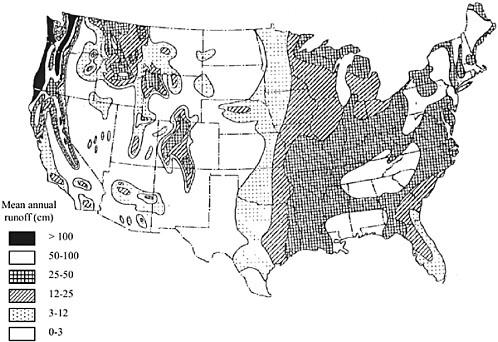
FIGURE 2-17 Mean annual runoff in the conterminous United States. SOURCE: Reprinted, with permission, from Satterlund and Adams (1992). © 1992 by John Wiley & Sons, Inc. NOTE: The white sections in the far Pacific Northwest and East fall into the mean annual runoff category of 50 to 100 cm, while those white sections located in the Great Basin, the arid Southwest, and the Plains states fall into the category of 0 to 3 cm.
zero and P–PET is negative, and consequently runoff percentages are typically well below 20 percent.
Seasonal variability in runoff reflects the timing and type of precipitation, the evapotranspiration rates throughout the year, and the intensity and duration of storms. Winter and early-spring storms in the East are longer in duration than in the West and occur when there is greater antecedent soil moisture, causing highest runoff in late winter or early spring despite fairly even distribution of precipitation throughout the year. Accumulation of snowfall and its subsequent melting are important factors influencing runoff patterns in cooler regions and where streams in the West have their headwaters in mountain ranges. Summer thunderstorms are dominant contributors to runoff in western mountains, the arid Southwest, and the Great Plains because winter precipitation is often low and because sparsely vegetated surfaces contribute a greater percentage of precipitation to overland flow. Together, these climatic factors determine patterns of peak runoff that become progressively later in the year from east to west across the United States. High-flow periods also tend to be shorter in duration the farther west one goes, as the dependence on summer thunderstorms increases. These general patterns of runoff variability do not include the Pacific Coast, where the timing of runoff is more similar to that of the Northeast.
The interannual variability of runoff, which in part determines the disturbance regimes characteristic of riparian areas, is highest in the Southwest (>100 percent), less in the Great Plains, Great Basin, and Midwest (>50 percent), and lowest in the Northwest, Rocky Mountains, and eastern United States (<30 percent) (Figure 2-18). Although in any given year and watershed the pattern of runoff can be relatively unique, over periods of multiple years, runoff patterns for specific watersheds tend to converge where lithology, soils, elevations, vegetation, and climatic inputs are similar.
Topographic and Geomorphic Patterns
In addition to climate and runoff features, there are important topographic and geomorphic patterns that lead to the observed variability in riparian areas across the United States. As shown in Figure 2-19, there are distinct types of riparian areas depending on the shape and steepness of the terrain, the composition of the underlying geologic materials, and their relative position within the stream network. For example, headwater streams tend to be associated with relatively steep watersheds and often have streams or rivers that are laterally constrained by hillslopes or geologic formations. Constrained streams have limited capability to adjust to changes in flow and sediment delivery rates. However, in a down-valley direction, the deposition of alluvial sediments along valley bottoms and lowlands decreases hillslope and geologic constraints on a stream’s lateral movement. Hence, unconstrained streams tend to have a greater sinuosity and are
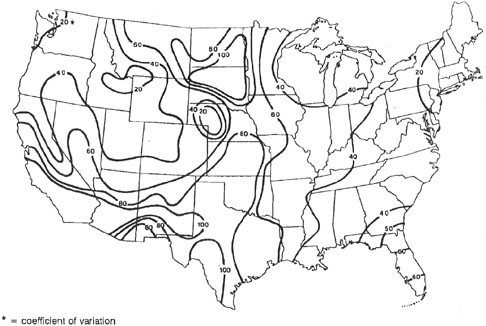
FIGURE 2-18 Interannual variability of runoff, as measured by coefficients of variation for runoff from the conterminous United States. SOURCE: Reprinted, with permission, from Patrick (1995). © 1995 by John Wiley & Sons, Inc.
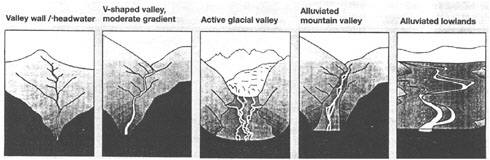
FIGURE 2-19 Generalized pattern of stream systems occurring across a range of terrain types. SOURCE: Reprinted, with permission, from Naiman et al. (1992). © 1992 by John Wiley & Sons, Inc.
more capable of adjusting to varying patterns of flow and sediment transport from upstream sources.
Position in the stream network plays a major role in riparian area structure because riparian areas are highly responsive both to the hydroperiod of the adjacent waterbody (e.g., depth, frequency, duration, etc.) and the source of the water (marine versus freshwater)—characteristics which differ among headwater streams, larger-order rivers, lakes, and estuaries. In many cases, these forces can be more important than climate in determining riparian area structure. For example, salt-influenced tidal marshes are more structurally and functionally distinct from floodplain forests within the same climate zone than are two salt-influenced tidal marshes, one in an arid climate and the other in a humid climate.
There have been various attempts at characterizing regions of distinct topography, hydrology, and geomorphology in order to provide an improved perspective regarding the regional structure of riparian areas. For example, Winter et al. (1998) and Winter (2001) define a “landscape unit” (called a basic building block of all landscapes) as simply an area of uplands adjacent to an area of lowlands, with the two areas being separated by an area of steeper terrain. The landscape unit’s hydrology is determined by hillslope gradients, soil permeability, dimensions and permeability of the geologic framework, and atmospheric-water exchange, which is controlled by climate. Conceivably, all of the more complex hydrologic landscapes that are evident in the United States are variations or multiples of these fundamental landscape units. Some examples include (1) mountainous terrain, with narrow lowlands and uplands separated by steep valley sides, (2) a basin and range landscape, with very wide lowlands separated from much narrower uplands by steep valley sides, and (3) plateaus and high plains consisting of narrow lowlands separated from very broad uplands by valley sides of various slopes (Winter et al., 1998).
The purpose of the hydrologic landscape unit classification is to characterize pathways and rates of water movement through landscapes. For example, if a landscape has low land slope and low-permeability soils, overland flow will be slow and recharge to groundwater will be limited. In contrast, if the soils are permeable, overland flow may be limited but subsurface flow and groundwater recharge will be high. The key variables used are the distribution of landscape relief (maximum minus minimum elevation), average slope, slope distribution (percent flatland and percent upland or lowland), geologic texture and permeability, and available atmospheric water exchange (annual average precipitation-potential evapotranspiration). Wolock (2001) used statistical analyses of the existing nationwide datasets (averaged over approximately 200-km2 watersheds) to classify landscapes across the United States, with the results clustering into 20 hydrologic landscape units (shown in Plate 2-4). Unlike with the life zone map (Plate 2-2), regional differences in hydrologic landscape units are significant across the entire country primarily because of the high variability in surface and subsurface properties.
Disturbance Regimes
A variety of natural, physical disturbances that span scales ranging from tectonic activity to localized erosion help shape riparian areas. Geologic uplift that increases or decreases channel slope can cause, respectively, greater channel incision or greater overbank flow frequencies (Burnett and Schumm, 1983). Even in tectonically stable areas, however, the primary influences on the structure of riparian plant communities are fluvial processes—in particular, floods and the associated transport of sediment within streams. Floods and overbank flows occur when stream discharge exceeds channel capacity. Where snowmelt is an important contributor, rain-on-snow events produce exceptional floods. At lower elevations, high-intensity thunderstorms are of great importance in generating floods, particularly in the Southwest and Great Basin. Floods along the Gulf of Mexico and the southern Atlantic Coast are often associated with landfall of hurricanes and tropical depressions.
As shown in Figure 2-20, fluvial disturbances play a significant role in determining the composition of riparian vegetation by controlling the germination and successful establishment of seedlings as well as their long-term survival. The recruitment of woody riparian species in particular is dependent on interannual variability in flooding, channel migration, and sediment deposition. For example, germination of cottonwoods and some willow species generally occurs on locally scoured beds following channel adjustments such as channel narrowing caused by reduced flows (Figure 2-20B) or a flood pulse (Figure 2-20D). Whether the seedlings are established and reproduce depends on future disturbance events; plants may perish under drought conditions or they could be scoured away during subsequent floods or winter ice flows. Only plant species capable of tolerating these disturbances are likely to survive over the long term. For example, the vertical accretion of flood-deposited sediment may result in burial of the root crowns of trees (Scott et al., 1996). This is a selective process that allows survival of tree populations that are capable of producing new root systems when their stems become buried. For actively meandering rivers (Figure 2-20C), large-scale sloughing occurs when flows locally undercut forest vegetation positioned on the outside of meander curves. At the same time, point bar formation and vertical accretion on the inside of meander curves provide substrates for seedling, and ultimately forest establishment (Friedman and Auble, 2000). Where lateral channel migration is active, the rate of movement can be calculated from increases in tree age along transects perpendicular to the inside of the meander curve (Everitt, 1968).
The microtopographic variation created by diverse fluvial processes supports a species richness that would not otherwise occur (Gregory et al., 1991). In more humid areas where precipitation is higher and more evenly distributed throughout the year, flood intensity is generally lower and flood duration longer than in arid climates. In such situations, vegetation tends to be more effective in
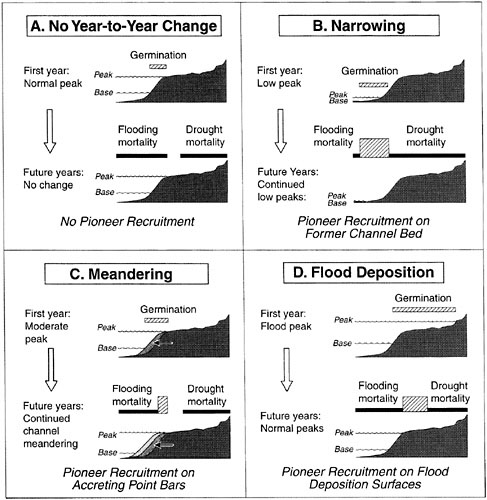
FIGURE 2-20 Hydrogeomorphic control of recruitment of woody pioneer species. Seed germination, early seedling mortality, and tree recruitment are shown in relation to annual high and low flow lines along four bottomland cross sections (A–D). In each of the four situations, the cross-hatched area in the upper part indicates the zone of seedling establishment, and the cross-hatched area in the lower part indicates the zone of long-term survival. (A) In the absence of interannual flow variability and channel movement, there is little or no tree recruitment. (B) On a narrowing channel, there is recruitment on the former channel bed. (C) Recruitment on point bars is typical of a meandering river. (D) Tree recruitment at high elevations is associated with infrequent floods and no channel movement. SOURCE: Reprinted, with permission, from Friedman and Auble (2000). © 2000 by Cambridge University Press.
stabilizing channel migrations, thus leading to a diminished zone of disturbance (Figure 2-21A). In contrast, rivers in more arid regions experience flood discharges that can be orders of magnitude greater than their base flows (which may be zero in some cases), resulting in significantly greater flow variation and physical disturbance. In arid regions disturbance-tolerant trees may be found along the entire cross section of riparian areas (Figure 2-21B).
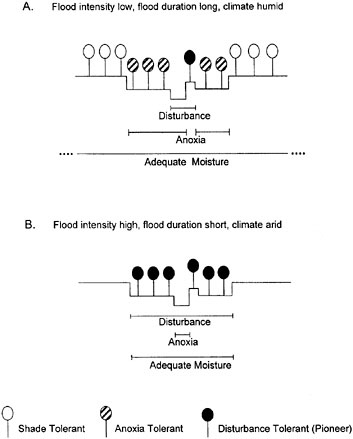
FIGURE 2-21 Influences of environmental stresses on the distribution of riparian trees. (A) Cross section of typical bottomlands in the low-gradient streams of the Coastal Plain of the southeastern United States. (B) Cross section typical of the bottomlands in the western Great Plains. Bars under cross sections show the extent of physical disturbance, anoxia, and adequate moisture for survival of trees. Although the three tree strategies are presented as distinct, many species combine strategies. The term “shade tolerant” refers to trees that are able to reproduce in the shade of other trees in the absence of physical disturbance. SOURCE: Reprinted, with permission, from Friedman and Auble (2000). © 2000 by Cambridge University Press.
Physical disturbance from trees, ice floes, boulders, and other solid objects in floodwaters can be effective in limiting vegetation to those species of very short stature, especially immediately adjacent to channels. Ice floes have a profound effect in boreal and arctic Alaska where ice floating downstream removes vegetation (Bliss and Cantlon, 1957; MacDonald and Lewis, 1973). During hurricane-generated flows that pass directly over the Luquillo Experimental Forest in Puerto Rico, slope failures and torrential stream discharge transport whole trees and meter-diameter boulders to riparian channels, creating disturbance that makes riparian forests less species-rich than surrounding upland forests (Ahmad et al., 1993; Scatena and Lugo, 1995). Nonetheless, it is clear that floodwater by itself can effectively eliminate vegetation, as evidenced by areas of sparsely colonized floodplains in the arid Southwest (Zimmerman, 1969). An extreme example of the long-term influence of fluvial processes is the elimination of forest areas by channel migration and the subsequent isolation of these forests in the tectonically active regions of upper Amazonia (Kalliola et al., 1991). These processes have been proposed as a speciation mechanism for forest plants on evolutionary time scales (Råsånen et al., 1987).
Fire is a physical disturbance predominantly in uplands, but it can become significant in arid riparian areas when drought conditions develop. In fact, fire is second only to flood events as a disturbance agent affecting riparian vegetation in northern Montana (Lee, 1983). In riparian areas along the southern California coast, where fire was historically only of minor importance, the introduction of a highly fire-tolerant grass (Arundo donax) in floodplains has led to the eventual elimination of the more fire-intolerant valley oaks that originally dominated (Boose and Holt, 1999). Such replacement tends to reduce the complexity of riparian forests with predictable consequences for nesting birds, shade-adapted plants, and other tree-dependent organisms.
Change in relative sea level is yet another, albeit more chronic, natural disturbance that causes changes in coastal riparian areas. Where sea level is rising in relation to the land surface, both inundation and salinity intrusion cause coastal riparian areas to change from forest to salt marsh vegetation (Brinson et al., 1995; Williams et al., 1999). As sea level rises, riparian areas migrate landward and replace upland forest while the estuarine shoreline erodes. Depending on the relative rates of forest replacement and shoreline erosion, the width of riparian areas can increase, decrease, or remain at steady-state. On islands or on coastlines where impediments to landward migration interfere, eroding shorelines caused by rising sea level will eventually eliminate riparian areas (Figure 2-22). The Mississippi alluvial valley represents a large-scale and long-time span case of downcutting and alluvial filling forced by multiple fluctuations in sea level during the Pleistocene (Fisk and McFarlan, 1955). Although the Mississippi example is largely irrelevant to most current ecological and socioeconomic concerns, it emphasizes how great a role sea level changes and tectonic activities played in eventually establishing today’s distribution of vegetation in estuarine
FIGURE 2-22 Response of coastal riparian areas to rising sea level. As the shoreline moves landward, riparian areas may migrate upstream or overland, or be eliminated as shorelines of islands erode. Vertical arrows represent the accumulation of sediment in pace with rising sea level, which must occur or riparian areas will be eliminated; horizontal arrows indicate shoreline erosion. SOURCE: Reprinted, with permission, from Brinson (1991). © 1991 by Dr. Douglas A. Wilcox, Editor-in-Chief, Wetlands.
riparian areas. Sea level changes and tectonic activity continue to shape the vegetation of high-latitude regions where continental margins are rising relative to sea level (Jordan, 2001). In these cases, coastal vegetation becomes increasingly isolated from tidal and salinity effects—the opposite of the more common landward “migration” of riparian areas.
Moisture Availability and Anoxia
Although physical disturbances are important in higher-order streams with strong flows and in headwater streams with steep gradients, soil moisture and
depth to the water table also influence the composition of plant communities. Riparian areas in humid climates at the lowest elevations commonly experience such frequent flooding that adequate moisture for plant growth is generally found across the entire riparian area (Figure 2-21A). In such cases, soil anoxia brought on by persistent flooding or saturation is a greater factor in controlling riparian species composition than is physical disturbance. Anoxia is particularly prevalent in the fine sediment grains typical of low-gradient downstream reaches, which tend to retain moisture much more efficiently than coarse sediment. Some of the most flood-tolerant species in the United States (e.g., Taxodium distichum, Nyssa aquatica) can tolerate constant soil saturation, although they require a drawdown period for seed germination and seedling establishment. Where trees cannot establish because of continuous flooding, marshes and submerged plant communities become established. Beaver may facilitate this process by both felling trees and creating continuous inundation. Thus, while structural differences in vegetation between riparian areas and uplands may not be readily apparent in humid climates, species composition is very different.
Riparian areas in arid regions, on the other hand, are in close proximity to the only significant sources of water available for tree growth. Thus, anoxia is extremely limited and is of negligible importance in influencing species composition compared to overall moisture availability and disturbance patterns (Figure 2-21B). The narrow band of available water along streams in arid regions results in a forested riparian area of pioneer species, such as willow and cottonwood, that stands in stark contrast to the surrounding more sparsely vegetated uplands.
The different gradients of plant species richness brought about by moisture availability are illustrated in Figure 2-23. In this case, the number of species found within the riparian area of arid Sierra Nevada streams is markedly higher than in adjacent uplands, while species richness throughout the more humid Cascades riparian areas and uplands is relatively uniform (although species composition is not identical).
Regional Riparian Vegetation
The characteristics of riparian vegetation vary substantially across the United States and correspond with geographic variation in climate, hydrologic regime, and associated geomorphology. For reasons discussed earlier, there are major differences between the riparian vegetation found in humid versus arid regions. In addition, within most of the geographic regions described, riparian vegetation types at low elevations differ from those in high-elevation mountainous regions. The vegetation of riparian areas also varies according to the kinds of terrestrial and aquatic ecosystems they connect. Salinity and tidal inundation influence riparian vegetation adjacent to estuarine and marine waterbodies. Shorelines of lakes often undergo interannual fluctuations in water level, forcing riparian plants to constantly shift their positions. In riverine settings, sedimentation and erosion,
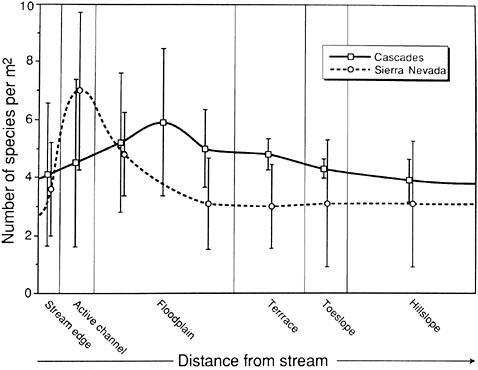
FIGURE 2-23 Gradients of plant species richness along lateral transects from the stream channel to upper hillslopes along three streams on the west slope of the Cascade Mountains of Oregon (more humid) and three streams in the Sierra Nevada of California (more arid). SOURCE: Reprinted, with permission, from Gregory et al. (1991). © 1991 by American Institute of Biological Sciences.
moisture availability, and the frequency, duration, and intensity of floods are selective factors that help shape the spatial patterns and species composition of riparian plant communities. The dynamic nature of many stream channels leads to cyclic changes in vegetation as the result of floodplain erosion and point bar deposition.
Riparian areas often support woody vegetation, and forests are a common community structure. There are notable exceptions, however, particularly along arid-zone, high-altitude, and high-latitude streams, where graminoids and shrubs are prevalent. Further, many forested riparian areas have herbaceous wetlands embedded within them and as a component of the forest floor. It is this complexity of vegetation types that contributes to the enormous richness of riparian areas in comparison to uplands, particularly in arid regions (Gregory et al., 1991; Brinson and Verhoeven, 1999). Thus, in the Great Plains and the arid and semiarid West, the contrast in species composition and physiognomic structure between riparian areas and uplands is large (Figure 2-23). In north-central Okla-
homa, riparian forests have a much higher species diversity than adjacent upland forests, while the opposite is true for corresponding studies in the eastern deciduous forest region (Collins et al., 1981). Outside the United States, riparian areas have also been shown to harbor more diverse plant communities than uplands on a regional scale. For example, at least 260 species (or 13 percent) of the total Swedish flora occur along the Vindel River, which holds the country’s record of 131 species per 200 m length (Nilsson, 1992). The diversity in riparian vegetation stems not so much from species richness within individual plant communities but rather from the existence of broad plant community types as well as age and structural diversity within plant communities. This large-scale heterogeneity is a result of the extreme conditions of disturbance, wetness and dryness, and other fluvial processes characteristic of riparian areas. For example, in the Saskat-chewan River delta in Canada, none of the plant communities is particularly species rich (Dirschl and Coupland, 1972). Yet the delta vegetation was classified into 11 broad plant community types according to physiognomy, with six of these consisting of woody vegetation (white spruce-hardwoods forest, black spruce-tamarack forest, tall willow-alder shrub, medium willow shrub, low willow shrub, and bog birch shrub). The remaining five community types were dominated by herbaceous wetland vegetation. Even in arid regions where riparian vegetation represents the only forest structure in the landscape, tree species diversity is low, but age class and structural diversity are typically high.
Because of these sources of biodiversity, regional differences among riparian areas are best illustrated by comparing the plant community and species compositions rather than determining that one region is more species-rich than another. The following section focuses on the dominant woody component of riparian areas to illustrate regional differences, organized around the broad geographical areas depicted in Figure 2-24. Because of the lack of equivalent and collated information on other riparian plant types, only trees are covered comprehensively. It should be noted that most riparian areas exhibit lateral zonation of species, something that cannot be addressed at large regional scales and as such is not part of this summary.
Boreal and Arctic Alaska
Boreal (interior) Alaska extends north from the maritime climate of the southeastern part of the state to the Brooks Range, which separates the interior from the Arctic, or North Slope. Although precipitation in the interior and the North Slope is relatively low, a combination of a short growing season, low evapotranspiration, and low topographic gradients results in excess moisture and saturated soil. Black spruce dominates the uplands of interior Alaska (Post, 1996), while riparian vegetation fluctuates in response to the dynamics of ice scour, active channel meandering, and sediment deposition. Where ice floes have removed vegetation, willow and poplar form low-growing communities (though
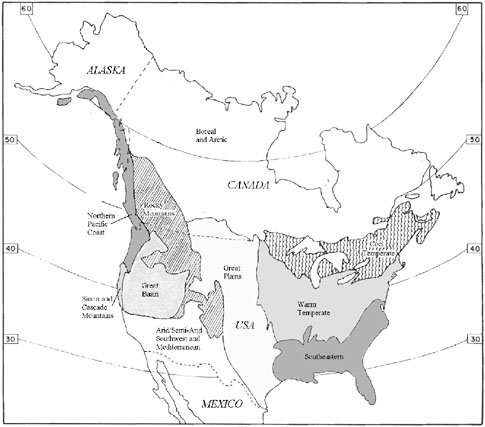
FIGURE 2-24 Vegetation regions of the United States.
willow is short-lived) (Walker et al., 1986). Alder also colonizes bare soil, but is eventually replaced by a continuous deciduous canopy of balsam poplar in the absence of further disturbances. Next, productive stands of white spruce develop on the rich alluvial soils, but eventually organic matter and a moss ground cover accumulate. This insulating layer subsequently modifies the soil microclimate so that permafrost develops and reduces rates of nutrient cycling, at which point, given enough time, a black spruce forest replaces white spruce, superficially resembling the upland forests (Van Cleve et al., 1991).
Riparian vegetation in arctic Alaska is generally shorter than that of the boreal zone. A range of community types are possible: a pioneer herbaceous community; a feltleaf willow stage (up to 6 m in height); a zone of deteriorating feltleaf willow with increasing prominence of greenleaf willows, mosses, and herbs; and an alder–willow stage, representing the oldest and most elevated condition (Bliss and Cantlon, 1957). Unvegetated sand and gravel bars are extensive
because of disturbance by ice floes, channel meandering, and rapid sediment deposition. Species richness of vegetation along arctic river corridors increases in a downstream direction, principally due to increased habitat heterogeneity (Gould and Walker, 1999).
Pacific Northwest and Coastal Mountains
The Pacific Northwest and coastal mountains support a diverse woody plant flora with a high percentage of endemic species, particularly in the California Floristic Province. The species dominating riparian forests in California are related to air temperature, groundwater depth and aeration, and frequency and intensity of disturbance (Holstein, 1984). At lower elevations and in the south, Fremont cottonwood is often the dominant riparian tree. At higher elevations or in the north, dominance shifts to black cottonwood. Where disturbance is more severe, however, narrowleaf willow or Goodding’s willow is common. White alder is abundant in areas of shallow, well-aerated groundwater in central California, particularly along montane streams. Further north, red alder forms dense riparian stands, but it can also grow in the uplands in the wetter climate. Where groundwater is deeper but still aerated, the dominant tree is often California sycamore. Where groundwater is deeper and not well aerated, California white oak—a deciduous oak endemic to the Central Valley and adjacent foothills—once grew in large stands.
Several other tree species grow in riparian areas of California and give the region a distinctive floristic character (Holstein, 1984). Boxelder grows primarily as a riparian species, while bigleaf maple, which is common in riparian areas, also grows in uplands. There are two species of riparian ash—velvet ash in the south and Oregon ash in the north. Another riparian species endemic to California is Hind’s walnut. In several areas, California laurel and coastal redwoods are abundant riparian species.
The riparian vegetation of western Oregon and Washington is closely related to that of northern California and southeast Alaska; one of the most characteristic trees is black cottonwood. Black cottonwood forests are especially prominent along large rivers, where several species of willow form a thick forest understory (Franklin and Dyrness, 1988). Oregon ash is also abundant in the poorly drained soils of riparian areas in the interior valleys of Oregon; these areas also include populations of bigleaf maple, Oregon white oak, red alder, and localized populations of California laurel. Buttonbush, which grows in riparian areas of the humid eastern United States, is represented by relictual riparian populations along the Cosumnes River in the southern Sacramento Valley. Water birch is a common riparian tree in a few restricted areas of the Klamath Mountains and the southeastern Sierra Nevadas.
Great Basin
The Great Basin is a vast area of the western United States characterized by predominantly basin topography, relatively low precipitation, and low streamflow. Warm summers and cold winters result in an extensive cover of sagebrush and associated shrubs, sometimes referred to as cold desert. In the lower elevations of the Great Basin, riparian forests are the only forests, and their structure varies substantially with latitude (Stine et al., 1984; Minshall, 1989). In the northwestern regions, riparian forests included black cottonwood, willow, hawthorn, water birch, chokecherry, and gray alder. In the southern Great Basin, Fremont cottonwood grows in association with several willow species and mesquite. Eastern Oregon and central Washington also have little-known riparian communities of willow and hawthorn or chokecherry (Lytjen, 1998). The riparian areas of the southern Great Basin have been increasingly invaded by saltcedar.
Arid and Semiarid Southwest
In the Mojave and Sonoran deserts and in Southern California, precipitation limits upland vegetation to scattered shrubs and occasional grasslands in the lowlands and evergreen woodlands and forests in the mountains. The structure, species composition, diversity, and productivity of riparian areas contrast sharply with adjacent uplands. The riparian vegetation in much of Arizona is dominated by a few species of trees (Lowe, 1961, 1964). “Wet” riparian areas in Arizona and across most of the arid Southwest support primarily Fremont cottonwood, Goodding’s willow, Arizona sycamore, velvet ash, and Arizona walnut. Netleaf hackberry is found to be a significant riparian tree species in the Catalina Mountains (Whittaker and Niering, 1965). Similar sets of species dominate the lower-elevation riparian areas of New Mexico and Southern California.
Stromberg et al. (1996) described a strong association between the distributions of these species along the San Pedro River of southern Arizona and depth to groundwater. One result of their analysis is a classification of species by the degree to which they are dependent on wetland environments. Goodding’s willow was classified as an obligate wetland species, Arizona walnut and Fremont cottonwood as facultative wetland species, velvet ash as a facultative species, and netleaf hackberry, Texas mulberry, and mesquite as facultative upland species.1 These species represent a sequence of drought tolerance, with Goodding’s willow being the least tolerant.
In a recent classification of New Mexico wetlands, 99 different forested and shrub–scrub wetland communities are described (Muldavin et al., 2000). It is
interesting that many of the same genera found in New Mexico are also found in the humid eastern United States, i.e., alder, willow, maple, dogwood, cottonwood, sycamore, and walnut. Species common to the two regions are boxelder and plains, or eastern, cottonwood.
Riparian areas of the arid and semiarid West are increasingly dominated by exotic tree species, especially saltcedar, Russian olive, Siberian elm, tree of heaven, and white mulberry (Crawford et al., 1993; Muldavin et al., 2000). The dry riparian areas along ephemeral washes in Arizona and Mexico are dominated by a very different group of woody plant species. In Arizona, these include mesquite, desert ironwood, blue paloverde, and desert willow (Johnson et al., 1989).
Rocky Mountains
The Rocky Mountains extend from Canada south to Mexico and are characterized by snowmelt-dominated stream flows. The riparian vegetation of the Rocky Mountains, as elsewhere, varies with elevation and latitude. The low valleys once had well-developed forests of narrowleaf cottonwood and strapleaf willow. The riparian association in the montane zone of the Colorado Rockies includes narrowleaf cottonwood, blue spruce, gray alder, water birch, and redosier dogwood (Baker, 1989). This association also occurs in eastern Idaho, western Wyoming, and southern Utah. In southern Colorado and northern New Mexico, the dominant riparian association consists of white fir, blue spruce, narrowleaf cottonwood, Rocky Mountain maple, redosier dogwood, and gray alder. Narrow-leaf cottonwood continues as an important riparian tree into the subalpine region of the central and southern Rocky Mountains. In the northern Rockies of Montana, black cottonwood is the dominant riparian tree. Other riparian trees or shrubs in the Rocky Mountains of regional importance include serviceberry, chokecherry, hackberry, and hawthorn. In the northern Rocky Mountains willows replace trees as the dominant riparian woody species on sites with gravelly silt loams, less than 42 percent coarse fragments, and a groundwater table within 0.6 m of the surface (Law et al., 2000).
Great Plains
The Great Plains, extending from the midsection of the country to the Front Range of the Rocky Mountains, are characterized by a strong moisture gradient from east to west and a steep change in temperature from south to north. Although thunderstorms dominate runoff in most of the region, the amount and timing are highly variable in comparison with winter storms. The riparian, or gallery, forests of the Great Plains show a great structural contrast with the adjacent uplands, which in their pre-agricultural state were dominated by short, mixed, or tall grass prairies. The western Great Plains show reduced diversity in
woody plants. For instance, on the Canadian River of central Oklahoma, the riparian forest was found to be composed of two native species—plains cottonwood and sandbar willow—and one introduced species—saltcedar (Ware and Penfound, 1949). The low diversity within the riparian forest was attributed to frequent flash flooding, shifting sands, high rates of evaporation, and intense heat. Tree diversity within Great Plains riparian areas appears to increase north-ward. Along Plum Creek, Colorado, the riparian vegetation is dominated by plains cottonwood and four willow species (Hupp and Osterkamp, 1996). These sites also support narrowleaf cottonwood and gray alder.
The eastern Great Plains show much greater riparian tree diversity, much like riparian communities farther east. For instance, the riparian forests of eastern Oklahoma support a rich tree flora, including black birch, sweet gum, water tupelo, American sycamore, bald cypress, hackberry, honey locust, elm, hickory, and boxelder (Bruner, 1931). In the northern Great Plains, the diversity of the riparian tree community is also greater than in the western plains. The riparian forests of the Missouri River in central North Dakota are composed predominantly of plains cottonwood, peachleaf willow, chokecherry, green ash, boxelder, American elm, netleaf hackberry, and bur oak (Johnson et al., 1976).
Cool Temperate East
Riparian forests from the western Great Lakes to New England are somewhat less diverse than those farther to the south (Heinselman, 1970; Wells and Thompson, 1974; Morris, 1977; Brinson, 1990). The trees include northern white cedar, black ash, tamarack, balsam fir, and white spruce. Speckled alder grows in forests where sufficient light penetrates that canopy. In riparian areas with poorly drained silty soils of high organic matter content, the dominant trees are slippery maple, eastern and black cottonwood, black cherry, and several species of willow. Well-drained silts generally support ash and American sycamore. On the sandy silt soils of frequently flooded point bars exposed to high flood energy, there is an association of slippery maple, sugar maple, black locust, and American elm. Bitternut hickory grows along with slippery elm and sugar maple on point bars of coarse sandy soils that are flooded with lower frequency. On riparian terraces exposed to the least flooding are forests of red maple, pines, black cherry, and oak.
An extensive listing of wetland tree species by the Minnesota Department of Natural Resources adds several tree species to the riparian flora characteristic of the cool temperate East (Henderson et al., 1998). The wettest areas of Minnesota support buttonbush and narrowleaf willow. In riparian areas that are flooded for shorter periods, tree communities include maple, alder, birch, dogwood, common winterberry, ash, tamarack, black cottonwood, eastern cottonwood, swamp white oak, several willow species, and northern white cedar.
Warm Temperate East
The warm temperate East includes the regions from southern New York and Delaware to Iowa and northern Missouri. In the westernmost part of this region, riparian forests of the Southeast (described below) extend up the Mississippi, Ohio, and Wabash Rivers where they are replaced gradually by those of the warm temperate East (Buell and Wistendahl, 1955; Wistendahl, 1958; Lindsey et al., 1961; Wolfe and Pittillo, 1977; Hughes and Cass, 1997). In the lower portions of floodplains, the historical forest was dominated by cottonwood, black willow, silver maple, ash, and American elm. The structure of these communities, however, was significantly altered with the introduction of Dutch elm disease. Less frequently flooded associations include hackberry, silver maple, sugar maple, redbud, American beech, and American elm. Other important riparian trees in this region include red maple, tulip tree, white ash, American hornbeam, black walnut, spice bush (a shrub), oak, and American sycamore. Along streams with steeper flow gradients and higher water velocities, the woody vegetation is dominated by hazel alder, silky dogwood, American witch hazel, possum haw, and black willow. On the narrow floodplains along streams in the western Appalachian Mountains, riparian forests are dominated by river birch or a more diverse association of bitternut hickory, red maple, tulip tree, black cherry, and black locust (Wolfe and Pittillo, 1977).
Southeast
The southeastern riparian region corresponds broadly to the distribution of bald cypress, though within the region, bald cypress is largely limited to the wettest and most deeply flooded sites (Mattoon, 1915). In these sites, bald cypress often occurs with water tupelo. Moving toward uplands from the wettest sites, bald cypress and water tupelo are replaced by a diverse forest that includes hickory, ash, willow, birch, cottonwood, sweet gum, hackberry, red maple, and oak (Brinson, 1990). Pines and oaks often dominate the forests in the least flooded sections of riparian areas.
Hupp and Osterkamp (1985) analyzed the distribution of woody species across the riparian landscape along Passage Creek, Virginia, dividing the riparian area of this high-gradient stream into depositional bars, channel shelf, floodplain bank, and floodplain. Species found mainly or largely restricted to channel shelves included hazel alder, silky dogwood, American sycamore, common winterberry, slippery elm, and southern arrowwood. Species restricted to the floodplains of the study area included black walnut, bitternut hickory, and American elm. A third group of species, found broadly across riparian areas, included boxelder, American hornbeam, green ash, eastern cottonwood, and American bladdernut. Only one tree species—black willow—occurred on depositional bars. In studying
lower-gradient rivers in Virginia, Hupp and Osterkamp (1996) found that the composition of riparian woody plant communities was similar to those found along the higher-gradient Passage Creek. However, the riparian areas of lower-gradient rivers included additional species—sweetgum, water hickory, silver maple, American beech, tulip tree, bald cypress, and water tupelo. Flood frequency and intensity were hypothesized to be the most important factors influencing the distribution of woody plant species within riparian landscapes.
Summary
The regional variation in riparian tree communities across North America is summarized in Table 2-1, which corresponds with the broad geographic areas in Figure 2-24. The only tree species listed are native trees with a wetland indicator status of obligate species, facultative wetland species, or facultative species for a particular region. One of the patterns discernable from the table is the higher regional diversity—from 26 to 33 genera—in riparian trees from the Great Plains eastward. West of the Great Plains, the number of riparian tree genera within regions ranges from 9 to 22. These results, however, are skewed toward areas where species reach tree stature, and thus do not thoroughly take into account riparian areas in arid climates and high altitudes that are dominated by shrubs and grasses. The purpose of making the comparison is not to determine which geographic areas have the highest species richness, but to show the distribution patterns of dominant genera. Despite the wide regional variation in riparian tree genera represented, a core of genera—alders, cottonwoods, and willows—grows in riparian areas across the continent. A second group of very widely distributed genera includes ash, birch, hackberry, hawthorn, and maple. These primary and secondary core genera are also associated with riparian areas across Europe and Asia. The remaining 37 genera in Table 2-1 are of regional significance and often dominate riparian tree biomass within their characteristic regions.
Table 2-2 attempts to identify the most important tree genera within each region in terms of their relative biomass or frequency of occurrence within riparian forests. The result is a list of five or six genera for eight of the regions and ten genera for the southeastern United States. The southeastern flora are more difficult to compress because riparian vegetation ranges from bald cypress and tupelo in the wettest portions of floodplains to less flood-tolerant taxa in the upper reaches of the same drainage system. In this respect, the Southeast is more diverse than other regions included in the analysis.
Influences of Riparian Vegetation
Riparian vegetation has profound effects on the microclimate of streams and floodplain forests. We have previously discussed how vegetation affects water cycling through evapotranspiration—an inevitable consequence of the growth of
TABLE 2-1 Distribution of Native Riparian Tree Genera Across the United Statesa
|
Primary core genera |
X |
|
|||||||
|
Secondary core genera |
X |
|
|||||||
|
Regionally important genera |
X |
|
|||||||
|
Riparian Tree Genera |
Boreal and Arctic |
Pacific Northwest and Coastal Mountains |
Great Basin |
Arid and Semiarid Southwest |
Rocky Mountains |
Great Plains |
Cool Temperate East |
Warm Temperate East |
Southeast |
|
Alder, Alnus |
X |
X |
X |
X |
X |
X |
X |
X |
X |
|
Ash, Fraxinus |
|
X |
X |
X |
X |
X |
X |
X |
X |
|
Bald cypress, Taxodium |
|
|
|
|
|
X |
|
X |
X |
|
Beech, Fagus |
|
X |
X |
||||||
|
Birch, Betula |
X |
X |
X |
|
X |
X |
X |
X |
X |
|
Bladdernut, Staphylea |
|
X |
X |
X |
X |
||||
|
Buckeye, Aesculus |
|
X |
|
X |
X |
||||
|
Buttonbush, Cephalanthus |
|
X |
|
X |
|
X |
X |
X |
X |
|
Calif laurel, Umbellularia |
|
X |
|
||||||
|
Cedar/arborvitae, Thuja |
X |
X |
|
|
X |
|
X |
X |
|
|
Cherry, Prunus |
|
X |
|
X |
X |
X |
X |
X |
|
|
Coastal redwood, Sequoia |
|
X |
|
||||||
|
Cottonwood, Populusb |
X |
X |
X |
X |
X |
X |
X |
X |
X |
|
Desert ironwood, Olneya |
|
|
|
X |
|
||||
|
Desert palm, Washingtonia |
|
|
|
X |
|
||||
|
Desert willow, Chilopsis |
|
|
|
X |
|
||||
|
Dogwood, Cornus |
X |
X |
X |
X |
X |
X |
X |
X |
X |
|
Elderberry, Sambucus |
|
X |
X |
X |
X |
X |
|
|
|
|
Elm, Ulmus |
|
X |
X |
X |
X |
||||
|
Fir, Abies |
|
X |
|
|
X |
|
X |
|
|
|
Hackberry, Celtis |
|
X |
X |
X |
X |
X |
X |
X |
X |
|
Hawthorn, Crataegus |
|
X |
X |
X |
X |
X |
X |
X |
X |
|
Hemlock, Tsuga |
X |
|
|||||||
TABLE 2-2 Dominant Tree Genera in Riparian Areas Across the United Statesa
|
Riparian Tree Genera |
Boreal and Arctic |
Pacific Northwest and Coastal Mountains |
Great Basin |
Arid and Semiarid Southwest |
Rocky Mountains |
Great Plains |
Cool Temperate East |
Warm Temperate East |
Southeast |
|
Alder, Alnus |
X |
X |
X |
X |
X |
|
|
|
|
|
Ash, Fraxinus |
|
X |
|
|
|
X |
X |
X |
X |
|
Bald cypress, Taxodium |
|
|
|
|
|
|
|
|
X |
|
Birch, Betula |
X |
|
X |
|
X |
|
|
|
|
|
Cottonwood,Populusb |
X |
X |
X |
X |
X |
X |
X |
X |
X |
|
Dogwood, Cornus |
X |
|
|
|
X |
|
|
|
|
|
Elm, Ulmus |
|
|
|
|
|
X |
X |
X |
|
|
Hawthorn, Crataegus |
|
|
X |
|
|
|
|
|
|
|
Hackberry, Celtis |
|
|
|
|
|
X |
|
|
|
|
Hickory, Carya |
|
|
|
|
|
|
|
|
X |
|
Maple, Acer |
|
X |
|
|
X |
X |
X |
X |
X |
|
Mesquite, Prosopis |
|
|
|
X |
|
|
|
|
|
|
Oak, Quercus |
|
|
|
|
|
|
|
|
X |
|
Cedar/arborvitae, Thuja |
X |
|
|
|
|
|
X |
|
|
|
Sweetgum, Liquidambar |
|
|
|
|
|
|
|
|
X |
|
Sycamore, Platanus |
|
X |
|
X |
|
|
|
X |
|
|
Tulip tree, Liriodendron |
|
|
|
|
|
|
|
|
X |
|
Tupelo, Nyssa |
|
|
|
|
|
|
|
|
X |
|
Walnut, Juglans |
|
|
|
X |
|
|
|
|
|
|
Willow, Salix |
X |
X |
X |
X |
X |
X |
X |
X |
X |
|
aData abstracted from the review of Brinson (1990). bAll Populus are considered in this row, not just cottonwood. |
|||||||||
riparian vegetation. As discussed below, there are numerous important ecological and geomorphic benefits of riparian vegetation, such as shading and temperature amelioration, provision of large wood, and stabilization of alluvial sediments that constitute the floodplain.
Thermal Regulation
Temperature in environmental systems can have important and profound effects upon a wide range of physical, chemical, and biological processes. For example, in aquatic systems the solubility of oxygen gas is inversely dependent upon water temperature; hence, higher water temperatures often lead to decreased levels of dissolved oxygen and increased stress for those organisms requiring adequate amounts of instream dissolved oxygen. The rates at which many chemical reactions proceed are also strongly influenced by temperature. As a result, basic processes such as transpiration, respiration, outgoing long-wave radiation, and others are strongly dependent upon environmental temperature regimes.
Vegetation has an important role in influencing the local thermal regime of streams and their adjacent riparian areas. It has been well established that riparian plant canopies—particularly those of forest vegetation—are effective interceptors of incoming solar radiation and can thus greatly reduce the amount of solar energy available to a stream or river (Brown, 1969). The shading effects of riparian plant canopies can prevent or retard the rate of stream warming during clear-sky conditions in summer months when streamflows are often low and high instream temperatures are of concern to aquatic biota (especially cold-water fish) (Gregory et al., 1991; McCullough, 1999; Naiman et al., 2000). For example, the only measured environmental variable that clearly distinguished between trout and non-trout streams in Ontario, Canada, was weekly maximum water temperature, which was inversely related to the percentage of upstream banks covered by forest (Barton et al., 1985). In Bear Creek, Oregon, the partial recovery of riparian vegetation following a change in grazing practices led to the trapping of sediments, increased storage of moisture in riparian soils, and conversion to perennial flow (see Box 5-1). Rainbow trout, a cold-water species that had been eliminated possibly by high temperatures and degraded habitat, returned to the stream and is present year round. On hot summer days in Glacier National Park, Montana, Hauer et al. (2000) observed temperatures as high as 25 °C in exposed channels of an alpine stream segment, whereas the stream did not exceed 10 °C further downstream where the stream was completely canopied by riparian forest. A large number of studies from various portions of the world have shown an increase in stream temperature as a result of decreased riparian vegetation (Figure 2-25).
Riparian vegetation can also influence local wind patterns, conductive and convective heat transfer, outgoing long-wave radiation, and other energy transfer processes in aquatic systems and the land occupied by riparian plants. The result
FIGURE 2-25 Changes in stream temperature in response to differences in riparian shade in empirical field studies throughout the world. Temperature change is expressed as the change in maximum temperature before and after vegetation removal or between reaches with and without vegetation. Locations are U.S. States, British Columbia (BC), New Zealand (NZ), and United Kingdom (UK).
is a local microclimate along streams and adjacent riparian systems that is more thermally moderate than adjacent upland environments. Although the effects of riparian plants upon stream temperatures has been widely studied (e.g., see review by Beschta et al., 1987b), their influences on other microclimate variables (atmospheric humidity, air temperatures, soil temperatures) have not.
In addition to the direct thermal moderation associated with riparian plant communities, there is an indirect, yet important, role that these plants can have on instream temperatures. The root strength associated with riparian plants often allows channels to remain relatively narrow (i.e., a small width-to-depth ratio). If such vegetation is removed and the root strength of the streambank is lost, channels over time may experience widening (i.e., a larger width-to-depth ratio) and become shallower. Given a specific amount of energy absorbed per unit surface area, a wide, shallow channel will experience a greater temperature increase than a narrow, deep channel. Furthermore, channel morphology can play an important role in the connectivity of instream flows with the hyporheic zone and groundwater. For example, alterations to riparian plant communities that contribute to channel incision may sufficiently modify hyporheic and groundwater connectivity such that thermal regimes and low-flow periods are significantly altered. Because streamside vegetation can influence both channel morphology and energy transfers in a variety of ways, alterations to riparian plant communities often have major influences on local microclimates and the thermal regimes of aquatic
systems. These effects occur not only in local reaches, but may also contribute cumulatively to downstream warming of a stream or river (Bach, 1995; Boyd et al., 1998).
In some areas, the hyporheic zone is also important in temperature regulation in streams. Mosley et al. (1983) observed no response of stream temperature to removal of riparian vegetation in a New Zealand stream (see Figure 2-25). The authors concluded that subsurface exchange of water was the major factor determining water temperature in this alluvial system. Schloz (2001) found that reaches of the Wenatchee River in Washington exhibited little response to riparian shading and that hyporheic exchange had a much greater influence on stream temperatures. Such studies and findings are scarce, but they indicate the potential for subsurface exchange, either from hyporheic flow or groundwater, to influence stream temperatures and moderate the influence of riparian shade on stream temperature. Emerging studies of hyporheic processes will provide the empirical basis for modeling the potential influences of alluvial and groundwater exchange for stream temperature.
Because of the presence of water (surface and/or subsurface) and potentially high transpiration rates, riparian areas typically have microclimates (e.g., thermal and relative humidity regimes) that are different than adjacent terrestrial environments. Nonetheless, investigation of the microclimate characteristics of such landscapes prior to the 1990s was limited. In the Pacific Northwest, forested riparian areas during summertime conditions have been found to have lower air temperatures (and less variance in air temperature) and higher daytime humidity (and less variance in humidity) than nearby terrestrial or upland areas (Danehy and Kirpes, 2000). This issue is particularly important to the use of riparian buffer strips alongside forestry operations for maintaining many of the functions of stream and riparian systems. That is, the effectiveness of such buffers with regard to protecting riparian microclimates has not been demonstrated (Brosofske et al., 1997; Dong et al., 1998). In general, little is known about how changes in riparian microclimates that might occur because of adjacent timber harvests may or may not affect riparian plant and animal communities. Perhaps even less is known regarding the microclimates of riparian areas in range and agricultural settings and the extent to which they have been altered by historical management practices. Additional microclimate research is clearly needed for a wide range of conditions.
Large Wood
Forested riparian areas contribute wood to streams, lakes, and wetlands. Several processes can recruit trees from riparian forests into adjacent bodies of water, including individual tree mortality, blowdown, bank erosion, and, in steep mountainous terrain, landslides. Large-wood accumulation in streams and rivers has historically been an important feature of forested riparian systems for all
stream sizes and in all geographical areas of North America (Sedell and Luchessa, 1982; Maser et al., 1988).
Large wood plays a critical role in maintaining and restoring physical habitat, biodiversity, and ecosystem processes in stream and river ecosystems at both local and landscape scales. Large wood creates roughness elements in flowing water, which shape pools and riffles, create variable velocities, and increase the residence time of water (Sedell and Beschta, 1991). Large wood helps streams and rivers slow the downstream routing of sediment and organic matter by providing increased hydraulic resistance to flow and encouraging the local deposition of these materials. Large wood also increases retention of dissolved nutrients and particulate material, and it supports microbial processes and nutrient transformation (Bisson et al., 1987; Maser et al., 1988). Wildlife benefit from the presence of large wood because it can serve as habitat, including refuge during floods and cover, and it provides an abundant but low-nutritional-quality food supply (i.e., it is a potential carbon source to some streams).
The amount of wood found in aquatic and riparian systems is a function of the rate of input, downstream transport, and rate of breakdown from decay and abrasion. As a result, standing stocks of wood differ from small streams to large rivers (Harmon et al., 1986; Maser and Sedell, 1994; Ralph et al., 1994) (Figure 2-26). Amounts of wood per unit area of streambed are greatest in small headwater streams where lateral input rates are high, flows are inadequate to transport
FIGURE 2-26 Standing stocks of wood in streams of different size in the McKenzie River Basin of Oregon. SOURCE: Reprinted, with permission, from Harmon et al. (1986). © 1986 by Harcourt International.

large pieces, and narrow channels resist movement of wood. In these small streams, wood plays a critical role in bed formation, storage of sediment and organic matter, habitat provision, and flood refuge. As streams increase in discharge and width downstream, flows can transport larger pieces, and wider channels provide less resistance to movement. The greater width of these streams also means that lateral inputs are distributed across a larger area, leading to lower standing stocks per unit area. In these larger systems, wood functions laterally along the river margins, and as part of major dams that accumulate during transport events. In floodplain rivers, high flows can move even the largest logs, and the extremely wide channels provide little resistance to movement. Wood outside the channel dissipates the power of overbank flows, traps sediment during flooding, serves as wildlife habitat and refuge, provides nursery logs for certain tree species that typically regenerate on downed wood, and supports microbial processes and transformation of critical nutrients (Maser et al., 1988).
Historically, large wood has often been cleared from streams, rivers, and riparian areas in order to protect bridges and roads from the effects of wood being transported downstream during high flows and to make rivers more navigable. As discussed in Chapter 5, the reintroduction of large wood to streams has become attractive and popular, particularly in the Pacific Northwest, as part of efforts to restore anadromous fish runs. These activities have seldom been evaluated in a systematic manner, however, and their level of success remains largely unknown.
Bank Stabilization
As early as 1885, it was understood that vegetation in riparian areas plays a critical role in stabilizing streambanks and thus supporting aquatic habitats (Van Cleef, 1885). Nearly a century later, Hickin (1984) identified five important mechanisms whereby vegetation influenced rivers in British Columbia: flow resistance, bank strengthening, bar sedimentation, formation of logjams, and concave bank (point bar) deposition. He identified a need “for studies to isolate the influence and quantity and, particularly, quality of vegetation on channel morphology and on lateral migration rates”—a need that continues today. Although the interconnection between soil, hydrology, and vegetation has widely been recognized in the jurisdictional delineation and understanding of wetland functions, an appreciation of how these components are intrinsically linked in riparian areas, particularly with regard to bank stability, has developed much more slowly (Sedell and Beschta, 1991).
For most streams and rivers, native plant communities are generally effective at protecting banks from the erosive effects of moving water. Aboveground plant parts (stems of trees, shrubs, graminoids, and forbs) tend to dampen turbulence and slow velocities at the water–streambank interface (Klingeman and Bradley, 1976). Root systems and soil organic matter help bind soil particles and alluvial sediments (Rutherford et al., 1999). Thus, streambanks can remain relatively stable even though they may experience considerable shear stress during periods of high flow. Although the shear stress imparted from flowing water is highly variable across a given channel cross section, such forces tend to be relatively high along the outside banks of meandering channels (Bathurst, 1979).
Different riparian plant communities may influence bank stabilization in different ways. For example, Trimble (1997) found that forested banks of some streams are more prone to periodic bank erosion compared with banks vegetated by herbaceous species, particularly grasses and sedges. Vegetated streambanks of any kind are typically more stable than unvegetated ones (Hicken, 1984).
Not only does vegetation provide stability to banks directly via its root system, but its ability to dissipate stream energy along a reach is also an extremely important function. As discussed previously, riparian vegetation plays a major role in creating relatively high roughness values for streambanks, thus retarding the potential for a bank to erode or the channel to migrate laterally. The establishment, growth, and succession of individual plants and groups of plants represent a mechanism by which riparian plant communities can continually respond to natural changes in streamflow and sediment loads and to local adjustments in channel morphology (Kalliola and Puhakka, 1988; Kalliola et al., 1991).
In addition to the immediate effects of resisting fluvial erosion of streambanks, vegetation plays a similarly important role with regard to floodplain development. Floodplain vegetation (including canopies, branches, stems, roots, and litter) not only protects the soil from direct rainfall impact and reduces the veloc-
ity of overbank flows (thereby preventing scour), but it also promotes deposition of fine sediments. In years without floods, riparian vegetation becomes established on exposed areas of floodplains, providing stability to these areas and promoting the vertical accretion of sediment during subsequent floods (Friedman et al., 1996a,b). Graminoids and other low-growing plants are particularly effective at protecting soils from surface erosion at high flows and are important for causing deposition of silts, although the amount of roughness associated with graminoids depends on species, stage of growth, and degree of disturbance (Wilson, 1967). Thus, riparian vegetation has had a fundamental role in the long-term maintenance of streambanks and floodplain landforms.
Other Effects of Vegetation
Other effects of vegetation on water cycling include the effects of root growth and decay in creating soil macroaggregates and consequently macropores, which increases soil permeability. Trees also modify the delivery of precipitation to the ground by intercepting, storing, and evaporating a portion of the incoming precipitation. In other instances, precipitation that is temporarily detained in the forest canopy but subsequently routed to the ground may be spatially redistributed by tree leaves, branches, and trunks. Root wads along the banks of streams reduce the rate of erosion, but also create channel complexity and instream habitat through overhangs and shading.
RIPARIAN AREAS AS HABITAT
The role of riparian areas in maintaining biodiversity is well known; their relative contributions greatly exceed the proportion of the landscape they occupy (Naiman et al., 1993, 2000; Crow et al., 2000). Scientific documentation of the importance of these areas for plants and animals comes from studies across the continent. In the Pacific Coast ecoregion, 60 percent of amphibian species, 16 percent of reptiles, 34 percent of birds, and 12 percent of mammals can be classified as riparian obligates (Kelsey and West, 1998). In the arid Southwest, 70 percent of threatened and endangered vertebrate species are listed as riparian obligates (Johnson, 1989); 60 percent of all vertebrate species are so defined (Ohmart and Anderson, 1982). Even in the relatively mesic environment of Wisconsin, 80 percent of plant and animal species on the state’s endangered list live all or part of their lives in the riparian area of lakes (Korth and Cunningham, 1999).
Habitat, which refers to the place an organism lives, comprises both abiotic and biotic factors (Odum, 1971) including the basic provisions of food and shelter. Clearly, many animals require more than one habitat type throughout their life cycle. In popular accounts and wildlife management literature, habitat commonly refers to places providing reproductive requirements. In the case of migra-
tory animal species, habitat also is used to refer to requirements of the animal in the non-breeding season, including stopover and wintering sites. Some animals use riparian areas as part of their home ranges or territories, moving through them on a short-term basis and over periods of hours and days. Indeed, the short-term movements of small mammals and birds within riparian areas have been shown to aid in the cycling of nutrients between the aquatic environment and adjacent uplands (Dobrowolski et al., 1993).
Riparian areas provide food resources for animals throughout the food web. Riparian vegetation along streams and lakes is critical as a primary food source to invertebrates from all the guilds (filter feeders, shredders, scrapers, and predators). It also provides a landing substrate for adult insects (such as midges, stoneflies, and mayflies) emerging from a waterbody (Benke and Wallace, 1990). As discussed below, the importance of this flux of insects in riparian areas to migratory and resident birds has received increased attention from ornithologists. The structural diversity of plant species in riparian areas creates a wide variety of feeding niches for herbivores and carnivores alike.
Various types of dispersal occur in riparian areas, including immigration, emigration, and migration. When applied to animals, migration connotes a cyclical movement, such as the spring and autumn migrations of North American songbirds. Applied to plants, migration refers to contraction or expansion of a population through time (Sauer, 1988). Hunter (1996) speaks of four basic types of movement: (1) daily movements of animals among patches of preferred habitat within their home range, (2) annual migrations, (3) dispersal movements of young animals and plant propagules, and (4) shifts in range in response to climate changes.
Although the following examples are limited to vertebrate animals and plants, it is well known that riparian areas also provide valuable habitat for invertebrates. The thermal regulation of streams and the supply of large wood afforded by riparian vegetation lead to characteristic invertebrate species both within streams and in their associated riparian areas (Hawkins et al., 1982; Anderson and Wallace, 1984; Benke and Wallace, 1990; Ward et al., 1999).
Fish
Much of the concern about riparian areas, particularly in the Pacific Northwest, had its genesis in maintaining viable fish populations (Hall and Lantz, 1969; Budd et al., 1987; Hall et al., 1987; Hartman et al., 1987). The major fish habitat elements that are influenced by riparian areas are water temperature, food supply, large wood, channel structure, and sediment (Wilzbach, 1985; Budd et al., 1987; Gillilan and Brown, 1997; Bisson et al., 2000). Temperature changes caused by the presence or absence of riparian vegetation have been shown to account for variability in trout populations (Barton et al., 1985; Wesche et al., 1987). Subsurface exchange between groundwater underneath riparian areas and
streams also plays a role in creating critical cold-water fish habitats (Stanford and Ward, 1993).
At least three decades of studies have emphasized the connection between aquatic nutrient input and processing and riparian areas. Studies have shown that in forested regions over 99 percent of energy and organic carbon in food webs of small headwater streams originate in adjacent forest ecosystems (Fisher and Likens, 1973). These terrestrial sources of food influence the growth and abundance of fish communities (Chapman, 1965, 1966; Mundie, 1969). Riparian vegetation also influences light availability, which affects the efficiency of prey capture by stream fish (Wilzbach and Hall, 1985). As forested streams increase in size from headwaters to large rivers, instream primary production contributes more to the food base of stream ecosystems, although floodplains and their forests still deliver large portions of the energy supply to large lowland rivers (Vannote et al., 1980; Minshall, 1988; Junk et al., 1989). Riparian forests with multiple canopy layers, species, and age classes offer a wider array of food resources and physical habitats than do simple uniform plant communities (Gregory et al., 1991; Swanson et al., 1992; Osborne and Kovacic, 1993; Moyle and Yoshiama, 1994). In particular, the diversity and density of aquatic macroinvertebrates—a primary fish food resource—are higher in streams with wider riparian areas (Newbold et al., 1980). Similar observations have been made in lakeshore fish communities (Bryan and Scarnecchia, 1992; Christiansen et al., 1996).
Riparian areas and floodplains are major habitat elements for fish assemblages in streams and rivers (Schlosser, 1991) and have a pronounced influence on fish movement, particularly on migration of anadromous fishes (Gillilan and Brown, 1997). River networks and their floodplains create distinctive landforms that lead to biological diversity among fish species. Tributary junctions and complex channels are some of the most diverse areas within the river network because of their array of depths, velocities, and channel edges (Gorman and Karr, 1978; Bayley and Li, 1992); they also support different fish species than those found in the main channel and adjoining tributary system. Floodplains, side channels, and alcoves can serve as lower-velocity refuges during floods and provide access to abundant food resources for fish communities during flooding (Schlosser, 1982; Angermeier, 1987; Bayley and Li, 1992).
Given the many roles that riparian areas play in supporting fish communities, it is not surprising that the abundance and distributions of fish populations have been shown to be affected by land-use practices and riparian forest conditions (Hall and Lantz, 1969; Murphy et al., 1981; Bisson et al., 1992; Frissell, 1993; Gregory and Bisson, 1996; Naiman et al., 2000). Habitat degradation and impaired riparian conditions have been associated with 90 percent of the observed extinctions and declines in anadromous salmonids in the Pacific Northwest (Nehlsen et al., 1991). Thus, it is clear that healthy and functional riparian areas are essential for the abundance and diversity of fish (NRC, 1996).
Herptiles
Most amphibian species and many reptiles are intimately associated with riparian areas and their waterbodies. A recent summary of herptile (reptile and amphibian) species in the eastern United States lists 121 species found in riparian areas of springs, streams, rivers, lakes, ponds, bogs, and ephemeral pools (Pauley et al., 2000). Because of this close association, herptiles have been used, in some cases, as indicators of riparian condition (Lowe, 1989; Wake, 1991).
Amphibians. Few terrestrial vertebrates are as closely tied to the presence of water as amphibians, which require water bodies for completion of their life cycles. Those amphibians living entirely within small streams, such as salamander species of the genera Desmognathus and Plethodon in the Southeast and the genera Dicamptodon and Rhyacotriton in the Pacific Northwest, depend (as do fishes) on riparian functions that perpetuate healthy aquatic ecosystems. Frogs, toads, and salamanders are closely associated with riparian areas year-round throughout the continent, and intact riparian areas between upland habitat and aquatic breeding habitat are crucial to their viability. Adults must effectively reach breeding areas in their seasonal migration, and emerging young need to disperse from the breeding waters into surrounding uplands. Analysis of data from five eastern states found that a buffer extending at least 164 meters from the water’s edge was required to perpetuate salamander populations (Semlitsch, 1998). For managed riparian areas to be effective in conserving pond-breeding species, they need to include a longitudinal component as well as sufficient width (Dodd and Cade, 1998). Microhabitat attributes of riparian areas are also important for meeting habitat requirements of herptiles. Work in eastern Texas revealed a positive relationship between a closed canopy and leaf litter ground cover and amphibian and reptile abundance in narrow streamside zones (Rudolph and Dickson, 1990). Degradation of riparian areas has led to the decline of some herptile species. For example, the tarahumara frog, which is restricted to southern Arizona streams with an intact community of sycamores and willows, is currently in danger of extinction (Ohmart and Anderson, 1978).
Reptiles. Most turtle species require functioning riparian areas to complete their life cycles. The wood turtle of the northeastern United States uses a variety of riparian habitats lying within about 360 meters of its home river for foraging, basking, and nesting (Vogt, 1981; Ewert, 1985). Other species of turtles, such as the snapping turtle, are almost exclusively aquatic but need the riparian area for nesting (Harding, 1997). Sea turtles of various families and species are completely dependent on coastal riparian dunes and beaches for nesting.
Snake species provide a less clear-cut example of obligatory dependence on riparian areas, although they are frequently seen hunting these biologically rich ecotones or seeking the cooler microclimate associated with them. In the Mid-

west, the queen snake is a riparian obligate that finds its major prey within riparian areas of streams, ponds, lakes, marshes, ditches, and canals (Harding, 1997). In the West, the western hognose snake is a denizen of river floodplains and even mountain canyons where there are alluvial deposits suitable for burrowing (Stebbins, 1966). Several species of lizards are strongly associated with the vegetative cover and organic material of southwestern riparian areas (Jones and Glinski, 1985; Warren and Schwalbe, 1985).
Birds
The importance of riparian areas as breeding habitat for birds is well known to birdwatchers and professional ornithologists alike. Because riparian areas are inherently diverse in plant species and varied in vertical and horizontal structure, they provide a variety of niches for birds, as documented by numerous studies (MacArthur, 1964; James, 1971; Karr and Roth, 1971; Whitmore, 1975; Rice et al., 1983, 1984). Riparian areas also provide a ready source of insects for breeding and migrating birds. Indeed, because of their close association with vegetation, and the relative ease of counting them, birds are often studied in assessments of riparian community functioning.
Perhaps nowhere is the relative importance of riparian areas vs. uplands to bird species as dramatic as in the arid southwestern United States. Avian density,

species richness, biodiversity, number of rare species, number of breeding pairs of birds, and biomass are extremely high in riparian areas compared to uplands (Szaro, 1991; Ohmart, 1996). These patterns are observed, sometimes to a lesser extent, across the country. In Iowa, wooded riparian areas provide habitat for 32 bird species as contrasted with only eight species in herbaceous areas (Stauffer and Best, 1980). Intact riparian areas of 75–175 m included 90 percent to 95 percent of Vermont’s bird species (Spackman and Hughes, 1995). Even in mesic forests of Canada, boreal riparian conifer stands have higher avian diversity and abundance than do the adjacent coniferous uplands (Larue et al., 1995). In some landscapes lacking contrast between riparian and upland vegetation, such as in parts of the Pacific Northwest, avian diversity in riparian areas has not been found to be significantly greater than in uplands, although some differences in community composition have been documented (McGarigal and McComb, 1992; Murray and Stauffer, 1995).
Migrating birds use riparian areas as navigational aids and for stopover sites (Faaborg, 1988; Gill, 1990; Helmers, 1992). Indeed, their suitability as stopover sites in terms of food availability, safety from predators, and vulnerability to environmental stresses is receiving increasing attention by researchers concerned about population declines (Moore and Simons, 1992; Ewert and Hamas, 1996). Midcontinent banding studies along the North Platte River have demonstrated the role of large-scale riparian areas in the migration of songbirds (Brown et al., 1996; Scharf and Kren, 1997). Even very tiny riparian areas can be crucial to
birds, as was illustrated in Wisconsin by the observation of 20 migratory avian species around vernal pools in a landscape that was otherwise virtually devoid of birds (Premo et al., 1992, and similar studies such as Long and Long, 1992).
Riparian areas have value not only for migratory birds, but also as habitat for juvenile birds in the post-fledging, premigration period (Faaborg et al., 1996) and for the dispersal of juveniles at the end of the breeding season (Machtans et al., 1996). As illustrated in Box 2-2, riparian areas in the sagebrush steppe have been found to be critically important to sage grouse species as brood-rearing habitat. Work along the lower Colorado River valley has highlighted the importance of riparian vegetative structure to wintering avian species, an often-overlooked habitat component (Anderson and Ohmart, 1977). Based on our knowledge of bird migration, riparian areas are critical to the conservation of avian diversity at the scale of regions, continents, and even hemispheres.
Mammals
Mammalian species with semi-aquatic habits, such as water shrew, star-nosed mole, beaver, river otter, and mink, clearly incorporate riparian areas as part of their habitats, finding in such areas critical food and shelter resources (e.g., see DeGraaf and Yamasaki, 2000). Examples of mammalian presence in riparian areas abound in the literature. The greatest activity for several species in Michigan, including white-tailed deer, bobcat, red squirrel, and snowshoe hare, occurred within 400 feet of a third-order stream (Rogers et al., 1992). Similarly, in Vermont “most movement” of white-tailed deer, coyote, raccoon, red fox, gray fox, snowshoe hare, and voles occurs within a few meters of the high-water mark (Spackman and Hughes, 1995). In the Southwest, over half the small mammals trapped in a larger desert study were from a desert wash (Szaro, 1991). In such arid areas, species like the cotton rat and harvest mouse are perhaps not so much attracted to the water itself as to the denser vegetation found where water is more abundant (Ohmart and Anderson, 1978).
Mammals also use riparian areas at all scales for movement, either as part of their home ranges on a short-term movement basis, or during dispersal. Studies of culvert use by small and medium-sized mammals underlie the importance of even small drainages as crossing points along highways (Clevenger and Waltho, 2000). Conservation biologists have long argued for protecting riparian areas as a mechanism for conserving far-ranging large mammals in landscapes with fragmented habitats, as they provide dispersal avenues and means for gene flow (Beier, 1993; Noss et al., 1996). Indeed, Florida continues to work on designing a linked reserve system for species such as Florida panther and Florida black bear (Hoctor et al., 2000).
Although many mammals depend on riparian areas to survive and thrive, perhaps no North American mammal is as influential in riparian areas as beaver, described in Box 2-3.
|
BOX 2-2 At the time of European settlement, sage grouse were abundant through most of the west. Since settlement, distribution of greater sage grouse (Centrocercus urophasianus) has decreased by about 50 percent, with abundance declining by 45 percent to 80 percent (Braun, 1998). In July 2000, Gunnison’s sage grouse (C. minimus), currently represented by only a few remnant populations in Colorado and southeastern Utah, was recognized as a distinct species (Young et al., 2000) and promptly became a candidate for federal listing as threatened. The U.S. Fish and Wildlife Service determined that listing as threatened is warranted but is precluded by other, higher administrative priorities. Similarly, a distinct population of greater sage grouse in the state of Washington recently was identified as a candidate for threatened status. There is some debate regarding the need to federally list the greater sage grouse throughout its range (Tweit, 2000). Concerns about precipitous population declines have resulted in considerable research on habitat requirements of both sage grouse species at all phases of their life cycles. Both greater and Gunnison’s sage grouse are critically dependent on riparian areas and wet sites within the sagebrush steppe for brood-rearing habitat. Within hours of hatch, females take their young broods from the sagebrush-dominated stands of their nesting sites to more mesic habitat, such as riparian areas. During their first two to four weeks, chicks forage for insects in riparian areas (Klebenow and Gray, 1968; Johnson and Boyce, 1990). Broods often use hay meadows in major drainages and frequently use the interface of two habitat types, such as sagebrush/wet meadow or willow–alder/wet meadow (Young, 1994). In addition to wet meadows, females with broods also use smaller mesic riparian areas around natural seeps and springs. Forbs that occur commonly in these areas constitute a significant portion of the diet of chicks between the ages of 2 and at least 12 weeks (Klebenow and Gray, 1968; Peterson, 1970; Johnson and Boyce, 1990). The importance of riparian areas as brood-rearing habitat appears to be further accentuated in drought years, when these moist areas may provide the only source of succulent forbs in the landscape (J. A. Crawford, Oregon State University, personal communication, 2001). Sufficient escape cover in the form of live, taller sagebrush that grows in riparian areas is important in reducing the depredation of sage grouse that use these sites (C. E. Braun, retired from Colorado Division of Wildlife, now Grouse, Inc., personal communication, 2001). Successful management for sage grouse will undoubtedly require a landscape-scale approach that includes perpetuation of ecologically functioning riparian areas.  |
|
BOX 2-3 Few would argue with the characterization of beaver (Castor canadensis), with its profound influence on riparian lands and aquatic ecosystems, as the quintessential riparian mammal. Its historic impact on North American economies and exploration eclipses that of most other wildlife. In addition, numerous scientific studies illustrate the critical role beaver play in riparian areas of North America at a wide range of temporal and spatial scales. By all available estimates, this influence was even greater in the past. Historical data from trapping harvests put the continental population of beaver before European settlement as high as 60–400 × 106 individuals (Seton, 1929), with a geographic range extending from the deserts of northern Mexico to the arctic tundra (Burt and Grossenheider, 1976; Jenkins and Busher, 1979). The explorer David Thompson, after crossing North America in 1784, stated that the continent “may be said to have been in the possession of two distinct races of beings, man and the beaver—with man occupying the highlands and the beaver in solid possession of the lowlands” (Rezendes and Roy, 1996). The present-day population has probably been reduced by a factor of ten-fold or more (Naiman et al., 1986), even after rebounding from extirpation in many portions of the species’ range (Bergerud and Miller, 1977). Beavers modify riparian areas and aquatic ecosystems by feeding and dam-building activities. These modifications create habitat for a variety of plant and animal species (Cook, 1940; Kirby, 1975; Hair et al., 1978; Malanson, 1993). For example, newly flooded forest ponds allow the formation of heron rookeries. The impounded nutrients result in a particularly rich aquatic system that lasts well beyond the life of the dam. In Oregon, the growth rate and size of juvenile salmonids in beaver ponds has been found to be superior to those of their same age cohorts in stream channel locations (Leidholt-Bruner et al., 1992). The influence of beaver on landscape patch dynamics is profound and well documented (Slough and Sadleir, 1977; Naiman et al., 1986; Remillard et al., 1987), causing more than one researcher to assign beaver the role of keystone species. The impounding of water (primarily on first- through fourth-order streams) has multiple effects, including modifications of channel geomorphology and hydrology, changes in riparian wetland types and vegetation, alterations in the aquatic invertebrate community, modifications in nutrient cycling, and changes in water chemistry (McDowell and Naiman, 1986; Naiman et al., 1986; Ford and Naiman, 1988). The positive effects of beavers on headwater streams (e.g., retention of runoff, groundwater recharge, and sediment trapping) have been particularly appreciated in the western states where beaver have been reestablished by natural resources managers as an integral component of watershed restoration efforts (Federal Interagency Working Group, 1998). Studies in the Southwest suggest that beaver may be an especially important factor in resisting erosional perturbations in first-order streams stemming from overgrazing at the turn of the twentieth century and that they should thus be used as a restoration management tool (Parker et al., 1987). Beavers affect the successional changes and species composition in the riparian vegetative community through differential feeding on woody plant species |
|
(Barnes and Dibble, 1988; Johnston and Naiman, 1990; Nolet et al., 1994). This influence is limited to the area in proximity to their aquatic homes since beavers generally do not forage for food much beyond 50–100 meters from their aquatic safety zone (Bradt, 1938; Jenkins, 1980; Belovsky, 1984). Their foraging distance from water has been shown to increase greatly, however, when large predators such as wolves are removed from the system, shrinking again when large predators return (R. Naiman, personal communication, University of Washington, 2000). In the Great Lakes region, early research found that a typical interval between abandonment and reoccupation of beaver ponds ranged from a few years to about 20 years (Lawrence, 1952). Other more recent work recognizes ontogenies of beaver ponds that can range from a year to many centuries (Naiman et al., 1988).  As beaver populations have rebounded in some portions of the country, conflicts with humans have often ensued. In spite of the demonstrated vital ecological roles played by beaver, the species’ ability to modify the environment frequently runs afoul of human goals. Most obviously and dramatically, beaver cut down vegetation that is desirable to people for its aesthetic or economic value. In addition, beaver activities flood lands that are economically valuable to people, such as forested or arable land. Road culverts provide perfect settings for beaver dams, resulting in flooded roads and added costs in maintenance and repair. Perhaps no conflict, however, matches the fervor of that between fish managers and beaver. In the Great Lakes states, beaver are often targeted for their deleterious impacts on populations of native trout, allegedly arising from effects of their dams on fish passage, streamflow, and water temperature. Yet few quantitative data are available to support this premise. A field study in Wisconsin (Avery, 1992), quantifying the effects of the removal of 546 beaver dams via “before and after” studies on a 9.8-mile branch of a major trout stream (Pemebonwon River) and on 22.7 miles of 14 smaller tributaries, had inconclusive findings that were |
|
statistically untested. For example, the effects of beaver dam removal on aquatic invertebrates were equivocal, with no detectable change as measured by the Hilsenhoff Biotic Index. Removal of dams appeared to have a cooling effect that was more noticeable on the tributaries, although no statistical testing was done. Finally, the study provided no direct evidence that dams acted as a physical barrier to trout movement in the river, although there appeared to be a trend (untested) toward increased trout numbers in the tributaries after dam removal. A more statistical study on the effects of beaver dams on streams in the Pestigo River watershed of northeastern Wisconsin, which examined effects of dam removal on downstream water temperature, found no significant simple reductions in temperature (McRae and Edwards, 1994). Instead, it found that large beaver impoundments may actually have a beneficial effect of dampening daily fluctuations in water temperatures downstream. The researchers concluded that local differences in the degree of vegetative and streambank shading, groundwater inflow, and stream volume make it difficult to generalize about the effects of beaver dams. They advocated that any beaver dam removal be selective and take such factors into account. Human land uses throughout a watershed also have a measurable influence on stream health (as measured by the Index of Biotic Integrity) (Wang et al., 1997), suggesting the need for larger-scale and more inclusive approaches to fish management. Overarching all beaver–human conflicts is the fact that most human activities and goals occur in a spatial-temporal framework that is out of sync with the patch dynamics of a beaver-influenced watershed. Beaver co-evolved with salmonids, making improbable an essentially negative relationship between these organisms. However, long-term population cycles of beaver and their predators, as well as ongoing beaver-induced successional changes in riparian vegetation and stream geomorphology, are not easily incorporated into short-term resource management plans. Even our longest-term baseline data are quite recent (within approximately the last 50 years) and come exclusively from a time after presettlement beaver populations were decimated and the larger landscapes were already greatly altered by agriculture and extensive logging. There certainly are some streams or reaches where beaver activity has been concentrated for an extended period of time by land management practices and thus has become a problem with regard to fish populations. And there are undoubtedly marginal salmonid streams where beaver activities, combined with factors such as riparian vegetative modification or siltation, may impair salmonid habitat. On some streams, fish passage may be temporarily prevented by beaver dams, and fish populations in a particular reach may decline in the short term. But in the long term, beavers eventually will deplete food resources in the area, their dams will degrade and be breached, and young salmonids will access the high-productivity areas (with respect to invertebrates, plants, and nutrients) upstream of dams as part of an ongoing cycle. As was pointed out over a half century ago by fisheries biologist Cook (1940), negative effects on salmonids are influenced by many specific circumstances (such as over-fishing, sedimentation, and channelization) associated with a particular stream, although beavers are often assigned the blame. A new management paradigm is needed that seeks to understand the beaver’s influence in terms of spatial-temporal patch dynamics and that recognizes the key and valuable ecological role played by this obligate riparian mammal (Naiman and Rogers, 1997). |
Plants
In addition to being characterized by unique assemblages of plant species (as discussed earlier), riparian areas frequently harbor rare plant species. Waterbodies and their riparian areas have microclimates and disturbance regimes that allow species to persist at the edges of their ranges. Furthermore, movement and regeneration of plant species are facilitated by floodplain disturbances, further enhancing the potential for perpetuation of plant diversity in riparian areas (Spackman and Hughes, 1995).
Riparian areas provide refugia for populations that may be relicts of former climatic periods. Prairie plant communities recorded along rivers in Miami County, Ohio, since the time of western settlement are illustrative of this phenomenon (Huston, 1978). Work in Europe has shown how landscape elements such as ditch edges are critical to the persistence of endangered plant species even in a highly modified landscape (Ruthsatz and Haber, 1981). Non-vascular plants such as bryophytes are disproportionately found in moist sites such as riparian areas (Andrus, 1990). In an extreme example of rarity associated with riparian areas, the Virginia round-leaf birch (Betula uber) is endemic to two creeks in Smythe County in southwestern Virginia (Ogle and Mazzeo, 1976). Rare and relic plant species found only along the Great Lakes shoreline, in addition to the many animal species using riparian areas in this region, are described in Box 2-4.
ENVIRONMENTAL SERVICES OF RIPARIAN AREAS
The biosphere is often called the human “life support” system because of its paramount importance to maintaining the atmosphere, oceans, and land resources that support human societies. Riparian areas are a subset of this life-support system. On a global scale, they are a major component of the conduits that transport water from the continents to the ocean (Schlesinger and Melack, 1981). They are highly valued by society because they provide sites for human settlement near ports, proximity to water supplies, convenient sites for waste disposal, and opportunities for water-based recreation.
Riparian areas associated with ephemeral channels and small intermittent streams contribute to society in more subtle ways, and they are often disregarded—in part because of the ease with which they can be converted to alternative uses. However, riparian areas of small streams warrant attention not only for their local contributions, but also for their great collective length (see Figure 2-15).
Because the fundamental ecological processes that riparian areas perform occur whether or not humans are present to take advantage of them, they can be discussed somewhat independently of human values that change over time and differ among cultures. Functions fall into three major categories: (1) hydrology and sediment dynamics, (2) biogeochemistry and nutrient cycling, and (3) habitat
|
BOX 2-4 The extensive Great Lakes riparian shoreline is a dominant feature on a continental scale. The state of Michigan alone has a coastal shoreline encompassing 3,222 miles—second only to Alaska. Islands contribute a striking proportion of coastal riparian area, as exemplified by the 948 miles of the Isle Royale archipelago shoreline, which is noted for its unusual assemblage of disjunct plant species (Voss, 1972). Small islands, in fact, may be nearly 100 percent riparian in composition, according to this report’s definition. In addition, the Great Lakes region is well known for its glaciated landscape punctuated with numerous lakes and streams of all sizes. In effect, large portions of this region’s landscape can be thought of as being almost entirely riparian. It is no wonder, then, that riparian areas play a central role in this region’s biodiversity. Great Lakes coastal riparian areas provide habitat for a variety of endemic plant species. Houghton’s goldenrod (Solidago houghtonii), first collected by Douglass Houghton in 1839, occurs nowhere in the world except along the northern shores of Lakes Michigan and Huron (Voss, 1996). Dwarf lake iris (Iris lacustris), another northern Great Lakes endemic, is particularly abundant on the rubble of old glacial beach ridges (Voss, 1972). Lake Huron tansy (Tanacetum huronense), first described by Thomas Nuttall in 1810 at the Straits of Mackinac, inhabits Great Lakes riparian dunes and upper beaches, tracking the fluctuations in lake levels (Voss, 1996). In the Apostle Islands National Lakeshore (Wisconsin), botanists have recorded 809 plant species. A European rush (Juncus squarrosus), known previously only from Greenland, a Eurasian sedge new to the Great Lakes (Carex ovalis), and an eastern North American sedge (C. tincta) are listed among its disjuncts and rarities (Judziewicz and Koch, 1993). In the upper peninsula of Michigan, major river systems such as the Menominee and Escanaba provide refugia for prairie plant species believed to be relicts from the widespread savannas of the warm, dry hypsithermal period (D. Henson, Independent Consulting Biologist, Tamarack Studios, personal communication, 2000) occurring roughly 6,000 years ago (Pielou, 1991; Davis et al., 2000). In general, relict and disjunct populations and endemic species represent pockets of genetic variability (Utter and Hurst, 1990) that may become even more important during periods of future climate change. Pollen analysis from coastal riparian areas of the Great Lakes (national parks and national lakeshores) has demonstrated a climate-modifying capacity of large water bodies that apparently has translated into refugia for plant species during past climate changes and would likely do the same for future changes predicted as a result of global warming (Davis et al., 2000). Great Lakes riparian areas are likewise important to the region’s animals. Work along the Lake Huron shoreline has emphasized the importance of Great Lakes shorelines as stopover points for early-spring migrant birds; birds are substantially more abundant and diverse at coastal sites than they are inland. The main attraction appears to be the rich source of food in the form of emerging aquatic insects such as midges and mayflies that light on the riparian vegetation (Dallman and Smith, 1995; Ewert and Hamas, 1996; Seefelt, 1997; Hyde, 1998; Smith et al., 1998). Riparian areas of rivers and tributaries flowing into the Great Lakes have |
|
enhanced diversity and habitat values resulting from their connectivity to these larger waterbodies. For example, small intermittent and first-order streams provide critical links between major river systems and numerous lakes and rivers on the landscape. Species as diverse as gray wolf (Canis lupus), coyote (Canis latrans), black bear (Ursus americanus), river otter (Lutra canadensis), red fox (Vulpes vulpes), and muskrat (Ondatra zibethicus) have been seen using seasonal drainages that connect with a third-order stream that is a major tributary of the Menominee River, the largest catchment in Michigan’s upper peninsula (E. Rogers and D. J. Tiller, White Water Associates, Inc., personal communication, 2000). Riparian areas of third-order streams flowing into Lake Huron likewise are important to foraging neotropical migrants as stopover sites (Wilson, 2000). |
and food web maintenance. Nine common examples are presented in Table 2-3, and many more could be added or derived from those listed. Present knowledge about these functions derives from an emerging but relatively large body of research conducted in the United States and elsewhere. In many cases, knowledge is sufficiently well developed such that indicators can be used as shortcuts to judge whether the functions are occurring at appropriate levels (see Chapter 5 for an in-depth discussion of assessment methods).
Riparian functions have both on-site and off-site effects, some of which may be expressed as goods and services available to society (Table 2-3). (This approach has been used to interpret the societal values of the functions of other ecosystem types [Christensen et al., 1996]). For example, functions related to hydrology and sediment dynamics include storage of surface water and sediment, which reduces damage from floodwaters downstream from the riparian area. Similarly, the function of cycling and accumulating chemical constituents has been measured in a number of studies on nitrogen and phosphorus cycling. These studies have shown that nutrients are intercepted, to varying degrees, as runoff passes through managed and natural riparian zones. The societal benefit is the buffering effect of pollutant removal, a service that has been a major motivation for protecting and managing riparian areas. Functions related to habitat and food web maintenance are the basis for many valued fisheries, not to mention that they contribute to human activities such as bird watching and wildlife enjoyment.
The hydrologic, nutrient cycling, and habitat/food web functions of riparian areas correspond to goods and services such as support of biodiversity, flood peak reduction, and removal of pollutants from runoff. Except for support of biodiversity, some of the environmental services of riparian areas can be provided by technologies, such as reservoirs for flood peak reduction and wastewater treatment plants for pollutant removal. However, these substitutions are directed at single functions rather than the multiple functions that riparian areas carry out simultaneously and with little direct costs to society.
CONCLUSIONS
Much of our understanding of riparian ecology (as reflected in this chapter) is based upon studies of streams and small shallow rivers because they are easier to study and are prevalent features of all landscapes. Nonetheless, large alluvial rivers have expansive riparian areas that are important as habitat for an enormous array of biota including endangered species, as natural sites for nutrient retention and pollution detoxification, and for other functions and values. Because large river floodplain–riparian areas have been substantially altered by human activities (see Chapter 3), their restoration will require a firm understanding of riparian structure and function at much larger watershed scales.
In a similar vein, our current understanding of the relationships between riparian vegetation and local environmental conditions is quite good. Even though the fundamental principles are universal, transferring specific information on riparian structure and function from one region to another can be problematic because of differences in vegetative composition and stature, discharge regimes, climate, geology and soils, and river network structure. Comparisons at the scale of climatic and geographic zones are seldom attempted even though such information is critical in making nationwide management decisions about riparian resources. Thus, in addition to the specific research recommendations noted throughout this chapter, it will be important to extend our knowledge of riparian functioning into large river systems and to those physical factors that strongly influence riparian systems (e.g., climate, geomorphology, and runoff) and the responses of biological components.
Riparian areas perform important hydrologic, geomorphic, and biological functions. These areas encompass complex above- and below-ground habitats created by the convergence of biophysical processes in the transition zone between aquatic and terrestrial ecosystems. Riparian areas encompass interactions in all three spatial dimensions of drainage corridors: laterally between stream channel and adjacent terrestrial zone, vertically between the surface and subsurface, and longitudinally between upstream and downstream reaches.
Riparian areas cannot be thought of in isolation from stream channels. The characteristic geomorphology, plant communities, and associated aquatic and wildlife species of riparian systems are intrinsically linked to the role of water as both an agent of disturbance and a critical requirement of biota. Although it is widely recognized that the aquatic portion of a riverine or lake system can have a pronounced effect on associated riparian areas, these riparian areas can have important influences on their aquatic systems.
Natural riparian systems have adapted to specific hydrologic disturbance regimes (i.e., flow frequency, magnitude, duration, timing and rate of
TABLE 2-3 Functions of Riparian Areas and Their Relationship to Environmental Servicesa
|
On-site or off-site Effects of Functions |
Goods and Services Valued by Society |
|
|
|
|
Attenuates downstream flood peaks |
Reduces damage from floodwaters (Daily, 1997) |
|
Maintains vegetation structure in arid climates |
Contributes to regional biodiversity through habitat (e.g., forest canopy) provision (Szaro, 1991; Ohmart, 1996; James et al., 2001) |
|
Contributes to fluvial geomorphology |
Creates predictable yet dynamic channel and floodplain dynamics (Beschta et al., 1987a; Klingeman et al., 1999) |
|
|
|
|
Provides energy to maintain aquatic and terrestrial food webs |
Supports populations of organisms (Gregory et al., 1991; Meyer and Wallace, 2001) |
|
Provides reservoirs for genetic diversity |
Contributes to biocomplexity (Szaro, 1991; Naiman and Rogers, 1997; Pollock et al., 1998) |
|
Intercepts nutrients and toxicants from runoff |
Removes pollutants from runoff (Bhowmilk et al., 1980; Peterjohn and Correll, 1984) |
|
Contributes to nutrient retention and to sequestration of carbon dioxide from the atmosphere |
Potentially ameliorates global warming (Van Cleve et al., 1991) |
|
|
|
|
Provides shade to stream during warm season |
Creates habitat for cold-water fish (Beschta et al., 1987b; McCullough, 1999) |
|
Allows daily movements to annual migrations |
Supplies objects for bird watching, wildlife enjoyment,and game hunting (Green and Tunstall, 1992; Flather and Cordell, 1995) |
|
Allows migratory fish to complete life cycles |
Provides fish for food and recreation (Nehlsen et al, 1991; Naiman et al., 2000) |
change) and corresponding sediment regimes (e.g., transport frequency and magnitude, particle sizes). Particularly in arid regions, the types of riparian vegetation present are largely determined by their tolerance of and adaptation to hydrologic disturbances. Managing riparian areas without regard to how they are influenced by the dynamic patterns of adjacent waterbodies ignores a fundamental aspect of how these systems function.
In addition to disturbance regimes, soil moisture plays a significant role in shaping the vegetative structure of riparian areas. In humid regions of the country, vegetation is abundant in both uplands and riparian areas, with species tolerant to high soil moisture and anoxia being more prevalent in riparian areas. In arid regions, plants are often concentrated along the band of adequate soil moisture provided by riparian areas.
A core of tree genera—alders, cottonwoods, and willows—grows in riparian areas across the continent. Gallery stands of these trees are predominant features of many riparian areas throughout the country. In much of the American West, the integrity of riparian areas in low-gradient meadow systems requires intact communities of shrubs (e.g., willows) and hydrophytic graminoids (sedges, rushes, and grasses).
Riparian areas, in proportion to their area within a watershed, perform more biologically productive functions than do uplands. Riparian areas provide a wide range of functions such as microclimate modification and shade, bank stabilization and modification of sedimentation processes, contributions of organic litter and large wood to aquatic systems, nutrient retention and cycling, wildlife habitat, and general food-web support for a wide range of aquatic and terrestrial organisms. Thus, even though they occupy only a small proportion of the total land base in most watersheds, they are uniquely positioned between the aquatic and terrestrial ecosystems to provide a wide range of functions critical for many aquatic and terrestrial species, for maintenance of water quality, for aesthetics, for the production of goods and services, and for a wide range of social and cultural values.
Riparian areas are effective in filtering and transforming materials (such as dissolved and particulate nonpoint source pollutants) from hillslope runoff. Pollutant removal in riparian areas is most effective along first- and second-order streams because a greater percentage of their flow derives from hillslope rather than upstream sources. However, where flows are concentrated into topographic depressions prior to entering a riparian area, the effectiveness of the riparian area for pollutant removal will be greatly reduced.
Because riparian areas are located at the convergence of terrestrial and aquatic ecosystems, they are regional hot spots of biodiversity and often exhibit high rates of biological productivity in marked contrast to the larger landscape. This is particularly dramatic in arid regions, as evidenced by the high number of plant and animal species that find crucial habitats along watercourses and washes. Riparian areas provide connectivity at all spatial and temporal scales, helping maintain landscape biodiversity by countering the negative ecological effects of habitat fragmentation.
REFERENCES
Abbe, T. B., and D. R. Montgomery. 1996. Large woody debris jams, channel hydraulics and habitat formation in large rivers. Regulated Rivers: Research & Management 12(2–3):201–221.
Addy, K. L., A. J. Gold, P. M. Groffman, and P. A. Jacinthe. 1999. Ground water nitrate removal in subsoil of forested and mowed riparian buffer zones. J. Environmental Quality 28(3):962–970.
Ahmad, R., F. N. Scatena, and A. Gupta. 1993. Morphology and sedimentation in Caribbean montane streams: examples from Jamaica and Puerto Rico. Sedimentary Geology 85:157–169.
Amoros, C., J. Rostan, G. Pautou, and J. Bravard. 1987. The reversible process concept applied to the environmental management of large river systems. Environ. Manage. 11(5):607–617.
Anderson, B. W., and R. D. Ohmart. 1977. Vegetation structure and bird use in the lower Colorado River Valley. Paper presented at the symposium on importance, preservation, and management of riparian habitat, Tucson, AZ.
Anderson, N. H., and J. B. Wallace. 1984. Habitat, life history, and behavioral adaptations of aquatic insects. Pp. 38–58 In: An introduction to the aquatic insects of North America. R. W. Merritt and K. W. Cummins (eds.). Dubuque, IA: Kendall/Hunt Publishing Co.
Andrus, R. E. 1990. Why rare and endangered byophytes? In: Ecosystem management: rare species and significant habitats. Mitchell, R. S., Sheviak, C. J. and D. J. Leopold (eds.). Proceedings of the 15th Annual Natural Areas Conference. New York State Museum Bulletin No. 471. Albany, NY.
Angermeier, P. L. 1987. Spatiotemporal variation in habitat selection by fishes in small Illinois streams. Pp. 52–60 In: Community and evolutionary ecology of North American stream fishes. W. J. Matthews and D. C. Heins (eds.). Norman, OK: University of Oklahoma Press.
Avery, E. L. 1992. Effects of removing beaver dams upon a northern Wisconsin brook trout stream. Study No. 406. Madison, WI: Wisconsin Department of Natural Resources, Bureau of Research, Fish Research Section.
Bach, L. 1995. River basin assessment: Upper/middle Grande Ronde River and Catherine Creek. Portland, OR: Oregon Department of Environmental Quality.
Baker, W. L. 1989. Classification of the riparian vegetation of the montane and sub-alpine zones in western Colorado. Great Basin Naturalist 49:214–228.
Bard, A. J., and R. L. Wilby. 1999. Eco-hydrology: plants and water in terrestrial environments. Routledge, London.
Barnes, W. J., and E. Dibble. 1988. The effects of beaver in riverbank forest succession. Can. J. Bot. 66:40–44.
Barton, D. R., W. D. Taylor, and R. M. Biette. 1985. Dimensions of riparian buffer strips required to maintain trout habitat in southern Ontario streams. North American Journal of Fisheries Management 5:364–378.
Bathurst, J. C. 1979. Distribution of boundary shear stress in rivers. Pp. 95–116 In: Adjustment of fluvial systems. D. D. Rhodes and G. P. Williams (eds.). Dubuque, IA: Kendall/Hunt.
Bayley, P. B. 1991. The flood pulse advantage and the restoration of river–floodplain systems. Regulated Rivers: Research and Management 6:75–86.
Bayley, P. B., and H. W. Li. 1992. Riverine fishes. Pp. 252–281 In: The rivers handbook: hydrological and ecological principles, Vol. 1. P. Calow and G. E. Petts (eds.). Oxford: Blackwell Scientific Publications.
Beier, P. 1993. Determining minimum habitat areas and habitat corridors for cougars. Conservation Biology 7:94–108.
Belovsky, G. E. 1984. Summer diet optimization by beaver. The American Midland Naturalist 111:209–222.
Benke, A. C., and J. B. Wallace. 1990. Wood dynamics in coastal plain blackwater streams. Can. J. Fish. Aquat. Sci. 47:92–99.
Bergerud, A. T., and D. R. Miller. 1977. Population dynamics of Newfoundland beaver. Can. J. Zool. 55:1480–1492.
Beschta, R. L., T. Blinn, G. E. Grant, G. G. Ice, and F. G. Swanson (editors). 1987a. Erosion and sedimentation in the Pacific Rim. International Association of Hydrological Sciences, Publication No. 165. 510 pp.
Beschta, R. L., R. E. Bilby, G. W. Brown, L. B. Holtby, and T. D. Hofstra. 1987b. Stream temperature and aquatic habitat: fisheries and forestry interactions. Pp. 191–232 In: Streamside management: forestry and fisheries comparisons. E. O. Salo and T. W. Cundy (eds.). Seattle, WA: University of Washington, Institute of Forest Resources, Contribution No. 57. 471 pp.
Bhowmilk, N. G., Bonini, A. P., Bogner, W. C., and Byrne, R. P. 1980. Hydraulics of flow and sediment transport to the Kankakee River in Illinois. Illinois State Water Survey Rep. of Investigation 98, Champaign, Ill., 170 p.
Bisson, P. A., T. P. Quinn, G. H. Reeves, and S. V. Gregory. 1992. Best management practices, cumulative effects, long-term trends in fish abundance in Pacific Northwest river systems. Pp. 189–232 In: Watershed management: balancing sustainability and environmental change. R. J. Naiman (ed.). New York: Springer-Verlag. 542 p.
Bisson, P.A., R. E. Bilby, M. D. Bryant, C. A. Dolloff, G. B. Grette, R. A. House, M. L. Murphy, K. V. Koski, and J. R. Sedell. 1987. Large woody debris in forested streams in the Pacific Northwest: Past, present, and future. Pp. 143–190 In: Streamside management: forestry and fishery interactions. E. O. Salo and T. W. Cundy (eds.). Seattle, WA: University of Washington, Institute of Forest Resources, Contribution No. 57.
Bisson, P. A., S. V. Gregory, and T. Nickelson. 2002. Long-term variation in anadromous and resident salmonid populations. In: Ecological and Environmental Research in the Alsea Watershed. J. Stednick (ed.). Springer-Verlag.
Bliss, L. C., and J. E. Cantlon. 1957. Succession on river alluvium in northern Alaska. American Midland Naturalist 58:452–469.
Bohlke, J. K., and J. M. Denver. 1995. Combined use of ground-water dating, chemical and isotopic analyses to resolve the history and fate of ground-water nitrate contamination in two agricultural watersheds, Atlantic coastal plain, Maryland. Water Resources Research 31: 2319–2339.
Boose, A. B., and J. S. Holt. 1999. Environmental effects on asexual reproduction in Arundo donax. Weed Research 39:117–127.
Boyd, M., B. Kasper, and A. Hamel. 1998 (draft). Tillamook basin temperature total maximum daily load (TMDL). Oregon Department of Environmental Quality, Portland. 42 pp. + appendices.
Boyer, E. W., G. M. Hornberger, K. E. Bencala, and D. M. McKnight. 1997. Response characteristics of DOC flushing in an alpine catchment. Hydrological Processes 11:1635–1647.
Bradt, G. W. 1938. A study of beaver colonies in Michigan. J. Mammalogy 19:139–162.
Braun, C. E. 1998. Sage grouse declines in western North America: What are the problems? Proceedings of the Western Association of Fish and Wildlife Agencies 78:139–156.
Brinson, M. M. 1990. Riverine forests. In: Forested wetlands. A. E. Lugo, M. Brinson, and S. Brown (eds.). New York: Elsevier.
Brinson, M. M. 1991. Landscape properties of pocosins and associated wetlands. Wetlands 11:441–465.
Brinson, M. M. 1993. Changes in the functioning of wetlands along environmental gradients. Wetlands 13:65–7.
Brinson, M. M. and J. Verhoeven. 1999. Riparian forests. In: Maintaining biodiversity in forested ecosystems. M. L. Hunter (ed.). Cambridge, England: Cambridge University Press.
Brinson, M. M., R. R. Christian, and L. K. Blum. 1995. Multiple states in the sea-level induced transition from terrestrial forest to estuary. Estuaries 18:648–659.
Brock, T. D., and M. T. Madigan. 1991. Biology of Microorganisms. Englewood Cliffs, NJ: Prentice Hall. 873 pp.
Brosofske, K. D., J.Chen, R. J. Naiman, and J. F. Franklin. 1997. Effects of harvesting on micro-climaic gradients from small streams to uplands in western Washington. Ecological applications 7(4):1188–1200.
Brown, C. R., M. B. Brown, P. A. Johnsgard, J. Kren, W. C. Scharf. 1996. Birds of the Cedar Point Biological Station area, Keith and Garden counties, Nebraska: Season occurrence and breeding data. Transactions of the Nebraska Academy of Sciences 23:91–108.
Brown, G. W. 1969. Predicting temperatures of small streams. Water Resources Research 5:68–75.
Bruner, W. E. 1931. The vegetation of Oklahoma. Ecological Monographs 1:99–188.
Bryan, M. D. and D. L. Scarnecchia. 1992. Species richness, composition, and abundance of fish larvae and juveniles inhabiting natural and developed shorelines of a glacial Iowa lake. Environ. Biol. of Fishes 35:329–341.
Budd, W. W., P. L. Cohen, and P. R. Saunders. 1987. Stream corridor management in the Pacific Northwest: I. Determination of stream corridor widths. Environmental Management 11:587–597.
Buell, M. F., and W. A. Wistendahl. 1955. Flood plain forests of the Raritan River. Bulletin of the Torrey Botanical Club 82:463–472.
Burnett, A. W., and S. A. Schumm. 1983. Alluvial-river response to neotectonic deformation in Louisiana and Mississippi. Science 222:49–50.
Burt, W. H., and Grossenheider. 1976. A field guide to the mammals. Boston, MA: Houghton Mifflin Company.
Carpenter, S. R., E. van Donk, and R. G. Wetzel. 1998. Effects of the nutrient loading gradient in shallow lakes. Pp. 393–396 In: The role of submersed macrophytes in structuring the biological community and biogeochemical dynamics in shallow lakes. Dordrecht, Netherlands: Kluwer Publishers.
Chapin, D. M., R. L. Beschta, and J. W. Shen. 2000. Flood frequencies required to sustain riparian plant communities in the Upper Klamath Basin, Oregon. Pp. 17–22 In: International conference on riparian ecology and management in multi-land use watersheds. P. J. Wiggington and R. L. Beschta (eds.). Middleburg, VA: American Water Resources Association.
Chapman, D. W. 1965. Net production of juvenile coho salmon in three Oregon streams. Trans. Amer. Fish. Soc. 94:40–52.
Chapman, D. W. 1966. Food and space as regulators of salmonid populations in streams. Amer. Nat. 100:345–357.
Cheng, J. D. 1989. Streamflow changes after clear-cut logging of a pine beetle-infested watershed in southern British Columbia, Canada. Water Resources Research 25:449–456.
Choi, J., and J. Harvey. 2000. Characterizing multiple timescales of stream and storage zone interaction that affect solute fate and transport in streams. Water Resources Research 36(6):1511–1518.
Christensen, N. L., A. M. Bartuska, J. H. Brown, S. Carpenter, C. D’Antonio, R. Francis, J. F. Franklin, J. A. MacMahon, R. F. Noss, D. J. Parsons, C. H. Peterson, M. G. Turner, and R. G. Woodmansee. 1996. The report of the Ecological Society of America committee on the scientific basis for ecosystem management. Ecological Applications 6:665–691.
Clevenger, A. P., and N. Waltho. 2000. Factors influencing the effectiveness of wildlife underpasses in Banff National Park, Alberta, Canada. Conservation Biology 14:47–55.
Collins, S. L., P. G. Risser, and E. L. Rice. 1981. Ordination and classification of mature bottomland forests in North Central Oklahoma. Bulletin of the Torrey Botanical Club 108:152–165.
Cook, D. B. 1940. Beaver-trout relations. J. Mammalogy 21:397–401.
Correll, D. L. 1997. Buffer zones and water quality protection. Pp. 7–20 In: Buffer zones: their processes and potential in water protection. N. E. Haycock,T. P. Burt, K. W. T. Goulding and G. Pinay (eds.). Harpenden, Hertfordshire, UK: Quest Environmental.
Cowan. W. L. 1956. Estimating hydraulic roughness coefficients. Agricultural Engineering 37:473–475.
Crawford C. S., A. C. Culley, R. Leutheuser, M. S. Sifuentes, L. H. White, and J. P. Wilber. 1993. Middle Rio Grande Ecosystem: Bosque Biological Management Plan. Albuquerque, NM: U.S. Fish and Wildlife Service, District 2. 291 pp.
Crow, T. R., M. E. Baker, and B. V. Barnes. 2000. Diversity in riparian landscapes. Pp. 34–66 In: Riparian management in forests of the continental eastern United States. E. S. Verry, J. W. Hornbeck, and C. A. Dolloff (eds.). New York: Lewis Publishers.
Dahm, C. N., N. B. Grimm, P. Marmonier, H. M. Valett, and P. Vervier. 1998. Nutrient dynamics at the interface between surface waters and groundwaters. Freshwater Biology 40:427–451.
Daily, G. C. (ed.). 1997. Nature’s services: Societal dependence on natural ecosystems. Washington, DC: Island Press.
Dallman, M., and R. J. Smith. 1995. Avian predation on Chironomids along the nearshore waters of Lake Huron. Michigan Birds and Natural History 2:201–204.
Daly, C., R. P. Neilson, and D. L. Phillips. 1994. A statistical-topographic model for mapping climatological precipitation over mountainous terrain. J. Appl. Meteor. 33:140–158.
Danehy, R. J., and B. J. Kirpes. 2000. Relative humidity gradients across riparian areas in eastern Oregon and Washington forests. Northwest Science 74(3):224–233.
Daniel, J. F. 1976. Estimating groundwater evapotranspiration from streamflow records. Water Resources Research 12(3):360–364.
Davis, M., C. Douglas, R. Calcote, K. L. Cole, M. G. Winkler, and R. Flakne. 2000. Holocene climate in the western Great Lakes national parks and lakeshores: implications for future climate change. Conservation Biology 14:968–983.
Dawson, T. E., and J. R. Ehleringer. 1993. Streamside trees that do not use streamwater. Nature 350:335–337.
DeGraaf, R. M., and M. Yamasaki. 2000. Bird and mammal habitat in riparian areas. Pp. 139–156 In: Riparian management in forests of the continental eastern United States. E. S. Verry, J. W. Hornbeck, and C.A. Dolloff (eds.). New York: Lewis Publishers.
Dillaha, T. A., R. B. Reneau, S. Mostaghimi, and V. O. Shanholtz. 1989. Vegetative filter strips for nonpoint source pollution control. Transactions of the ASAE 32(2):491–496.
Dingman, S. L. 1984. Fluvial hydrology. New York: W. H. Freeman.
Dirschl, H. J., and R. T. Coupland. 1972. Vegetation patterns and site relationships in the Saskatchewan River delta. Canadian Journal of Botany 50:647–675.
Dobrowolski, K. A., A. Kozakiewicz, and B. Leznicka. 1993. The role of small mammals and birds in transport of matter through the shore zone of lakes. Hydrobiologia 251:81–93.
Dodd, C. K., Jr., and B. S. Cade. 1998. Movement patterns and the conservation of amphibians breeding in small, temporary wetlands. Conservation Biology 12:331–339.
Dong, J., J. Chen, K. D. Brosofske, and R. J. Naiman. 1998. Modeling air temperature gradients across managed small streams in western Washington. Journal of Environmental Management 53:309–321.
Dunne, T., and R. D. Black. 1970. Partial area contributions to storm runoff in a small New England watershed. Water Resources Research 6: 1296–1311.
Dunne, T. 1978. Field studies of hillslope flow processes. Pp. 227–293 In: Hillslope hydrology. M. J. Kirkley (ed.). Chicester: John Wiley.
Everitt, B. L. 1968. Use of cottonwood in an investigation of the recent history of a flood plain. American Journal of Science 266: 417–439.
Ewert, M. A. 1985. Second Year Report: Assessment of the current distribution and abundance of the Wood Turtle (Clemmys insculpta) in Minnesota and along the St. Croix National Scenic Riverway in Wisconsin. Unpublished report for the Minnesota Nongame Wildlife Program and Minnesota Field Office of the Nature Conservancy, 37 p.
Ewert, D. N., and M. J. Hamas. 1996. Ecology of migratory landbirds during migration in the Midwest. Pp. 200–208 In: Management of Midwestern landscapes for the conservation of neotropical migratory birds. Gen. Tech. Rep. NC–187. F. R. Thompson III (ed.). St. Paul, MN: USDA Forest Service North Central Experiment Station.
Faaborg, J. 1988. Ornithology, an ecological approach. Englewood Cliffs, NJ: Prentice Hall. 470 pp.
Faaborg, J., A. D. Anders, M. E. Baltz, and W. K. Gram. 1996. Non-breeding season considerations for the conservation of migratory birds in the Midwest: Post breeding and wintering species. Pp. 189–199 In: Management of Midwestern landscapes for the conservation of neotropical migratory birds. Gen. Tech. Rep. NC–187. F. R. Thompson III (ed.). St. Paul, MN: USDA Forest Service North Central Experiment Station.
Federal Interagency Working Group. 1998. Stream corridor restoration. principles, processes, and practices. Washington, DC: National Technical Information Service, U.S. Department of Commerce.
Findlay, S. 1995. Importance of surface–subsurface exchange in stream ecosystems: the hyporheic zone. Limnology and Oceanography 40: 159–164.
Fisher, S. G., N. B. Grimm, E. Marti, R. M. Holmes, and J. B. Jones. 1998. Material spiraling in stream corridors: a telescoping ecosystem model. Ecosystems 1:19–34.
Fisher, S. G., and G. E. Likens. 1973. Energy flow in Bear Brook, New Hampshire: an integrative approach to stream ecosystem metabolism. Ecological Monographs 43(4): 421–439.
Fisk, H.N., and E. McFarland, Jr. 1955. Later Quaternary deltaic deposits of the Mississippi River. Geological Society of America Special Paper 62:279–302.
Flanagan, L. B., J. R. Ehleringer and T. E. Dawson. 1992. Water sources of plants growing in woodland, desert and riparian communities: Evidence from stable isotope analysis. In: Proceedings of the symposium on ecology and management of riparian shrub communities. Gen. Tech. Rep. INT-289. Clary, W. P., E. D. Durant McArthur, D. Bedunah and C. L. Wambolt (eds.). Ogden, UT: USDA Forest Service Intermountain Research Station.
Flather, C. H., and H. K. Cordell. 1995. Outdoor recreation: historical and anticipated trends. Pp. 3–16 In: Wildlife and recreationists: historical and anticipated trends. R. L. Knight and K. J. Gutzwiller (eds.). Washington, DC: Island Press.
Ford, T. E., and R. J. Naiman. 1988. Alteration of carbon cycling by beaver: methane evasion rates from boreal forest streams and rivers. Can. J. Zool. 66:529–533.
Franklin, J. F., and C. T. Dyrness. 1988. Natural vegetation of Oregon and Washington. Corvallis, OR: Oregon State University Press.
Friedman, J. M., and G. T. Auble. 2000. Floods, flood control, and bottomland vegetation. Pp. 219–237 In: Inland flood hazards: human, riparian, and aquatic communities. E. Wohl (ed.), Cambridge, England: Cambridge University Press.
Friedman, J. M., W. R. Osterkamp, and W. M. Lewis, Jr. 1996a. Channel narrowing and vegetation development following a Great Plains flood. Ecology 77(7):2167–2181.
Friedman, J. M., W. R. Osterkamp, and W. M. Lewis, Jr. 1996b. The role of vegetation and bed-level fluctuations in the process of channel narrowing. Geomorphology 14:341–351.
Frissell, C. A. 1993. A new strategy for watershed restoration and recovery of Pacific Salmon in the Pacific Northwest. Eugene, OR: Pacific Rivers Council, Inc.
Garten, C. T., Huston, M. A., Thomes, C. A. 1994. Topographic variation of soil nitrogen dynamics at Walker Branch Watershed, Tennessee. Forest Science 40(3):497–512.
Gatewood, J. S., J. W. Robinson, B. R. Colby, J. D. Hem, and L. C. Halpenny. 1950. Use of water by bottomland vegetation in the Lower Safford Valley, Arizona. U.S. Geological Survey Water-Supply Paper 1103.
Geraghty, J. J., D. W. Miller, F. van der Leeden, and F. L. Troise. 1973. Water Atlas of the United States. Third edition. Port Washington, NY: Water Information Center.
Gibert, J., J. A. Stanford, M.-J. Dole-Oliver, and J. V. Ward. 1994. Basic attributes of groundwater ecosystems and prospects for research. Pp. 7–40 In: Groundwater ecology. J. Gibert, D. L. Danielopol, and J. A. Stanford (eds.). San Diego, CA: Academic Press, Inc.
Gill, F. B. 1990. Ornithology. New York: W. H. Freeman and Co. 660 pp.
Gilliam, J. W., J. E. Parsons, and R. L. Mikkelsen. 1997. Nitrogen dynamics and buffer zones. Pp. 54–61 In: Buffer zones: their processes and potential in water protection. N. E. Haycock, T. P. Burt, K. W. T. Goulding and G. Pinay (eds.). Harpenden, Hertfordshire, UK: Quest Environmental.
Gillilan, D., and T. Brown. 1997. Instream Flow Protection: Seeking a Balance in Western Water Uses. Washington, DC:Island Press.
Goodrich, D. C. et al. 2000. Seasonal estimates of riparian evapotranspiration using remote and in situ measurements. Agricultural and Forest Meteorology 105(1–3):281–310.
Gorman, O. T., and J. R. Karr. 1978. Habitat structure and stream fish communities. Ecology 59:507–515.
Gould, W. A., and M. D. Walker. 1999. Plant communities and landscape diversity along a Canadian arctic river. Journal of Vegetation Science 10:537–548.
Green, C. H., and S. M Tunstall. 1992. The amenity and environmental value of river corridors in Britain. In: River conservation and management. New York: John Wiley & Sons.
Gregory, S. V., and P. A. Bisson. 1996. Degradation and loss of anadromous salmonid habitat in the Pacific Northwest. Pp. 277–314. In: Pacific salmon and their ecosystems: status and future options. D. Stouder and R. J. Naiman (eds.). New York: Springer-Verlag.
Gregory, K. J., and D. E. Walling. 1973. Drainage basin form and process. New York: John Wiley and Sons. 456 pp.
Gregory, S. V., F. J. Swanson, W. A. McKee, and K. W. Cummins. 1991. An ecosystem perspective of riparian zones. BioScience 41:540–551.
Groffman, P. M., A. J. Gold, and R. C. Simmons. 1992. Nitrate dynamics in riparian forests: microbial studies. J. Environ. Qual. 21:666–671.
Hair, J. D., G. T. Hepp, L. M. Luckett, K. P. Reese, and D. K. Woodward. 1978. Beaver pond ecosystems and their relationships to multi-use natural resource management. Pp. 80–92 In: Strategies for protection and management of floodplain wetlands and other riparian ecosystems. Proceedings of the Symposium, Dec. 11–13, 1978, Callaway Gardens, GA. Washington, DC: USDA and USFS.
Hall, F. R. 1968. Base-flow recessions—a review. Water Resources Research 4(5):973–983.
Hall, J. D., and R. L. Lantz. 1969. Effects of logging on the habitat of coho salmon and cutthroat trout in coastal streams. Pp. 355–375 In: Salmon and trout in streams. T. G. Northcote (ed.). H. R. MacMillan Lectures in Fisheries. Vancouver, British Columbia: University of British Columbia.
Hall, J. D., G. W. Brown, and R. L. Lantz. 1987. The Alsea watershed study: a retrospective. Pp. 399–416 In: Streamside management: forestry-fisheries interactions. E. O. Salo and T. W. Cundy (eds.). Seattle, WA: University of Washington Institute of Forest Resources.
Harding, J. H. 1997. Amphibians and reptiles of the Great Lakes region. Ann Arbor, MI: The University of Michigan Press. 378 pp.
Harmon, M. E., J. F. Franklin, F. J. Swanson, P. Sollins, S. V. Gregory, J. D. Lattin, N. H. Anderson, S. P. Cline, N. G. Aumen, J. R. Sedell, G. W. Lienkaemper, K. Cromack, Jr., and K. W. Cummins. 1986. Ecology of coarse woody debris in temperate ecosystems. Advances in Ecological Research 15:133–302.
Hartman, G. F., J. C. Scrivener, L. B. Holtby, and L. Powell. 1987. Some effects of different streamside treatments on physical conditions and fish population processes in Carnation Creek, a coastal rain forest stream in British Columbia. Pp. 330–372 In: Streamside management: forestry-fisheries interactions. E. O. Salo and T. W. Cundy (eds.). Seattle, WA: University of Washington Institute of Forest Resources.
Harvey, J. W., et al. 1991. Preliminary investigation of the effect of hillslope hydrology on the mechanics of solute exchange between streams and subsurface gravel zones. Pp. 413–418 In: U.S. Geological Survey Toxic Substances Hydrology Program—Proceedings of the technical meeting, Monterey, California, March 11–15, 1991. G. E. Mallard and D. A. Aronson (eds.). Water Resources Investigations Report 91-4034.
Harvey, J. W., and K. E. Bencala. 1993. The effect of streambed topography on surface-subsurface water interactions in mountain catchments. Water Resources Research 29:89–98.
Hauer, F. R., J. A. Stanford, J. J. Giersch, and W. H. Lowe. 2000. Distribution and abundance patterns of macroinvertebrates in a mountain stream: an analysis along multiple environmental gradients. Verh. Internat. Verein. Limnol. 27:1–4.
Hawkins, C. P., M. L. Murphy, and N. H. Anderson. 1982. Effect of canopy, substrate composition, and gradient on the structure of macroinvertebrate communities in Cascade Range streams of Oregon. Ecology 63:1840–1856.
Hayes, J. C., and J. E. Hairston. 1983. Modeling the long-term effectiveness of vegetative filters as on site sediment controls. ASAE Paper No. 83-2081. St. Joseph, MI: Am. Soc. Agric. Engrs. 27 pp.
Heinselman, M. L. 1970. Landscape evolution, peatland types, and the environment in the Lake Agassiz Peatlands Natural Area, Minnesota. Ecological Monographs 40:235–261.
Helmers, D. L. 1992. Shorebird management manual. Manomet, MA: Western Hemisphere Shorebird Reserve Network. 58 pp.
Henderson, C. L., C. C. Dindorf, F. J. Rozumalski. 1998. Landscaping for Wildlife and Water Quality. Minneapolis-St. Paul, MN: Minnesota Department of Natural Resources.
Hewlett, J. K., and A. R. Hibbert. 1967. Factors affecting the response of small watersheds to precipitation in humid areas. Pp. 275–290 In: International symposium on forest hydrology. W. E. Sopper and H. W. Lull (eds.). Oxford, England: Pergamon Press. 813 pp.
Hickin, E. J. 1984. Vegetation and river channel dynamics. Canadian Geographer 28:111–126.
Hicks, B. J., R. L. Beschta, and D. Harr. 1991. Long-term changes in streamflow following logging—western Oregon and associated fisheries implications. Water Resources Bulletin 27(2):217–226.
Hinkle, S. R., J. H. Duff, F. J. Triska, A. Laenen, E. B. Gates, K. E. Bencala, D. A. Wentz, and S. R. Silva. 2001. Linking hyporheic flow and nitrogen cycling near the Willamette River—a large river in Oregon, USA. J. Hydrol. 244:157–180.
Hoctor, T. S., M. H. Carr, and P. D. Zwick. 2000. Identifying a linked reserve system using a regional landscape approach: the Florida ecological network. Conservation Biology 14:984–1000.
Holstein, G. 1984. California riparian forests: deciduous islands in an evergreen sea. Pp. 2–22 In: California riparian systems. R. E. Warner and K. M. Hendrix (eds.). Berkeley, CA: University of California Press.
Hughes, J. W., and W. B. Cass. 1997. Pattern and process of a floodplain forest, Vermont, USA: predicted responses of vegetation to perturbation. Journal of Applied Ecology 34:594–612.
Hunt, R. J., D. P. Krabbenhoft, and M. P. Anderson. 1996. Groundwater inflow measurements in wetland systems. Water Resources Research 32(3):495–507.
Hunter, M. L., Jr. 1996. Fundamentals of Conservation Biology. Cambridge, MA: Blackwell Science. 482 pp.
Hupp, C. R. 2000. Hydrology, geomorphology and vegetation of Coastal Plain rivers in the southeastern USA. Hydrological Processes 14:2991–3010.
Hupp, C. R., and W. R. Osterkamp. 1985. Bottomland vegetation distribution along Passage Creek, Virginia, in relation to fluvial landforms. Ecology 66:670–681.
Hupp, C. R., and W. R. Osterkamp. 1996. Riparian vegetation and fluvial geomorphic processes. Geomorphology 14:277–295.
Huston, S. L. 1978. Prairie remnants along the Stillwater River in Miami County, Ohio. In: The Prairie Peninsula—in the “Shadow” of Transeau. Proceedings of the Sixth North American Prairie Conference, Ohio State University, Columbus, Ohio. Ohio Biological Survey Biological Notes No. 15.
Hyde, D. 1998. Stopover ecology of migratory birds along the central Lake Michigan shoreline. M.S. thesis, Central Michigan University, Mt. Pleasant, MI.
Inamdar, S. P., R. R. Lowrance, L. S. Altier, R. G. Williams and R. K. Hubbard. 1999. Riparian ecosystem management model (REMM): II. Testing of the water quality and nutrient cycling component for a Coastal Plain riparian system. Trans. Am. Soc. Agr. Eng. 42:1691–1707.
James, A., K. J. Gaston, and A. Balmford. 2001. Can we afford to conserve biodiversity? BioScience 51:43–52.
James, F. C. 1971. Ordination of habitat relationships among breeding birds. Wils. Bull. 83:215–236.
Jenkins, S. H. 1980. A size-distance relation in food selection by beavers. Ecology 61:740–746.
Jenkins, S. H., and P. E. Busher. 1979. Castor canadensis. Mammalian Species 120:1–9.
Johnson, A. S. 1989. The thin green line: riparian corridors and endangered species in Arizona and New Mexico. Pp. 35–46 In: Preserving communities and corridors. G. Mackintosh (ed.). Washington, DC: Defenders of Wildlife.
Johnson, G. D., and M. S. Boyce. 1990. Feeding trials with insects in the diet of sage grouse chicks. J. Wildl. Manage. 54:89–91.
Johnson, R. R., P. S. Bennett, and L. Haight. 1989. Southwestern woody riparian vegetation and succession: an evolutionary approach. Pp. 135–139 In: Proceedings of the California Riparian Systems Conference. D. L. Abell (ed.). USDA Forest Service General Technical Report PSW—110.
Johnson, W. C., R. L. Burgess, and W. R. Keammerer. 1976. Forest overstory vegetation and environment on the Missouri River floodplain in North Dakota. Ecological Monographs 46:59–84.
Johnston, C. A., and R. J. Naiman. 1990. Browse selection by beaver: effects on riparian forest composition. Can. J. For. Res. 20:1036–1043.
Jones, K. B., and P. C. Glinski. 1985. Microhabitats of lizards in a southwestern riparian community. Pp. 342–346 In: Riparian ecosystems and their management: reconciling conflicting uses. USDA Forest Service Gen. Tech. Bull. RM-120.
Jones, J. B., and P. J. Mulholland. 2000. Streams and ground waters. San Diego, CA: Academic Press.
Jordan, J. W. 2001. Late Quaternary sea level change in Southern Beringia: postglacial emergence of the Western Alaska Peninsula. Quaternary Science Reviews 20:509–523.
Judziewicz, E. J., and R. G. Koch. 1993. Flora and vegetation of the Apostle Islands National Lakeshore and Madeline Island, Ashland and Bayfield Counties, Wisconsin. The Michigan Botanist 32 (2):43–193.
Junk, W. J., Bayley, P. B., and Sparks, R. E. 1989. The flood pulse concept in river floodplain systems. Pp. 110–127 In: Proceedings of the International Large River Symposium. D. P. Dodge (ed.). Can. Spec. Publ. Fish. Aquat. Sci. 106.
Kalliola, R., and M. Puhakka. 1988. River dynamics and vegetation mosaicism: a case study of the River Kamajohka, northernmost Finland. Journal of Biogeography 15:703–719.
Kalliola, R., J. Salo, M. Puhakka, and M. Rajasilta. 1991. New site formation and colonizing vegetation in primary succession on the western Amazon floodplains. Journal of Ecology 79:877–901.
Karr, J. R., and R. R. Roth. 1971. Vegetation structure and avian diversity in several new world areas. Am. Nat. 105:423–435.
Kauffman, J. B., and W. C. Krueger. 1984. Livestock impacts on riparian ecosystems and streamside management implications. Journal of Range Management 37:430–437.
Kelsey, K. A., and S. D. West. 1998. Riparian wildlife. Pp. 235–258 In: River ecology and management: lessons from the Pacific Coastal ecoregion. R. J. Naiman and R. E. Bilby (eds.). New York: Springer-Verlag.
Keppeler, E. T., and R. R. Ziemer. 1990. Logging effects on streamflow-water yield and summer low flows at Caspar Creek in northwestern California. Water Resources Research 26:1669–1679.
Kirby, R. E. 1975. Wildlife utilization of beaver flowages on the Chippewa National Forest, North Central Minnesota. The Loon Winter:180–181.
Klebenow, D. A., and G. M. Gray. 1968. Food habits of juvenile sage grouse. J. Range Manage. 21:80–83.
Klingeman, P. C., R. L. Beschta, P. D. Komar, and J. B. Bradley. 1999. In: Gravel-bed rivers in the environment. Highland Ranch, CO: Water Resources Publications. 832 pp.
Klingeman, P. C., and J. B. Bradley. 1976. Willamette River basin streambank stabilization by natural means. Corvallis, OR: Oregon State University Water Resources Research Institute.
Korth, R., and P. Cunningham. 1999. Margin of error? Human influence on Wisconsin shores. Wisconsin Lakes Partnership (Wisconsin Association of Lakes, Wisconsin Department of Natural Resources, and University of Wisconsin-Extension). UWEX Lakes, University of Wisconsin, Stevens Point, WI.
Krabbenhoft et al., 1990. Estimating groundwater exchange with lakes—1. the stable isotope mass balance method. Water Resources Research 26(10):2445–2453.
Kratz, T. K., K. E. Webster, C. J. Bowser, J. J. Magnuson, and B. J. Benson. 1997. The influence of landscape position on lakes in northern Wisconsin. Freshwater Biology 37:290–17.
Labaugh, J. W., Winter, T. C., Swanson, G. A., Rosenberry, D. O., Nelson, R. D., and Euliss, N. H. 1996. Changes in atmospheric circulation patterns affect midcontinent wetlands sensitive to climate. Limnology and Oceanography 41(5):864–870.
Larkin, R. G., and J. M. Sharp. 1992. On the relationship between river-basin geomorphology, aquifer hydraulics, and ground-water flow direction in alluvial aquifers. Geological Society of America Bulletin 104:1608–1620.
Larue, P. L. Belanger, and J. Huot. 1995. Riparian edge effects on boreal balsam fir bird communities. Can. J. For. Res. 25:555–566.
Law, D. J., C. B. Marlow, J. C. Mosley, S. Custer, P. Hook and B. Leinard. 2000. Water table dynamics and soil texture of three riparian plant communities. Northwest Science 74(3):233–241.
Lawrence, W. H. 1952. Evidence of the age of beaver ponds. J. Wildlife Management 16:69–78.
Lee, D., T. A. Dillaha, and J. H. Sherrard. 1989. Modeling phosphorus transport in grass buffer strips. J. Environ. Eng. 115:409–427.
Lee, L. C. 1983. The floodplain and wetland vegetation of two Pacific Northwest river ecosystems. Ph.D. Dissertation, University of Washington, Seattle, WA.
Leidholt-Bruner, K., D. E. Hibbs, and W. C. McComb. 1992. Beaver dam locations and their effects on distribution and abundance of coho salmon fry in two coastal Oregon streams. Northwest Science 66:218–223.
Leopold, L. B., M. G. Wolman, and J. P. Miller. 1964. Fluvial Processes in Geomorphology. San Francisco, CA: W. H. Freeman and Co.
Li, R., and H. W. Shen. 1973. Effect of tall vegetation on flow and sediment. American Society of Civil Engineers Journal of the Hydraulics Division 99:793–814.
Lindsey, A. A., R. O. Petty, D. K Sterlin, and W. Van Asdall. 1961. Vegetation and environment along the Wabash and Tippecanoe Rivers. Ecological Monographs 31:105–156.
Long, C. A., and C. F. Long. 1992. Some effects of land use on avian diversity in a Wisconsin’s oak– pine savanna and riparian forest. Passenger Pigeon 54:125–136.
Lowe, C. H. 1961. Biotic communities in the sub-Mogollon region of the inland Southwest. Journal of the Arizona Academy of Science 2:40–49.
Lowe, C. H. 1964. The Vertebrates of Arizona. Tucson, AZ: The University of Arizona Press. 270 pp.
Lowe, C. H. 1989. The riparianness of a desert herptofauna. USDA Forest Service General Technical Report PSW-110. Washington, DC: USDA Forest Service.
Lowrance et al., 1995. Water quality functions of riparian forest buffer systems in the Chesapeake Bay watershed. Report No. EPA-R-95-004. Annapolis, MD: Chesapeake Bay Program. 67 pp.
Lugo, A. E., S. L. Brown, R. Dodson, T. S. Smith, and H. H. Shugart. 1999. The Holdridge life zones on the conterminous United States in relation to ecosystem mapping. Journal of Biogeography 26:1025–1038.
Lytjen, D. 1998. Ecology of woody riparian vegetation in tributaries of the upper Grande Ronde River basin, Oregon. M.S. thesis, Oregon State University, Corvallis. 76 pp.
MacArthur, R. H. 1964. Environmental factors affecting bird species diversity. Am. Nat. 98:387–397.
MacDonald, B. C., and C. P. Lewis. 1973. Geomorphic and sedimentologic processes of rivers and coast, Yukon Coastal Plain. Geological Survey of Canada. Cat. No. R72-12173. Ottawa, Canada: Information Canada.
Machtans, C., M. Villard, and S. J. Hannon. 1996. Use of riparian buffer strips as movement corridors by forest birds. Conservation Biology 1366–1379.
MacNish, R. D., C. L. Unkrich, E. Smythe, D. C. Goodrich, and T. Maddock, III. 2000. Comparison of riparian evapotranspiration estimates based on a water balance approach and sap flow measurements. Agricultural and Forest Meteorology 105(1–3):271–280.
Malanson, G. P. 1993. Riparian Landscapes. Cambridge Studies in Ecology. Cambridge, UK: Cambridge University Press.
Maser, C., and J. R. Sedell. 1994. From the forest to the sea: the ecology of wood in streams. Delray Beach, FL: St. Lucie Press. 200 p.
Maser, C., R. F. Tarrant, J. M. Trappe, and J. F. Franklin (eds.). 1988. From the forest to the sea: a story of fallen trees. General Technical Report PNW-GTR-229. Portland, OR: USDA Forest Service. 153 pp.
Mattoon, W. R. 1915. The Southern Cypress. Bulletin 272. Washington DC: USDA. 74 pp.
McCullough, D. A. 1999. A review and synthesis of effects of alterations to the water temperature regime of freshwater life stages of salmonids, with special reference to chinook salmon. Water Resource Assessment EPA 910-R-99-010. Portland, OR: Columbia River Inter-Tribal Fish Commission. 291 pp.
McDowell, D. M., and R. J. Naiman. 1986. Structure and function of a benthic invertebrate stream community as influenced by beaver (Castor canadensis). Oecologia 68:481–489.
McGarigal, K., and W. C. McComb. 1992. Streamside versus upslope breeding bird communities in the central Oregon coast range. Journal of Wildlife Management 56:10–23.
McRae, G., and C. J. Edwards. 1994. Thermal characteristics of Wisconsin headwater streams occupied by beaver: implications for brook trout habitat. Transactions of the American Fisheries Society 123:641–656.
Meade, R. H., T. R. Yuzyk, and T. J. Day. 1990. Movement and storage of sediment in rivers of the United States and Canada. Pp. 255–280 In: Surface water hydrology. M. G. Wolman and H. C. Riggs (eds.). Boulder, CO; Geological Society of America.
Meyer, J. L., and J. B. Wallace. 2001. Lost linkages and lotic ecology: rediscovering small streams. Pp. 295–317 In: Ecology: achievement and challenge. M. C. Press, N. J. Huntly, and S. Levin (eds.). Oxford: Blackwell Scientific Publications.
Midwest Plan Service. 1985. Livestock Waste Facilities Handbook. Ames, IA: Iowa State University.
Minshall, G. W. 1988. Stream ecosystem theory: a global perspective. Journal of the North American Benthological Society 7:263–288.
Minshall, G. W. 1989. The ecology of stream and riparian habitats of the Great Basin region: a community profile. U.S. Fish and Wildlife Service Biological Report 85(7.24).
Moench, A. F., Sauer, V. B., and M. E. Jennings. 1974. Modification of routed streamflow by channel loss and base flow. Water Resources Research 10(5):963–968.
Montgomery, D. R., T. B. Abbe, J. M. Buffington, N. P. Peterson, K. M. Schmidt, and J. D. Stock. 1996. Distribution of bedrock and alluvial channels in forested mountain drainage basins. Nature 381:587–589.
Moore, F. R., and T. R. Simons. 1992. Habitat suitability and stopover ecology of neotropical landbird migrants. Pp. 345–355 In: Ecology and conservation of neotropical migrant landbirds. J. M. Hagan, III, and D. W. Johnston (eds.). Washington, DC: Smithsonian Institution Press.
Moran, M. S., and P. Heilman (eds.). 2000. Special Issue: Semi-Arid Land-Surface-Atmosphere (SALSA) Program. Agricultural and Forest Meteorology 105(1–3). 323 pp.
Morris, L. A. 1977. Evaluation, classification and management of the floodplain forest of south central New York. M.S. Thesis, SUNY College Environmental Science and Forestry, Syracuse, N.Y.
Mosley, M. P. 1983. Variability of water temperatures in the braided Ashley and Rakaia rivers. New Zealand Journal of Marine and Freshwater Research 17:331–342.
Moyle, P. B., and R. M. Yoshiama. 1994. Protection of aquatic biodiversity in California: a five-tiered approach. Fisheries 19:6–19.
Muldavin, E., P. Durkin, M. Bradley, M. Stuever, and P. Melhop. 2000. Handbook of wetland vegetation communities of New Mexico. Albuquerque, NM: New Mexico Heritage Program.
Mundie, J. H. 1969. Ecological implications of diet of juvenile coho in streams. Pp. 135–152 In: Symposium on salmon and trout in streams. T. G. Northcote (ed.). H. R. MacMillan Lectures in Fisheries, Univ. of British Columbia.
Murphy, M. L., C. P. Hawkins, and N. H. Anderson. 1981. Effects of canopy modification and accumulated sediment on stream communities. Transactions of the American Fisheries Society 110:469–478.
Murray, N. L., and Stauffer, D. F. 1995. Nongame bird use of habitat in central Appalachian riparian forests. J. Wildl. Manage. 59:78–88.
Naiman, R. J., and K. H. Rogers. 1997. Large animals and system-level characteristics in river corridors. BioScience 47:521–529.
Naiman, R. J., C. A. Johnston, J. C. Kelley. 1988. Alteration of North American streams by beaver. BioScience 38:753–761.
Naiman, R. J., D. G. Lonzarich, T. J. Beechie, and S. C. Ralph. 1992. General principles of classification and the assessment of conservation potential in rivers. Pp. 93–123 In: River conservation and management. P. Boon, P. Calow, and G. Petts (eds.). Chichester, UK: Wiley and Sons.
Naiman, R. J., H. Décamps, and M. Pollock. 1993. The role of riparian corridors in maintaining regional biodiversity. Ecological Applications 3:209–212.
Naiman, R. J., J. M. Melillo, and J. E. Hobbie. 1986. Ecosystem alteration of boreal forest streams by beaver (Castor canadensis). Ecology 67:1254–1269.
Naiman, R. J., R. E. Bilby, and P. A. Bisson. 2000. Riparian ecology and management in the Pacific coastal rain forest. BioScience 50:996–1011.
National Research Council (NRC). 1995. Wetlands: characteristics and boundaries. Washington, DC: National Academy Press.
NRC. 1996. Upstream: salmon and society in the Pacific Northwest. Washington, DC: National Academy Press.
Nehlsen, W., J. E. Williams, and J. A. Lichatowich. 1991. Pacific salmon at the crossroads: stocks at risk from California, Oregon, Idaho, and Washington. Fisheries 16:4–21.
Newbold, J. D., D. C. Erman, and K. B. Roby. 1980. Effects of logging on macroinvertebrates in streams with and without buffer strips. Can. J. Fish. Aquat. Sci. 37:1077–1085.
Newbold, J. D., J. W. Elwood, R. V. O’Neill, and W. Van Winkle. 1982. Nutrient spiraling in streams: implications for nutrient limitation and invertebrate activity. American Naturalist 120:628–652.
Nilsson, C. 1992. Conservation management of riparian communities. Pp. 352–372 In: Ecological principles of nature conservation. L. Hansson (ed.). London, England: Elsevier Applied Science.
Nilsson, C., R. Jansson, and U. Zinko. 1997. Long-term responses of river-margin vegetation to water-level regulation. Science 276:798–800.
Nolet, B. A., A. Hoekstra, and M. M. Ottenheim. 1994. Selective foraging on woody species by the beaver Castor fiber, and its impact on a riparian willow forest. Biological Conservation 70:117–128.
Noss, R. F., H. B. Quigley, M. G. Hornocker,T. Merrill, and P. C. Paquet. 1996. Conservation biology and carnivore conservation in the Rocky Mountains. Conservation Biology 10:949–963.
Odum, E. P. 1971. Fundamentals of Ecology. Philadelphia, PA: W. B. Saunders Company.
Ogle, D. W., and P. M. Mazzeo. 1976. Betula uber, the Virginia round-leaf birch, rediscovered in southwest Virginia. Castanea 41:248–256.
Ohmart, R. D. 1996. Historical and present impacts of livestock grazing on fish and wildlife resources in western riparian habitats. Pp. 245–279 In: Rangeland wildlife. P. R. Krausman (ed.). Denver, CO: Society for Range Management. 440 pp.
Ohmart, R. D., and B. W. Anderson. 1978. Wildlife use values of wetlands in the arid southwestern United States. Pp. 278–295 In: Wetland functions and values: the state of our understanding. Proceedings of the National Symposium on Wetlands. P. E. Greeson, J. R. Clark, and J. E. Clark (eds.). Minneapolis, MN: American Water Resources Association.
Ohmart, R. D., and B. W. Anderson. 1982. North American desert riparian ecosystems. Pp. 433–479 In: Reference handbook on the deserts of North America. G. L. Bender (ed.). Westport, CT: Greenwood Press. 594 pp.
Osborne, L. L., and D. A. Kovacic. 1993. Riparian vegetated buffer strips in water-quality restoration and stream management. Freshwater Biology 29:243–258.
Parker, M., F. J. Wood, Jr., B. H. Smith, and R. G. Elder. 1987. Erosional downcutting in lower order riparian ecosystems: have historical changes been caused by removal of beaver? Pp. 35–38 In: Riparian ecosystems and their management: reconciling conflicting uses. USDA Forest Service Gen. Tech. Bull. RM-120.
Parkin, T. B. 1987. Soil microsites as a source of denitrification variability. Soil Science Society of America Journal 51(5):1194–1199.
Patrick, R. 1995. Rivers of the United States. Volume II: Chemical and Physical Characteristics. New York: John Wiley and Sons, Inc.
Pauley, T. K. J. C. Mitchell, R. R. Buech, and J. J. Moriarty. 2000. Ecology and management of riparian habitats for amphibians and reptiles. Pp. 169–206 In: Riparian management in forests of the continental eastern united States. E. S. Verry, J. W. Hornbeck, and C. A. Dolloff (eds.). New York: Lewis Publishers.
Peterjohn, W. T., and D. L. Correll. 1984. Nutrient dynamics in an agricultural watershed: observations on the role of a riparian forest. Ecology 65:1466–1475.
Peterson, J. G. 1970. The food habits and summer distribution of juvenile sage grouse in central Montana. J. Wildl. Manage. 34:147–154.
Petryk, S., and G. Bosmajian III. 1975. Analysis of flow through vegetation. American Society of Civil Engineers Journal of the Hydraulics Division 101:871–884.
Phillips, J. D., Denver, J. M., Shedlock, R. J., and P. A. Hamilton. 1993. Effect of forested wetlands on nitrate concentrations in groundwater and surface water of the Delmarva Peninsula. Wetlands 13:75–83.
Piegay, H., and A. M. Gurnell. 1997. Large woody debris and river geomorphological pattern: examples from S.E. France and S. England. Geomorphology 19(1–2):99–116.
Pielou, E. C. 1991. After the ice age. Chicago: University of Chicago Press. 366 pp.
Pinay, G., A. Fabre, P. Vervier, and F. Gazelle. 1992. Control of C, N and P distribution in soils of riparian forests. Landscape Ecology 6:121–132.
Pinder, G. F., and S. P. Sauer. 1971. Numerical simulation of flood wave modification due to bank storage effects. Water Resources Research 7(1):63–70.
Poff, N. L., J. D. Allan, M. B. Bain, J. R. Karr, K. L. Prestegaard, B. D. Richter, R. E. Sparks, and J. C. Stromberg. 1997. The natural flow regime: a paradigm for river conservation and restoration. BioScience 47(11):769–784.
Pollock, M. M., R. J. Naiman, and T. A. Hanley. 1998. Plant species richness in riparian wetlands—a test of biodiversity theory. Ecology 79:94–105.
Post, R. A. 1996. Functional profile of black spruce wetlands in Alaska. EPA 910/R-96-006. Seattle, WA: U.S. Environmental Protection Agency.
Premo, D., Premo, B, Rogers, E. I., Tiller, D. J. 1992. The woodland vernal pond: an oasis of diversity. In: Total ecosystem management strategies Vol. 1(3), Amasa, MI: White Water Associates, Inc.
Ralph, S. C., G. C. Poole, L. L. Conquest, and R. J. Naiman. 1994. Stream channel morphology and woody debris in logged and unlogged basins of western Washington. Canadian Journal of Fisheries and Aquatic Sciences 51:37–51.
Råsånen, M. E., J. S. Salo, and R. J. Kalliola. 1987. Fluvial perturbance in the western Amazon basin: regulation by long-term sub-Andean tectonics. Science 238:1398–1401.
Reifsnyder, W. E., and H. W. Lull. 1965. Radian energy in relation to forests. USDA Forest Service, Technical Bull. No. 1344. 111 pp.
Remillard, M. M., G. K. Gruendling, and D. J. Bogucki. 1987. Disturbance by beaver (Castor canadensis) and increased landscape heterogeneity. Pp. 103–122 In: Landscape heterogeneity and disturbance. Ecological Studies, Vol. 64. M. G. Turner (ed.). New York: Springer-Verlag.
Rezendes, P., and P. Roy. 1996. Wetlands, the web of life. A Sierra Club Book. Burlington, VT: Verve Editions.
Rice, J., B. W. Anderson, and R. D. Ohmart. 1984. Comparison of the importance of different habitat attributes to avian community organization. J. Wildl. Manage. 48:895–911.
Rice, J., R. D. Ohmart, and B. W. Anderson. 1983. Habitat selection attributes of an avian community: a discriminant analysis investigation. Ecological Monographs 5:263–290.
Rogers, E. I., D. Tiller, and D. Premo. 1992. Mammal tracking study along the West Fence River in Iron County, Michigan. TEMS Research Brief. Total Ecosystem Management Program Annual Report. (In house report.) Amasa, MI: White Water Associates, Inc.
Rothacher, J. 1970. Increases in water yield following clear-cut logging in the Pacific Northwest. Water Resources Research 6(2): 653–658.
Rudolph, D. C., and J. G. Dickson. 1990. Streamside zone width and amphibian and reptile abundance. The Southwestern Naturalist 35:472–476.
Rutherford, I., B. Abernethy, and I. Prosser. 1999. Pp. 61–78 In: Riparian land management technical guidelines, volume one: principles of sound management. S. Lovett and P. Price (eds.). Canberra, Australia: Land and Water Resources Research Development Corporation.
Ruthsatz, B., and W. Haber. 1981. The significance of small-scale landscape elements in rural areas as refuges for endangered plant species. Proc. Int. Congr. Neth. Soc. Landscape Ecol. Veldhoven, Pudoc, Wageningen.
Rutledge, A. T., and T. O. Mesko. 1996. Estimated hydrologic characteristics of shallow aquifer systems in the Valley and Ridge, the Blue Ridge, and the Piedmont Physiographic Provinces based on analysis of streamflow recession and base flow. U.S. Geological Survey Professional Paper 1422-B. 54 pp.
Satterlund, D. R., and P. W. Adams. 1992. Wildland watershed management. New York: John Wiley & Sons, Inc. 436 pp.
Sauer, J. D. 1988. Plant migration: the dynamics of geographic patterning in seed plant species. Berkeley, CA: University of California Press. 298 pp.
Scatena, F. N., and A. E. Lugo. 1995. Geomorphology, disturbance, and the soil and vegetation of two subtropical wet steepland watersheds of Puerto Rico. Geomorphology 13:199–213.
Schade, J. D., S. G. Fisher, N. B. Grimm, and J. A. Seddon. 2001. The influence of a riparian shrub on nitrogen cycling in a Sonoran desert stream. Ecology 82:3363–3376.
Scharf, W. C., and J. Kren. 1997. Summer diet of Orchard Orioles in southwestern Nebraska. Southwestern Naturalist 42:127–131.
Schlesinger, W. H., and J. M. Melack. 1981. Transport of organic carbon in the world’s rivers. Tellus 33:172–187.
Schlosser, I. J. 1982. Fish community structure and function along two habitat gradients in a headwater stream. Ecological Monographs 52(4):395–414.
Schlosser, I. J. 1991. Stream fish ecology: a landscape perspective. BioScience 41:704–712.
Schloz, J. 2001. The variability in stream temperatures in the Wenatchee National Forest and their relationship to physical, geological, and land management factors. M.S. Thesis. University of Washington, Seattle, WA.
Schultz, R. C., J. P. Colletti, T. M. Isenhart, W. W. Simpkins, C. W. Mize, and M. L. Thompson. 1995. Design and placement of multi-species riparian buffer strip system. Agroforestry Systems 29:201–226.
Schultz, R. C., J. P. Colletti, T. M. Isenhart, C. O. Marquez, W. W. Simpkins, and C. J. Ball. 2000. Riparian forest buffer practices. Pp. 189–281 In: North American agroforestry: an integrated science and practice. Madison, WI: American Society of Agronomy.
Schumm, S. A. 1960. The shape of alluvial channels in relation to sediment type. U.S. Geological Survey Professional Paper 352B, 17–30.
Scott, M. L., J. M. Friedman, and G. T. Auble. 1996. Fluvial process and the establishment of bottomland trees. Geomorphology 14:327–339.
Sebestyen, S. D., and R. L. Schneider. 2001. Dynamic temporal patterns of nearshore seepage flux in a headwater Adirondack lake. Journal of Hydrology 247(3–4):137–150.
Sedell, J. R., and K. J. Luchessa. 1982. Using the historical record as an aid to salmonid habitat enhancement. Pp. 210–223 In: Acquisition and utilization of aquatic habitat inventory information. N. B. Armantrout (ed.). Portland, OR: Western Division, American Fisheries Society.
Sedell, J. R., and R. L. Beschta. 1991. Bringing back the “bio” in bioengineering. American Fisheries Society Symposium 10:160–175.
Seefelt, N. 1997. Foraging behaviors and attack rates of American redstarts and black–throated green warblers. M.S. Thesis, Central Michigan University, Mt. Pleasant, MI.
Semlitsch, R. D. 1998. Biological delineation of terrestrial buffer zones for pond-breeding salamanders. Conservation Biology 12:1113–1119.
Seton, E. T. 1929. Lives of game animals. Vol. 4, Part 2. Rodents, etc. Garden City, New York: Doubleday.
Slough, B. G., and R. M. F. S. Sadleir. 1977. A land capability classification system for beaver (Castor canadensis Kuhl). Can. J. Zool. 55:1324–1335.
Smith, R., M. Hamas, M. Dallman, and D. Ewert. 1998. Spatial variation in the foraging of the black-throated green warbler along the shoreline of northern Lake Huron. The Condor 100:474–484.
Spackman, S. C., and J. W. Hughes. 1995. Assessment of minimum stream corridor width for biological conservation: species richness and distribution along mid-order streams in Vermont, USA. Biological Conservation 71:325–332.
Squillace, P. J. 1996. Observed and simulated movement of bank-storage water. Ground Water 34(1):121–134.
Stanford, J. A., and F. R. Hauer. 1992. Mitigating the impacts of stream and lake regulation in the Flathead River catchment, Montana, USA: An ecosystem perspective. Aquatic Conservation: Marine & Freshwater Ecosystems 2(1):35–63.
Stanford, J. A., and J. V. Ward. 1993. An ecosystem perspective of alluvial rivers: connectivity and the hyporheic corridor. Journal of the North American Benthological Society 12:48–60.
Stanford, J. A. 1998. Rivers in the landscape: introduction to the special issue on riparian and groundwater ecology. Freshwater Biology 40(3):402–406.
Stauffer, D. F., and L. B. Best. 1980. Habitat selection by birds of riparian communities: evaluating effects of habitat alterations. J. Wildl. Manage. 44:1–15.
Stebbins, R. C. 1966. A field guide to western reptiles and amphibians. Boston, MA: Houghton Mifflin Company. 279 pp.
Stine, S., D. Gaines, and P. Vorster. 1984. Destruction of riparian systems due to water development in the Mono Lake watershed. Pp. 528–533 In: California riparian systems. R. E. Warner and K. M. Hendrix (eds.). Berkeley, CA: University of California Press.
Strahler, A. N. 1952. Hypsometric (area-altitude) analysis of erosional topography. Bulletin of the Geological Society of America 63:1117–1142.
Stromberg, J. C., R. Tiller, and B. Richter. 1996. Effects of groundwater decline on riparian vegetation of semiarid regions: the San Pedro, Arizona. Ecological Applications 6:113–131.
Swanson, F. J., S. H. Johnson, S. V. Gregory, and S. A. Acker. 1998. Flood disturbance in a forested mountain landscape. BioScience 48(9):681–689.
Swanson, F. J., S. M. Wondzell, and G. E. Grant. 1992. Landforms, disturbance, and ecotones. Pp. 304–323 In: Landscape boundaries: consequences for biotic diversity and ecological flows. A. J. Hansen and F. di Castri (eds.). New York: Springer Verlag.
Szaro, R. C. 1991. Wildlife communities of southwestern riparian ecosystems. Pp. 174–200 In: Wildlife habitats in managed landscapes. J. E. Rodiek and E. G. Bolen (eds.). Washington, DC: Island Press.
Tockner, K., F. Mallard, and J. V. Ward. 2000. An extension of the flood pulse concept. Hydrological Processes 14:2861–2883.
Trimble, S. W. 1997. Stream channel erosion and change resulting from riparian forests. Geology 25(5):467–469.
Triska, F. J., V. C. Kennedy, R. J. Avanzino, G. W. Zellweger, and K. E. Bencala. 1989. Retention and transport of nutrients in a third-order stream in northwestern California: hyporheic processes. Ecology 70:1893–1905.
Troendle, C. A. 1983. The potential of water yield augmentation from forest management in the Rocky Mountain Region. Water Resources Bulletin 19:359–373.
Tweit, S. J. 2000. The next spotted owl? Audubon Nov.–Dec.
U.S. Department of Agriculture (USDA). 2000. Conservation buffers to reduce pesticide losses. Washington, DC: USDA Natural Resources Conservation Service. 21 pp.
USDA NRCS. 2001. Plants Database. Baton Rouge, LA: National Plant Data Center.
Utter, J. M., and A. W. Hurst. 1990. The significance and management of relict populations of Chamaelirium luteum (L.) Gray. In: Ecosystem management: rare species and significant habitats. R. S. Mitchell, C. J. Sheviak, and D. J. Leopold (eds.). Proceedings of the 15th Annual Natural Areas Conference. New York State Museum Bulletin No. 471. Albany, NY.
Van Cleef, J. S. 1885. How to restore our trout streams. Transactions of the American Fisheries Society 14:50–55.
Van Cleve, K., F. S. Chapin III, C. T. Dyrness, and L. A. Viereck. 1991. Element cycling in taiga forests: state-factor control. BioScience 41:78–88.
Vannote, R. L., G. W. Minshall, K. W. Cummins, J. R. Sedell, and C. E. Cushing. 1980. The river continuum concept. Canadian Journal of Fisheries and Aquatic Sciences 37:130–137.
Vervier, P. 1990. Hydrochemical characterization of the water dynamics of a karst system. Journal of Hydrology 121:103–117.
Vervier, P., M. Dobson, and G. Pinay. 1993. Role of interaction zone between surface and ground waters in DOC transport and processing: considerations for river restoration. Freshwater Biology 29:275–284.
Vervier, P., J. Gibert, P. Marmonier, and M.-L. Dole-Olivier. 1992. A perspective on the permeability of the surface freshwater–groundwater ecotone. Journal of the North American Benthological Society 11:93–102.
Vogt, R. C. 1981. Natural history of amphibians and reptiles in Wisconsin. Milwaukee, WI: The Milwaukee Public Museum. 205 pp.
Voss, E. G. 1972. Michigan flora, Part I, gymnosperms and monocots. Cranbrook Institute of Science Bulletin 55 and University of Michigan Herbarium.
Voss, E. G. 1996. Michigan Flora, Part III, Dicots (Pyrolaceae-Compositae). Cranbrook Institute of Science Bulletin 61 and University of Michigan Herbarium.
Wake, D. B. 1991. Declining amphibian populations. Science 253.
Walker, L. R., J. C. Zasada, and F. S. Chapin, III. 1986. The role of life history processes in primary succession on an Alaskan floodplain. Ecology 67:1243–1253.
Wang, L., Lyons, J., Kanehl, P, Gatti, R. 1997. Influences of watershed land use on habitat quality and biotic integrity in Wisconsin streams. Fisheries 22:6–12.
Ward, J. R., K. Tockner, and F. Schiemer. 1999. Biodiversity of floodplain river ecosystems: ecotones and connectivity. Regulated Rivers: Research and Management 15:125–139.
Ward, J. V. 1998. A running water perspective of ecotones, boundaries and connectivity. Verh. Internat. Verein. Limnol. 26:1165–1168.
Ward, J. V., K. Tockner, P. J. Edwards, J. Kollmann, A. M. Gurnell, G. E. Petts, G. Bretschko, and B. Rossaro. 2000. Potential role of island dynamics in river ecosystems. Verh. Internat. Verein. Limnol. 27.
Ware, G. H., and W. T. Penfound. 1949. The vegetation of the lower levels of the floodplain of the south Canadian River in Central Oklahoma. Ecology 30:478–484.
Warren, P. L., and C. R. Schwalbe. 1985. Herpetofauna in riparian habitats along the Colorado River in Grand Canyon. Pp. 347–354 In: Riparian ecosystems and their management: reconciling conflicting uses. USDA Forest Service Gen. Tech. Bull. RM-120.
Wells, J. R., and P. W. Thompson. 1974. Vegetation and flora of Keweenaw County, Michigan. The Michigan Botanist 13:107–151.
Wesche, T. A., C. M. Goertler, and C. B. Frye. 1987. Contribution of riparian vegetation to trout cover in small streams. North American Journal of Fisheries Management 7:151–153.
Wetzel, R. G. 1999. Plants and water in and adjacent to lakes. In: Eco-hydrology: plants and water in terrestrial and aquatic environments. A. J. Baird and R. L. Wilby (eds.). London: Routledge.
Wetzel, R. G., and M. Søndergaard. 1998. Role of submersed macrophytes for the microbial community and dynamics of dissolved organic carbon in aquatic ecosystems. Pp. 133–148 In: Role of submersed macrophytes in structuring the biological community and biogeochemical dynamics in shallow lakes. Dordrecht, Netherlands: Kluwer Publishers.
Wetzel, R. G. 2001. Limnology: lake and river ecosystems. San Diego, CA: Academic Press.
Wharton, C. H., Kitchens, W. M., and Sipe, T. W. 1982. The ecology of bottomland hardwood swamps of the southeast: a community profile. FWS/OBS-81/37. U.S. Fish and Wildlife Service. 133 pp.
Whitmore, R. C. 1975. Habitat ordination of passerine birds of the Virgin River Valley, southwestern Utah. Wils. Bull. 87:65–74.
Whittaker, R. H., and W. A. Niering. 1965. Vegetation of the Santa Catalina Mountains, Arizona: a gradient analysis of the south slope. Ecology 46:429–452.
Williams, K. K. C. Ewel, R. P. Stumpf, F. E. Putz, and T. W. Workman. 1999. Sea-level rise and coastal forest retreat on the west coast of Florida, USA. Ecology 80:(6)2045–2063.
Williamson, K. J., D. A. Bella, R. L. Beschta, G. Grant, P. C. Klingeman, H. W. Li, and P. O. Nelson. 1995. Gravel disturbance impacts on salmon habitat and stream health, Volume 1: Summary report. Corvallis, OR: Oregon Water Resources Research Institute, Oregon State University. 52 pp.
Wilson, L. G. 1967. Sediment removal from flood water by grass filtration. Transactions of the American Society of Civil Engineers 10:35–37.
Wilson, T. 2000. Vernal use of riparian habitats by neotropical migrants. M.S. Thesis, Central Michigan University, Mt. Pleasant, MI.
Wilzbach, M. A. 1985. Relative roles of food abundance and cover in determining the habitat distribution of stream-dwelling cutthroat trout (Salmo clarki). Can. J. Fish. Aquat. Sci. 42:1668–1672.
Wilzbach, M. A., and J. D. Hall. 1985. Prey availability and foraging behavior of cutthroat trout in an open and forested section of stream. Verh. Int. Ver. Limnol. 22:2516–2522.
Winter, T. C., J. W. Harvey, O. L. Franke, and W. M. Alley. 1998. Ground water and surface water: a single resource. USGS Circular 1139. Denver, CO: USGS.
Winter, T. C. 2001. The concept of hydrologic landscapes. Journal of the American Water Resources Association 37(2):335–349.
Wistendahl, W. A. 1958. The flood plain of the Raritan River, New Jersey. Ecological Monographs 28:129–153.
Wolfe, C. B., Jr., and J. D. Pittillo. 1977. Some ecological factors influencing the distribution of Betula nigra L. in western North Carolina. Castanea 42:18–32.
Wolock, D. M. 2001. USGS. Personal Communication. Reston, VA.
Wroblickly et al. 1998. Seasonal variation in surface-subsurface water exchange and lateral hyporheic area of two stream-aquifer systems. Water Resources Research 34(3):317–328.
Young, J. R. 1994. The influence of sexual selection on phenotypic and genetic divergence of Sage Grouse. Ph.D. diss. Purdue Univ., West Lafayette, Indiana.
Young, J. R., Braun, C. E., Oyler-McCance, S. J., Hupp, J. W., and T. W. Quinn. 2000. A new species of sage-grouse from southwestern Colorado. Wils. Bull. 112:445–453.
Zimmerman, R. C. 1969. Plant ecology of an arid basin: Tres Alamos-Redington area, southeastern Arizona. U.S. Geological Survey Professional Paper 485-D. 47 pp.































































































