
3
Present Conditions: Vegetation
VEGETATION OF YELLOWSTONE’S northern range, a mosaic of different forest and nonforest communities, is the result of interactions among many environmental factors. To understand how management decisions may affect vegetation of the northern range, the committee reviewed the present conditions of the major vegetation types. In this chapter, the current state of a type of vegetation—for example, sagebrush or aspen communities—is described, followed by a discussion of how the driving variables may affect these conditions. In several cases, where changes in driving variables may alter ecosystem components, the nature of these changes and their consequences is discussed. Recent modification of environmental drivers is discussed to help explain the significance of recent changes of the ecosystem components. Although this chapter emphasizes the dominant plants, such as sagebrush or aspen, the concern over loss or degradation of these systems is not only for the dominant species, but also for the biodiversity—plants, animals, fungi, and microbes— that they support.
UPLAND SHRUBLANDS AND GRASSLANDS OF THE NORTHERN RANGE
Shrublands
Big sagebrush-Idaho fescue is the most abundant sagebrush-grassland type. It occurs on sites with thin cobble soils to well-developed loams, gener-
ally at elevations of 1,800 to 2,400 m within the 40- to 75-cm precipitation zone. It is distributed throughout the park but is most common in the Gardner and Lamar River drainages (Despain 1990). The habitat type is dominated by mountain big sagebrush (Artemisia tridentata ssp. vaseyana), although Wyoming big sage (A. tridentata ssp. wyomingensis) may also be present. Identification of big sagebrush subspecies is particularly important because of differences in palatability and preference to ungulates. Idaho fescue (Festuca idahoensis) dominates the understory with Agropyron spicatum and Koeleria macrantha also present. Forbs (broad-leaved herbaceous plants), such as Geum triflorum, are abundant.
Primary production (the amount of carbon fixed by photosynthesis) varies widely in big sagebrush-Idaho fescue habitat depending on rainfall and temperature (Mueggler and Stewart 1980). A 50% difference in production may occur on any given site over a 3-year period. Production varied across the type from 560 kg/ha (Mueggler and Stewart 1980) to 1,610 kg/ha with grasses contributing 21% to 42% of the production, forbs 38% to 56%, and shrubs 10% to 41%. Between 88% and 98% of the shrub production is from big sagebrush.
Big sagebrush-Idaho fescue habitat, which is heavily grazed in winter by ungulates, and the grassland habitat type (Idaho fescue-bearded wheatgrass) account for slightly more than half of all the nonforested vegetation in the park and on the northern range (Houston 1982). These two types probably furnish most of the forage for the large number of grazing animals in the park (Despain 1990).
Wyoming big sagebrush-bluebunch wheatgrass (A. spicatum, now Pseudoroegneria spicatum) habitat type occurs in the Gardner River canyon in small areas on southern and western slopes, often between big sagebrush-Idaho fescue and other grasslands on ridgetops and upper slopes. It occurs on shallow to moderately deep soils formed over several parent materials.
Mountain big sagebrush is the dominant shrub, although basin big sagebrush (Artemisia tridentata ssp. tridentata) may occur on deeper soils in drainages. Low shrubs, such as A. frigida and Gutierrezia sarothrae, are usually present. In addition to bluebunch wheatgrass, other conspicuous grasses include K. macrantha, Poa secunda, and Stipa comata. Production varies between 670 and 1,120 kg/ha with high variability between sites but not between years (Mueggler and Stewart 1980). This type is heavily grazed in winter by ungulates in the Gardiner area. Big sagebrush receives enough browsing to reduce the size of its canopies.
In the Gardiner area, Wambolt and Sherwood (1999) also describe a Wyoming big sagebrush-bluebunch wheatgrass (A. spicatum) habitat type, as did Mueggler and Stewart (1980) and Houston (1982). Associated species include sprouting shrubs like rubber rabbitbrush (Chrysothamnus nauseosus), green rabbitbrush (Chrysothamnus viscidiflorus), and gray horsebrush (Tetradymia canescens). Prairie junegrass (K. macrantha) and Sandberg bluegrass (P. secunda) are also common.
Grasslands
Idaho fescue-bearded wheatgrass (Agropyron caninum) habitat type is a highly productive mesic grassland with high species diversity. It occurs on gentle slopes at elevations of 2,000 to 2,600 m, within the 46- to 76-cm rainfall zone. It is dominated by grasses but contains a higher proportion of forbs (30% to 70%) than other western Montana habitat types. It has a short growing season and is used by native ungulates in winter (Houston 1982).
Idaho fescue-Richardson’s needlegrass (Stipa richardsonii) habitat type generally occurs at elevations of 1,100 to 2,100 m on gentle slopes and deep soils. It is a moderately mesic and productive grassland type dominated by Festuca idahoensis, S. richardsonii, Danthonia intermedia, Stipa occidentalis, and G. viscosissimum. This grassland is summer range for sheep or cattle, and it receives substantial winter grazing by native ungulates.
Idaho fescue-bluebunch wheatgrass is the most common xeric (Houston 1982) or moderately mesic (35 to 50 cm of precipitation) grassland type in the Greater Yellowstone Ecosystem (GYE) (Mueggler and Stewart 1980). It is found on intermediate mountain slopes at elevations of 1,400 to 2,300 m and occurs on a wide variety of parent materials.
Other grasses include K. macrantha, P. sanbergii, and either S. comata or S. occidentalis. Forbs cover from 10% to 60% of the area and include Achillea millefolium, Antennaria rosea, Arenaria congesta, and possibly Phlox hoodii. Medium shrubs such as A. tridentata and C. viscidiflorus are occasionally present. Annual primary production is highly variable depending on the weather. The grasses are used by elk and deer at the lower elevations for winter range and by pronghorn year-round. At the highest elevation, this type is summer range for elk and deer. At middle elevations, it is used as spring and fall range by all ungulates and as winter range by bighorn sheep and mountain goats.
Bluebunch wheatgrass-Sandberg bluegrass, or A. spicatum-P. sanbergii, is usually found between 900 and 1,800m, especially on gravelly soils on steep southern slopes. It is a moderately arid type in the 35- to 50-cm precipitation zone. Shrub and forb cover is low, and rhizomatous grasses are generally absent.
Needle-and-thread-blue grama (S. comata-Bouteloua gracilis) habitat type is usually found on broad alluvial benches and valley floors. Houston found this grass type in Yellowstone National Park (YNP) in the boundary line area, upstream on the Yellowstone River to the Black Canyon. It generally occurs below 1,500m and is the driest grassland habitat type (20 to 35 cm precipitation). The type is floristically simple, containing grasses and a low cover of forbs and shrubs. Needle-and-thread grass is a bunchgrass that dominates late seral stages of the community but decreases under heavy grazing pressure. Blue grama, the other dominant sod-forming grass in the community, increases under heavy grazing.
The terms decreaser, increaser, and invader refer to a plant’s response to grazing (Dyksterhuis 1949). Decreaser plants are most preferred by grazing animals and with continued heavy grazing are the first kinds of plants to decline in cover in a community. Increaser plants initially increase in cover in a community under ungulate grazing pressure because the preferred decreaser plants are declining and opening up space for increasers to grow. Eventually, as heavy grazing pressure continues, the increaser plants also decline, opening up sites for invader species of low palatability and generally low nutritional value. Common shrubs that increase with overgrazing include A. frigida, G. sarothrae, and Opuntia polycantha.
Factors Influencing Present Conditions of Sagebrush and Grasslands
Sagebrush: Ungulate Use
Big sagebrush is a particularly important food plant for several Yellowstone ungulates, especially in winter. Consequently, lower sites where there is little snow or where snow does not deeply cover shrubs are heavily grazed. During other seasons use is less, although big sagebrush may be an important source of protein for elk during the gestation period and in summer because grasses alone do not meet their protein needs (Wambolt et al. 1997).
Not all ungulates in the northern range use big sagebrush to the same
extent and not all subspecies of big sagebrush are equally used by the ungulates that feed on it. Big sagebrush is an important component of elk, mule deer, and pronghorn diets but not of bighorn sheep, bison, and mountain goats (Houston 1982). Mule deer and elk strongly prefer mountain big sagebrush to Wyoming big sagebrush and basin big sagebrush (Wambolt 1996). Wyoming and basin big sagebrush are much preferred over black sagebrush (A. nova), but during severe winters, all subspecies of sagebrush are browsed.
Ungulate browsing in low-elevation sagebrush sites near the park boundary has resulted in significant negative effects on big sagebrush (Wambolt 1996). In some cases, up to 91% of the leaders were removed and unbrowsed plants had higher productivity (45 g per plant) and seed-head production (60.3 seed heads per plant) than browsed plants (10 g per plant and 0.08 seed heads per plant) (Hoffman and Wambolt 1996). Up to 35% of plants were killed between 1982 and 1992, and many plants that survived had high percentages of dead crown (Wambolt 1996). Wambolt’s exclosure work (1998) demonstrates elk-induced decreases of sagebrush even in areas where there were no other ungulates. Wambolt and Sherwood (1999) came to similar conclusions. However, according to the National Park Service (YNP 1997), on 97% of the northern Yellowstone winter range sagebrush is stable or increasing and only 3% of the land shows sagebrush decline. In general, less browsing damage is observed at higher elevations (Singer and Renkin 1995, YNP 1997). Thus, elk appear to affect sagebrush at lower elevations—including the 3% of the winter range described by NPS as having declining sagebrush—but not at higher elevations.
Grasslands: Ungulate Use
Grasses are important components of the diet of most YNP ungulates except pronghorns and moose (Singer and Norland 1994). The importance of grasses in the diets of most ungulates on the northern range of YNP was shown by Singer and Norland (1994) through microhistological analyses on feces, comparison with earlier published work based on rumen analyses, and, for bighorn sheep, examination of feeding sites. Mean percentage diet compositions assessed by these methods were, as follows: elk, 75% to 79%; bison, 53% to 54%; mule deer, 19% to 32%, pronghorn, 10% to 4%; and bighorn sheep, 65% to 58%. Bison also included a high proportion of sedges in their diets (32% to 56%).
As in many other grasslands, ungulates in YNP move nonrandomly over
the landscape, feeding preferentially on grasses that are at particular stages of development (Frank et al. 1998). Yellowstone elk migrate elevationally as grasses produce new growth in spring (Houston 1982, Frank and McNaughton 1992). Some areas are intensively grazed but they recover as animals move to other patches. Timing of feeding is critical because feeding on vegetative material can have less impact than removal of growing points or reproductive structures. Despain (1996) compared one exclosure with the surrounding area. He found that elk fed heavily on the highly palatable bluebunch wheatgrass but moved off before the grass flowered so that there was little difference between exclosures and the surrounding areas in the total amount of green biomass of all species at the end of the growing season.
The grazing and migration pattern in the northern winter range results in modest spring and summer grazing on the lower ranges that receive heavy winter pressure and more intense grazing at higher elevations as snow recedes and green-up occurs (Singer and Harter 1996).
There are visually apparent effects of grazing on YNP grasslands. The question is whether those effects are signs of damage induced by feeding populations that exceed the carrying capacity of those rangelands. NPS perspective at YNP (YNP 1997) is that there is a perceptional problem—observers who see YNP rangelands make comparisons with commercial livestock rangeland and interpret the differences to indicate overgrazing (Coughenour and Singer 1991, 1996a).
Grasses are generally adapted to grazing and may even respond positively to appropriate grazing levels (Huff and Varley 1999). Grazing in YNP may cause enhanced plant protein (Singer 1996) and nitrogen content (Coughenour 1991, Mack and Singer 1993), and grazed plants may produce taller leaf and seed stalks (Singer and Harter 1996). Also, it is possible that animal movements, deposition of urine and feces, and the physical effects of hooves combined with plant responses to grazing could result in dense, short grass stands of enhanced above-ground growth (Frank and McNaughton 1992). Plant diversity on grazed sites is often higher than on ungrazed or heavily grazed sites (Wallace et al. 1995). Some authors have suggested that there are alarming decreases in plant diversity on Yellowstone’s northern range, implying that overgrazing is occurring on some sites (Wagner et al. 1995).
Grazing in the northern range does not appear to reduce root biomass (Coughenour 1991) or soil moisture content, even though there is an increase in soil bulk density (Lane and Montagne 1996). Changes have been reported in forb biomass (Singer 1995) and soil nutrients (Lane and Montagne 1996).
Confounding Factors: Fire, Pocket Gophers, and Nonnative Plants
Three other factors affect shrublands and grasslands in the northern range, but they have received little study.
Usually, fire is not common in grass or big sagebrush communities in the northern range (Despain 1990), but unusual increases in sagebrush since the 1870s (Houston 1982) may have caused sufficient fuel buildup to carry fires in this fire-sensitive vegetation type. After fire, numerous seedlings may establish, although there is little evidence that these plants survive and reproduce, especially for mountain big sagebrush (Wambolt et al. 1999). Possibly, the lush, young greenery draws ungulates to the site, increasing herbivory and further decreasing sagebrush population recruitment (Wambolt et al. 1999). Fire has fewer negative effects on grasslands and may stimulate community renewal.
Pocket gophers (Thomomys talpoides) are common, active, fossorial rodents that move masses of soil wherever there is sufficient below-ground consumable biomass to support them. Their digging activities, and those of bears, create patches in YNP grasslands with high densities of forbs that replace grasses and add to community diversity (Despain 1990). Pocket gophers can alter the structure of a community significantly and change the time-course of succession (Chase et al. 1982). Activities of elk and other ungulates might affect Thomomys populations by altering vegetation cover and soil compaction and consequently indirectly influence vegetation characteristics. If this had occurred, it would complicate the assessment of ungulates’ effects on vegetation, but we are not aware of relevant data for the northern range.
Nonnative plants are abundant in Yellowstone’s northern range, especially in big sagebrush and grassland habitats. Three grass species, timothy (Phleum pratense), crested wheatgrass (Agropyron cristatum), and cheat grass (Bromus tectorum), occur in the area and might alter, to an unknown degree, ecosystem productivity, susceptibility to fires, and ecosystem nutrient dynamics as well as other integrated measures of ecosystem processes (Huff and Varley 1999).
Conclusions
Not enough data are available for the committee to evaluate the claim that abiotic factors in the northern range are currently more influential than biotic factors on vegetation (YNP 1997, Frank et al. 1998), or the reverse. Certainly, many processes, especially those in the soil, are strongly mediated by animals
in YNP (Frank and Groffman 1998). The available data indicate that over short periods (decades), browsing and, in some areas, grazing have caused declines in plant populations and productivity.
Sagebrush
Big sagebrush at higher elevations in the northern range appears to be at relatively high abundance and vigor. These areas are important for ungulates during the nonwinter portions of the year. Because snow is usually deep and plants are relatively protected from elk browsing, these areas do not show sagebrush decline and may even show increases. They are not obviously of immediate management concern.
Lower-elevation big sagebrush stands, which have been very heavily used by elk, are decreasing in density and productivity, especially near the northern park boundary. Those sites are a critical winter range for a variety of other ungulates, especially pronghorn. It appears that, without extensive and intensive management to offset the damage done by elk browsing and grazing, the sites will continue to be degraded as resources for pronghorns and other ungulates (Wambolt and Sherwood 1999), especially near the northern boundary of the park.
Grasslands
Grasslands do not appear to have been altered as much by grazing as low-elevation shrublands have been by browsing. However, the few comprehensive reviews of the literature do not factor in the amount of biomass or other integrated measures of ecosystem characteristics contributed by nonnative species. The few studies available do not indicate that biodiversity is declining or that these systems are near a threshold value for some characteristic that is critical to any ecosystem process that currently appears to be within normal, long-term variations of the system. However, the committee would have more confidence if there were more data and analyses available.
FOREST TYPES ASSOCIATED WITH THE NORTHERN RANGE
YNP forests range from lower elevation woodlands through dense forest to timberline woodlands of whitebark pine and spruce and fir krummholz.
Knight (1994) describes seven forest and woodland types in Wyoming, six of which are found in YNP. The seven include limber-pine woodland, ponderosapine forest, Douglas-fir forest, aspen forest, lodgepole-pine forest, spruce-fir forest, and whitebark-pine woodland. There are many associations within these forest types, based on the composition of dominant and subordinate understory plants. Despain (1990) uses these associations to describe many forest habitat types. Of the seven types described by Knight, only ponderosapine forests are not found in YNP, and limber-pine woodlands are not common. The lack of ponderosa pine is considered to be caused by the predominance of rhyolitic soils on the Yellowstone Plateau, soils that create water stress conditions too extreme for ponderosa pine in the elevational zone where ponderosa pine might have established. The area may also lack the higher summer precipitation and warmer and longer growing season temperatures required by ponderosa pine. Of the six YNP forest or woodland types described by Knight (1994), aspen forests are discussed separately in this chapter because of the importance of their growth and reproductive response to changing environmental conditions in the northern range.
The forests of northern YNP and adjacent areas exist in a mosaic of forests and meadows (or parks). The causes for this mosaic mostly relate to moisture availability, whether influenced by soils, topography, or other factors (Patten 1963). The forests of YNP are continuously changing as stands mature and external factors, such as long-term climatic changes (Whitlock 1993), cause decline or loss of existing stands. Additionally, because of natural or anthropogenic environmental changes, such as climate change or fire suppression, many nonforest areas have been invaded by trees (Patten 1969, Jakubas and Romme 1993). Historical photograph comparisons show that many slopes throughout the northern range have more conifer forests now than in the past (Meagher and Houston 1998). These photographic comparisons also show a decline of aspen stands throughout the area, a phenomenon also seen in remotely sensed data that show aspen changes inside and outside northern YNP (Ripple and Larsen 2000a). Invasion by conifers of sagebrush and other nonforested areas continues to occur throughout the GYE. Sagebrush areas are becoming forest (Patten 1969), and forests are invading subalpine meadows (Jakubas and Romme 1993).
The most common forest type in the lower elevations of the northern range is dominated by Douglas fir (Pseudotsuga menziesii). It occurs below the lodgepole pine zone (1,800 to 2,300 m) in dense stands on cooler sites and sparse stands often mixed with Rocky Mountain juniper on drier or warmer sites. Douglas fir develops a thick bark, which makes mature trees relatively
fire tolerant. In the Yellowstone and Lamar River valleys (a major portion of the northern range), Douglas fir is the most common tree, with snowberry a common understory shrub in warmer sites and pine grass common in cooler sites. Aspen and lodgepole pineare often associated with Douglas fir in these areas. Douglas-fir forest with shiny-leaf spirea and other short woody shrubs as understory is common in the northern range on upper slopes and ridges (Despain 1990).
The extensive zone above Douglas-fir forest in the northern range is dominated by lodgepole pine. Although Despain (1990) described a few pure lodgepole-pine habitat types in YNP, none were in the northern range. However, using a cover classification, he described many cover types based on different successional stages of lodgepole forests with different understory recovery phases of climax species (e.g., subalpine fir). Knight (1994), however, described the lodgepole-pine forest as the most common forest type in Wyoming, occurring in northern Wyoming from 1,800 to 3,200m. He pointed out that, although lodgepole pine is primarily a fire successional species, climax lodgepole pine can occur on cool, nutrient-poor sites where other Rocky Mountain conifers cannot survive.
The forest zone above the Douglas-fir zone includes other subalpine species such as subalpine fir, Engelmann spruce, and, at higher elevations, whitebark-pine woodlands. Spruce, fir, and sometimes whitebarkpine form stunted krummholz “forest” stands on the ecotone between forest and alpine communities. In most cases, the krummholz is on exposed ridges or rocky outcrops. The elevation of these woody communities is well above the northern range especially the northern winter range.
Factors Influencing Present Conditions of Northern Range Forests
The present condition of forests on the northern range has been determined primarily by changes in management of fire and ungulates
Fire
Fire management policy has changed over the past several decades in YNP as well as in surrounding national forest wilderness areas. Before the 1970s, all fires were extinguished regardless of their location or intensity.
During the 1970s, the benefits of fire were recognized, and fires that were unlikely to damage human activities or structures were left to burn. This was very successful as most fires in the park during the decades leading up to 1988 burned a few to several hundreds of hectares in Yellowstone’s conifer forests, and the forest mosaic normally formed by disturbance processes was gradually returning to a more “natural” landscape. Fires did not occur in the sagebrush-grasslands during this time. (The fires of 1988 would have occurred even if decades of controlled burns had preceded them.)
Forests of the northern range are a mosaic of burned and unburned stands, most of which burned in 1988 (Despain et al. 1989). Before 1988 the forests were pure lodgepole-pine or mixed lodgepole-Douglas-fir and lodgepole-spruce-fir forests (Keigley 1997a). Most of the burned forests are recovering as nearly pure stands of young lodgepole pine.
Ungulate Use
Many forest stands in the northern winter range and in the upper Gallatin River drainage are heavily browsed and highlined (i.e., a browsing pattern on trees caused by ungulates removing foliage and live twigs as high as they can reach, thus creating a high line usually a few meters above the ground) (Kay 1990). Stunted conifers may not be browsed during mild winters and thus grow into branched trees (see following paragraph for analysis of tree architecture). All conifer species are browsed. Spruce, fir, and Douglas-fir trees are highlined, and adventitious branches, which grow on tree stems, are also browsed. In the upper Gallatin River drainage near the YNP boundary, trees are highlined throughout most of the area, but highlining decreases or disappears several miles south of the park boundary at higher elevations or several miles north of the boundary, where ungulates migrate but few overwinter (committee observations). The forest-ungulate interaction found on the Gallatin may represent a microcosm of the northern winter range. This interaction demonstrates the influence of ungulates on the forest under winter conditions where ungulates now stay at higher elevations in areas that once were primarily summer range (Patten 1963).
Branch architecture and growth form have been used for determining the intensity of browsing of shrubs and trees (Keigley 1997a). Young trees branch after the terminal shoots are browsed and then may grow into branched rather than single-stem tall trees if browsing pressure is reduced. Dating of the origin of branching and other tree-architectural anomalies have been tied to periods
of elevated ungulate numbers, while periods of “release” of terminal branches to form tall trees appear to correspond with periods when ungulate numbers were low (Keigley 1998).
Weather and Hydrology
Changes in forest patterns may be caused by altered hydrological conditions. Certainly, some years in the decade of the 1980s had warmer, drier weather than the decades preceding it, and apparently than the decade succeeding it. Consequently, when temperatures were warmer and winds were greater than normal, small fires became big fires and the extensive fires of 1988 occurred. Such cyclical changes in climate cause short, sporadic changes in many variables that affect forest ecosystems and therefore may temporarily establish external conditions for change. These cycles are typical; thus, disturbances and changes in forests of the northern range as influenced by hydrological fluctuations are normal. How hydrological cycles influence other factors such as ungulate behavior may be an important compounding effect in determining causes for alterations of the forests and individual trees within the forests and woodlands of the northern range.
Diseases and Infestations
Disease and infestations of insects and parasites are major factors that modify the forests of the YNP and the surrounding area. Several infestations of native species such as spruce budworm and pine-bark beetle have killed many hectares of forest, sometimes producing barren slopes, whereas at other locations only individual trees die. Reduction of tree health by these infestations, especially pine-bark beetle on lodgepole pine, makes trees susceptible to other pathogenic epiphytes such as dwarf mistletoe.
Recently, invasion of nonnative white-pine blister rust into the Rocky Mountains has caused mortality in whitebark pine and limber pine (Kendall and Schirokauer 1997, Kendall and Asebrook 1998). Proximity of Ribes spp., the secondary host for the fungus, as well as appropriate environmental conditions are necessary for blister rust to invade pine stands. In areas where blister rust infection is very high, such as Glacier National Park, conditions are moist; in YNP, drier conditions tend to retard the spread of the rust (Kendall 1998,
Kendall and Keane 2001). Elevation also plays a role in amount of infection, as whitebark-pine stands at higher elevations, and perhaps greater distances from Ribes communities, show less infection and mortality than lower-elevation stands. White-pine blister rust infection continues to expand in the GYE; if whitebark pine is lost in that area, their seeds, a major food source for grizzly bears, will also be lost (Kendall 1983, Mattson et al. 2001).
Conclusions
Forests associated with the northern range form a mosaic with nonforested areas. Over the past century, probably because of fire suppression, many non-forested areas have been invaded by trees, converting many hectares of the northern range into savanna-type forests or closed-canopy forest stands. These forests have been browsed by ungulates to a limited extent, mostly where ungulates are short of winter food. This use does not appear to significantly affect forest advance or the present distribution of the forests in the northern range.
ASPEN COMMUNITIES OF THE NORTHERN RANGE
Aspen (Populus tremuloides) is the most widely distributed native North American tree species (Fowells 1965) and an important component of landscapes in the intermountain west. Aspen is the only native upland deciduous tree that occurs in YNP. Aspen stands support high numbers and diversity of breeding birds (DeByle and Winokur 1985) and provide habitat for other wildlife. They are an important source of forage for browsing ungulates, especially during winter (Olmstead 1979). Aspen stands also are considered prime areas for livestock grazing and can be extensive enough to provide a quality watershed and attractive scenery (DeByle and Winokur 1985). However, many aspen stands throughout the west, including YNP, appear to have declined during the twentieth century as old trees died and little recruitment took place. The causes and consequences of this decline have received considerable discussion (Krebill 1972, Loope and Gruell 1973, Schier 1975, Hinds and Wengert 1977, Olmstead 1979, Bartos and Mueggler 1981, Hinds 1985, Boyce 1989, Kay 1990, Bartos et al. 1994, Romme et al. 1995, Baker et al. 1997).
Like other species of the genus Populus (poplars), aspen are single-
trunked, deciduous trees that spread clonally by means of root-borne sucker shoots (Eckenwalder 1996). They are among the fastest-growing temperate trees, and their shoots continue to grow after bud-burst by initiating, expanding, and maturing leaves throughout the growing season. Aspen is dioecious, having separate male and female trees that flower before leaf emergence in the spring. The seeds are born on catkins and are wind dispersed, often over long distances. Seeds remain viable for only a few weeks after their release in late spring (Moss 1938). Aspen usually regenerates via vegetative suckering from the residual root system after a disturbance that kills the mature trees. These adventitious shoots grow rapidly and are supported by the parental root system for growth for at least the first 25 years (Zahner and DeByle 1965). The new suckers develop new roots within the first few years, but the parent root system remains alive and functioning for 40 to 50 years (Pregitzer and Friend 1996). Individual stems are relatively short lived. For example, most stems in the Colorado Front Range are less than 75 years old. Few stems reach 200 years of age. The wood of poplars lacks terpenoids and other compounds that resist decay, so the centers of large mature trees often are much rotted before they fall (Eckenwalder 1996).
Aspen has a broader range of environmental tolerances than most of its associated species and can grow in most mountain vegetation zones (Daubenmire 1943, Fowells 1965). In the intermountain region, however, aspen is confined to sites with moist soils with at least 38 cm of annual precipitation and cold winters with deep snows (Jones and DeByle 1985). Subsurface moisture from seeps, or other factors that concentrate water, characterize aspen sites. In western Wyoming, the modal elevation of aspen stands is about 2,000m. The upper elevation limits may be determined by growing season length and the lower limits by evapotranspirational demands (Mueggler 1988). Aspen in the northern range of YNP occurs in small- to medium-sized stands growing on moist areas of the landscape. However, aspen stands in YNP are not as robust as stands in the mid-intermountain regions of Utah and Colorado (Mueggler 1988). In the Rocky Mountains, aspen is primarily a clonal species that reproduces almost exclusively by root sprouting and produces large stands composed of stems from one or a few genetic individuals (Barnes 1966, McDonough 1985, Tuskan et al. 1996). The individual clones respond differently to environmental conditions across the landscape. This is obvious in the fall when stands change color at different times and to different shades; differences may also be evident in responses to moisture stress and other environmental factors.
Reestablishment of aspen by seed is believed to have occurred infrequently in the Rocky Mountains since the last glaciation because climatic conditions have not been suitable for widespread germination and seedling establishment (Einspahr and Winston 1977, Cook 1983). However, rare episodes of seedling recruitment have occurred (Jelinski and Cheliak 1992), among them the widespread establishment of aspen seedlings in YNP after the 1988 fires (Kay 1993, Romme et al. 1997, Stevens et al. 1999). It is difficult to age aspen clones, as they are long-lived and may be much older than the oldest live canopy tree (Grant 1993). Existing large trees in an aspen clone represent “recruitment events” for tree-sized stems—that is, periods when conditions were suitable for seedlings or root sprouts to develop into tall trees.
Aspen occurs both as successional and as climax vegetation. Aspen root sprouts readily after disturbances such as fires that kill the overstory trees (Mueggler 1988, Romme et al. 2001). Fire enhances this recruitment, but any event, including aging, senescence, and death (e.g., by girdling) of the overstory trees, that reduces the apical dominance that typically suppresses root sprout development and growth may trigger extensive root sprouting. The extensive aspen stands throughout the central Rockies are believed to be partly a result of wildfires, and some investigators have suggested that the elimination of fire reduced regeneration of aspen (Loope and Gruell 1973, Houston 1982). Root sprouts tend to grow faster than seedlings, and thus aspen sprouts can easily outcompete other species, such as conifers, that must regenerate through seeds (Despain 1990). Aspen is not shade-tolerant, so if conifers start to grow under aspen stands, they may eventually shade out the aspen. In other locations, aspen can remain as the climax vegetation, continuing to regenerate through root sprouts when the individuals that compose the canopy become senescent (Despain 1990).
Aspen, like most Populus species, is preferentially browsed by ungulates when in leaf and during winter (Olmstead 1979). Elk eat the tips of aspen sprouts and the bark of mature trees, except where the smooth white bark has been replaced by thick, black, corky bark in response to prior injury. Aspen root sprouts and seedlings may be severely browsed by elk during winter or during spring and fall migrations between the summer and winter ranges. In many parts of YNP and the GYE, elk commonly browse nearly all root sprouts in aspen stands (Romme et al. 1995, Ripple and Larsen 2000a). Several investigators have suggested that excessive elk browsing is the major reason for the lack of regeneration of aspen stands (Krebill 1972; Beetle 1974, 1979; Kay 1990; Bartos et al. 1994). Indeed, ungulate browsing can increase mortality
of aspen saplings and suckers reduce or eliminate root suckers (which prevents regeneration of large stems), and contribute to increased disease in larger trees (DeByle and Winokur 1985, Hinds 1985).
Aspen stands are scattered throughout the northern range where moisture conditions are suitable. Historical photographs suggest that aspen thickets occurred on the northern range during the early twentieth century (Houston 1982, Kay 1990, Meagher and Houston 1998). Kay (1990) and Keigley and Wagner (1998) show photographs of aspen stands in the 1800s with no browse line, whereas Houston (1982) describes a browse line for this period. In the late 1800s, there were approximately 6,000 ha of aspen on the northern winter range, whereas today aspen cover only about 2,000 ha, and many of them are in a shrub form (Renkin and Despain 1996). Because much of the northern winter range is sagebrush-grassland or wet bottomlands, locations where aspen can develop are limited. This scarcity of sites is common in semi-arid environments such as the lower elevations of the northern winter range. In the forested areas of the northern range, aspen represents a localized vegetation type, possibly relicts of past fire regimes (Houston 1982).
Except for aspen clones in exclosures, most aspen stands on the northern range contain aging or senescent trees with a relatively dense understory of root sprouts mixed with a herbaceous ground cover. About 85% of the large aspen alive today on the northern range originated before 1920 (Romme et al. 1995, Ripple and Larsen 2000b). Romme et al. (1995) obtained increment cores from 15 aspen stands, which revealed a period of aspen regeneration in the 1870s and 1880s. Ripple and Larsen (2000b) developed a more comprehensive age structure for northern range aspen by obtaining 98 readable increment cores from 57 different stands. Results of this study revealed that 85% of the sampled aspen stems originated between 1871 and 1920, with 10% originating before 1871 and only 5% from 1921 to 1998. The relative paucity of older aspen (Romme et al. 1995, Kay 1997, Ripple and Larsen 2000b) is consistent with the relatively short lifespan of individual aspen stems. It is clear that very few of the stems became established in the twentieth century. However, quantifying the establishment of tree-sized aspen earlier in the nineteenth century is difficult because many trees have already died and some older trees are not sound enough for aging.
A historical data set in which the diameter at breast height (DBH) was recorded for aspen trees in 20 separate riparian stands in 1921 and 1922 (Warren 1926) has provided a valuable insight on northern-range aspen in the eighteenth and nineteenth centuries. Ripple and Larsen (2000b) used these data to infer historical age distribution. New increment cores were collected in
1998 from 30 northern range aspen and 19 aspen from the Gallatin National Forest to determine the relationship between aspen age and DBH. The resulting regression was then applied to the largest diameter aspen in each of Warren’s (1926) stands. This analysis revealed that the aspen present in 1921 and 1922 had originated between 1750 and 1920.
Northern range aspen have produced root sprouts during recent decades, but root spouts have not been able to grow higher than the herbaceous layer. Sprouts are regularly browsed back by ungulates to the approximate depth of winter snowpack (Romme et al. 1995). Browsing intensity is high. For example, between 50% and 70% of aspen root sprouts in both burned and unburned stands on the northern range were browsed in 1990 and 1991 (Romme et al. 1995), and Ripple and Larsen (2000b) found that 89% of aspen stems in the northern range showed evidence of browsing in 1997. As the large, older aspen stems die, they are not replaced by new recruitment into the canopy, and shrubby aspen, grasses, and forbs may come to dominate (Kay 1990). However, root sprouts that have been protected from browsing within an exclosure have grown several meters, which suggests that climatic conditions are suitable for such growth. Removal of the exclosure exposes them to stem girdling by ungulates. These taller individuals then die, returning the clone to aged trees and herb-height root sprouts.
Multiaged or multiheight aspen clones are scarce in Yellowstone, especially in the northern parts of the park. In contrast, multiheight aspen clones exist at mid-elevations in areas within the GYE but outside YNP (e.g., Centennial Valley and Grand Teton National Park). Some areas of transitional range outside YNP may receive less browsing because migrating elk may not be present long enough or densities of ungulates in these areas may be lower than in YNP. Studies from Yellowstone and elsewhere in the Rocky Mountains provide insight into the regional variation in the factors that explain these differences, including fire, elk browsing, and other environmental drivers (e.g., DeByle and Winokur 1985, Romme et al. 1995, Baker et al. 1997, Kay 1997, White et al. 1998).
Factors Influencing Present Conditions of Aspen Communities
Precipitation
Rain and snow, by recharging subsurface soil moisture and protecting woody plants, affect the availability of winter forage and hence migration of
ungulates. Together or separately, these may influence ungulate utilization of aspen root sprouts, shrub-aspen, and aspen bark. For example, in some reports on the northern winter range, long-term sprout height appeared to be a function of snow depth (Kittams 1952, Barmore 1980). Annual precipitation has fluctuated over the past century but without drying or wetting trends. During this time there has been no reduction in stream flows in general in YNP or in the Lamar River, as measured at a gage within the northern winter range. However, the 1930s and 1980s were drier decades than others in this century. Data from the 1980s have been used to infer a drying trend (YNP northern range report), but if there was a trend, it ended in the 1990s. Snow-water equivalent depth in April for two snow courses in the northern range shows a decline in the 1980s, a period that also was below normal for annual peak flows. The 1990s, however, had peak flows above normal, and heavy snowpacks in 1996 and 1997 produced back-to-back 100-year floods on the Yellowstone River.
The conclusions that can be drawn from the hydrological information are that there have been periods of above or below normal precipitation and snow cover during the past century, but there is no consistent pattern of declining precipitation or snow cover in the northern range. Consequently, snow through its influence on elk migration or protection of aspens, cannot be the one factor that produced changes in aspen stand conditions on the northern range. Precipitation as a source of subsurface moisture also has probably not changed sufficiently to affect aspen condition. The fact that aspen clones continue to produce prolific root sprouts and maintain shrub-aspen stands indicates that there probably is little moisture stress for these communities, although seedlings may experience greater moisture stress.
Balling et al. (1992b) examined the climate record for YNP (1890 to 1990) with regard to the relationship between climate and wildfire. Although they did not focus on snowpack, their analyses demonstrated a significant increase in summer temperatures during the century-long record along with a decline in January to June precipitation levels. Taken together, these changes suggest an increase in summer drought conditions in YNP during the past century. The period before the historical record coincides with the end of the Little Ice Age, an extended period of cooler global climate conditions, whereas the period of the Balling et al. (1992a) study ended before the wetter 1990s. Although these differences are not sufficient to explain the decline of aspen stands in YNP during the twentieth century, the climatic changes suggest the possibility of increased moisture stress as a contributing factor, although the twentieth century fluctuations in annual indices of drought stress fall within
normal fluctuations over the past several centuries. However, in Yellowstone and throughout the Rocky Mountains, climatic change does not appear to be closely associated with aspen regeneration. In Rocky Mountain National Park, Baker et al. (1997) found little evidence for a relationship between aspen regeneration and climate, and aspen have not regenerated in Yellowstone during the climatically favorable conditions that have occurred since the 1920s (Ripple and Larsen 2000b). More important, their dendrochronological data show that aspen regeneration occurred during both favorable and unfavorable climatic conditions between 1871 and 1920 (Ripple and Larsen 2000b). Moreover, many aspen clones in the GYE where there is little winter use by ungulates show normal recruitment and expansion of clonal boundaries (committee member observations).
Fire
Fire is a major factor that enhances aspen recruitment (Loope and Gruell 1973, Brown and DeByle 1987). Typically, fire removes competing overstory conifers, triggering profuse aspen root sprouting and potentially producing extensive clonal stands. Reduction in fire frequency consequently may reduce the number and extent of aspen clones. Clones that do not have extensive successional understory development of conifers may continue to exist for many “generations” through root sprouting as the older trees senesce.
On the northern range, fire frequency has been reduced since the 1800s through fire suppression in YNP and adjacent national forests. Fire recurred every 20 to 25 years in northern YNP before the initiation of fire control in the late 1800s; from 1900 to 1988, there were almost no fires in Yellowstone’s northern range (Houston 1973). Consequently, some of the aspen clones are becoming mixed aspen-conifer stands (Mueggler 1988). Others, however, remain as relatively pure aspen clones, especially in locations either some distance from conifer seed dispersal or on locations that may not be suitable for conifer recruitment. Examples of both pure aspen clones, some contained within exclosures, and aspen-conifer successional stands are found on the northern range. In both situations, there is often extensive aspen clonal root sprouting if the conifer canopy is not too dense. Thus, a reduction in fire frequency may reduce the expansion of existing aspen clones and also may prevent development of new clones through sexual recruitment. Fire suppression could be one factor contributing to the lack of recent recruitment of tree-sized stems. However, Hessl (2000) found no significant difference in aspen-sucker
density in response to fire in the Gros Ventre watershed south of Yellowstone, and, even after the 1988 fires, aspen shoots have not grown enough to escape being eaten by elk in northern Yellowstone.
The 1988 Yellowstone fires burned approximately 22% of the northern range. Mature aspen stands that were burned in 1988 produced hundreds of thousands of root sprouts per hectare, but the 1988 fires resulted in a reduction in the number of aspen within shrub-aspen communities on the northern range (Kay and Wagner 1996). The density of shrub-aspen before fire (1986) was about 18,000 stems per ha, and shortly after the fire (1989) it was about 19,000 stems per ha, but by 1992 the number had dropped to about 10,000 stems per hectare. Extensive seeding of aspen in burned areas also occurred after the 1988 fires (Kay 1993, Romme et al. 1997). These seedling aspen are genetically diverse but are not elongating rapidly in YNP (Stevens et al. 1999). Browsing intensity on seedling aspen remains high, and even unbrowsed individuals attained maximum heights of only about 1.5m by 1999 (M.G.Turner, personal observation). Thus, despite extensive root sprouting and seedling establishment after the 1988 fires, regeneration of large aspen is not yet occurring.
The accumulation of postfire coarse woody debris may, however, favor aspen regeneration. As conifers killed by fire fall, extremely dense piles of coarse woody debris may accumulate and provide refugia from ungulate browsing. After the 1988 fires, seedling aspen growing in areas with a lot of fallen conifers had not been browsed by ungulates since 1993 (Turner and Romme 1994). In burned areas on the northern range, Ripple and Larsen (2001) found that aspen suckers protected by fallen conifer barriers were, on average, twice the height of unprotected suckers.
Ungulate Use
Dendrochronological studies in the GYE and elsewhere in the Rocky Mountain region suggest a strong relationship between elk density and regeneration of aspen cohorts—stands containing a predominant size class or distinct patches of different size classes that are spatially segregated within a clone (Baker et al. 1997). In Rocky Mountain National Park, Baker et al. (1997) found only a weak correspondence between regeneration of aspen cohorts since 1875 and climatic and hydrologic fluctuations; aspen regenerated in both cool and warm periods and during periods with less than average precipitation. However, aspen regeneration seemed to be related to fluctuations in elk den-
sity, with cohorts regenerating when elk density was probably low and exhibiting no regeneration when elk density was high (Baker et al. 1997). Notably, aspen regeneration appeared to have been sporadic when elk density was lower than today (e.g., late 1800s to early 1900s), although older cohorts may have already senesced. The complete absence of recruitment of aspen trees since the late 1970s is anomalous in their record. Based on mortality levels of suckers and established trees, sucker density, and the reduced height and extensive branching of browsed suckers, Baker et al. (1997) concluded that the aspen population in the elk winter range in Rocky Mountain National Park is declining, largely in response to elk browsing. In the Gros Ventre watershed, located south of Yellowstone and managed by the Bridger-Teton National Forest, Hessl (2000) examined aspen regeneration since 1850 in relation to fire, elk browsing, climatic variation, and land management regimes. Regeneration of aspen cohorts in the Gros Ventre has been episodic and largely corresponds to periods when elk density was lower (e.g., late 1800s and 1940s and 1950s) or browsing pressure on native vegetation was reduced (1970s) because of artificial winter feeding in the National Elk Refuge located near Jackson, Wyoming. Current patterns of aspen regeneration appear to be related to human-induced gradients of elk use, with browsing intensity highest near the winter feeding grounds and declining with distance from the feeding grounds.
Ripple and Larsen (2000a) compared the percentage of aspen stems browsed in stands located in the northern range and in two locations in the nearby Gallatin National Forest: the Sunlight Basin, close to the northern range, and Clarks Fork Basin, which does not receive heavy ungulate use in the winter. Their initial data revealed that the highest browsing intensity occurred on the northern range (89.6% of 1,100 observed ramets were browsed), but browsing intensity was also high in the Sunlight (85.6% of 611 ramets) and Clarks Fork (76.7% of 322 ramets) Basins. However, the Clarks Fork site contained a more even distribution of aspen in a wide range of size classes compared with the northern range, in which all aspen stands contained individuals of #DXGT#20 cm DBH and none of #DXLT#11 cm DBH. In the Sunlight Basin, 90% of the stands contained aspen of #DXGT#11 cm DBH, but 11.5% of the stands contained individuals of 1 to 5 cm DBH, and 27.8% of the stands had individuals of 6 to 10 cm DBH. Ripple and Larsen are also using aerial photos to examine changes through time in aspen stands in northern Yellowstone and the Gallatin and Shoshone National Forests, but the results of these analyses are not available.
There is indisputable evidence that ungulates, primarily elk, are browsing
aspen intensively on the northern range and likely have done so throughout the twentieth century. Ungulates are a major factor contributing to the current absence of recruitment of tree-sized aspen in YNP (Kay 1993, Romme et al. 1995, Ripple and Larsen 2000b) and in other locations in the Rocky Mountains (Baker et al. 1997, Hessl 2000). What is not known, however, is how ungulate numbers and patterns of habitat use before park establishment might have influenced the spatial and temporal dynamics of herbivory on aspen. This perspective is important because an understanding of the dynamic interaction between elk and aspen would help us interpret the current condition.
There is controversy about the numbers, or even the presence, of ungulates in the northern range in the pre-Columbian period. Evidence supports the presence of ungulates (primarily elk) in the northern range when YNP was established (Schullery and Whittlesey 1992). However, available evidence does not suggest that extensive herds of ungulates wintered in what is now the northern winter range in the period immediately before establishment of YNP (see Appendix A). Certainly, photographic evidence presented by Kay (1990) and Keigley and Wagner (1998) suggests that there was little woody plant utilization by ungulates in the mid- to late 1800s. This could mean that (1) no or few ungulates were there any time of year (Kay’s position), or (2) that no or few ungulates used the northern range when forage was scarce and woody plants were one of the few food sources available because herds had migrated to lower elevations, conditions expected during severe winters (Keigley and Wagner’s position).
It is unlikely that woody plants, including aspen stands with little highlining and extensive tall willow communities, could have existed if large numbers of ungulates used the northern range in winter in the late 1800s. Data collected by Barmore (1965, presented by Renkin and Despain 1996) when the elk herd was reduced in the late 1950s and early 1960s show that levels of elk use as low as 10 elk-use days per acre resulted in over 75% utilization of aspen sprout leaders, and elk-use days no greater than 25 per acre resulted in 100% utilization. Consequently, there appears to be a change in the ungulate use pattern of the northern range over the past century and a half, with more animals staying in the park’s northern range in the winter. This change may be due to several factors, none of which has been thoroughly tested. These include increased human activities and development within the original winter range and winter migration routes as well as development of a regular hunting season that created hunting pressures on the YNP northern border. Cole (1969) used these factors to account for altered migration patterns and forage use of elk in southern YNP-Grand Teton National Park elk herds.
Chemical composition, especially secondary compounds, may influence herbivory and the survival of aspen. Some authors have suggested that willows and aspen in YNP have inadequate secondary compounds to defend against elk browsing (e.g., Singer et al. 1994), but the chemical composition of aspen and its relationship to browsing have not been studied in Yellowstone. The dominant secondary metabolites of aspen are phenolic products of the shikimic acid pathway. These include condensed tannins, phenolic glycosides (the salicylates salicin, salicortin, tremuloidin, and tremulacin), and coniferyl benzoate (Palo 1984, Lindroth and Hwang 1996). Tannins and phenolic glycosides occur in leaf, stem, and root tissues, whereas coniferyl benzoate occurs only in flower buds. The influence of tannins and phenolic glycosides on feeding by insect herbivores, particularly Lepidoptera, has received considerable investigation. Tannins are ineffective defenses, whereas phenolic glycosides are effective at moderate to high concentrations against many aspen-feeding insects (Bryant et al. 1987, Hemming andLindroth 1995, Lindroth and Hwang 1996, Hwang and Lindroth 1998). Coniferyl benzoate provides protection against herbivorous birds such as grouse (Jakubas et al. 1989). However, little is known about the influence of aspen secondary metabolites on the foraging behavior of mammals, particularly ungulates. Phenolic glycosides and their derivatives, but not tannins, deter feeding by hares (Tahvanainen et al. 1985, Reichardt et al. 1990). Aspen tannins may be similarly ineffective against browsing ungulates, a prediction consistent with the production of tannin-binding salivary proteins in such animals (Austin et al. 1989, Hagerman and Robbins 1993). Jelinski and Fisher (1991) found significant variation in secondary compounds among aspen clones but concluded that these concentrations did not appear to inhibit palatability or digestibility for members of the deer family. Erwin et al. (2001) concluded that for aspen seedlings and clones in Yellowstone, foliar phenolic glycosides and tannins were not active defenses induced in response to browsing by large mammals.
Several exclosures in the northern range and the Gallatin range (northwestern YNP) demonstrate that, with protection from ungulates, aspen can still grow tall and root sprouts can produce multiple age classes of above-ground stems. Intensive ungulate browsing on aspen may result from factors that concentrate the animals near the aspen clones, and/or from the ungulate herds being sufficiently large that nearly all aspen is utilized. If ungulates are “prevented” from migrating to portions of their historical winter range, the density of ungulates on the remaining available winter range will be unnaturally high. Hunting and human developments (e.g., throughout Paradise Valley, north of YNP) can prevent long-distance migration. Truncation of migration routes may
also hasten the spring return of ungulates to the summer range at times when herbaceous plant biomass is still low and woody forage must be consumed.
Interrelated Changes
Aspen communities no longer produce root sprouts that grow into tall trees in the northern range. Root sprouting continues, which indicates adequate local abiotic conditions for this form of recruitment and suggests that other factors must be contributing to prevent the development of tall aspen clones. Fire suppression has prevented reduction of competition and enhancement of conditions for tall growth. However, this cannot explain the lack of regeneration, particularly given that the 1988 fires resulted in profuse root sprouting that was intensively browsed. Ungulate browsing clearly contributes to the lack of regeneration, although the long-term dynamics of aspen in northern YNP are not well understood. Romme et al. (1995) hypothesized that during the late 1800s, when they thought the last recruitment of tree-sized aspen occurred, might have been characterized by several co-occurring factors, including intensive market hunting for elk, more frequent fires, generally moist growing conditions, and reduction in beaver populations, which all contributed to reduced browsing pressure. However, recent work by Ripple and Larsen (2000b) indicates that aspen regeneration did occur as recently as 1921. In interpreting these patterns, Ripple and Larsen (2000b) hypothesized that the disruption of natural predator-prey relationships may have contributed to the observed differences in aspen regeneration. Furthermore, they suggest that the reestablishment of wolves in 1995 may benefit aspen in the long term. In addition to reducing elk population size, wolves may also influence ungulate movement and browsing patterns. In Jasper National Park, White et al. (1998) reported that a new cohort of aspen sprouts regenerated into trees ranging from 3 to 5 m tall after wolves were reintroduced. Top-down control on ungulate herbivory has also been suggested in Isle Royale National Park, a system with no long-range ungulate migration (McLaren and Peterson 1994).
Postfire coarse woody debris (i.e., fallen trees) also can protect aspen from ungulate browsing and has contributed to greater elongation of aspen suckers in burned stands on the northern range (Ripple and Larsen 2001). Ripple and Larsen (2001) have also suggested that park managers might create experimental “jackstraw piles” of dead conifers to create barriers to browsing and hence facilitate aspen regeneration.
Conclusion
All data sources indicate that the abundance of large aspen in northern Yellowstone has declined during the twentieth century, and most indicate that this decline is due primarily to ungulate browsing. Although climate and fire may influence aspen dynamics, these factors have not been the main drivers for the changes in aspen observed during the recent century. Rather, the ungulate population, both in size and behavior, appears to be most strongly correlated with aspen regeneration in the GYE. Large aspen trees are likely to regenerate on the northern range under current conditions only if they are protected from ungulates, either through physical barriers (which may develop naturally with postfire coarse woody debris or which may be provided artificially) or through behavioral changes, as might be induced by the presence of predators such as wolves, unless much more extensive lower-elevation winter range becomes available.
RIPARIAN COMMUNITIES OF THE NORTHERN RANGE
Riparian ecosystems are the transition from stream to upland. They occupy a very small part of the landscape, often less than 1%, yet they play an important role in stream dynamics, wildlife ecology, and biodiversity of the region (Naiman et al. 1993, Naiman and Decamps 1997, Patten 1998). In most cases riparian ecosystems occur on alluvial sediment deposits where river and alluvial groundwater supplements water available from precipitation (Gregory et al. 1991). Riparian ecosystems may also be found near springs and seeps where the groundwater surfaces to create wet areas or surface flows. Riparian ecosystems are frequently disturbed by periodic flooding and thus are in a continual process of succession (Malanson 1993). They also exhibit a high level of resiliency after termination or removal of disturbances or stressors—for example, after floods, after the return of normal stream flows, or after removal of grazing (Stromberg et al. 1997; Kaufmann et al. 2000; Patten 1998, 2000).
Most western North American riparian vegetation communities are a result of recruitment and survival in response to seasonal hydrological events, variation in groundwater depth, and flood-generated gravel bars. For example, most western cottonwood species recruit along streams on bare, moist surfaces during the decline of spring high flows (Friedman et al. 1995, Scott et al.
1997, Stromberg et al. 1997). Survival of these trees depends on maintenance of a high water table in the floodplain and avoidance of scouring by floods and ice flows. Mortality, or inability to survive after recruitment may result if the water table drops below that tolerated by young or maturing plants (Rood and Mahoney 1995).
Willows are a common woody component of many western riparian communities. Although their recruitment may also coincide with spring floods, availability of damp soils may be sufficient for them to establish and survive. Factors that help maintain an elevated water table enhance willow community growth and expansion. For example, beaver activity may elevate the water table and create suitable sites for willow community expansion, whereas stream incising may lower the water table and cause stress to willows.
In the Rockies, valley geomorphology directly influences the extent and type of riparian communities (Patten 1998). Streams that flow through broad valleys with low gradients may be lined by both woody and herbaceous riparian vegetation. If the water table is shallow, wetland herbaceous plants (e.g., sedges and wetland grasses) may extend for some distance from the river. These areas often are devoid of woody species because the herbaceous plants may prevent establishment of willows or cottonwoods. Willows and sometimes cottonwoods may occur near the stream, where floods enhance their recruitment. Once established, these species may spread asexually and expand within the floodplain, often occurring away from the stream as it migrates across the floodplain.
Riparian communities in the northern range have developed in response to many of the conditions discussed above. Both low-gradient and relatively steep valleys exist in the northern range. In the lower reaches of the northern range, the Yellowstone and Gardner Rivers cut through canyons, providing only narrow areas for cottonwood stands and shrub-willow communities. Twenty-four species of willow are found in the northern range, and species composition changes, in part, with elevation as well as geomorphic setting (YNP 1997). For example, false mountain willow (Salix pseudomonticola) is more common at lower elevations; Drummond, Farr, and Barclay willows (S. drummondiana, S. farrii, S. barcleyi) are more common at higher elevations; wolf willow (S. wolfii) is common on broad floodplains; and sandbar willow (S. exigua) is common on sandy, exposed-stream meander lobes. Other woody species, such as shrubby cinquefoil (Potentilla fruticosa) and water birch (Betula glandulosa), are also found in the riparian zone (Singer 1996).
Reaches of the Lamar River and Soda Butte and Cache Creeks at mid-elevation in the northern range extend across low-gradient floodplains where willow and wetland herbaceous communities are common. Narrow leaf cottonwood (Populus augustifolia) occurs in limited stands near the channel on present and past river meander lobes (point bars) (Keigley 1997b). At higher elevations these rivers flow through both narrow- and steep-gradient valleys and shallow low-gradient floodplains. Cottonwoods drop out and willow and herbaceous communities dominate on low-gradient reaches. Conifers become co-dominants along the streams as the range changes from winter/ summer to only summer ungulate range (the upper elevations of the northern range). Throughout the northern range are several seeps and springs that also support deep-rooted communities, primarily willow. These are not truly riparian communities because they do not occur along the edge of streams or lakes, but the shallow water table that creates the spring or seep also creates suitable habitat for riparian and wetland species. Also, in some areas aspen is a riparian species. Warren (1926) found aspen along streams and ponds, especially where there was extensive beaver activity, but most of those stands are gone. Aspen can still be found in riparian conditions but in limited situations, especially along smaller streams. Most aspen stands in the northern range are in moist upland areas.
Riparian communities in the northern range exist, or have existed, because conditions were suitable (1) for recruitment of the characteristic riparian species, (2) for maintenance of established species because the alluvial water table was sufficiently shallow to maintain plant growth and survival, and (3) because factors, such as loss of adequate groundwater availability, changes in surface hydrology, plant community modification through browsing and grazing activities, and other human modifications of the landscape exerted only moderate pressures. If any of these conditions—which helped create the riparian communities—change through natural or anthropogenic actions, the riparian community is stressed and may degrade or be lost from places on the northern range where they formerly occurred.
The riparian ecosystems along the northern borders of YNP and adjacent areas of ungulate winter use, including the northern range and the Gallatin River valley in the northwestern corner of the park, show evidence of degradation in response to stressors (Patten 1968, Singer et al. 1994, Keigley 1997b, YNP 1997). In these areas, woody riparian vegetation has many dead branches, often with new growth emanating from low on the plant. In the upper Gallatin River drainage there are no cottonwoods, and so evidence of
stress exists only in willows, whereas in the lower and mid-elevations of the northern range both willows and cottonwoods show signs of stress. In some cases, where factors that maintain riparian communities have changed substantially during the past 100 years or more, riparian communities are absent. In other situations, woody riparian vegetation appears to be stunted or maintained in a short-stature growth form. This is true of both cottonwoods and willows. Evidence of periodically successful recruitment of young cottonwoods and of the existence of cottonwood canopy layers representing several age classes or recruitment events is almost lacking along most of the streams in the northern range. Mature and overmature cottonwoods occur in several locations, and hedged cottonwoods with large stems, which clearly are old, are sometimes found in these mature cottonwood galleries (Keigley 1997b). Cottonwood seedlings are found on good recruitment sites some years, but these seedlings currently are not surviving to become mature trees. Although narrow-leaf cottonwood can root-sprout and this is occurring along the streams in the northern range, root sprouts also do not appear to be reaching maturity or growing tall enough to contribute to a mid-height or tall canopy within the cottonwood galleries.
On broad, low-gradient floodplains, extensive willow communities still exist, but they are generally overtopped by the associated herbaceous vegetation. In other places where willow occurs, such as at seeps or along steeper gradient streams, willows also appear to be prevented from growing to normal stature for the species. Willow species differ in their normal height growth. For example, wolf willow generally is of low stature (1 to 1.5 m), whereas Geyer, Booth, and Bebb willow generally attain heights of several meters. Many willow species can reproduce asexually, and spread along the streams where moisture conditions are favorable. Recruitment of shrubby willow species is not tied as closely to hydrological events as that of cottonwood. Seed dispersal may not depend strongly on spring floods, and recruitment may occur on any available moist substrate that is accessible to the seeds. Willows normally require and are more tolerant of wetter soils than cottonwood. Consequently, their establishment in near-wetland conditions close to streams, instead of on more elevated locations (0.5 to 1.5 m above baseflow) where cottonwoods grow, shows they could grow along a river or on any wet soil with persistent alluvial groundwater regardless of surface events such as spring floods. Willows are also less tolerant of drought conditions and lower water tables than cottonwoods (Stromberg et al. 1996).
Factors Influencing Present Conditions of Riparian Communities
Hydrology
Several hydrological factors influence riparian vegetation recruitment, growth, and maintenance. We discuss these factors separately, as changes in any one may significantly influence riparian communities.
Precipitation
Precipitation influences riparian ecosystems through its influence on runoff, stream flow, and groundwater recharge. Variability in precipitation for the past several decades falls within normal variation over the period of record, including expected periods of drought and above-normal precipitation. If precipitation were the cause of changes in riparian vegetation in the northern range, stream flow magnitudes should be outside the normal range. An analysis of peak and annual flows in the Yellowstone River near Corwin Springs shows that both peak flows and annual discharge volumes were generally below average in the earlier part of the twentieth century (Figures 3–1 and 3–2). Peak flows are analyzed in addition to annual discharge because they often closely represent snowpack conditions, the conditions that recharge and maintain elevated water tables in the watershed. Corwin Springs was used because it is the first U.S. Geological Survey (USGS) gage outside YNP on the Yellowstone River. The Yellowstone River drains most of the watershed of the northern range in addition to southern watersheds in Yellowstone, and data from the Corwin gage integrate most of the watershed output. Discharge patterns at Corwin Springs also correlate well with discharge patterns of the Lamar River, which primarily drains the northern range (Mowry 1998). Precipitation gages and snow measurements are spotty, and precipitation in mountainous terrain tends to be quite heterogeneous. Thus, an integrating measurement is more useful.
YNP used Yellowstone’s northern range data to argue that there was a decline in precipitation and stream flows from 1982 to 1994 caused by drying conditions that stressed the range, forest, and riparian ecosystems (YNP 1997). However, this is too short a period to demonstrate unusual variation. Peak flows and annual discharge averaged below normal during the 1980s, but
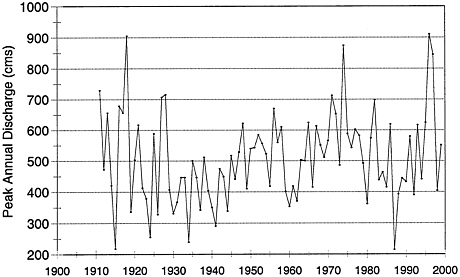
FIGURE 3–1 Annual peak stream flow in cubic meters per second (cms) at Corwin Springs on the Yellowstone River a few miles downstream from Gardiner, MT. USGS gage 06191500.
willows declined (i.e., reduced stature and loss) during the earlier decades of the twentieth century (Smith et al. 1915, Warren 1926). Therefore, it is necessary to use hydrological data from most of the century to detect whether hydrological changes have caused willow decline.
Increased precipitation at high elevations has been used to explain the vigor of higher-elevation willow communities (YNP1997); however, willows occur along stream courses where they have access to groundwater and soils wetted from the stream or from capillary rise of water from the water table (Dawson and Ehleringer 1991, Busch et al. 1992, Flanagan et al. 1992). Consequently, the amount of local precipitation has very little influence on willow growth and survival, except when it is in the form of a snow cover (see below).
Snow Accumulation
Annual snow accumulation during the twentieth century was within normal variability for the period of record based on spring runoff data (Farnes 1998).
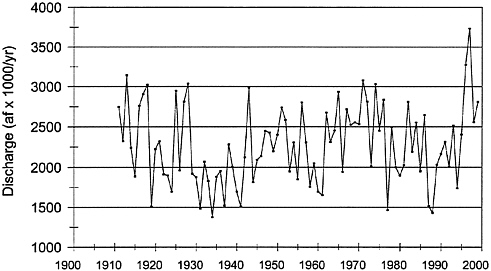
FIGURE 3–2 Annual discharge volumes (thousands of acre-feet per year) at Corwin Springs on the Yellowstone River a few miles downstream from Gardiner, MT. USGS gage 06191500.
There has been considerable variation in the past few years, with 1996 and 1997 having high accumulations, and 1998, 2000, and 2001 having accumulations well below the mean. Some evidence shows that runoff from snow melt peaked earlier, by three days on average, after the 1988 fires (Farnes 1998) and that peak flows may have been higher, but peak discharge data do not support the latter conclusion. Also, runoff appears to have returned to normal for the snowfall amounts within a decade after the fires. Snow accumulation may play a more important role in riparian vegetation structure through protection of riparian shrubs as pointed out by Singer (1996). Not only does deep snow accumulation around riparian woody plants reduce browsing use, it also may prevent use of higher-elevation riparian vegetation by ungulates. Conversely deep snow can cause greater use of riparian vegetation by browsers if herbaceous plants are buried and woody stems are the only forage exposed. This is especially true when a thaw-freeze cycle creates an impenetrable ice layer over herbaceous vegetation. The present condition of riparian vegetation in the northern range may be caused by heavy utilization when it is one of the few sources of forage in deep snow years or years with significant thaw-freeze events.
Stream Discharge (Hydrographs)
Streams often recharge adjacent groundwater, although in some cases groundwater movement may be toward the stream rather than away from it. Consequently, reduction in stream flows combined with reduced groundwater movement from uplands when the precipitation is below normal may reduce groundwater availability to riparian vegetation. However, the USGS daily historical hydrological data for rivers flowing from the northern range (e.g., Yellowstone at Corwin Springs) do not indicate any unusual reduction or increase in stream flows over the past century beyond normal variation. Consequently, surface hydrological changes do not appear to be sufficient to have directly affected riparian availability of groundwater, although if some streams incised their channels during the past several decades, that could have lowered the surface flow enough to lower the alluvial water table to levels that could stress riparian vegetation.
Spring floods can scour channel margins and, through overbank flows, scour and flood the adjacent floodplain to produce suitable sites for recruitment. High spring flows that could enhance recruitment of riparian plants may occur only every 5 or 10 years, resulting in spaced age-classes of the woody riparian species, which is especially evident in large woody species such as cottonwood. The lack of evidence of regular recruitment events in the northern range indicates that perhaps there have been fewer spring flood events during the past few decades. However, USGS peak flow data for rivers coming from the northern range watershed show periodic high or flood flows, sufficiently high to produce a recruitment event and probably high enough to have seedling establishment above either ice scour or smaller scouring flood events. Therefore, the lack of periodic recruitment by riparian plants in the northern range cannot be attributed to absence of flood events.
Groundwater (Alluvial Water Table): Streams, Seeps, and Beavers
Groundwater monitoring data obtained by the committee show that groundwater in the floodplains, where most of the riparian vegetation in the northern range occurs, has not changed sufficiently in the past several decades to cause stress on riparian vegetation. Data from one well north of Gardiner showed water table decline in the 1980s (YNP1997), but the 1980s were a dry decade and domestic use of groundwater in the area of the well probably increased as well. Groundwater monitoring wells within the northern range are needed if
seasonal and annual fluctuations in groundwater levels are to be determined.
The existence of riparian vegetation, especially willow, along most of the rivers and on the floodplains in the northern range indicates that the water table is still high enough to maintain these communities. The occurrence of seeps and springs is additional evidence that the water table is near enough to the surface to maintain riparian vegetation. Groundwater change appears to have been an important factor in those few areas where riparian vegetation once occurred but is no longer present because the beaver ponds were abandoned.
Stem xylem water potential can indicate water stress in woody plants. Measurements of xylem water potential of short (browsing suppressed) and tall stature willows near exclosures showed water potential averages of -1.17±0.68 and -2.4±0.85 bars, respectively. Comparisons between short and intermediate stature willows were -1.71±0.68 and -2.86±1.35 bars, respectively (Singer et al. 1994). Thus, tall and intermediate willows with greater canopies had greater water stress than short willows, but these values are much less than wilting-point values (i.e., -15 bars) for mesic plants. Loss of willows during the 1988 drought period may be partly explained by water stress (Singer et al. 1994) but may be better explained by possible water table declines and warm, dry winds that occurred during that period.
Beavers, whose activity currently helps maintain riparian vegetation along many streams in the GYE, are currently rare or absent in most of Yellowstone’s northern range. The reasons for beaver decline and the consequences for willows have recently been controversial (YNP 1997; Singer et al. 1998, 2000; Keigley 2000). An early theory attributed beaver decline to reductions due to ungulate browsing of woody plants (willow and aspen), which are beaver food and building material (Bailey 1930, Wright and Thompson 1935, Jonas 1955). In the absence of beavers, water tables in areas elevated by beaver dams may decline so that riparian vegetation cannot survive. Singer et al. (1998, 2000) argued that this process is exemplified by the healthy stands of willow at Willow Meadows (a location in the transition zone from winter to summer northern range use) that occur on a broad floodplain where the water table is less than a meter deep are maintained by beavers.
Geomorphology: Stream Banks and Channels
Many of the streams in the northern range on the broader floodplains now produce braided channels where formerly they produced single or multiple meandering channels (Meyer et al. 1995). The braided channels are dynamic
and have moved considerably over the past several decades (Mowry 1998). Changes in the width of Soda Butte Creek measured when it was full to the banks near the upper edge of the winter range were related to hydrological history and perhaps vegetation cover (Mowry 1998). Locations with tall willows and little winter ungulate use remained stable (i.e., no statistically significant changes) during a relatively constant hydrological period from 1954 to 1987, whereas channels with grass-covered banks and reaches with little willow bank stability narrowed over that time. That demonstrates, for these sections of Soda Butte Creek upstream of most wintering ungulates, that vigorous woody riparian vegetation tends to maintain bank stability under most hydrological conditions whereas non-woody banks tend to aggrade, a process explained by Lyons et al. (2000) from research on Great Plains rivers.
However, the floods of spring 1996 (one of two back-to-back hundred-year floods on the Yellowstone River and its tributaries) altered the willow-stabilized banks more than banks stabilized by grass or low willows. That happened because woody riparian species (e.g., willows) stabilize banks only to a flood-discharge threshold, beyond which shear stress scours and uproots the plants, so that banks collapse (e.g., Stromberg et al. 1997). Stream banks that are stabilized only by vegetation that offer low resistance to flowing water (e.g., grasses and low willows), may withstand high-velocity floods better than banks with woody plants. Also, if one high-flood year is followed by another, as was the case in 1996 and 1997 in Yellowstone, the instability created by the first year may allow the subsequent flood to alter the channel morphology more than it would if it had occurred alone. For example, the 1997 hundred-year flood on the main stem of the upper Yellowstone River downstream from YNP removed many hectares of mature cottonwood trees that had been made unstable by the 1996 flood (committee observation).
The sandy deposits in several of the rivers of the northern range, primarily the Lamar River, are thought by some observers to be a product of increased erosion along the river channels (R.Beschta, Oregon State University, comments to committee, 1999) and high sediment supply from steep erodible terrain and tributary streams (Rosgen 1993). Rosgen suggested that poor riparian conditions due to excess browsing cause unstable channel conditions and increased sediments in the Lamar River valley. In addition, Chadde and Kay (1991) claimed that the Lamar River valley channels have incised, perhaps as much as several meters, because of heavy ungulate use. If incision has occurred, the shallow riparian water table needed to sustain riparian communities and vigorous growth response to browsing will not be maintained. For example, willows growing along an apparent water table gradient away from the
Gallatin river have decreased growth and recovery to browsing with distance from the river (Patten 1968).
Floodplains with shallower alluvial deposits, or steeper low-elevation valleys, tend to have bedrock conditions nearer the surface that prevent decline of the water table even if stream flows are reduced. In such places, willow vigor is maintained even when browsing occurs.
Ungulate Use
Several studies have assessed the response of willows to protection from browsing (Kay 1990, 1994; Chadde and Kay 1991; Singer et al. 1994, 1998). Some of these studies have used measurements that show that frequency and occurrence of willows inside and outside exclosures do not differ significantly (YNP1997). These measurements count the number of individuals present, regardless of their size or vigor. Therefore they cannot detect changes in size, cover, or productivity—key vegetation components. Other studies have measured willow cover (Kay 1990, 1994; Singer et al. 1994; Singer 1996), which is a better metric for determining shrub vigor because it measures actual areal cover of the canopy. Singer (1996) showed that, during the three decades after construction of upland exclosures, total canopy areas for several species of willow inside the exclosure were 200% to 600% greater than those outside. Also, annual biomass production was 5 to 10 times greater inside the exclosure than outside (Kay 1994, Kay and Chadde 1992). During the period between measurements of willow canopy cover in the 1950s and 1960s and measurements in the 1980s, cover increased many fold inside and hardly changed outside exclosures (Kay 1994). Data from exclosures showed that only willows protected from browsing reproduced (Kay 1994). No seed-producing catkins were found outside, but they averaged over 300,000 per m2 inside. Ungulate browsing also removes pollen-producing catkins. Near Geode Creek, no catkins were found on willows below the browse height, but many were found above the browse height (Kay 1994). This reduction in reproduction of willows in the northern range may account for the reduction in willow pollen in recent lake sediments (Barnosky et al. 1988).
Although exclosure studies show that browsing has a dramatic effect on willows, exclosures represent the extreme of no browsing—rare in ecosystems that have browsers, as YNP does. Thus, the best way to determine how ungulate populations within and outside YNP cause changes in willow communities is to compare natural willow communities inside and outside YNP but still
within the northern range. Unfortunately, this type of study has only been done for aspen (Kay 1990), although several recent publications comparing early and recent park photos show that tall-willow communities have declined in the northern range (Kay 1990, YNP 1997, Keigley and Wagner 1998, Meagher and Houston 1998). Explanations for the differences between early and recent photos include excessive browsing by ungulates (Kay 1990, Keigley and Wagner 1998) and climate change, especially during the 1930s drought (YNP 1997, Meagher and Houston 1998). Groundwater conditions, which have not been measured in any of the studies of ungulate utilization of riparian shrubs, should be measured at all sites.
The lack of cottonwood recruitment may be due to ungulate browsing. When ungulate numbers were reduced in the northern range in the 1950s and 1960s, cottonwoods were temporarily “released,” put on new shoots, and grew taller than during the preceding or following decades (Keigley 1998).
Concentrations of defensive or secondary chemicals in riparian vegetation may influence the magnitude of ungulate use. Several studies were designed to determine whether low-stature willows have lower concentrations of defensive chemicals and thus are more palatable and more heavily used by herbivores than taller willows (Singer et al. 1994). Extremely short stature is not a natural condition for most willows. Consequently, very short shrubs must be a result of browsing. A controversy has developed about whether the level and composition of certain secondary chemicals is a result of short stature or a response to browsing. This controversy has resulted in a series of commentary papers following Singer et al. (1994) (Singer and Cates 1995, Wagner et al. 1995).
Studies on secondary chemicals in willows generally show that low-elevation, short willows tend not to produce the concentrations of defensive secondary chemicals (e.g., tannins) found in higher-elevation and tall willows. This supports the argument that the tall willows have greater defenses against ungulate browsing and thus remain tall because ungulates avoid them. However, many of the tall willow communities are at higher elevations where snow cover or other conditions prevent heavy winter ungulate use. Thus, tall willows occur in areas with few wintering elk. Also short willows might not produce as high a concentration of secondary chemicals as tall willows because growth after browsing, the only type of shoot these short willows produce, tends not to produce as many of these chemicals. One question that is not answered in the many studies dealing with secondary chemical defense is whether the concentrations of secondary chemicals found in tall-stature willows are sufficient to prevent browsing even by ungulates that are starving. Starving ungu-
lates will eat almost any woody plant that is available if other food such as grass and forbs are buried and unavailable. Browsed highlines on lodgepole pine, a low-quality food, are an example of this type of ungulate use. The evidence that lower levels of defensive chemicals increases utilization of willows or other riparian shrubs by browsers is insufficient to explain the decline in stature or loss of willows and riparian vegetation in the northern range during this century.
Conclusions
During the first six decades of the twentieth century the NPS was concerned that ungulates were reducing the riparian communities; however, since then, factors other than ungulate browsing, such as climate change, have been hypothesized to explain the loss or reduction in stature of the riparian communities of the northern range (Houston 1982, YNP 1997). The committee concludes that some riparian losses may be due to changed hydrological conditions in addition to responses of vegetation to ungulate use. Flooding events and water tables are still suitable for recruitment of the dominant riparian vegetation. However, channel incision might have lowered the water table, reducing the ability of willows to recover from browsing. The increase in ungulate browsing over the past century, as evidenced by hedged willows and lost willow stands throughout lower elevations of the northern range, has caused most of the reduced willow cover and lowered willow reproduction this area. Ungulate use also appears to be the primary factor preventing cottonwoods from recruiting, because seedlings do appear on good seedbed sites along the rivers of the northern range after appropriate hydrological events, but they fail to survive.
WETLAND VEGETATION OF THE NORTHERN RANGE
Wetlands are a prime source of biodiversity in YNP (Elliot and Hektner 2000). Several types of wetlands, such as natural depressions, beaver dam wetlands, thermal wetlands, and wetlands along rivers and creeks (i.e., riparian wetlands), are found on the northern range. Chadde et al. (1988) defined 62 wetland communities in the northern range. These included wetlands dominated by trees (e.g., spruce [Picea] and aspen), willows, shrubs (e.g., cinquefoil [Potentilla fruticosa] and silver sagebrush [Artemisia cana]), and gram-
inoids (e.g., grasses and sedges). Most wetlands in YNP have been mapped under the U.S. Fish and Wildlife Service’s National Wetlands Inventory mapping program. Although wetlands are important across YNP, they are hardly mentioned in the park’s documents on the northern range (YNP 1997) or in a vegetation description of YNP (Despain 1990).
There have been limited studies of wetlands in the northern range. Houston (1982) sampled five highly productive wetland meadow sites with humic soil types dominated by sedges. Relatively few species were sampled (1 to 13 species on the five sampled sites). The sites were dominated by sedges and rushes and graded to species characteristic of mesic grasslands.
One wetland type was identified as tufted hairgrass-sedge type, described by Mueggler and Stewart (1980). This habitat type dominated by Deschampsia caespitosa, is found on high-elevation valley bottoms between 2,000 and 3,000m (Mueggler and Stewart 1980). The soils are deep and poorly drained, with water standing on the soil surface at least part of the growing season. D. caespitosa is the dominant grass but sedges (Carex spp.) are always present. Other grasses include Danthonia intermedia, Phleum alpinum, and Agrostis and Juncus spp. Forb species present include Potentilla gracilis, Polygonum bistortoides, and Antennaria corymbosa. This is a highly productive habitat type with annual production reaching 2,900 kg/ha.
Other wetlands were on alkaline soils formerly dominated by alkali grass and meadows cut for hay, now dominated by introduced timothy grass. All wetland communities receive substantial winter grazing by elk and especially bison, a species that may consume large quantities of sedges.
Brichta (1987) studied 21 of the 62 northern-range wetland community types defined by Chadde et al. (1988). He also measured soil types, surface-soil saturation status, and groundwater depth and chemistry. Of the 180 plots studied, 24 mostly were aspen or willow stands in exclosures that excluded herbivores. Depth of soil organic horizon was highly variable, ranging from less than 5 cm for about half of the study plots to more than 60 cm for many sedge-dominated sites.
Community types with abundant sedges had saturated soils all summer. Types with aspen, as well as some graminoid communities, were saturated for part of the summer; community types with spruce, and cinquefoil, as well as other graminoid-dominated communities, were dry most of the summer.
Water tables were variable among study sites. For example, mean depth of water below soil surface was more than 100 cm for spruce wetlands around 100 cm for aspen wetlands, about 20 to 40 cm for willow wetlands and as low as 100 cm, from 40 to 100 cm for cinquefoil wetlands, and for graminoid wet-
lands from near surface to below 100 cm. Many water tables remained steady throughout the summer; others dropped from near surface to below 100 cm. Several sites with declining water tables were near glacial ponds. Water chemistry was also variable with most pHs near or slightly below neutral. Water chemistry differences did not appear to influence plant diversity, but availability of shallow water through the growing season did. Wetland community types typically changed across moisture gradients.
All of the aspen wetlands and some willow sites studied by Brichta (1987) were within exclosures. At the Junction Butte exclosure, a Salix geyeriana/Deschampsia cespitosa community was inside the exclosure, but a Potentilla fruticosa/Deschampsia cespitosa community was immediately outside it. Brichta concluded that “the influences of grazing and succession upon wetland community type distribution should be further studied. Water regimes and soils were similar for paired plots on either side of the exclosures, therefore differences in soils and water levels could not account for the marked difference in vegetation that existed inside and adjacent to exclosures.” Brichta’s study was during a dry period that eventually resulted in the 1988 fires. Perhaps water tables would have been higher and more sites would have had saturated soils throughout the summer had the study been done during wet years.
Only four species of amphibians are known with certainty to occur in YNP now: boreal toad (Bufo boreas), tiger salamander (Ambystoma tigrinum), boreal chorus frog (Pseudacris maculatd), and Columbia spotted frog (Rana luteiventris). Two other species have been reported but not verified in recent times. The population sizes of the boreal toad have declined significantly, those of the spotted frog less so, and the chorus frog and tiger salamander appear to vary within normal limits. Wetlands are critical habitat for amphibians, which are often considered indicator species of environmental health (e.g., EPA1998 Star Grant: Environmental Factors That Influence Amphibian Community Structure and Health as Indicators of Ecosystems). Habitat changes resulting from the interplay between vegetation, hydrology, elk, and beaver could influence available wetland habitat for amphibians, but none of the decreases is clearly related to the known direct or indirect effects of elk population size or feeding.
Conclusions
Hydrological changes in the northern range are the most likely cause of changes in wetland communities, although evidence from exclosures indicates
that hydrological factors do not account for the differences between plant communities inside and outside of exclosures. Lowering water tables from stream incision and loss of beaver ponds may reduce wetland habitat. There may be drying of wetland depressions but there are no long-term data on shallow groundwater levels in northern range locations where drying may be occurring to explain changes in these depressions. Some of the depressions may also be filling in, reducing the amount of area available for wetland species (committee observation, Yellowstone National Park northern range, June 1999). Wetlands in the northern range that support herbaceous vegetation may be grazed, but use of these areas probably is not as detrimental to their long-term sustainability as potential changes in groundwater availability. However, wetlands dominated by woody plants appear to be significantly degraded by browsing.








































