
4
Present Conditions: Animals
POPULATION DYNAMICS OF NORTHERN RANGE UNGULATES
YELLOWSTONE’S NORTHERN RANGE supports a rich community of native ungulates. Ungulates in the park were subject to market hunting until the 1880s, and the park’s wildlife was not seriously protected until the U.S. Army was assigned administration of the park in 1886 (YNP 1997). By this time, most ungulate populations had been greatly reduced. The Army (and later NPS) managed and protected the resident ungulates and diligently controlled predators, which resulted in greatly reduced populations of coyotes, bears, and mountain lions, and the extirpation of wolves from Yellowstone National Park (YNP). YNP adopted a policy of “natural regulation” in 1968, which led to increased populations of elk and bison. Throughout the twentieth century, management of ungulates has been controversial and great concern has been expressed by the public and park officials about the “correct” management objectives and the actions needed to achieve them.
Density Dependence and Natural Regulation
The concept of density-dependent regulation of population sizes has figured prominently in the controversy over management of elk and bison in YNP
(Houston 1982, YNP 1997). For a population to be regulated by density-dependent factors, some combination of the following processes must operate. As population density increases, mortality and emigration rates increase and the rate of reproduction decreases. Increases in mortality can result from depletion of food supplies because individuals find it increasingly difficult to obtain adequate nutrition. Diseases, whose transmission is facilitated by high population densities, and predators also can increase mortality (Sinclair 1989, Royama 1992, Begon et al. 1996). Rates of reproduction may decrease because females cannot obtain enough food to support high rates of pregnancy and because offspring may be born at lower weights and less appropriate times than when food supplies are good. These rates may change gradually with population density, or there may be thresholds at which major changes occur (Fowler 1987; McCullough 1990, 1992).
The combination of these processes tends to cause population densities to decline when they are high and to increase when they are low. However, this does not guarantee that population densities will stabilize or reach some equilibrium because changes in rainfall, snow accumulation, fires, and other abiotic events may cause large fluctuations in the capacity of the landscape to support the population (Soether et al. 1997). In other words, because the environmental conditions in the landscape may vary considerably, the magnitude of variation in the density of a population by itself cannot be used to assess the importance of density-dependent factors in regulating the size of a population. Most populations of larger herbivores are subject to a combination of stochastic and density-dependent processes that lead to large variation in rates of juvenile survival and subsequent changes in population growth rates (McCullough 1990, Sinclair and Arcese 1995, Soether 1997, Gaillard et al. 1998).
The conceptual basis of density-dependent population regulation is simple, and there are many examples of ungulate populations in which fecundity declines or mortality increases as population density increases. However, no single statistical method identifying density dependence has emerged, despite vigorous discussion (Strong 1986, Pollard et al 1987, Turchin 1990, Dennis and Taper 1994, Soether 1997, White and Bartmann 1997, Shenk et al. 1998, Bjornstad et al. 1999). Many unharvested ungulates are regulated, at some point, by density dependence (McCullough 1979, Sinclair 1979, Fowler 1981, Gaillard et al. 1998), but populations are always subject to a multitude of factors and it can be difficult to distinguish the effects of density from those of other influences. The best evidence for density dependence comes from direct measures of changes in population processes such as mortality, fecundity, and migration (Shenk et al. 1998).
Elk
The intense public debate since the 1920s over management of the northern range elk population has stimulated numerous studies of the Yellowstone elk population (Barmore 1980; Houston 1982; Chase 1986; Merrill and Boyce 1991; Coughenour and Singer 1996a; Singer et al. 1997, 1998). Nevertheless, reliable data on population size and distribution exist for only the past several decades, and population size estimates in published reports differ (e.g., compare Houston [1982] with Lemke et al. [1998]). Elk are highly mobile. Inmild winters, they are widely dispersed and make extensive use of forested habitats where they are difficult to count; aerial counts during harsh winters, when elk move to lower elevations, are therefore more reliable than counts during mild winters. Hunting also alters distribution and the number of animals in the population. Aerial surveys of elk initiated in 1956 marked the beginning of reasonably reliable estimates of elk in YNP (Houston 1982). Comparisons of population estimates are confounded by changes in survey technique, differences in the time of survey (e.g., before or after harvest), and vagaries of weather that influence animal movements and visibility.
Harvest and Movement
Historical records of northern range elk illustrate the dominant influence of intense management before 1968 and the effects of management and natural processes since 1968 (Figure 4–1). Public concerns about overgrazing of the northern range resulted in herd reduction by the park at rates that kept the elk population relatively low and stable from the 1920s until 1968 (Houston 1982). After 1967, when elk harvest stopped, the elk population increased (Figure 4–1). From 1968 through 1975, hunter harvests outside the park from the northern herd dropped from about 1,500 elk per year to fewer than 200 (Houston 1982).
The winter late hunt north of YNP resumed in 1967 amid concerns that disturbances due to hunting would inhibit movements of elk from YNP to the historical winter range north of park boundaries. Until the severe winter of 1988, relatively few elk were observed north of Dome Mountain (approximately 16 km north of YNP) (Lemke et al. 1998). The northern range elk population, which had expanded to about 20,000 animals, responded to heavy snows in the winter of 1988–1989 by moving to a lower-elevation winter range en masse. More than 3,000 elk were observed in the area of Dome Mountain,
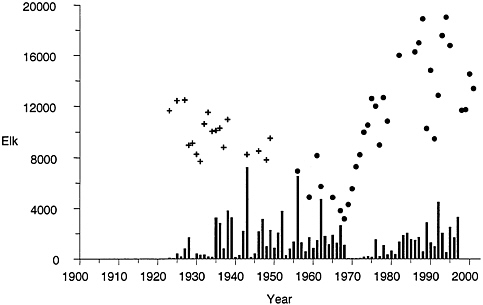
FIGURE 4–1 Elk population counts and harvest of northern range elk. Vertical bars show the number of elk harvested. Filled circles are counts from aerial surveys; crosses are ground counts. Sources: YNP 1997, Lemke et al. 1998, Lemke 1999.
a substantial proportion of the 7,000 to 8,000 elk that migrated north out of YNP that year (Figure 4–2) (Lemke et al. 1998). This event marked a major change, or restoration, in behavior of northern range elk. A significant proportion of the population has consistently migrated to the area of Dome Mountain in subsequent years. From 1975 to 1988, an average of about 200 elk per year wintered north of Dome Mountain; the average increased to about 2,800 per yearfrom 1989 to 2001 (Lemke et al. 1998; Lemke 1999; T.Lemke, Montana Fish, Wildlife and Parks, personal communication, 2001). From 1989 to 1999, an average of 5,600 elk (range, 1,533 to 8,626) wintered outside the northern border of the park, including the area of Dome Mountain (Lemke et al. 1998; Lemke, personal communication, 2001). The many elk wintering outside YNP boundaries are using an expanded winter range. Houston (1982) estimated that during the 1970s the elk winter range consisted of 109,000 ha (as measured by Lemke et al. [1998]), whereas current winter distribution typically includes around 153,000 ha of winter range, an increase of 41% (Lemke et al. 1998). Most of the increase in winter range is outside YNP, where the area utilized increased from 22,000 to 53,000 ha, including 9,200 ha north of Dome
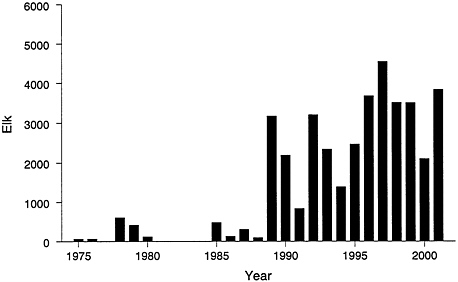
FIGURE 4–2 Number of elk counted north of Dome Mountain, Montana. Sources: Lemke et al. 1998, Lemke 1999.
Mountain. In response to the apparent need for a lower-elevation winter range, Montana Fish, Wildlife and Parks and the Rocky Mountain Elk Foundation collaboratively acquired 3,500 ha of key winter range on Dome Mountain (Lemke et al. 1998). Grazing by domestic livestock was discontinued on this land to provide an enhanced supply of winter forage for wildlife (Lemke et al. 1998).
Regulation of Elk Populations
The northern range elk herd is strongly influenced by both density-dependent and density-independent factors. Mortality of juveniles varies widely from year to year and is positively correlated with population density (Barmore 1980, Houston 1982, Merrill and Boyce 1991, Coughenour and Singer 1996a, Singer et al. 1997, Taper and Gogan 2002). Increased juvenile mortality rates at high elk densities are caused by grizzly bears, black bears, and coyotes (Singer et al. 1997), and, recently, wolves (Smith et al. 1999a). As elk density increases, pregnancy rates (Houston 1982) decline, and a larger proportion of elk calves are born later and at a lower birth weight. These calves survive less well
(Singer et al. 1993), which reduces the rate of population growth (Houston 1982, Merrill and Boyce 1991, Coughenour and Singer 1996a, Singer et al. 1997, NRC 1998). Together, these factors produce a negative correlation between population growth and population size (NRC 1998, Taper and Gogan 2002), a clear indication of density dependence.
Weather has also had a major influence on the current migratory pattern of northern range elk. Deep snow restricts the area available for feeding by ungulates, and their response is to seek better foraging conditions at lower elevations (Coughenour and Singer 1996b). One result of the large migration of elk in response to the severe winter of 1988 was that some elk learned the landscape and continued to leave the park with greater frequency after 1989 (Figure 4–2). Elk migrations out of YNP were clearly influenced by weather. The size of the migration out of YNP is correlated with snow water equivalent (SWE),1 a rough measure of snow depth (NRC 1998). With more recent data (1989 to 1999), the correlation between the number of elk migrating from the northern range and SWE remains significant (Y =-2579.1+380.8; r2=0.41, p=0.03). Similarly, the number of elk killed in the late Gardiner hunt from 1976 to 1999 is correlated with SWE (Y=-333.3+71.5; r2=0.54, p=0.012). Neither the number of elk leaving YNP nor the take from the Gardiner hunt was correlated with population size (p>0.3). Although regression equations suggest that no elk will leave the park when SWE is less than 5 or 7 inches (12.7 to 17.8 cm), the lowest SWE recorded since 1949 was 10 inches (25.4 cm). Therefore, elk are likely to migrate from YNP in even the mildest of winters.
The postulated effects of severe winter weather on elk have been corroborated by simulation modeling. Landscape-scale simulations of northern range elk identified winter severity as the primary cause of major mortality events (Turner et al. 1994b, Coughenour and Singer 1996b, Wu et al. 1996). Thus, one of the most important factors determining variation in elk population size is one over which managers have no control.
The current pattern of winter use of Dome Mountain may be influenced by artificial feeding. In 1989, Montana Fish, Wildlife and Parks leased agricultural property on Dome Mountain that was used to produce alfalfa. The lease stipulated that only the first cutting of hay would be removed; all subsequent
growth would be left as winter forage for wildlife. From 1992 through 1996, an estimated 209 tons of hay per year was produced on this property (Montana Fish, Wildlife and Parks 1997 lease2), a desirable resource within easy reach of northern range elk. Although severe weather may have initially motivated elk movements out of YNP, the existence of high-quality and abundant forage has likely encouraged them to return to the area. A change in the relationship between SWE and elk north of Dome Mountain in winter is apparent from a comparison of data before and after 1989 (Figure 4–3).
A key issue surrounding the natural regulation policy is the size around which the population will fluctuate and the impact of those populations on the northern range. In the absence of wolves, recent estimates of the largest elk populations the winter range can support range from 16,000 to 22,000 animals (Coughenour and Singer 1996a, NRC 1998, Taper and Gogan 2002), a large increase from the early estimates of 5,000 to 11,000 elk (Grimm 1938, 1939; Cooper 1963; Cole 1969). The increases resulted from a better understanding of elk dynamics and from a large increase in the area of available winter range (Lemke et al. 1998). Because only a fraction of the total maximum population is actually observed, these estimates translate to a total population of 20,000 to 22,000 elk that could be supported under the environmental conditions of the past few decades.
Natural Variation in Elk Population Size
Wildlife managers typically prescribe actions that reduce variation in population size or resources. Management of northern range elk has been no different, and observed population fluctuations would likely have been greater in the absence of annual herd reductions, acquisition of winter range, and provision of supplemental forage. Annual harvests prevented elk from attaining high densities that could have exacerbated intraspecific competition, led to more severe nutritional deprivation and adversely affected the range. DelGiudice et al. (1991) found that, during the relatively mild winter of 1987, elk on the northern range showed signs of hunger by midwinter. Nutritional deprivation was significantly associated with declines in cow/calf ratios. During the harsh winter of 1988, winter mortality of elk was severe, and NPS
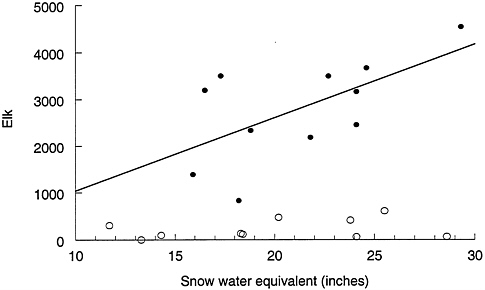
FIGURE 4–3 Number of elk counted north of Dome Mountain, Montana, as a function of SWE. Open circles are data from 1969 to 1988; filled circles are data from 1989 to 1999. Sources: Farnes et al. 1999, Lemke et al. 1998, Lemke 1999.
estimated that more than 4,000 elk died (Singer et al. 1989). Mortality during the 1988–1989 winter may have been increased by the previous summer’s drought combined with the large elk population, although a similarly severe winter die-off was reported in 1919 (reviewed by Houston 1982) and less severe die-offs occurred in 1974 and 1996 (YNP 1997; T.Lemke, Montana Fish, Wildlife and Parks, personal communication, January 18, 2000). Large mortality events may be relatively common among mammals, and their periodic occurrence is expected for Yellowstone’s ungulates (Young 1994, Erb and Boyce 1999).
Potential Impact of Wolves on Elk
Reintroduction of wolves to YNP in 1995 marked the restoration of the primary predator in the system. Wolves are probably a keystone species (i.e., a species that influences community structure out of proportion to their numbers) (Paine 1966) in the northern range and their activities could touch virtually every aspect of northern range ecology. Wolves regulate herbivore popu-
lations in other systems, with consequent effects on landscape and ecosystem processes (McLaren and Peterson 1994, Messier 1994, NRC 1997).
The ability of wolves to regulate Yellowstone’s elk population depends on how wolves’ consumption of prey changes with prey availability and whether the elk killed by wolves would have died from other causes anyway. Most predators increase their consumption of prey as food becomes more available, thereby reducing the population growth rate of the prey and stabilizing population fluctuations. However, all predators exhibit satiation at some point, and if wolves become satiated when elk are highly abundant, then wolves are likely to have a destabilizing effect, exacerbating population fluctuations caused by severe winters or other factors. An additional, critical consideration is whether wolves kill animals that otherwise would not have died. If wolves kill animals that otherwise would have died from old age or starvation, they add little to the rate of mortality.
Data necessary to develop detailed models of wolf-elk dynamics are unavailable, but simple models have been constructed to evaluate the likely range of effects of wolves on elk, and to a lesser extent, on the northern range ecosystem. Based on early models of wolf-elk dynamics, we could reasonably expect wolves to reduce the elk population by 5–20% (Boyce and Gaillard 1992, Mack and Singer 1993). So far, only Boyce and Anderson’s (1999) model includes stochastic variation due to predator behavior and vegetation dynamics. Boyce and Anderson’s (1999) model was intentionally simplistic in an effort to make the results interpretable, so their results are most useful for identifying qualitative trends in system responses. Boyce and Anderson compared effects of variation introduced at the bottom (vegetation) or top (predator functional response) of the system. When stochasticity was introduced by varying vegetation production, 95% of the variance in herbivore numbers was explained by vegetation alone. For this model, only 28% of the variance in population dynamics was explained by the number of predators. When variation entered the system at the top, via variation in the functional response of wolves, vegetation and predators alone accounted for 21% and 75% of the variation in herbivore numbers, respectively. These results identify a key problem for future research on interaction of wolves and herbivores: the source of variation in the system can have a profound effect on evaluation of the relative role of regulating factors.
Weather has had a very large impact on dynamics of elk in Yellowstone, and wolves are unlikely to change this. On the other hand, a bad year for elk is likely to be a good year for wolves; therefore, variation in elk-wolf dynamics will almost certainly result from variation due to both changes in forage pro-
duction and in predation by wolves. Severe weather per se is unlikely to have any direct effect on wolves, but severe weather may make it easier for wolves to kill elk, thereby driving greater variation in prey populations (Post et al. 1999).
Bison
Management of bison in and around YNP also has been controversial, but for different reasons. Bison in YNP are infected with the bacterium Brucella abortus, the causative agent of brucellosis (Meagher and Meyer 1994), a disease that causes abortion in cattle and is of significant economic and political interest to the livestock industry. Efforts to prevent transmission of brucellosis from free-ranging bison to nearby cattle have resulted in the slaughter of more than 2,000 bison on the boundaries of YNP, which has created a public outcry. The Yellowstone herd is the only free-ranging bison herd that avoided extermination in the late 1800s, and for many people, bison are a symbol of the American West. Americans continue to care passionately about management of Yellowstone’s bison. A recent draft management plan (NPS 1998) evoked more than 60,000 written comments from the public.
Bison were widely distributed in North America before YNP was created in 1872, but by 1900, only about two dozen free-roaming bison survived in YNP (Meagher 1973). Bison taxonomy remains controversial, but according to Reynolds et al. (1982) and Meagher (1973) plains bison (Bison bison bison) inhabited North America east of the Rocky Mountains; the wood or mountain bison (Bison bison athabascae) lived in grasslands in mountain valleys, parks, and northern boreal woodlands and tundra (Reynolds et al. 1982). Yellowstone’s remnant bison herd consisted of mountain bison. Park management of wildlife in the early 1900s involved supplemental feeding in winter, protection of bison within enclosures, and culling of weak individuals (Meagher 1973). Bison from domestic herds were transported to YNP to supplement the size of the tiny herd of wild bison, but many of those domestic animals were plains bison. The total number of bison in YNP in 1902 was 44 (Figure 4–4) (YNP 1997). With protection and intensive management, the bison population increased to more than 1,000 animals by the mid-1920s. Harvests were initiated then and conducted most years to keep the bison population at about 1,500 animals until the 1960s. By 1968 the population was 400 (Figure 4–4), after which harvest was stopped (Meagher 1973). In the absence of harvest after 1968, the bison population rapidly and consistently
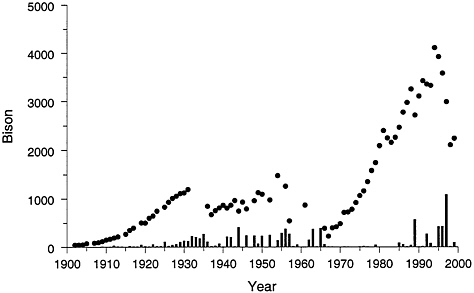
FIGURE 4–4 Total bison count (filled circles) and removals (vertical bars) in Yellowstone National Park. Sources: Dobson and Meagher 1996, YNP1997, Taper et al. 2000.
increased until the severe winter of 1988, by which time about 3,000 bison lived in YNP. Concerns over transmission of brucellosis from bison to domestic cattle had heightened by 1988, and more than 500 bison were slaughtered as they migrated out of YNP seeking suitable winter range. In the winter of 1996–1997, which had deep snows, more than 1,000 bison were slaughtered when they migrated outside the park.
Bison are gregarious and naturally live in nomadic herds. As the bison population in YNP expanded, bison eventually formed more or less discrete herds. Through 1968, when there were about 400 bison in YNP, bison formed three winter herd subunits and two summer breeding populations (Meagher 1973). Major wintering areas were the northern range, central YNP (Hayden Valley/Mary Mountain), and the Madison/Firehole area in western YNP.
Natural Regulation of YNP Bison
Does the YNP bison population exhibit density dependence, and if so, how many bison is the park likely to support? Relevant data are available for only two periods of protracted growth of the YNP bison population. The first was
from 1902 to 1931, when the population grew from fewer than 100 to more than 1,000 animals (Figure 4–4). Analyses of this period are complicated by management actions, which included fencing, artificial feeding, and castration of male calves (Meagher 1973). From 1931 to 1967 the size of the bison population was controlled by intense culling. The second period of protracted growth was from 1968 to 1994, which ended with the killing of more than 2,000 bison between 1994 and 1998. Bison in YNP were never allowed to achieve a population that appears to be in equilibrium, dynamic or otherwise. The analysis of bison population dynamics is further complicated by the nomadic and migratory habits of bison. Given the opportunity, bison travel long distances, and movements of up to 240 km were reported in a northern population (Soper 1941). Hornaday (1889, in Reynolds et al. 1982) documented migratory bison movements of several hundred miles. In YNP, bison clearly responded to increased population density by moving to new areas, both inside and outside park boundaries (Meagher 1989, NRC 1998, Taper et al. 2000). These traits complicate predictions of bison behavior in the Yellowstone ecosystem and suggest that management to restrict bison to park lands will need to be intense.
Given the tendency for bison to expand into new areas as population density increases, identifying density-dependent processes is difficult. Nevertheless, if the bison population exhibited density dependence, its per capita growth rate would diminish as density increases. From 1969 to 1981, the population grew at a more consistent and higher rate than after 1981 (Figure 4–5). Was the diminished growth rate after 1981 solely the result of park herd reductions, or is it an indication of density dependence?
The NRC (1998) examined this question, first focusing on growth of YNP bison from 1972 to 1995, a period with few artificial removals of bison. Over this period, the average annual increment to the population was 145 individuals (NRC 1998), which suggests a continuous decline in per capita reproduction throughout the range of population sizes. This would be a rather unusual pattern of density dependence for an ungulate population (McCullough 1990). The NRC (1998) suggested that this pattern was most likely to arise where female dominance strongly influenced calf survival or where there were few good habitats in which females successfully raised calves.
An alternative interpretation is that since 1968 the bison population has experienced two distinct growth phases. The first phase, from 1968 to 1981, was characterized by rapid and consistent growth rates. The second phase, from 1982 to 1999, has been characterized by high variance in growth rates, density-dependent effects on dispersion and movement, and a much more prominent effect of weather. Most data support this interpretation.
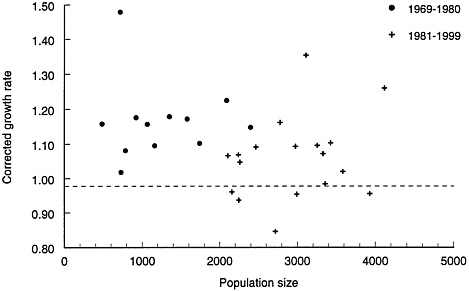
FIGURE 4–5 Growth rate (corrected for removals) of Yellowstone bison. Sources: YNP1997; Taper et al. 2000; P.Gogan, Montana State University, personal communication, May 12, 1999.
If suitable habitat for bison in Yellowstone was relatively abundant before 1982 and limited thereafter, annual per capita recruitment and growth rates should differ between these periods. Per capita growth rates can be estimated from the annual increment to the population, corrected for harvest. The corrected annual increment to the population is the difference in population size between years, with harvest added back (Nt+1-Nt+Ht). The average corrected increment does not differ between the two periods (160 vs. 162 bison), but there is a huge difference in variance between the two periods (Figure 4–6). Mean SWE was similar during 1982 to 1999 and 1969 to 1981 (19.9 vs. 22.3 inches [50.55 vs. 56.64 cm]), as was the number of above-average snow years (SWE#DXGT#21 inches [53.34 cm]; 8 vs. 7 years). Although this single index of winter severity fails to incorporate many important factors, SWE has thus far been the best predictor of bison movements. It reveals no differences in weather between the two periods, but growth rates were different. Overall, corrected annual growth rate was lower from 1982 to 1999 than from 1968 to 1981 (6% vs. 16%; p#DXLT#0.01), and the coefficient of variation was greater (0.20 vs. 0.47).
These analyses are subject to the usual uncertainties about population
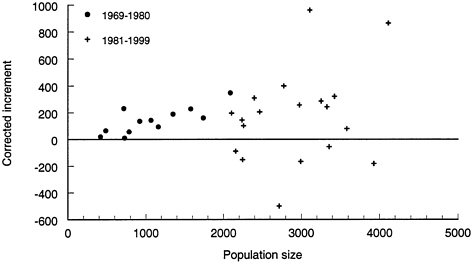
FIGURE 4–6 Annual increment in the YNP bison population corrected for harvest (corrected increment) as a function of population size. Sources: YNP 1997; Taper et al. 2000; P.Gogan, Montana State University, personal communication, May 12,1999.
estimates, but the trends are too strong to be ignored. They suggest that the Yellowstone bison grows at a constant per capita recruitment until a threshold size is reached, at which point density-dependent factors become important and growth rate, on average, declines (Stubbs 1977, Fowler 1987, McCullough 1990). Such a model suggests that competition has little influence on population processes until resources are largely depleted. For bison this makes sense. Intake rate is an asymptotic function of forage availability (Hudson and Frank 1987, Gross et al. 1993), thus competition is unlikely to influence survival until forage availability is reduced below a threshold level. Reduced survival of calves is most likely to be the first indication of intraspecific competition (Soether 1997, Gaillard et al. 1998), and there is good evidence that this occurs at higher population levels (reviewed by NRC [1998]).
Because weather is highly variable in Yellowstone and influences survival and recruitment, growth of the bison herd is subject to both density-dependent and density-independent influences. At low densities, bison have access to adequate forage in all but the most extreme winters, and their growth rate is stable and high. At high densities, overwinter mortality may be high because good habitat is fully occupied and some proportion of the population is forced
into poor habitat, significantly reducing growth rates. As density increases, the importance of both density-dependent (e.g., competition) and density-independent factors (i.e., weather) increases. Both factors contribute to stabilizing the population, but weather tends to add noise to the system and prevent the bison population from stabilizing.
It follows that Yellowstone bison were relatively unconstrained by habitat quality before about 1980. The density of bison in YNP during that period was relatively low, and bison had high and consistent rates of growth (Figure 4–4). Similarly, during the same period the Yellowstone elk population had rapid and consistent growth (Figure 4–1). By about 1980, bison appear to have reached a density where most habitat was fully occupied and, in a typical year, winter habitat was limiting. Bison responded by seeking better habitat, and mortality became density dependent. Data on bison movements are consistent with this hypothesis. Meagher (1989) noted that year-round use of the Blacktail area began in 1980 and that the winter of 1982 marked the beginning of annual use by bison of the area from Mammoth to Gardiner.
Other investigators have also concluded that Yellowstone bison have shown signs of density-dependence limitation. Taper et al. (2000) evaluated the most extensive set of bison observations made by Meagher from spring 1970 to fall 1997. The data came from surveys from more than 160 flights during which the location and size of more than 20,000 bison groups were recorded. Bison used the smallest area during rut and the largest area during winter. As population size and density increased, there was a large increase in the area used by bison during winter, from about 200 km2 in 1970 to more than 600 km2 in the 1990s. If intraspecific competition were an important factor regulating the size of the bison population, growth rate would be expected to decline as population size increased, after accounting for the effects of area. Taper et al. (2000) thus examined changes in growth rate as a function of density by adjusting for the area actually used by bison in winter. As density increased, growth rate declined (p#DXLT#0.05), although the predictive value of the regression was very low (r2=0.29) and much of the variation in growth rate is not explained by density alone. This suggests that density-independent factors, such as severe weather, were strongly influencing rates of recruitment or mortality.
Taper et al. (2000) estimated the number of bison that YNP might support on a sustained basis. To do so, they determined the area used by bison during winter within YNP (the season when bison are most dispersed) and the density of bison at which the annual growth rate was zero (recruitment=mortality).
These calculations led to an estimated population of about 3,200 bison. Taper et al. (2000) carefully stated that this estimate was a crude approximation and should be used only with great caution. The estimate assumed bison would not leave YNP (i.e., would not migrate as a response to increased density), and it implicitly included the effects of recent management actions that may have modified bison behavior, such as winter grooming of roads and control of bison movements at park borders.
The first response of bison to increased density is to expand the area used, especially during winter. Therefore, one would expect bison to frequently migrate to other areas outside park boundaries. Taper et al. (2000) accounted for this behavior by estimating the density at which bison are unlikely to expand their range beyond park boundaries. That density was approximated by the population of bison in 1984, when management actions began to remove significant numbers of bison at park borders. Bison used about 675 km2 of winter range at a population size of about 2,800, which may be the population size at which bison might remain inside YNP without management intervention.
Taper et al. (2000) invoked a rather complicated process to arrive at the estimate of 2,800 bison, but other lines of evidence yield estimates that are nearly the same. The NRC (1998) examined the number of bison removed by harvest at park boundaries as a function of population size. In general, bison begin regularly leaving park boundaries when the population exceeds about 2,500 animals; when the population was more than 3,000, the number of bison leaving YNP was highly related to SWE (NRC 1998). Finally, the bison population numbered about 2,400 animals in 1981, which seems to be a pivotal year in terms of its growth and movement. Taken independently, each of these lines of evidence is weak, but taken together, they provide a consistent picture of the response of bison to increasing density within YNP.
Brucellosis
Because of the importance of brucellosis, much attention and controversy has been directed at the disease (NRC 1998). Brucellosis is endemic in bison and elk herds in YNP. In elk (Thorne et al. 1997) and bison (Williams et al. 1997) brucellosis typically causes abortion of the first pregnancy after infection in most infected females. Epididymitis and orchitis may occur in bison bulls (Williams et al. 1993, Rhyan et al. 1997).
Population consequences of brucellosis on elk and bison on the northern
range are not specifically known. Loss of 7% to 12% of the calf crop has been estimated for elk in the Jackson elk herd, where brucellosis is endemic (Herriges et al. 1989, Smith and Robbins 1994). Nevertheless, elk and bison herds infected by B. abortus increase at about the same rates as herds without the disease (Peterson et al. 1991, Dobson and Meagher 1996, Williams et al. 1997, NRC 1998), because the effects of brucellosis have been overshadowed by density-dependent factors and climatic effects such as severe winters. Although it is unlikely to be important on the northern range, transmission of brucellosis from elk to bison is suspected to have occurred on the National Elk Refuge (NER) in Jackson Hole, Wyoming (Petersen et al. 1991, Williams et al. 1993, NRC 1998).
Elk on native winter ranges do not appear to maintain brucellosis (Morton et al. 1981, Thorne and Herriges 1992, Toman et al. 1997). Brucellosis was probably present in transplant stock moved from the NER to historical ranges, but with one possible exception (Robbins et al. 1982; E.T.Thorne, personal communication, 2001), it was not maintained.
Population density is important for understanding the dynamics of brucellosis in elk and bison and the likelihood that it will be maintained. At one end of the density spectrum is the situation on the NER and various state feedgrounds where elk are concentrated over artificial feed during the winter and early spring (Boyce 1989, Smith et al. 1997, Toman et al. 1997), when abortions due to brucellosis occur. In severe and even during normal winters, elk are essentially confined to feedgrounds because of limited native winter range or because deep snows preclude them from leaving. Because animals stand body-to-body while eating, a birthing or abortion event of an infected elk could expose large numbers of animals to B. abortus. The threshold density of elk required for brucellosis to be maintained within the northern range elk population is not known.
Management of brucellosis will remain a contentious issue, because no management alternative can simultaneously meet YNP’s goal of minimizing interference by humans and the livestock industry’s goal of brucellosis eradication. Management actions that might eventually eradicate brucellosis are limited to test and slaughter programs and vaccination, but the absence of a highly efficacious vaccine and a way to easily administer a vaccine to free-roaming wildlife are major difficulties. Risk management includes temporal and spatial separation of wild ungulates and livestock and vaccination of nearby cattle (Montana Department of Livestock and Montana Fish, Wildlife and Parks 2000).
Current management of brucellosis in YNP bison relies on the establishment of three management zones and a combination of hazing, test, and slaughter; vaccination when a safe vaccine has been approved for use; and acceptance of some free-ranging seronegative bison outside the park (Montana Department of Livestock and Montana Fish, Wildlife and Parks 2000). The primary purpose of this management plan is to reduce the risk of transmission of brucellosis to nearby cattle by removing the source of infection. The second purpose is to provide negative reinforcement to bison that leave YNP, thereby reducing future risk by diminishing the likelihood that bison will continue to leave YNP. Simulation results consistently show that test and slaughter procedures alone will not reduce the prevalence of brucellosis in bison (Gross et al. 1998).
Other Ungulates
Populations of mule deer, white-tailed deer, pronghorn, bighorn sheep, mountain goats, and moose inhabit the northern range. Because all these species are present in the park in small numbers, the natural regulation policy has not affected them directly. However, the status of pronghorn is sufficiently tenuous that they deserve special attention, regardless of the management of other species.
Mule Deer
Mule deer (Odocoileus hemionus hemionus) are the third most abundant ungulate on the northern range. Mule deer populations appear to have fluctuated on the northern range from perhaps fewer than 200 to more than 2,500 individuals in the past 95 years (Barmore 1980). Different counting techniques throughout this period make it difficult to track the population fluctuations with much accuracy. However, it appears that the population both inside and outside the park boundary on the northern range may have grown to 1,000 individuals by 1911 and oscillated near that number until 1987, when the population increased to more than 2,000.
Singer and Renkin (1995) report that minimum mule deer density in lower elevations during the period 1965 to 1968 averaged 4 deer per km2. By the late 1980s, minimum deer density averaged 2 per km2. Interestingly, Singer
and Renkin’s data (1995) do not indicate a correlation between minimum mule deer population density and minimum elk population density. Minimum average elk density increased from 6 per km2 to 16 to 19 per km2 during the same time period. In contrast, YNP (1997) indicates that mule deer increased during the 1980s simultaneously with increases in the elk population. Singer and Norland (1994) report that mule deer populations increased from approximately 1,000 to 2,200 individuals between 1979 and 1988. Northern range data suggest that the population fluctuated between 1,600 and 2,500 individuals from 1987 to 1999 (Gogan et al. 1999). The population is currently declining and will continue to decline as long as adult female survival is less than 85%, winter survival of fawns is less than 45%, and early winter fawn/100 doe ratios are less than 66. Lemke (1999) reports, however, that segments of the population (e.g., Region 3, east of the Yellowstone River) have increased in the past year.
Approximately 70% of the mule deer in the Gardner Basin migrate out of the park during the winter. The remaining 30% move to winter ranges within the park boundaries at lower elevations (YNP 1997). Some individuals migrate 80 km to their winter range (Wallmo 1978).
Mule deer overlap in habitat use with bison and elk, but they overlap little in their diets (Hudson et al. 1976, McCullough 1980, Singer and Norland 1994). Estimates of percent shrub, determined by microhistological analyses of feces collected on the northern range during the mid-1980s, show that almost 50% of the mule deer diet is composed of shrubs. Elk and bison diets, in contrast, are only 4.1% and 1.6% shrubs, respectively (Singer and Renkin 1995).
Park policy and harvest decisions made in southern Montana are likely to affect mule deer if their population begins to expand. The current levels of harvest and lack of growth suggest that either the harvest is additive (i.e., the animals harvested would not have died otherwise) or that density-dependent factors are becoming strong enough to limit their population growth. Fawn/doe ratios did not decline when populations increased, but ratios have not increased while the population has decreased. This suggests that density-dependent factors are not currently influencing this population. As with other mule deer populations in the Rockies, winter severity, a density-independent factor, probably plays the most important role in determining population fluctuations.
White-Tailed Deer
White-tailed deer have never been abundant in YNP even though they are native to the northern Rocky Mountains. They were apparently abundant in
the 1800s in the region around the park, but populations declined until the early 1900s because of overexploitation and habitat change due to forest fires, severe winters (Pengelly 1961, Allen 1971), and heavy browsing of their winter habitat by elk (Murie 1951). Population fluctuations occur, but historical accounts in the Greater Yellowstone ecosystem (GYE) suggest that white-tailed deer have always occurred in low numbers except in the presence of agriculture, and are an “occasional inhabitant” of the northern range (YNP 1997).
White-tailed deer have long been known to exhibit density-dependent population regulation (McCullough 1979). In the YNP region, adult females produce 1.8 to 2.0 fetuses per year, whereas yearling does are less fecund, producing 1.2 to 1.33 fetuses per year. Survivorship of fawns is less than 60% but may be slightly higher in lower elevations and riparian areas. Adult females generally have a survival rate of 65% and a mean life expectancy of 4.2 years. The small population on the northern range suggests that park policy will have little to no effect on white-tailed deer.
Pronghorn
If historical records are accurate, the current population of about 250 pronghorn in the northern range is less than 15% of that in the early 1900s (YNP 1997) (Figure 4–7). Regardless of early population estimates, Yellowstone’s pronghorn population is small, isolated, and at severe risk of extirpation (Goodman 1996, YNP 1997, Lemke 1999). Furthermore, northern range pronghorn are of special interest because their population may contain a unique genetic heritage (Lee et al. 1994).
Pronghorn in the northern range occur in lower elevations, primarily along the Yellowstone River from Devil’s Slide, north of the park border, to areas east of Tower Junction and up the Lamar Valley. Pronghorn are intolerant of deep snow and their distribution in the northern range is restricted in winter to lower windswept areas where food is exposed throughout the winter, particularly sagebrush grassland (Barmore 1980). The area of pronghorn summer range is considerably greater than suitable winter range, but about 75% of northern range pronghorn are thought to reside in the same area throughout the year, and only about 25% migrate seasonally to a higher-elevation summer range (Caslick 1998).
Estimates of the number of pronghorn in the northern range were up to
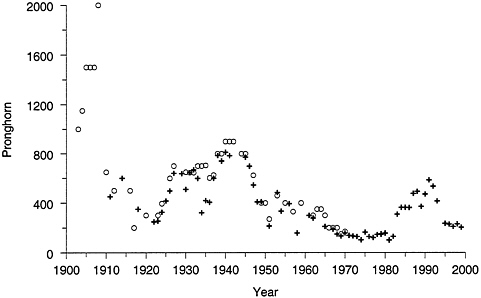
FIGURE 4–7 Pronghorn counts in the northern range. Open circles are estimated population sizes and crosses are actual counts. Sources: YNP 1997, Lemke 1999.
“thousands” during early European settlement, although it appears likely that no more than 1,000 to 1,500 pronghorn actually occupied the area near Gardiner (YNP 1997) (Figure 4–7). The population probably remained at that level until about 1910, after which estimates declined to as few as 200 in 1916 (YNP 1997). By the mid-1920s the pronghorn population recovered and remained relatively stable at 600 to 1,000 animals until 1950. Harvest of 258 animals in 1951 reduced the population to 300 to 400, and it remained at that level until 94 animals were harvested in 1966 (Chase 1986, YNP 1997). Surveys the following year observed only 188 pronghorn (YNP 1997). The harsh winter of 1967 killed about 42% of the herd (Barmore 1980). Withtheadoption of the policy of natural regulation, harvest of pronghorn within YNP was abolished in 1968. However, the population did not recover from the 1966 harvest and, in fact, declined to 100 to 200 until the early 1980s. Apparently favorable conditions during the 1980s, perhaps related to a series of mild winters, permitted the population to increase to the maximum recent count of nearly 600 in 1991. Over the past decade, the population has again declined, and since 1995 aerial surveys have counted no more than 235 pronghorn (YNP
1997, Lemke 1999). In spite of the small population, permits to hunt northern Yellowstone pronghorn when outside the park have been issued continuously since 1985.
Predation and winter nutrition are the factors most likely to restrict growth of the Yellowstone pronghorn population. Pronghorn in North America generally occupy open prairie grasslands, which may contain a high proportion of sagebrush (Artemisia spp.), often a major component of the pronghorn diet (e.g., 78% by rumen analysis, 93% by “utilization”) (Bayless 1969). In Yellowstone, the diet of pronghorn on their winter range is similar to the diets of elk, bison, and deer, so these species may come into direct competition during winter (Wambolt 1996). Overlap is less in the other seasons (Schwartz et al. 1977, Schwartz and Ellis 1981). In northeastern California and northwestern Nevada, more than 90% of the winter diets of both deer and pronghorn consisted of woody browse, including sagebrush species (Bayless 1969, Hanley and Hanley 1982).
Pronghorn may be particularly sensitive to severe winter conditions or competition because of their exceptionally high reproductive effort (Byers and Moodie 1990). Pronghorn at the National Bison Range in Montana always produced twins, and the total mass of offspring averaged 17% of the mother’s body mass (Byers and Moodie 1990). The rate of twinning by Yellowstone pronghorn is unknown, but pregnancy rates have been high (O’Gara 1968). Nutritional deprivation during winter can cause resorption of one or both fetuses (Barrett 1982) and lead to high overwinter mortality of fawns. Barrett (1982) estimated fawn winter mortality at 53.3% during a severe winter in Alberta. Similarly, severe winters resulted in adult mortality rates of 38%, 50%, and 62% in Saskatchewan, Alberta, and Montana, respectively (Pepper and Quinn 1965, cited in Martinka 1967; Barrett 1982). During a severe winter in Glasgow, Montana, ratios dropped from 90 to 110 per 100 does to 39 to 55 per 100 does at a sagebrush-poor site, whereas at a nearby site with abundant sagebrush there was no such depression (Martinka 1967). Access to prime winter feeding grounds can therefore be particularly critical for pronghorn. In Yellowstone, this was formally recognized in 1932 with the addition of 7,600 acres (3,077 ha) of winter range on the northern edge of the park “as an antelope preserve” (Caslick 1998). The large decline in the pronghorn population before 1961 “would probably have occurred without artificial reductions due to the decline of big sagebrush (Artemisia tridentata) in the park and, possibly, cessation of predator control in the park” (Barmore 1980). Big
sagebrush is the dominant food item in the winter diet of northern range pronghorn (Singer and Norland 1994).
Pronghorn are the smallest ungulate in YNP and thus they may be especially susceptible to smaller predators, such as coyotes and bobcats, which have difficulty capturing larger ungulates (Byers 1997). Records from the past 40 years indicate that fawn survival rates in Yellowstone are only about 25% (Barmore 1980, Caslick 1998). Caslick attributed most of the mortality to predation, with coyotes, bobcats, and golden eagles accounting for 12% to 90% of fawn mortalities (Caslick 1998). Of the 10 fawns radiocollared in 1991, eight were apparently killed by coyotes within 35 days (Scott 1991). Coyotes are clearly important predators on fawns, and Bruns (1970) specifically noted that coyotes did not appear to prey selectively on wounded or aged animals but fed on all age classes. Byers (1997) documented the extermination of pronghorn on the National Bison Range as a result of predation by coyotes.
Several factors may have increased the density of predators in critical pronghorn breeding areas. By 1998, reintroduced wolves were reported to have killed 6 pronghorn (1 adult and 5 fawns) (Caslick 1998), and wolves may have indirectly increased predation by coyote by displacing coyotes from areas farther from human development to areas where pronghorn live. In addition, late hunts for elk leaving YNP have created a source of abundant food for predators during winter because successful hunters leave animal waste in the field. Predators that consume hunters’ waste may switch to pronghorn in the spring.
Bison management may also affect pronghorn because the Stephen’s Creek bison facility, where bison are tested for brucellosis and then released or slaughtered, was built in key winter pronghorn range to hold bison that leave the park. Caslick and Caslick (annual reports 1995 to 1999) conducted weekly surveys for pronghorn during winter, recording locations where pronghorn were observed and factors that might influence pronghorn behavior or distribution. They recorded the status of gates (open or closed), observations of predators (coyotes, wolves, and domestic dogs), and disturbances such as operation of the Stephen’s Creek bison facility or the presence of visitors in pronghorn habitat. Pronghorn were never present in counting blocks containing the bison facility when it was in operation but were commonly observed in those areas when the facility was unused. Visitors also influenced pronghorn distributions, and pronghorn were much more likely to cross a fenceline via a gate (50% of Scott’s 1992 observations) than they were to go under the wire
or through buck and pole fences. Scott (1992) also noted that pronghorn were turned away by 28% of fence encounters, whereas pronghorn were detained within the structure of the fence 6% of the time. Because bison movements out of YNP are related to winter snow (NRC1998), the bison facility is more likely to be used during winters with deep snow, a period when pronghorn are also more likely to be stressed.
Human harvesting of YNP pronghorn has affected this population for more than 50 years, generally correlating with declining population numbers. More than 1,200 pronghorn were removed from the population between 1947 and 1968, a period when the herd declined from 625 to 200 animals. The population continued to decline after hunting ceased and a minimum count of 103 occurred in 1974. By 1985, 365 pronghorn were observed during population surveys, and repeated complaints from local ranchers led to the establishment of a game damage hunt for pronghorn in 1985 (Lemke 1998). The hunt was established by issuing permits to harvest 25 pronghorn annually to control perceived game damage to agricultural crops, but the area was also opened to hunters with a multidistrict bow hunting permit. Although the exact number of pronghorn killed is unknown, estimates are that up to 51 pronghorn per year were harvested from this area (Lemke 1999). In 1995, the number of damage hunt licenses issued each year was reduced to 5, but 12 animals were taken the following year (bow hunters took the balance). The population has again declined to about 200 animals. Effective in fall 2000, Montana Fish, Wildlife and Parks restricted hunting of this pronghorn population to five annual damage permits for an early season. Because few pronghorn use this area during hunting season, the projected harvest is one to three animals per year (T. Lemke, Montana Fish, Wildlife and Parks, personal communication, June 14, 2001).
The ability of the herd to recover from removals and hunting depends largely on factors that affect the production and survivorship of fawns. Fawn production depends on the ability of female pronghorn to maintain good condition, particularly during the winter, and this depends on the severity of the winter and the availability of food. An increasing elk population is reducing the availability of sagebrush, a key winter forage for pronghorn. In addition, winter use of YNP by people has dramatically increased, first because of snow-mobilers and more recently because of wildlife watching, especially for wolves. This pronghorn herd faces a serious risk of extinction. The risk could be reduced by eliminating harvest, by increasing harvest of coyotes and bobcats, by restricting recreational access to critical winter range, by reducing
disturbances within the winter range, and by management actions that result in enhanced growth and vigor of sagebrush within winter range. Survival of fawns begins with the condition of their dams but is most critically determined by predation pressure. Predators currently appear to be major contributors to fawn mortality.
Moose
Moose inhabit riparian and forested areas from northern Colorado through Alaska, where they typically feed on shrubby plants, especially willows. Many of the preferred foods of moose are early-successional plants; thus, disturbances such as fire and floods may be important for maintaining a suitable habitat. In general, moose are not migratory, although summer and winter ranges may differ in elevation. Moose are reputed to be the only ungulate in YNP whose winter range is higher than its summer range. Moose tend to spend most of the year as solitary animals or as female-offspring pairs (Miquelle et al. 1992).
Although relatively little is known about the ecology of moose that inhabit the northern range, the population clearly is small. Recent surveys resulted in estimates of about 200 animals, but the accuracy of the estimates is uncertain (YNP 1997). There are currently no surveys for moose because of the difficulty and expense associated with attempting to estimate the size of a widely dispersed population that inhabits heavily forested and rugged terrain.
Yellowstone’s northern range probably contains relatively little suitable habitat for moose. Tyers (1981) found that about half the forage consumed by northern range moose consisted of subalpine fir and lodgepole pine, their preferred winter forage. He found that most browsing took place in old forests. Although moose are large, they are selective feeders that choose higher-quality forage during summer and they prefer deciduous to conifer browse (Belovsky 1981, Renecker and Hudson 1988, Shipley et al. 1998). In the absence of predators, moose prefer summer riparian habitats with abundant willow. These observations suggest that moose in the northern range are surviving in marginal habitat and that the northern range is unlikely to support a substantial moose population. The abundance of moose has probably declined with the loss of willows.
Reintroduction of wolves is likely to affect moose, but because so little is known about northern range moose, we are unlikely to be able to tell whether
wolves have any effect on them. In other areas, moose populations have been regulated by the combined effects of predation by wolves and bears (reviewed by NRC 1997). Elk are likely to remain the primary prey of wolves, but the existence of a high-density wolf population presents a high risk to moose, especially calves.
Mountain Goats
Mountain goats were introduced by Montana Fish, Wildlife and Parks to the Absaroka Mountains northeast of YNP; they are exotic to YNP (Laundre 1990). As the introduced population increased, it expanded and colonized habitats on the border of YNP and, to a limited extent, within the park. Although diets of mountain goat and bighorn sheep overlap substantially (Laundre 1994), these species tend to live in very different habitats in YNP (Varley 1994, 1996). Outside park boundaries, mountain goat populations are controlled by hunting. The area of suitable habitat within the park is limited (Laundre 1990). Mountain goats are a management concern because in some habitats, especially in the absence of large predators, they have the potential to increase very rapidly (Hayden 1989) and achieve densities that may result in habitat degradation (Pfitsch et al. 1983), but this does not yet appear to have occurred in YNP.
Bighorn Sheep
Historical accounts suggest that the bighorn sheep population on the northern range was formerly much larger than in recent times (Schullery and Whittlesey 1992). The causes of the decline before 1981 are unknown but may include disease transmission from domestic livestock or competition with other ungulates. Although there is substantial diet overlap between elk and bighorn sheep (Houston 1982, Singer and Norland 1994), the species tend to use different habitats, and the extent of competition that might occur is unknown.
Small bighorn sheep populations can be highly susceptible to a variety of diseases that can result in catastrophic population crashes (Bunch et al. 1999); an epidemic of keratoconjunctivitis (pinkeye) occurred in the northern range bighorn population in 1981. This event was associated with a major population decline: bighorn counts declined from about 500 to fewer than 200 animals (Figure 4–8). Small bighorn sheep populations may be highly susceptible to
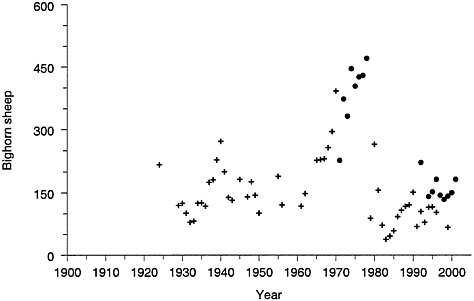
FIGURE 4–8 Bighorn sheep counts in the northern range. Filled circles are aerial counts and crosses are ground counts. Note that estimates from the methods differ, and the apparent large changes in population size are partly due to different survey methods and area surveyed. Sources: YNP (1997) for details of surveys; data from YNP (1997) and Lemke (1999).
regulation by predation (Wehausen 1996, Ross et al. 1997), and ongoing studies of mountain lions may provide a better indication of the relative importance of factors that may control the bighorn sheep population.
MAJOR PREDATORS IN THE YELLOWSTONE ECOSYSTEM
Four large terrestrial carnivores prey on Yellowstone’s ungulates: grizzly and black bears (Ursus arctos and U. americanus), mountain lions (Puma concolor), and wolves (Canis lupus). Coyotes (Canis latrans), medium-sized carnivores (Buskirk 1999), also prey on ungulates. Humans also kill substantial numbers of ungulates and may be considered a sixth major predator in the GYE.
Large terrestrial carnivores have large home ranges and move long distances, often ignoring political boundaries. For example, the range of Yellowstone’s grizzly bears “links most of the habitats, and associated species of the GYE” (Clark et al. 1999), and mountain lions radiocollared in the northern
GYE have dispersed to distant areas in Idaho, Wyoming, and Montana (Murphy 1998). As with most other mammals (Greenwood 1980), young males of these terrestrial carnivores typically disperse farther than females (Craighead et al. 1999).
Large carnivores typically utilize a variety of habitat types, and although they may specialize on particular species or sizes of prey, they consume a variety of prey and other foods as the opportunity or need arises (Johnson and Crabtree 1999). Bears eat primarily plant material, although Yellowstone’s grizzlies consume relatively more ungulates than most other grizzly populations (Jonkel 1987, Mattson et al. 1991, Knight et al. 1999). Large carnivores and their prey are intelligent and adaptable animals (Berger et al. 2001), which makes their interactions complex.
Among the influences on populations of large carnivores are changes in the abundance and relative abundance of different prey species; natural forces such as fire and weather, which influence the abundance, distribution, and availability of prey; human activities such as habitat alteration and hunting of prey populations; and changes in the abundance and behavior of other prey and predators.
Large carnivores inhabited Yellowstone long before the first European explorers arrived. Bones of wolves, coyotes, and grizzly bears were found in various strata during excavations at Lamar Cave, whose strata extend back about 3,000 years. Multiple Euro-American observers reported wolves, coyotes, grizzly and black bears, and mountain lions in the GYE before 1882 (Schullery and Whittlesey 1999).
Historical Human Impacts on Carnivore Populations
Predator eradication was a major goal in late nineteenth- and early twentieth-century America. Predators were easily poisoned with strychninelaced carcasses, and by 1880, wolf, coyote, and mountain lion populations in the GYE were greatly reduced (Schullery and Whittlesey 1999). When the U.S. Cavalry arrived in 1886, predators were at first protected along with other animals. However, to protect game species, poisoning of coyotes was resumed in 1898, and in 1907 army personnel were directed to kill coyotes, wolves, and mountain lions (Schullery and Whittlesey 1999). From 1904 to 1935, predator control in Yellowstone resulted in the killing of at least 4,352 coyotes, 136 wolves, and 121 mountain lions (Schullery and Whittlesey 1999). However, by the 1930s, there was mounting opposition among ecologists and
conservationists to predator control in national parks. In 1936, official park policy changed to protect native predators, including coyotes, although park managers could still kill individual predators deemed “harmful” (Schullery and Whittlesey 1999). However, wolves were effectively extinct in Yellowstone by then (Smith et al. 1999b) and predator control probably had eradicated mountain lions as well (Craighead et al. 1999).
Bears fared better than wolves and mountain lions. Bear populations were probably reduced by widespread poisoning of ungulate carcasses and by reductions in ungulate populations due to uncontrolled market hunting in the 1870s (Schullery and Whittlesey 1992). However, once they received army protection in 1886, bears began to feed at garbage dumps near hotels, where they soon became a popular tourist attraction (Schullery 1992, Knight et al. 1999).
YNP personnel supported and expanded the bear feeding program and bears ultimately were fed at numerous garbage dumps as well as along roadsides. Grizzlies dominated the dumps, although male black bears used them when grizzlies were not present; female and subadult black bears tended to beg for food from tourists on park roads (Knight et al. 1999). Large numbers of bears became habituated to humans, and they injured people and damaged property. These “problem” bears were then removed from the park. From 1930 to 1969, 46 people were injured by black bears, and an average of 24 black bears were removed from the park per year (Schullery 1992). Although the viewing of bears feeding at dumps was immensely popular with tourists, opposition to the practice grew and the park responded by closing the last public-viewing area at a dump during World War II, although it did not actually close the dumps until the late 1960s and early 1970s (Knight et al. 1999). Closing the dumps led to very high grizzly bear mortality; 229 grizzly bears were removed from the GYE between 1967 and 1972 as bears that previously fed in dumps began to seek food in campgrounds and threatened human safety (Knight et al. 1999). Black bears begging for food remained a common sight along roads until the late 1960s. By 1975, park managers had effectively eliminated this sight by improving sanitation, enforcing a no-feeding policy, and removing begging bears (Knight et al. 1999).
The states surrounding Yellowstone gradually reduced or prohibited hunting of grizzly bears, beginning with a prohibition by Idaho in 1946. However, a complete moratorium on hunting grizzly bears anywhere in the GYE was not imposed until 1974. The GYE grizzly bear was listed as threatened under the Endangered Species Act in 1975 (Knight et al. 1999).
Current Population Dynamics of Large Carnivores
Grizzly Bears
Yellowstone’s grizzly bears are the best-studied bears in the world, yet their status and future prospects continue to be subjects of vigorous controversy. Obtaining accurate estimates of the size and dynamics of a grizzly bear population is inherently difficult. Grizzlies are long-lived and females do not reproduce until they are about 6 years old. Grizzly bears are not easily observed as they are mostly solitary and travel over large home ranges in remote, mountainous, and largely forested country. There are additional logistical difficulties in reaching many parts of the study area, recapturing animals, and maintaining operative radiocollars on individual females to obtain long-term reproductive data.
Grizzly bear research began in YNP in 1959. An interagency Grizzly Bear Team was established in the early 1970s. At first, park authorities added to natural logistical difficulties by prohibiting radiocollaring of bears until 1975; then they allowed only one or two bears per year to be collared (Knight et al. 1999). By 1982 there were enough data for researchers to conclude that adult female mortality was high and that the reproductive rates were so low that the population was declining at about 2% per year (Knight and Eberhardt 1985). In 1983, an Interagency Grizzly Bear Committee was formed and charged with devising management strategies to reverse the population decline. Agencies began to manage habitat for grizzlies by eliminating sheep allotments within grizzly areas, increasing efforts to prevent illegal killing of bears, and changing policies such as food storage, garbage disposal, and removal of problem bears to minimize the need to legally kill bears (Knight et al. 1999).
Population analyses suggest that the Yellowstone grizzly bear population was relatively stable from about 1959 to 1993, with periods of slight increase or decrease (Eberhardt et al. 1994, Pease and Mattson 1999, Boyce et al. 2001). Pease and Mattson (1999) predicted a large (~15%) decline in the size of the grizzly bear population from 1993 to 1996 because of widespread failure of whitebark pine, a key food resource. Other models were less sensitive to this factor and suggested a slight increase in the size of the grizzly bear population over the same period. Counts of individual females with young cubs and survival rates also indicated a positive trend in grizzly bear numbers (Knight et al. 1999).
All recent estimates of the size of the Yellowstone grizzly bear population are in the low hundreds (Craighead et al. 1999). These include an estimate of
390 based on marked females; an estimate of 339 based on known families of bears; a statistical (bootstrapped) estimate of 344 with a 90% confidence interval of 280 to 610 bears (Eberhardt 1995); and an interagency review committee estimate of at least 245 bears, of which 67 were adult females (Eberhardt and Knight 1996).
The Yellowstone grizzly bear population is currently isolated from other grizzly populations and is not large enough to avoid loss of genetic variation in the short term (Harris and Allendorf 1989). The current genetic effective population size is only 13 to 65 bears (Paetkau et al. 1998). The genetic effective population size of a wild population, which is generally much less than the census size of the wild population, is defined as the size of a hypothetical population that would have the same rate of decrease in genetic diversity by genetic drift (or increase in inbreeding) as the focal wild population (Hedrick 1983). Loss of genetic diversity reduces population fitness and the probability of long-term survival; thus YNP’s grizzly bears probably need more protected habitat and dispersal corridors to preserve genetic diversity (Craighead et al. 1999). The best habitat for possible future population expansion appears to be in Wyoming. The greatest single threat to the population is increasing development of private lands, which not only decreases habitat but also greatly increases the potential for human-bear conflicts and consequent death of the bears involved (Knight et al. 1999).
Grizzly bears require a diverse habitat with minimal human disturbance to cope with climatic changes, alterations in the availability of different foods, human impacts, and changes in the abundance of other wildlife populations. Grizzlies can exploit marginal habitat to some degree but they require time to learn new habitat-use patterns when conditions change (Jonkel 1987).
In Yellowstone, grizzlies feed on weakened and winter-killed elk and bison from March through May, and they kill newborn elk calves during May and June (Singer et al. 1997, Knight et al. 1999). A few individual bears kill healthy elk during the summer, and bull elk become more susceptible during the fall rut. Seeds of whitebark pine (Pinus albicaulis), currently threatened by white-pine blister rust (Cronartium rubicola) in the GYE, are an important food for Yellowstone’s grizzly bears. About 30% of the most productive whitebark pine areas burned during the 1988 fires (Knight et al. 1999).
Black Bears
Very little is known about the numbers and population dynamics of Yel-
lowstone’s black bears; research has been directed disproportionately at grizzly bears. The population has probably decreased since dumps were closed and feeding bears along roadsides was stopped. However, the only study of Yellowstone’s black bears was conducted more than 30 years ago. Barnes and Bray (1967) estimated a minimum density of 0.07 bear per km2 in their Gallatin Mountain study area. Estimates of black bear densities in other areas of North America range from 0.1 to 1.3 bears per km2; however, densities are generally lower at higher altitudes (i.e., YNP) because of the shorter foraging season and poorer soils (Kolenosky and Strathearn 1987). Cole (1976) estimated that there were 650 black bears within YNP by extrapolating the Barnes and Bray estimate over a larger area. Craighead et al. (1999) estimated that there are currently fewer than 2,000 black bears in the GYE. The status of the population is uncertain because of the lack of data, although the population is generally presumed to be stable. About 1,000 black bears are legally killed in Montana each year (Craighead et al. 1999).
The ecology of black bears is known from many studies in other areas (Kolenosky and Strathearn 1987). Black bears are habitat and feeding generalists (Johnson and Crabtree 1999). They are usually forest dwellers, and the best black bear habitat is mixed forests that contain a variety of tree and shrub species of different ages (Kolenosky and Strathearn 1987). Black bears in Yellowstone typically prefer spruce-fir habitats and adjacent meadows but were often observed in lodgepole-pine forests along roadsides during the era of roadside feeding (Barnes and Bray 1967). Black bears are basically vegetarians and their diet appears to be largely determined by local food availability. Bears adapt to new sources of food and change their foraging habits accordingly. Black bears can kill young ungulates, which are vulnerable for a 2 to 4-week period after birth (Kolenosky and Strathearn 1987), and also eat carrion and insects. Killing young ungulates appears to be a learned behavior, and once learned, may continue to be part of an individual’s foraging routine (Kolenosky and Strathearn 1987). Carrion provided by cougars and other predators may be a significant food source for black bears in Yellowstone (Crabtree and Sheldon 1999a, Murphy et al. 1999).
Ecological conflicts exist between black and grizzly bears and may be depressing populations of both species (Jonkel 1987). However, interactions between Yellowstone’s black and grizzly bear populations have not been studied. Where both species coexist in Alaska, grizzly bears dominate black bears (Miller et al. 1997). In interior Alaska, grizzlies are most commonly associated with alpine tundra habitats, whereas black bears frequent forest and lowland areas (Klein et al. 1998). In the absence of grizzly bears in northern Quebec,
black bears have moved into tundra habitats normally occupied by barren ground grizzlies west of Hudson Bay (J.Huot, Faculty of Science and Genetics, University of Laval, personal communication, January 18, 2001).
Mountain Lions
Mountain lions have excellent long-distance dispersal abilities (Murphy et al. 1999). Young males have dispersed as far as 480 km from their natal home range (Craighead et al. 1999). Presumably, these dispersal abilities enabled mountain lions to recolonize YNP after they were locally eradicated by predator control programs. The GYE mountain lion population ranges over some 2,200 ha of relatively contiguous habitat (Murphy et al. 1999). Craighead et al. (1999) estimated that there are fewer than 500 adult mountain lions in the GYE, but they gave no basis for this estimate. Because of their high female survivorship and fecundity (litters of up to six kittens), mountain lion populations are more resilient than those of some other large predators, such as bears (Weaver et al. 1996).
Human-caused mortality is an important influence on most mountain lion populations, including those in the GYE outside the boundaries of YNP (Murphy et al. 1999). Most of this mortality is due to legal hunting: 48% of mortality among adult and subadult radiocollared mountain lions in the northern Yellowstone ecosystem was due to hunting; another 48% was attributed to natural causes (Murphy et al. 1999). Approximately 500 mountain lions were killed by hunters in Montana in 1998 (Craighead et al. 1999). However, mountain lion populations appear to have stabilized or even increased in many areas of the northern Rockies, including in the northern range area (Craighead et al. 1999), despite hunting pressure (Murphy et al. 1999).
Mountain lions occupy a wide range of habitats, although they prefer areas of steep and rugged topography (Lindzey 1987). Abundant cover is important to them as it provides security from enemies, including other predators and humans, and increases hunting success as mountain lions typically hide and ambush their prey. Thus, the structural characteristics of the local vegetation appear to be more important than the dominant plant species (Lindzey 1987).
Mountain lions are almost totally carnivorous and can kill all the YNP ungulates except adult bison. The diet of mountain lions varies with the abundance and availability of prey seasonally and geographically. Deer compose a major portion of their diet in most areas, although mountain lions also kill large numbers of small prey, such as snowshoe hares (Lepus americanus)
when they are abundant (Lindzey 1987). In the northern GYE, elk calves are a major source of food for mountain lions (Murphy 1998). However, because there are relatively few mountain lions and they kill far fewer ungulates than human hunters, Murphy (1998) concluded that they have little direct effect on the size of the elk and deer populations. Mountain lions are responsible for about 3% of the elk and 4% of the mule deer deaths in the northern GYE (Murphy 1998). Mountain lions kill about 12% of the buck mule deer, 9% of the elk calves, and 1% of the bull elk, but less than 5% of the other age-sex classes of elk and deer.
Wolves
Wolves in the northern Rocky Mountains were listed as endangered under the federal Endangered Species Act in 1974. Many scientists favored reintroduction over natural recolonization as a means of restoring wolves to the GYE. However, because of extensive controversy, wolves were not reintroduced to YNP until more than 20 years after they were listed as endangered (Bangs et al. 1998). In 1995, 14 wolves in three packs captured in Alberta, Canada, were introduced to YNP. In 1996, another 17 wolves in four packs captured in British Columbia, Canada, were introduced to YNP. In 1997, 10 pups and 3 adults from a pack captured in northwestern Montana (because they were chasing livestock) were released in the park (Bangs et al. 1998).
The wolves have thrived and are being extensively monitored. Ten packs in 1997 and seven packs in 1998 produced pups; by fall 1998, the population estimate was 116 wolves (Bangs et al. 1998). According to the Yellowstone wolf project report for 1998 (Smith 1998), litter size in 1998 averaged 5.5 pups and 81% of the 44 pups born survived to the end of 1998. Fifteen wolves died in 1998: five pups, four yearlings, and six adults. About half this mortality was due to natural causes, including wolves killed by other wolves, avalanches, and elk. The remaining mortality was due to human activities, including wolves killed by control actions, illegal shooting, and vehicles. Some assumptions made before the initiation of the wolf reintroduction program have been validated (Smith et al. 1999b): no preexisting wolves were found in the GYE and it was necessary to impose land-use restrictions around wolf dens. However, wolves have killed more than the predicted 120 ungulates per year and fewer than the predicted 19 cattle and 15 sheep per year. Many more visitors than expected have seen wolves.
It was predicted that wolves would travel outside the experimental area of
YNP and immediately adjacent neighboring GYE lands, but this had not occurred through the end of 1998 (Smith et al. 1999b). Wolves regularly disperse long distances: male wolves disperse a mean of 85 km and a maximum of 917 km from their natal home ranges (Craighead et al. 1999). Some wolves had dispersed out of the park as far south as the Jackson area (which is still part of the primary wolf recovery area) (Smith 1998), but none had dispersed from the area designated as the “Yellowstone Nonessential Experimental Population Area,” which includes all of Wyoming, Montana as far north as the Missouri River, and part of Idaho (Bangs et al. 1998).
Wolves in different geographical locations rely on different species of prey. Wolves in North America prey largely on ungulates and beavers, but also take other types of prey (Carbyn 1987). Elk have been the wolves’ major prey in the GYE to date, which is not surprising given the abundance of elk. However, the wolves are known to have killed every ungulate species present except for bighorn sheep (Smith et al. 1999b). In 1998, wolf researchers found 109 definite and 121 probable wolf kills (Smith 1998). Eighty-six percent of these kills were elk (198), followed by 3% each of mule deer (7), pronghorn (6), and coyotes (7); 2% of bison (5); and 1% each of moose (3), wolves, and unidentified prey. The age composition of the elk kill was 43% calves, 21 % cows, 19% bulls, and 16% unknown. Packs on the northern winter range killed an average of one ungulate every two to three days during March and one every three to four days during November and December.
Wolves can adjust to changing prey abundance and vulnerability (Messier 1995), although in multiple prey systems they may be slow to change their favored prey species (Dale et al. 1995). In some instances, wolves have switched to different age classes as their proportion in the prey population changed as a consequence of previous predation (Dekker et al. 1995) or winters with deep snow (Mech et al. 1995). Wolves have also learned to kill new types of prey (Klein 1995). However, it is difficult to predict how or when there may be substantial changes in the prey taken by Yellowstone’s wolves.
The Yellowstone wolves have been returned to an ecosystem that existed without them for much of the past century. Adjustment of the system to the presence of wolves is likely to take many years, as the wolves and the other components of the ecosystem adjust to one another (Klein 1995, Berger et al. 2001). Wolves will probably alter their patterns of prey selection, pack structure, movements, and population dynamics as the density, distribution, predation avoidance behavior, and population structure of their prey species change. A similar adjustment can be expected between wolves and other carnivores. A return to relative stability of predator-prey relationships within the Yellowstone
ecosystem may take longer than the period that wolves were missing from the system. Long-term monitoring of the predators and ungulates within the Yellowstone ecosystem provides a unique opportunity to greatly expand knowledge about interspecies relationships among upper trophic levels.
Coyotes
Coyotes are the smallest and most numerous of Yellowstone’s major predators. Although they were the first of the park’s predators to be intensively studied (Schullery and Whittlesey 1999), there was a long gap between the publication of Adolph Murie’s classic monograph on coyotes (1940) and the modern era of coyote research. The latter began in 1989, six years before wolves were reintroduced to the ecosystem (Crabtree and Sheldon 1999a). Reintroduction of wolves was expected to have a negative effect on Yellowstone’s coyote populations, as larger species of canids tend to compete for prey with, kill, and physically displace their smaller competitors.
Crabtree and Sheldon (1999a, 1999b) recently summarized data collected during these new studies of the coyote on Yellowstone’s northern range. Coyotes live in all GYE habitats below about 2,400m except for very steep, rocky areas and areas with deep snow. Crabtree and Sheldon (1999b) estimated an average pre-wolf density of 0.45 adult coyote per km2 on the northern range and densities from 0.1 to 0.4 coyote per km2 over much of the forest habitat in the GYE. Higher densities, sometimes exceeding 1 coyote per km2, occur in some of the more open grassland and shrub habitats. Craighead et al. (1999) estimated that there are fewer than 3,000 adult coyotes in the GYE.
Yellowstone’s coyotes are protected from hunting and trapping and inhabit an environment with good food resources. Crabtree and Sheldon (1999a, 1999b) argued that these factors explain many of the differences between Yellowstone coyotes and those studied in other areas, including larger pack size, greater social stability, higher adult survival rates (91% per year), higher mean age of adults, and lower dispersal rates of juveniles.
Coyotes on the northern range have a social system similar to that of wolves, living in territorial packs with an average size of six adults. However, unlike wolves, members of coyote packs often travel alone. Coyote territories are contiguous (no intervening spaces between them), nonoverlapping, and average 10.1 km2. Territory boundaries in the Lamar Valley and the Blacktail Plateau were very stable from 1990 through 1995, changing little from year to year.
Even though female coyotes become sexually mature around 10 months of age, Yellowstone females do not breed until they are 2 to 5 years old. An average of 5.4 pups are born per territory but an average of only 1.5 pups survive to 1 year of age. Principal causes of pup mortality are disease and starvation.
Coyotes feed primarily on voles and elk carcasses and the diet varies seasonally (Murie 1940, Crabtree and Sheldon 1999b). Ungulates provide about 45% of the coyotes’ annual biomass consumption, most of which is consumed as carrion during the five winter months.
The reintroduction of wolves in 1995 had dramatic effects on the coyote population (Crabtree and Sheldon 1999b). Wolves killed from 25% to 33% of the coyote population during each of the 1996–1997 and 1997–1998 winters, especially in the core areas used by wolves. Almost all these coyotes were killed near the carcasses of elk killed by wolves. The coyote population in the Lamar Valley dropped from 80 coyotes in 12 packs in 1995 to 36 coyotes in 9 packs in 1998 (Crabtree and Sheldon 1999b), and coyotes failed to recolonize their traditional territories in core wolf areas.
However, coyotes are flexible and adaptable animals and have already begun to travel in larger groups (Crabtree and Sheldon 1999b). Some of the surviving coyote packs are smaller and are producing larger, healthier pups with higher survival rates. Coyote packs on the fringes of wolf territories have experienced little mortality and are able to benefit from elk carcasses killed by neighboring wolves.





































