
6
Intraspecific Variability in Fertility and Offspring Survival in a Nonhuman Primate: Behavioral Control of Ecological and Social Sources
Jeanne Altmann and Susan C. Alberts
The great variability and complexity of human vital rates (age-specific survival and fertility rates) and of behaviors affecting these rates are topics of major investigation in many disciplines. The extent and nature of such variability for our closest nonhuman relatives are only beginning to be elucidated. Our goal in the present chapter is to investigate fertility and family behavior from the perspective of life histories—the schedules of vital rates—in a natural primate population. We do this by evaluating the potential fitness consequences, magnitude, and sources of variability in life histories, particularly of females, and in the behaviors affecting them.
Before doing so, we pause to place the life histories of both baboons (the focus of this chapter) and humans in a comparative mammalian perspective. Most evolutionary studies of life histories have been comparative within or among orders of mammals or even at higher taxonomic levels, and the answers to questions about life history variability and the behaviors affecting it are often quite different depending on the taxonomic level being investigated—vertebrates, mammals, primates, or a single species such as baboons or humans. Our brief comparative notes below are restricted to mammals. (In addition to the references that follow, the interested reader is referred to chapters in Boyce, 1988; Stearns, 1992; Charnov, 1993; Lee, 1999; many in Kappeler and Pereira, 2003; and references therein.)
INTERSPECIFIC, COMPARATIVE VARIABILITY IN MAMMALIAN LIFE HISTORIES
In studies that take a classical comparative approach, patterns of variability or constancy among species or higher taxonomic levels such as
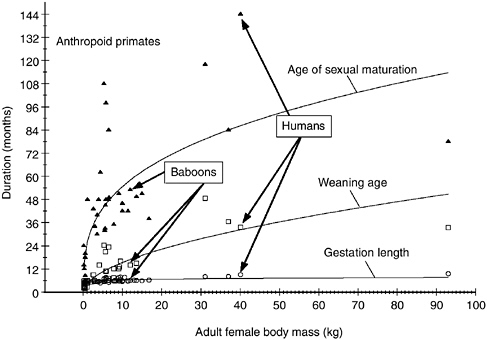
FIGURE 6-1 Duration of several life history stages as a function of body size in anthropoid primates.
SOURCE: Data primarily from compilations in Smuts et al. (1987), updated in Lee (1999) and references therein.
genera are analyzed. Consequently, single species such as humans or baboons represent at most a single point in the analyses. From that coarse perspective, several generalities can be made about the life history traits of various mammals. First, these traits covary and cluster along a continuum, such that some species have “fast” life histories—rapid offspring growth rates, early maturation, high rates of reproduction, and short reproductive spans (high adult mortality). At the other extreme are species with “slow” life histories—low rates of offspring growth, late maturation, low rates of reproduction and adult mortality. Second, large-bodied mammals tend to have slow life histories, small-bodied ones fast life histories (see Figure 6-1 for anthropoid primates). Third, although this pattern of a fast-slow continuum can be seen in all orders of mammals, the tendency toward slow or fast life histories also differs greatly among the various orders of mammals—carnivores versus primates, for example. Mammals of the same size in different orders differ fairly consistently in slow or fast life history style, and primates, mainly anthropoid primates, have particularly slow life histories. Finally, another interesting feature of the fast-slow continuum is that many life history traits, such as growth rates and adult mortality rates
(Charnov, 1993), remain strongly related to each other even when the effects of body size are removed; that is, constraints of size are not what lead to the correlation among traits, as was often assumed in many studies prior to the mid-1980s (see historical review in Harvey and Purvis, 1999).
Both humans and baboons exhibit slow life histories; those of baboons are basically as expected for a primate of their size, whereas some aspects of human life histories tend to be slower than expected (but see, e.g., Hrdy, 1999, and Hawkes, 2002, regarding human “hyperfertility”). That is, primates in general and anthropoid primates in particular have life histories characteristic of much larger nonprimate mammals. They also have particularly long periods of immaturity. Our human quality-based lifestyle runs deep in our phylogenetic history, and we come from a lineage, a family tree, that has at each branch exaggerated or extended the slow lifestyle—to a considerable extent a trade-off of quantity for quality. What explains these patterns, the differences among mammalian orders and the correlations found among life history variables at higher taxonomic levels? Diverse answers to those questions have been proposed, both historically and currently, and the interested reader is referred to Charnov (1993, 2001), Kozlowski and Weiner (1997), the historical review and perspective provided by Harvey and Purvis (1999), and an application to human life history evolution based on Charnov’s approach in Hawkes (2002) and Hawkes et al. (2003).
Most important, however, from the perspective of the current volume and the topic of this chapter—variability within a species, whether humans or baboons—is that good explanations of life history variability and correlations are not necessarily the same for all taxa or at all levels of investigation. The relationship among life history variables within species or populations often is, and is expected to be, different in direction, as well as strength, from that among orders (see, e.g., Lande, 1979; Harvey and Clutton-Brock, 1985; Emerson and Arnold, 1989; Lee et al., 1991; Worthman, this volume, for humans). For example, as a result of ecological sensitivity within, rather than among, species, large body size is often associated with large litters, early maturation, high reproductive rates, and low adult mortality rates, in striking contrast to the relationship of these variables among species of a given mammalian order. Life history theories that apply at one level cannot simply be extrapolated from that level to another—for example, from across mammalian orders or from differences among species within an order to variability within species (see, e.g., Lande, 1979; Emerson and Arnold, 1989; Kozlowski and Weiner, 1997).
BABOON LIFE HISTORIES: LIFE HISTORY PATTERN AND VARIABILITY
Comparative studies serve to anchor our perception of human or non-human primate traits to our shared biological history and some basic relationships such as those among body size, phylogeny, and life histories. They leave unanswered, however, questions about current dynamic patterns that are shaping behavior, life history variability, ecological responses, and evolutionary potential within species. These require analyses of lifetimes and of the factors that influence them. Here we present two analyses of life history variability in savannah baboons. First, we use matrix demography models to examine the relative strength of selection on different vital rates. In particular, we examine the sensitivity of fitness to comparable changes in infant survival and adult fertility. Second, we evaluate the variability existing in a natural population and the extent to which behavior, particularly choice of habitat and social environment, affects vital rates for both females and males.
Until recently, humans were the only primate species for which the requisite life history data were available for detailed analysis of life history variability (see Blurton-Jones et al., 1999, and Kaplan and Lancaster, this volume, and references in both). However, for a small handful of species, these data are accumulating for at least some life history components, and we provide here one of the first such analyses for the large, sexually dimorphic, predominantly terrestrial and highly social baboon, Papio cynocephalus. Selective omnivores, baboons are widespread throughout Africa and occupy a broad range of habitats from mountain through woodland and savannah to semidesert. The data presented here derive primarily from a study underway for more than three decades of the Amboseli baboon population, which resides in the basin to the north and west of Mount Kilimanjaro.
Baboons
Baboons live in discrete social groups. Members of a group forage during the day and sleep at night in much closer proximity to each other than to members of other groups, and virtually all social interactions are among members of the group of residence. A female usually spends her whole life in the group into which she was born, whereas a male leaves his natal group around the time he attains full adulthood at 8 years. Although groups are sometimes in close proximity, the boundaries are usually very clear, spatially as well as behaviorally. The amount of time that groups spend in close proximity is of relatively short duration and can be somewhat tense, even in habitats or years in which these encounters are rela-
tively more frequent (e.g., Cheney and Seyfarth, 1977; Shopland and Altmann, 1987). Within groups, adult females form clear dominance hierarchies that are predominantly stable both within and across generations as juvenile daughters assume the “family rank” about a year before menarche. Dominance rank in males, in contrast, is much more highly dependent on size and strength and is highly age dependent and unstable (Alberts et al., in press; Packer, 1979; Packer et al., 2000).
The larger African carnivores—leopards, lions, and hyenas—prey on baboons and are a particular a risk at night. In each habitat where they are found, members of a baboon group sleep close together either on cliff edges or high in those trees in their habitat that would be the most difficult for a predator to climb. Of the two major tree species in Amboseli, for example, baboons prefer fever trees, Acacia xanthopholea, to umbrella trees, A. tortilis; fever tree branches are higher off the ground, smoother, and more vertical. For baboons the distribution of sparsely scattered nighttime roosts, as well as of potable water and food resources, affects patterns of encounters between groups, daily travel, and seasonal variability in these patterns. In Amboseli, baboons of the fully wild-foraging groups awaken and descend from their sleeping trees shortly after dawn. For the next 11 to 12 hours, they spend almost 75 percent of their time foraging—feeding or traveling to food—across their short-grassland savannah habitat, approximately 10 percent socializing, and the remainder resting, often in a midday siesta.
Baboon infants weigh a little less than 1 kg at birth. In the first few months of life the infant clings to its mother’s ventrum and thereby obtains continuous nipple access and transportation during the 8 to 10 km of daily travel. Gradual nutritional and locomotor independence develops during the next year until the infant’s mother conceives again when the infant is about 18 months old and weighs approximately 3 to 4 kg. Although adult male baboons are approximately double the body mass of adult females, infant and juvenile females are very close in size to their male age peers, and almost all of the sexual dimorphism in body size arises during an adolescent growth spurt in males after females reach menarche between 4 and 5 years of age.
Life History Patterns in Amboseli
To analyze baboon life histories and life history variability, we used data collected from 1971 to 1999 for approximately 600 individuals living in completely wild-foraging groups of baboons (Alberts and Altmann, 2003). We constructed life tables with 1-year age classes and the corresponding survivorship and fertility entries of a population projection matrix, shown for females in Table 6-1 and males in Table 6-2. The tables also
TABLE 6-1 Per Annum Vital Rates for Wild-Foraging Amboseli Baboon Females (1971-1999) by One-Year Age Classes for Analysis in Projection Matrix Models
|
Female Age Class |
Entries in Population Projection Matrix |
Elasticity |
Age-Specific Birth Rate |
||
|
Survivorship |
Fertilitya |
Survival |
Fertility |
||
|
1 |
0.7910 |
0 |
0.0972 |
0 |
0 |
|
2 |
0.8884 |
0 |
0.0972 |
0 |
0 |
|
3 |
0.9366 |
0 |
0.0972 |
0 |
0 |
|
4 |
0.9688 |
0 |
0.0972 |
0 |
0 |
|
5 |
0.9529 |
0.1281 |
0.0906 |
0.0066 |
0.0083 |
|
6 |
0.9439 |
0.2719 |
0.0778 |
0.0128 |
0.3056 |
|
7 |
0.9481 |
0.2747 |
0.0661 |
0.0117 |
0.3500 |
|
8 |
0.9483 |
0.2639 |
0.0558 |
0.0103 |
0.3085 |
|
9 |
0.9427 |
0.2590 |
0.0466 |
0.0092 |
0.3256 |
|
10 |
0.9233 |
0.2230 |
0.0395 |
0.0072 |
0.2973 |
|
11 |
0.8910 |
0.1975 |
0.0338 |
0.0057 |
0.2429 |
|
12 |
0.8943 |
0.2303 |
0.0281 |
0.0057 |
0.2459 |
|
13 |
0.9419 |
0.2607 |
0.0226 |
0.0055 |
0.3273 |
|
14 |
0.9160 |
0.2588 |
0.0177 |
0.0050 |
0.3000 |
|
15 |
0.8701 |
0.2833 |
0.0129 |
0.0048 |
0.3333 |
|
16 |
0.8731 |
0.3000 |
0.0086 |
0.0042 |
0.3784 |
|
17 |
0.7880 |
0.2647 |
0.0055 |
0.0032 |
0.3704 |
|
18 |
0.6329 |
0.2162 |
0.0035 |
0.0020 |
0.3158 |
|
19 |
0.6278 |
0.2433 |
0.0022 |
0.0013 |
0.3000 |
|
20 |
0.8648 |
0.1832 |
0.0016 |
0.0006 |
0.4286 |
|
21 |
0.7455 |
0.2391 |
0.0009 |
0.0007 |
0.0000 |
|
22 |
0.5517 |
0.3207 |
0.0003 |
0.0006 |
0.7500 |
|
23 |
0.6090 |
0.1302 |
0.0001 |
0.0001 |
0.0000 |
|
24 |
0.4174 |
0.2138 |
0 |
0.0001 |
0.5000 |
|
25 |
0.3796 |
0 |
0 |
0 |
0 |
|
26 |
0.5000 |
0 |
0 |
0 |
0 |
|
27 |
0 |
0 |
0 |
0 |
0 |
|
a The fertility entries in the third column are the elements in the first row of the Leslie matrix, computed from the age-specific birth rates in the last column along with the person-years-lived entries from the estimated life table. The fertility entries take into account both adult survival in the interval and birth rate to individuals in that interval; it is based on the full cohort that enters an age class, whether they survive the age class or not. Because baboons, like humans, do not have a distinct birth season, our calculations are based on a birth flow model. For details, see Caswell, (2001), Alberts and Altmann (2003). |
|||||
contain age-specific birth rates and elasticities, which will be discussed shortly. As evident in the tables, Amboseli baboons experience high infant mortality, much lower female mortality during the late juvenile and early adult years, and then gradually increasing mortality in the latter portion of
TABLE 6-2 Per Annum Vital Rates for Wild-Fraging Amboseli Baboon Males (1971-1999) by One-Year Age Classes for Analysis in Projection Matrix Models
|
Male Age Class |
Entries in Population Projection Matrix |
Elasticity |
Age-Specific Birth Rateb |
||
|
Survivorship |
Fertilitya |
Survival |
Fertility |
||
|
1 |
0.7825 |
0 |
0.1066 |
0 |
0 |
|
2 |
0.9122 |
0 |
0.1066 |
0 |
0 |
|
3 |
0.9337 |
0 |
0.1066 |
0 |
0 |
|
4 |
0.9167 |
0 |
0.1066 |
0 |
0 |
|
5 |
0.9588 |
0 |
0.1066 |
0 |
0 |
|
6 |
0.9427 |
0.3734 |
0.0948 |
0.0117 |
0 |
|
7 |
0.9390 |
0.7759 |
0.0743 |
0.0205 |
0 |
|
8 |
0.9416 |
0.7791 |
0.0570 |
0.0173 |
0.0667 |
|
9 |
0.9311 |
0.7663 |
0.0428 |
0.0143 |
0.5754 |
|
10 |
0.9112 |
0.7423 |
0.0313 |
0.0115 |
0.8155 |
|
11 |
0.8944 |
0.7221 |
0.0222 |
0.0091 |
0.6493 |
|
12 |
0.8820 |
0.7074 |
0.0151 |
0.0071 |
0.8176 |
|
13 |
0.8456 |
0.6652 |
0.0099 |
0.0052 |
0.5543 |
|
14 |
0.8295 |
0.6467 |
0.0060 |
0.0038 |
0.4919 |
|
15 |
0.7816 |
0.5950 |
0.0034 |
0.0026 |
0.3007 |
|
16 |
0.6665 |
0.4747 |
0.0020 |
0.0015 |
0.3326 |
|
17 |
0.6251 |
0.4334 |
0.0012 |
0.0008 |
0.0776 |
|
18 |
0.8002 |
0.6252 |
0.0005 |
0.0006 |
0.0647 |
|
19 |
0.7500 |
0.5859 |
0.0001 |
0.0004 |
0.0485 |
|
20 |
0.3333 |
0.2131 |
0 |
0.0001 |
0.0776 |
|
21 |
0 |
0 |
0 |
0 |
0 |
|
a The fertility entry takes into account both adult survival in the interval and birth rate to individuals in that interval; it is based on the full cohort that enters an age class, whether they survive the age class or not. Because baboons, like humans, do not have a distinct birth season our calculations are based on a birth flow model. For details, see Caswell (2001) and Alberts and Altmann (2003). b Our calculation of birth rate (and therefore fertility) for males is based on the proportion of mating attributable to males of that age class (see text and Alberts and Altmann, 2003, especially p. 78 and Appendix 4.1 for reproductive rate terminology). |
|||||
the second decade of life (see also Bronikowski et al., 2002). Only a small proportion of individuals live into their third decade. At all three long-term field sites—Gombe and Mikumi, Tanzania, and Amboseli—maximum recorded longevity is 26 to 27 years (Gombe: Packer et al., 1995, and Bronikowski et al., 2002; Mikumi, estimated: Rhine et al., 2000; Amboseli: Bronikowski et al., 2002, and Alberts and Altmann, 2003). Mortality rates for subadult and adult males are somewhat higher than those for like-aged
females, reflecting a common mammalian sex difference. For the Amboseli baboons, this sex difference probably derives from a combination of intrinsic and extrinsic causes of senescence and mortality, including the mortality risk of dispersal (Alberts and Altmann, 1995). Known-age males of 15 to 18 years look much older and more frail than their female age peers.1
Menarche in baboons is followed by a period of somewhat abnormal sexual cycles and adolescent sub-fertility, followed by several normal cycles, conception, and then a 6-month gestation period; as a result, females produce their first offspring (Table 6-1) approximately 18 months after menarche. A long period of relatively steady birth rates follows until early in the third decade of life for the few females who live that long. These patterns are identifiable from near-daily records of menstruation, probabilistic visual correlates of ovulation, and other aspects of reproduction that are readily observed in baboons (Altmann et al., 1977, and references therein). Interbirth intervals after a surviving offspring are almost 2 years at various sites; if an infant dies, the interval is much shorter as its mother resumes cycling within a month and conceives after only one or two cycles.
Estimating male offspring production is more problematic and requires more caution in interpretation. In baboons, male dominance status is highly age related, dominance is a good predictor of mating behavior when a female is fertile, and observed mating behavior is a good predictor of genetic paternity (Altmann et al., 1996; Alberts et al., in press, and references therein). We used age-specific mating behavior to make proportional paternity assignments for males of each age; the total conception rate of males is constrained to and determined by that of females. Male offspring production declines much more rapidly with age than do birth rates of females, and the sex difference in this decline is much greater than that for survival (Tables 6-1 and 6-2).
A General Approach to Evaluating the Relative Strength of Selection on Different Vital Rates: Perturbations of Female Life Histories Using Matrix Models
If one were to compare two family lineages within a population, one lineage in which investment is successfully directed toward increasing infant survival and another in which it is successfully devoted to increasing birth rates, which would be more effective in enhancing population growth
or lineage fitness? Would enhanced parental care be favored or enhanced mating effort? The answer depends in part on the species’ basic schedule of mortality and birth rates, on the extent and style of fast or slow life history. Focusing the question on anthropoids, given the general pattern of age-specific mortality and fertility found in humans and other large primates (a slow life history), what would be the relative impact on biological fitness of two different behavioral changes, each of which appeared in some individuals, one a behavioral change that produced a small proportional change in infant mortality, the other a behavioral change that produced a small proportional change in birth rates? Demographic matrix models are a useful tool for exploring this question. (For a fuller discussion of these models, see references, below, and Alberts and Altmann, 2003, for an introduction to matrix models in the context of primate life history analysis.)
A matrix model is based on age-specific survival and fertility rates (from life tables). The model used is the usual Leslie matrix projection model, with one-year-wide age groups, familiar to demographers. Notation follows Caswell (2001). The model generates two results that are of particular interest for our purposes. The first result, λ, is a measure of the projected population growth rate. This measure equals er, where r is Lotka’s intrinsic rate of natural increase. The measure is also equivalent to the relative fitness of the life history described by the vital rates (Lande, 1982a, 1982b; see also McDonald and Caswell, 1993, and Caswell, 2001). The second result is elasticity (or sensitivity) measures, which provide a simple means to explore effects on λ of variability or small perturbations in the initial vital rates. These analyses are of particular relevance to a consideration of the life history consequences of fertility and parental behavior. Each vital rate (each age-specific mortality or fertility rate) in a matrix model will have a characteristic sensitivity, which is an estimate of the impact on λ of a small change in that vital rate (the slope of the vital rate function at that point) with all others held constant. Sensitivities cannot be compared directly because they are based on different rates. Elasticities are more useful; they are sensitivities that have been scaled so that their sum for both fertility and survival across all age classes is 1; they can be directly compared (see, e.g., Benton and Grant, 1999). A vital rate (in our case fertility or survival in a 1-year interval) with large elasticity is one for which small changes result in a relatively large change in λ compared to the effect on λ of a small change in the other vital rates. During the past two decades, matrix models have been greatly extended and many constraining assumptions have been relaxed (Caswell, 2001, and chapters in Heppel et al., 2000, and references therein). Concurrently these models have increasingly been applied to natural populations in studies of population viability and conservation (e.g., Heppel et al., 1994; Crooks et al., 1997; Mills et al.,
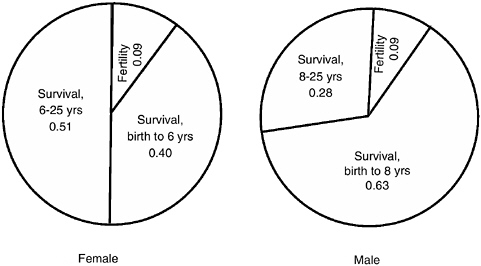
FIGURE 6-2 Proportion of elasticity in the matrix model attributable to immature survival, fertility, and adult survival.
NOTE: The greater proportion attributable to immature versus mature survival for males derives solely from the later maturity of males than females.
SOURCE: Data from Tables 6-1 and 6-2. See text and Alberts and Altmann (2003) for details.
1999), ecology (e.g., chapters in Heppell et al., 2000), and the evolution of behavior (McDonald, 1993; McDonald and Caswell, 1993).
If we return to the two lineages, one that invests in effective infant care versus the other that invests in an effective increase in fertile matings, we can compare—for example, in the Amboseli case—survival elasticities of young infants (age class 1) to fertility elasticities of adults (Table 6-1, Figure 6-2 ). For baboon females in Amboseli, a small increase in an infant’s survival during the first year of life will have a much greater impact on λ—up to two orders of magnitude greater—than a proportional increase in female fertility in any age during adulthood. Specifically, the elasticity of survival for age class 1 is 0.0972, whereas the elasticity of fertility is 0.0128 at its highest, in age class 6, and declines to an order of magnitude less, 0.0013, by age 19. The importance of survival versus fertility is evidenced by the fact that, for Amboseli baboons, survival accounts for 91 percent of the total elasticities and fertility for only 9 percent, for both males and females (Figure 6-2; see also Alberts and Altmann, 2003), a common pattern in long-lived species (discussion in McDonald, 1993, for avian species). The implication of these results for, say, an individual making alternative behavioral or investment decisions is complicated. Interpretation depends partially on whether these are lifetime decisions affecting at once fertility or
offspring survival for all years, flexible year-to-year options, or perhaps age-specific decisions (e.g., by young adult or old females). Interpretation also depends on the mechanisms and costs of each type of change; and on whether a key assumption of sensitivity analysis, independence among vital rates, is violated.
One example illustrates some of the potential issues in the case of baboons, although it is applicable to many other species. For an older female, the balance would always seem to favor investment in a current infant’s survival rather than in increased fertility (producing another infant). In contrast, for a young adult female in a particular year, a small increase in fertility that year might seem to balance an increase in survival of that single year’s infant; however, if infant survival is dependent on maternal care, that is, the independence assumption of sensitivity analysis is violated, the mother’s increase in fertility is likely to produce a decrease in the current infant’s survival rather than leaving it unchanged. For wild-foraging baboons, this is the case even if the current infant is in its second year of life. If the current infant is less than a year old, its death is almost assured if its mother dies or is caring for another infant. Moreover, as we shall see in the next section, the calculus may vary by ecological and social conditions (see also Hrdy, 1999; Ellison, 2001; Worthman and Kaplan and Lancaster chapters in this volume, for discussion of this topic in humans).
This example highlights several limitations or cautions that apply to interpretation of this simple matrix model and of sensitivity analyses in particular. One is the aggregate approach to the vital rates; that is, age classes are treated as groups of homogeneous individuals. Another is that vital rates are assumed to be independent of each other. Both of these assumptions are surely violated to varying degrees. A particularly important example of lack of independence is the case of male fertility rates; male fertility has been calculated by apportioning total female fertility across male age classes according to observed age-specific patterns of mating. Thus, an increase in fertility for one male age class is necessarily accompanied by a decrease in fertility for another. Another example is the situation in which trade-offs between female survival and reproduction or between future reproduction and survival of current offspring (example above) are significant. A third concern about sensitivity analysis is that infinitesimal, independent changes from one set of initial values may not be predictive of responses to larger and otherwise more realistic changes or for changes from a different set of initial vital rates because the fitness function for a vital rate is often not linear (e.g., Pfister, 1998; see also papers in Heppel et al., 2000).
An important complement to sensitivity analysis is direct perturbations of the matrix. Direct perturbation permits manipulation of a matrix to create hypothetical scenarios that are within the range, magnitude, and
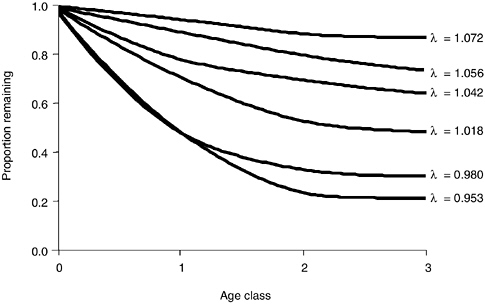
FIGURE 6-3 Projected population growth (or relative fitness) for different levels of first-year survivorship holding all other survival and fertility values to those in Table 6-1 (Amboseli fully wild-foraging groups, 1971-1999).
NOTE: The curve producing λ = 1.042 results from first-year survival in Table 6-1. The lowest value, λ = 0.953, results from 0.5 first-year survival obtained in a small sample of wild-foraging individuals during a period leading to a decline in the Amboseli population in the 1960s and early 1970s. The highest value, λ = 1.072, results from 0.9 first-year survival, obtained during a decade-long study of the lodge group, a “food-enhanced” group that obtained much of its nutrition from a tourist lodge grounds and the discarded foodstuffs at a lodge garbage pit. Published data for first-year survival from other sites fall within the range of survival explored here.
correlation structure realistic for the population of interest (see discussion and application in Mills et al., 1999). We implemented such a perturbation for females by using the basic model built from the female survival and fertility values in Table 6-1 and then simply changed first-year survival within a range from 0.5 through 0.9. This range of values includes all those reported for baboons or observed in Amboseli (see caption for Figure 6-3). This perturbation alone produced a large range of estimates of fitness, or λ, from a value below replacement to an appreciable expansion rate. That is, variability solely in first-year infant survival that is within the observed range for baboons has major consequences for estimates of relative fitness of both sexes and for the future of the population.
Clearly, the fitness potential is great for variability in effective family and fertility behaviors. Is such variability realized and, if so, how?
Variability in a Natural Population: Life History Plasticity and the Behavioral Ecology of Fertility, Offspring Survival, and Offspring Quality in Amboseli Baboons
What magnitude of variability is observed in the different components of the Amboseli baboons’ life histories, and what are the social and ecological sources of this variability? For our first analysis, we consider not just the fully wild-foraging baboon groups in Amboseli but also individuals from a group in the Amboseli population that has established its range and does some of its foraging around a nearby tourist lodge in the park (lodge group, Figure 6-4). In the subsequent analyses we again restrict analyses to data for the fully wild-foraging groups. All the analyses below require partitioning of the overall dataset in varying ways by time periods or subsets of individuals. Therefore, to retain reasonable sample sizes, we collapse the vital rates from 1-year age intervals into a few meaningful and manageable
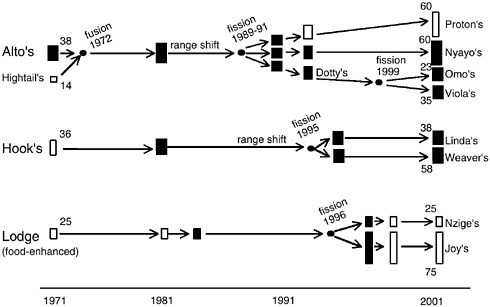
FIGURE 6-4 Amboseli study groups that contributed data to the present analyses.
NOTE: Although some demographic data are available for all groups in all years, detailed estimates of all vital rates are available only for years in which a group is indicated as a study group (darkened boxes). Approximate dates that Alto’s group and Hook’s group made major shifts in home range are indicated; years of group fusion and fission are indicated by arrows. The lodge group and its fission products are food-enhanced groups; all other study groups are completely wild-foraging.
components—infant survival, age at reproductive maturity, and reproductive rate throughout adulthood.
Furthermore, we focus on females for reproductive analyses. Data are better for females than males, because females are the nondispersing sex and because maternity can be assigned with greater certainty than can paternity. We examine variability in two reproductive parameters: reproductive maturity (measured as age at menarche) and fertility or reproductive rate (measured as interbirth interval for mothers with surviving offspring). Infant survival in this analysis is taken as survival through 2 years because interbirth intervals, and maternal investment, are approximately 1.5 to 2 years in duration in fully wild-foraging groups. As in the life tables and basic matrix model, for this first exploration and for consistency with the matrix model, data are aggregate, treated as independent, and pooled across all births in the subsets being compared. In addition, for simplicity and because of power limitations, interactions between ecological and social environments are not explored in the present analyses.
Choosing a Place to Live: Fertility, Offspring Survival, and Offspring Quality Are Food Limited
Home ranges differ in the resource base they provide for individuals or groups. Yet an individual’s home range is not something completely outside its control, a condition in which it must just do its best given the circumstances (Worthman, this volume, emphasizes this point for humans). In a heterogeneous environment, groups or individuals can move, for example, and thereby alter their foraging opportunities for the foreseeable future; that is, they may simultaneously alter many age-specific vital rates. Such an action has potential costs and benefits. The costs include short-term ones, such as dealing with a new environment in which the habits of predators and the locations of food and water are poorly known, and also perhaps long-term ones when, for example, a move may entail a trade-off between higher rates of predation and improved foraging. The benefits may outweigh the costs, however, and the move may in the balance be worthwhile if vital rates are highly food limited and if a move can improve foraging conditions.
For evaluating the extent to which offspring survival, offspring quality, and adult fertility are food limited for the Amboseli baboons, we made comparisons for two instances in which baboon groups chose living situations that changed food availability. First, we compared the totally wild-foraging groups in the Amboseli population with a group of baboons that moved into an area near the park’s tourist lodges and supplemented their diet with discarded food scraps from the lodge (Altmann et al., 1993; Altmann and Muruthi, 1988; Hahn et al., 2003; Kemnitz et al., 2002;
Muruthi et al., 1991). This situation is common wherever cercopithecine primates live near human habitation (see, e.g., Fa and Southwick, 1988). Second, we compared the fully wild-foraging groups in two time periods— the 1970s and the 1990s—before and after, respectively, the groups moved from an area of low food availability into a richer natural habitat, one in which baboon foods and shelter were more abundant (see Figure 6-4).
Food-Enhanced Groups
Adult females in the lodge group, the group that established a range near human habitation, experienced higher rates of reproduction and their offspring experienced much higher rates of survival and earlier menarche (Table 6-3, rows 1and 2; Table 6-4, row 1). The greatest proportional increase was in offspring survival. The home range and consequent lifestyle chosen by these females required tolerance of close association with humans, of higher density of baboons in close proximity, and of higher rates of aggressive interactions (Altmann et al., 1993; Kemnitz et al., 2002; Muruthi, 1989). Those individuals that tolerated these conditions were able to improve opportunities for their offspring and their fertility. Adult males, unlike females, usually leave their group of birth and disperse into other groups where they reproduce. Therefore, to achieve the advantages of life in a food-enhanced group, most Amboseli males would need to tolerate these altered conditions despite not having grown up in them, perhaps a much greater challenge than that experienced by females, who could adapt to these conditions during ontogeny. In fact, despite the much higher food availability and consequent lower foraging demand experienced by the lodge group, dispersal into this group did not occur over the first 16 years after they moved into close human association. Interestingly, dispersal out
TABLE 6-3 Impact of Foraging Environment on Various Life History Components
|
Foraging Environment |
Offspring Survival to 2 Years |
Offspring Growth |
Age of Maturation (years) |
Reproductive Rate (interbirth interval after surviving offspring) |
|
Food enhanced |
0.89 |
9 g/day |
3.72 |
1.28 |
|
Wild |
0.70 |
5 g/day |
4.52 |
1.72 |
|
Before move to better habitat (1970s) |
0.51 |
|
4.69 |
1.81 |
|
After move to better habitat (1990s) |
0.70 |
|
4.31 |
1.62 |
TABLE 6-4 Impact of Ecological and Social Factors on Various Life History Components (Percent change)
|
|
Offspring Survival to 2 Years |
Age of Maturation |
Reproductive Rate |
|
Food enhanced |
+27% |
+18% |
+26% |
|
Wild-foraging: Improved foraging (1970s-1990s) |
+38% |
+8% |
+15% |
|
Reduced density (fewer adult females) |
? |
|
+13% |
|
Improved social status (high ranking) |
= |
+8% |
+20% |
of the group by males that matured in the lodge group was reduced in comparison to males from wild-foraging groups; perhaps many males were unprepared or unwilling to face the hardships of a fully wild-foraging lifestyle despite the advantages of access to less closely related mates. As a result, an increasing proportion of potential mates in the lodge group were close maternal and paternal relatives. Although mate avoidance between maternal siblings is strong in many cercopithecines, including baboons, avoidance among paternal siblings is much less so (Alberts, 1999). Consequently, baboons from the lodge group were subject to an increased chance of inbreeding, and behavioral evidence of inbreeding has been associated with reduced offspring survival in wild baboons (Alberts and Altmann, 1995; see also Packer, 1979 for a group in Gombe).
Moving to a Better Wild-Foraging Environment
Between the 1970s and 1990s, the fully wild-foraging study groups moved from their original home range as the habitat degenerated into a richer woodland savannah area to the south and west of the original range, one in which baboon foods and shelter trees were much more abundant (Figure 6-4). The impact of this behavior on the vital rates was similar to that experienced by the lodge group (Table 6-3, rows 3 and 4), and the percentage improvement from the 1970s to the 1990s (Table 6-4, row 2) was again greatest for offspring survival.
In both cases of improved foraging environment, rapid offspring growth rates may be the life history variable mediating the improved survival and fertility that resulted. We were able to examine this possibility indirectly for the food-enhanced contrast; youngsters in the lodge group grew at almost double the rates of those in the wild-foraging groups, 9 versus 5 g per day
(growth data were available for only 5 years in the late 1980s, precluding a similar comparison over time in the wild-foraging groups).
Choosing a Social Environment: Fertility, Offspring Survival and Offspring Quality Are Density and Socially Limited
For individuals in the wild-foraging groups, we evaluated social sources of baboon life history variability. In particular we examined the effects of group size as a measure of immediate or experienced density. We also examined the effects of an individual female’s social status as study groups changed in size over time. The analyses were performed using a stepwise general linear model, after taking into account the effects of the change in foraging environment. When females were in larger groups (partial r2 = 0.05; slope, 0.0053; p = 0.0005), and particularly when those groups had more adult females (partial r2 = 0.08; slope, 0.0212, p = 0.0001), interbirth intervals after the production of a surviving offspring were longer, which lowered fertility for females with surviving offspring (see Figure 6-5). Each additional 10 adult females in a group resulted in an increase of approximately 0.2 years (2.5 months) in interbirth intervals after surviving offspring. In other words, living in small groups enhances reproduction but not offspring survival.
Although we suspected that high density inflicts a slight cost to infant survival, we did not have the power to detect such a cost in the present analyses. In the aggregate, fertility rates do not appear to exhibit density dependence. We postulate a scenario in which in high-density conditions the lower reproductive rates for females with surviving offspring are offset by the higher fertility that occurs when infants die, resulting in no effect on the aggregate fertility measurements used in our matrix models. We do not currently have a clear answer, but the situation highlights the need to look below the surface of overall fertility rates to evaluate density dependence. For baboons in several populations, life in large groups may sometimes entail costs to offspring survival and reproduction (Bulger and Hamilton, 1987; Rhine et al., 1988; Wasser and Starling, 1988).
The impact of large group size on vital rates is probably mediated at least partly through altered hormone levels or indirectly through physiological effects of reduced foraging efficiency from “scramble competition” without direct contest (van Schaik and van Noordwijk, 1988). This is suggested by data from Amboseli (Bronikowski and Altmann, 1996) as well as from other cercopithecine populations (Dunbar, 1996; van Schaik and van Noordwijk, 1988). One theory of primate social diversity and its evolution also assumes that higher rates of direct contest competition will be a source of life history variability in populations such as baboons with clear female dominance hierarchies (van Schaik, 1983, 1989), a mechanism that
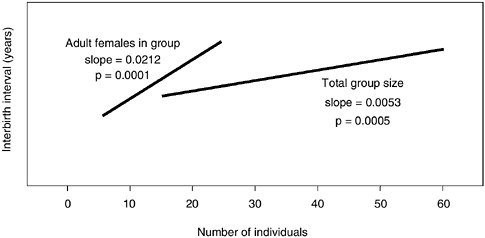
FIGURE 6-5 High density results in lower reproductive rate (longer interbirth intervals following a surviving offspring).
NOTE: Data pooled for all individuals in wild-foraging study groups. Interbirth interval following birth of a surviving offspring is a function of immediate experienced density as measured by group size (total group size and number of adult females in the group) after controlling for change in foraging environment. Although both measures are significant, a change in the number of adult females has a much greater impact (see also Figure 6-6). The range of numbers of adult females and total group sizes depicted here are approximately those observed in wild-foraging Amboseli groups. Interbirth intervals following infant death are excluded from analyses of interbirth intervals in this chapter because baboons, like humans, are not seasonal breeders and, with the loss of a suckling infant, a female rapidly resumes cycling and becomes pregnant in only one or two cycles rather than the usual three to five, resulting in much shorter interbirth intervals.
may occur under some conditions in Mikumi baboons (Rhine et al., 1988; Wasser and Starling, 1988). Future detailed analysis of both behavioral and hormonal variability among individuals may facilitate elucidating selective forces on primate social structure.
Can nonhuman females control group size? In species that exhibit female dispersal in addition to, or rarely instead of, male dispersal, females can control group size as males generally do, through dispersal, and the same is the case for fission-fusion societies (e.g., Wrangham, 1980; Dunbar, 1988). For species exhibiting matrilocality, those in which females remain for life in their group of birth, density may not at first seem to be a variable that females can control. However, control is possible through the rare occurrences of fission (van Schaik, 1996). Although fission of baboon or macaque groups has been reported primarily for groups that have experienced very rapid growth in group size through food enhancement, data on
fission of wild-foraging groups are also slowly accumulating (e.g., Nash, 1976; Ron et al., 1994). Amboseli females in fully wild-foraging groups have engaged in such fission events three times in the past three decades (Figure 6-4), resulting in a return to smaller groups and higher reproductive rates.
The costs of group living may not impact all adults equally, particularly in species with a social structure that is relatively hierarchical rather than egalitarian, including most cercopithecines (e.g., Wrangham, 1980; van Schaik, 1983). In baboon groups, inequality is evident in agonistic dominance hierarchies, a pervasive feature of baboon life (e.g., Hausfater, 1975; Altmann et al., 1988) and is predictive of access to various ecological and social resources (Barton, 1993; Barton and Whiten, 1993; Post et al., 1980; Silk, 1987, review for other primates). Adult female rank is very stable throughout adulthood in baboons and macaques, and daughters assume their family’s rank prior to adolescence (Hausfater et al., 1982; Pereira and Fairbanks, 1993; Walters, 1980). Despite widespread, though not ubiquitous (e.g., Watts, 1996), findings of dominance effects on foraging in a number of primate species, and despite long-postulated fitness consequences of dominance status, adequate data for rigorous tests remain scarce (van Schaik, 1983, and sequelae; Sterck et al., 1997).
In Amboseli, after taking into account effects of changing environment and group size, rank affects female reproductive rate (partial r2 = 0.07, slope = 0.013, p = 0.0037) and age of maturation (partial r2 = 0.08, slope = 0.02, p = 0.0025; see Figure 6-6). As with the impact of home range quality, these rank effects may be mediated by differences in offspring growth rate. Offspring of high-ranking females grew more rapidly, and rapid growth was predictive of early maturity. A 10-rank difference in dominance status was associated with a 0.2-year (2.5-month) difference in age of menarche and a 0.13-year (1.5-month) difference in interbirth interval. Busse (1982) reported a similar rank-associated difference in interbirth interval in chacma baboons in Botswana, and rank impacted a number of fitness components in anubis baboons in Gombe, Tanzania (Packer et al., 1995), and under some conditions in Mikumi baboons (Rhine et al., 1988; Wasser and Starling, 1988). Thus, through attainment and maintenance of high status, baboon females accrue fitness benefits from both enhanced fertility and offspring quality. Although the relative status of females in a group is ordinarily transmitted with great fidelity between generations (see Figure 6-7), a low-ranking female can reduce the number of others who dominate her or her daughters by participating in a group fission and thereby escape the costs of low status.
Male fertility is also density dependent in Amboseli and elsewhere (Alberts and Altmann, 1995; Alberts et al., in press; Altmann, 2000). Male mating opportunities are related to the number of females and the number
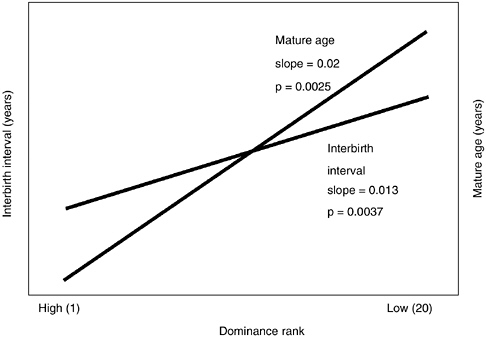
FIGURE 6-6 Age of reproductive maturity and reproductive rate (measured as in Figure 6-5) are functions of social status.
NOTE: High-ranking females experience higher reproductive rates and their offspring mature at younger ages than do low-ranking females (after controlling for changes in foraging environment and group size).
of other adult males in the group. As a consequence, adult males disperse among groups. That is, the costs entailed by living with many other adult males are often avoided or at least mitigated by dispersal to groups with more favorable demographic makeup (Alberts and Altmann, 1995; Altmann, 2000). The relationship between dominance status and mating success in male cercopithecines has a long, controversial, and yet well-documented history. Across species, among baboon populations, and within a single population over time (Amboseli), the relationship accounts for approximately 50 percent of the variance in male mating success but is highly variable (Alberts et al., in press). In most species and populations, male dominance status is strongly negatively associated with age (Alberts et al., in press; Packer et al., 2000). Males who stay in a large group or a group in which they are not of high rank sometimes use coalitions and other social means of enhancing fertility (Alberts et al., in press; Noë and Sluijter, 1990, 1995; Packer, 1977). Some older males who had previously been top-ranking and the father of many offspring also stay in groups in which
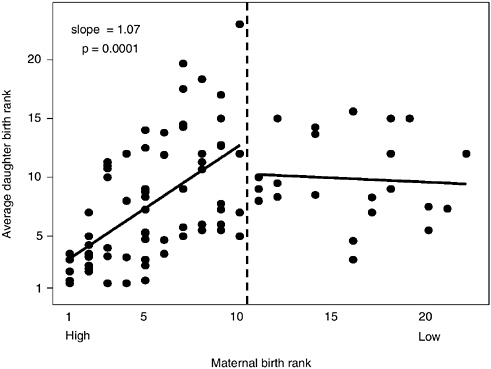
FIGURE 6-7 Daughters’ social status as predicted by that of their mothers.
NOTE: For maternal birth ranks 1 through 10 (left side of the figure), this effect is clear and strong (double that expected for a completely heritable trait). No effect occurs beyond rank 10 because group fission occurs beyond this size; females of different matrilines retain the predicted rank relative to each other in the fission products, but at the time of fission the new smaller groups have had 10 or fewer adult females in the wild-foraging groups (three fission events).
their fertility is low. They often provide care and protection for their likely offspring and are thought to reduce the likelihood of infanticide by other males (most recently reviewed in van Schaik and Janson, 2000, and Palombit, 2003), potentially gaining a few matings through female choice rather than dominance status (e.g., Strum, 1982; Smuts, 1985) but also potentially choosing enhancement of offspring survival over greater mating opportunities.
In summary, by altering their home range and their social environments through dispersal (males) or group fission (females), both males and females use behavior to change their physical and social environments. In the process, they often mitigate the effects of low social status, in ways that enhance fitness and sometimes in ways that particularly enhance offspring survival and quality (see Figure 6-8).
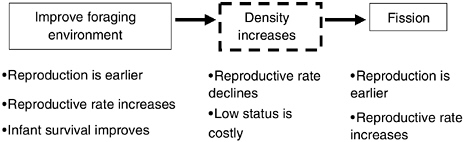
FIGURE 6-8 Overview of behaviors affecting fertility and offspring survival by fully wild-foraging baboons in Amboseli over three decades.
NOTE: The baboons shifted home range when the habitat in their previous range degenerated. Offspring then experienced higher rates of survival and matured at younger ages, and females reproduced more often if their offspring survived. As a result, immediate experienced density (group size and number of adult females per group) increased and reproductive rate then declined as a result of socially constrained reproduction. Group fission followed, removing the density and social constraints.
Covariance of Life History Components
We started with elasticity analyses that assumed independence among life history components. Violations of this assumption may sometimes be significant, and future analyses will need to explore the dependences. From a broad evolutionary perspective, strong linkages among life history variables in the form of life-history “invariants,” are central to Charnov’s comparative (interspecific) approach to the evolution of life histories (e.g. Charnov and Berrigan, 1993; Charnov, 2001, applications to human evolution in Hawkes, 2002, and references therein). Some of the same correlations may pertain at the level of intraspecific variability as well. However, life history correlations are usually weaker or even in the opposite direction in populations or species than at higher taxonomic levels (see Harvey and Clutton-Brock, 1985; Emerson and Arnold, 1989; Kozlowski and Weiner, 1997; Worthman, this volume). Within a population, some life-history components may be positively correlated because of shared underlying processes (discussed with particular focus on humans in Hrdy, 1999, and Ellison, 2001, and references in both). Under those conditions, behavioral changes will result in several rates increasing or decreasing together. This was seen for infant survival and early maturity in the Amboseli analyses. In other instances, individuals face trade-offs. For instance, producing another infant may inflict higher mortality risk on the mother herself or on her current infant (e.g., Altmann et al., 1988, for baboons). For the Amboseli baboons, we find that these trade-offs seem to be ecologically contingent;
they are evident only in the most stringent ecological habitats or years (Altmann et al., 1988).
Not only might vital rates be correlated within individuals, but the actions of one individual can affect reproduction and offspring survival of another. The most obvious instance is the effect of either parent’s behavior on the successful reproduction of the other, particularly in socially monogamous or polyandrous species. The phenomenon is more general, however, as in the case of alloparental care by siblings, grandparents, or other, perhaps more distant, relatives in which one individual may enhance the reproduction of another at a cost to its own reproduction. Enhanced fitness through the actions of others has been postulated for the evolution of alloparental care among some primate species and more specifically as an explanation for humans having “faster” reproductive rates than expected for their body size and otherwise slow life history components (see particularly Hrdy, 1999; Ross and McLarnon, 2000; Hawkes, 2002, and references therein). Covariance and other constraints, within and among individuals, are even more complicated for males. If males cannot appreciably affect total offspring production—that is, if offspring production is determined solely or even primarily by females— increased paternity in one age class will be offset by decreased paternity at other ages, and realistic perturbation analyses for males will need to take this trade-off into account.
The analyses presented here provide clear evidence of behavior that enhances fertility, offspring survival, and offspring quality. Changes in reproductive rates were most pervasive but not of greatest magnitude. Changes in foraging environment affected each parameter but most greatly affected offspring survival. In addition, daughters of low-status females matured later. The matrix model perturbation results raised the possibility that female baboons faced with alternatives might benefit from biasing decisions in favor of offspring survival and quality rather than enhanced fertility. The extent to which baboons are actually faced with such behavioral and life history trade-offs and behave as predicted is not yet clear. Both the simple models themselves and departures from their assumptions, such as population substructure, covariance, and stochastic variability, serve to guide future models and empirical investigations. Furthermore, that the social and environmental factors to which the baboons respond covary and feed back on each other is obvious (Figure 6-8) and is of necessity a topic of future theoretical and empirical investigation. Natural populations of nonhuman primates are relatively small and therefore present challenges to investigations of complexity and change over time, challenges that compound those already inherent in study of species with slow life histories.
CONCLUSIONS
Over a three-decade period, encompassing at least part of the lives of six generations of baboons and just surpassing the full lifetime of the longest living of the animals, individuals experienced great diversity of environmental and social conditions. These included normal wet and dry seasons; years of abundance and others of drought; many close relatives for some individuals or at certain times and few for others; habitat degeneration in the original home area and availability of not-too-distant areas with rich, new opportunities; groups with many competitors or with few. Groups of individuals, and also individuals acting somewhat independently, altered the physical and social conditions in which they lived and thereby considerably changed, through their own behavior, their own reproductive lives and the opportunities provided to their offspring.
Environmental variability has been proposed as the critical environmental context of human evolution (e.g., Potts, 1998). We suggest that adaptation to variable and changing environments is likely to have been an important feature of primate evolution more broadly. Success at responding to environments that change on various temporal and spatial scales and with varying degrees of predictability may be a recurrent theme for the most enduring and widespread primate lineages that we see today and for those likely to persist into the future. Because differences in survival of immature young will have the greatest impact on population growth and individual fitness for species with life histories characteristic of human and nonhuman primates, understanding fertility, parental behavior, offspring behavior, and the mechanisms producing variability in each must of necessity hold a central place in understanding primate adaptation and human origins.
For nonhuman primates as for humans, pregnancy, parturition, and offspring rearing occur within a complex ecological and social context. Few would question that the context of human fertility and parental care behaviors is highly variable and that humans both adjust their behavior to context and are often the agents of altering that context (e.g. Hrdy, 1999; Ellison, 2001; Worthman this volume; and references in each of these). That such variability and agency may also be significant in the lives and evolutionary history of our nonhuman primate relatives has not received comparable status. The opportunity to begin to do so offers considerable potential both for understanding primate behavioral ecology and evolution and for providing a window into human origins and diversity.
Together, the present analyses provide just one piece in beginning to elucidate the conditions, extent, mechanisms, and individual variability in richness of behavioral complexity related to fertility and offspring care in
natural primate societies. Future research will benefit from the use of more complex matrix demography models—for example, ones that incorporate temporal heterogeneity and covariance among life history components (Caswell, 2001). Studies of physiological mechanisms, ontogenetic effects, the effect of life history trade-offs on observed behavior (see above), and heritable differences in fertility and parental behaviors will also be essential to the agenda of elucidating ecological and evolutionary perspectives on fertility and parental care behaviors in nonhuman primates. The origin of both an absolutely and a relatively long period between birth and maturation in humans is seen among other large anthropoids. That this period of immaturity historically held and currently holds great opportunities for evolution is unsurprising. Increased links between studies of human and nonhuman primates, and of mechanisms and behavioral ecology, are essential to enhancing the research agenda of each.
ACKNOWLEDGMENTS
We thank the Office of the President of the Republic of Kenya and the Kenya Wildlife Service for permission to work in Amboseli over the years. We also thank the Institute of Primate Research of the National Museums of Kenya for institutional sponsorship in Kenya and the wardens and staff of Amboseli National Park and the local communities of the Amboseli/ Longido region for cooperation and hospitality. Particular appreciation goes to the Amboseli fieldworkers who contributed to the data over the years, especially R.S. Mututua, S.N. Sayialel, J.K. Warutere, P.M. Muruthi, and A. Samuels; to K.O. Pinc for database programming in the creation of BABASE and to the series of assistants who organized BABASE entry and extraction of BABASE data—D. Shimizu, S.L. Combes, A. Mosser, and J.M. Zayas. S.A. Altmann, R.A. Bulatao, K. Hawkes, H. Kaplan, J.W. Lynch, K. Wachter, and an anonymous reviewer provided valuable comments on the manuscript. We gratefully acknowledge financial support from the National Science Foundation (IBN-9985910 and its predecessors) and the Chicago Zoological Society.
REFERENCES
Alberts, S.C. 1999 Paternal kin discrimination in wild baboons. Proceedings of the Royal Society of London-Biological Sciences 266:1501-1506.
Alberts, S.C., and J. Altmann 1995 Balancing costs and opportunities: Dispersal in male baboons. American Naturalist 145:279-306.
2003 Matrix models for primate life history analysis. In Primate Life History and Socioecology. P. Kappeler and M.E. Pereira, eds. Chicago: University of Chicago Press.
Alberts, S.C., H.E. Watts, and J. Altmann In press Queuing and queue-jumping: Long term patterns of reproductive skew among male savannah baboons. Animal Behaviour. Available online at <http://www.duke.edu/~alberts/AlbertsLab/Queuing.pdf>
Altmann, J. 2000 Predicting male distribution among primate groups: Models of outcome and process. Pp. 236-247 in Primate Males. P. Kappler, ed. Cambridge: Cambridge University Press.
Altmann, J., and P. Muruthi 1988 Differences in daily life between semi-provisioned and wild-feeding baboons. American Journal of Primatology 15:213-222.
Altmann, J., S.A. Altmann, G. Hausfater, and S.A. McCuskey 1977 Life history of yellow baboons: Physical development, reproductive parameters and infant mortality. Primates 18:315-330.
Altmann, J., G. Hausfater, and S. Altmann 1988 Determinants of reproductive success in savannah baboons (Papio cynocephalus). Pp. 403-418 in Reproductive Success. T.H. Clutton-Brock, ed. Chicago: University of Chicago Press.
Altmann, J., D. Schoeller, S.A. Altmann, P. Muruthi, and R.M. Sapolsky 1993 Body size and fatness of free-living baboons reflect food availability and activity levels. American Journal of Primatology 30:149-161.
Altmann, J., S.C. Alberts, S.A. Haines, J. Dubach, P. Muruthi, T. Coote, E. Geffen, D.J. Cheesman, R.S. Mututua, S.N. Saiyalel, R.K. Wayne, R.C. Lacy, and M.W. Bruford 1996 Behavior predicts genetic structure in a wild primate group. Proceedings of the National Academy of Sciences 93:5797-5801.
Barton, R.A. 1993 Sociospatial mechanisms of feeding competition in female olive baboons, Papio anubis. Animal Behaviour 46:791-802.
Barton, R.A., and A. Whiten 1993 Feeding competition among female olive baboons, Papio anubis. Animal Behaviour 46:777-789.
Benton, T.G., and A. Grant 1999 Elasticity analysis as an important tool in evolutionary and population ecology. Trends in Ecology and Evolution 14:467-471.
Blurton-Jones, N., K. Hawkes, and J.F. O’Connell 1999 Some current ideas about the evolution of the human life history. Pp. 140-166 in Comparative Primate Socioecology. P.C. Lee, ed. Cambridge: Cambridge University Press.
Boyce, M.S. 1988 Where do we go from here? Pp. 351-361 in Evolution of Life Histories of Mammals: Theory and Pattern, M.S. Boyce, ed. New Haven, CT: Yale University Press.
Bronikowski, A., and J. Altmann 1996 Foraging in a variable environment: Weather patterns and the behavioral ecology of baboons . Behavioral Ecology and Sociobiology 39:11-25.
Bronikowski, A.M., S.C. Alberts, J. Altmann, C. Packer, K.D. Carey, and M. Tatar 2002 The aging baboon: Comparative demography in a nonhuman primate. Proceedings of the National Academy of Sciences 99:9591-9595.
Bulger, J., and W.J. Hamilton, III 1987 Rank and density correlates of inclusive fitness measures in a natural chacma baboon troop. International Journal of Primatology 8:635-650.
Busse, C.D. 1982 Social dominance and offspring mortality among female chacma baboons. International Journal of Primatology 3:267.
Caswell, H. 2001 Matrix Population Models. Sunderland, MA: Sinauer Associates.
Charnov, E.L. 1993 Life History Invariants. Oxford: Oxford University Press.
2001 Evolution of mammal life histories. Evolutionary Ecology Research 3:521-535.
Charnov, E. L., and D. Berrigan 1993 Why do female primates have such long lifespans and so few babies? or life in the slow lane. Evolutionary Anthropology 1:191-194.
Cheney, D.L., and R.M. Seyfarth 1977 Behavior of adult and immature male baboons during intergroup encounters. Nature 269:404-406.
Crooks, K.R., M.A. Sanjayan, and D.F. Doak 1997 New insights on cheetah conservation through demographic modeling. Conservation Biology 12:889-895.
Dunbar, R.I.M. 1988 Primate Social Systems. New York: Cornell University Press.
1996 Determinants of group size in primates: A general model. Proceedings of the British Academy 88:33-57.
Ellison, P.T. 2001 On Fertile Ground. Cambridge, MA: Harvard University Press.
Emerson, S., and Arnold, S. 1989 Intra- and interspecific relationships between morphology, performance and fitness. Pp. 295-314 in Complex Organismal Functions: Integration and Evolution in Vertebrates. D.B. Wake and G. Roth, eds. New York: S. Bernhard.
Fa, J.E., and C.H. Southwick 1988 Monographs in Primatology: Ecology and Behavior of Food-Enhanced Primate Groups. New York: Alan R. Liss.
Hahn, N.E., D. Proulx, P.M. Muruthi, S.C. Alberts, and J. Altmann 2003 Gastrointestinal parasites in free-ranging Kenyan baboons (Papio cynocephalus and Papio). International Journal of Primatology 24(2): April.
Harvey, P.H., and T.H. Clutton-Brock 1985 Life history variation in primates. Evolution 39:559-581.
Harvey, P.H., and A. Purvis 1999 Understanding the ecological and evolutionary reasons for life history variation: Mammals as a case study. Pp. 232-248 in Advanced Ecological Theory: Principles and Applications. J. McGlade, ed. London: Blackwell Science.
Hausfater, G. 1975 Dominance and Reproduction in Baboons (Papio cynocephalus): A Quantitative Analysis. Basel: S. Karger.
Hausfater, G., S. Altmann, and J. Altmann 1982 Long-term consistency of dominance relations among female baboons (Papio cynocephalus). Science 217:752-755.
Hawkes, K. 2002 Grandmothers and the evolution of human longevity. Paper presented at the symposium Strategies for Reproductive Success, Human Biology Association, 13 April.
Hawkes, K., J.F. O’Connell, and N.G. Blurton Jones 2003 The evolution of human life histories: Primate tradeoffs, grandmothering socioecology, and the fossil record . In Primate Life Histories and Socioecology. P. Kappeler and M.E. Pereria, eds. Chicago: University of Chicago Press.
Heppel, S., J.R. Walters, and L.B. Crowder 1994 Evaluating management alternatives for red-cockaded woodpeckers: A modeling approach. Journal of Wildlife Management 58:479-487.
Heppel, S., C. Pfister, and H. deKroon, eds. 2000 Elasticity analysis in population biology: Methods and applications. Ecology 81:605-708.
Hrdy, S. B. 1999 Mother Nature: A History of Mothers, Infants, and Natural Selection. New York: Pantheon.
Kappeler, P.M., and M.E. Pereira 2003 Primate Life Histories and Socioecology. Chicago: University of Chicago Press.
Kemnitz, J.W., R.M. Sapolsky, J. Altmann, P.M. Muruthi, G.E. Mott, and M.L. Stefanick 2002 Effects of food availability on serum insulin and lipid concentrations in free-ranging baboons. American Journal of Primatology 57:13-19.
Kozlowski, J., and J. Weiner 1997 Interspecific allometries are by-products of body size optimization. American Naturalist 149:352-380.
Lande, R. 1979 Quantitative genetic analysis of multivariate evolution, applied to brain:body size allometry. Evolution 33:402-416.
1982a Elements of a quantitative genetic model of life history evolution. Pp. 21-29 in Evolution and Genetics of Life Histories. H. Dingle and J. P. Hegmann, eds. New York: Springer-Verlag.
1982b A quantitative genetic theory of life history evolution. Ecology 63:607-615.
Lee, P.C. 1999 Comparative Primate Socioecology. Cambridge: Cambridge University Press.
Lee, P.C., P. Majluf, and I.J. Gordon 1991 Growth, weaning and maternal investment from a comparative perspective. Journal of Zoology of London 225:99-114.
McDonald, D.B. 1993 Demographic consequences of sexual selection in the long-tailed manakin. Behavioral Ecology 4:297-309.
McDonald, D.B., and H. Caswell 1993 Matrix methods for avian demography. Pp. 139-185 in Current Ornithology. D.M. Power, ed. New York: Plenum Press.
Mills, L.S., D.F. Doak, and M.J. Wisdom 1999 Reliability of conservation actions based on elasticity analysis of matrix models. Conservation Biology 13:815-829.
Muruthi, P. 1989 Food intake and energy expenditure in savannah baboons. MSc. thesis, University of Nairobi.
Muruthi, P., J. Altmann, and S. Altmann 1991 Resource base, parity, and reproductive condition affect females’ feeding time and nutrient intake within and between groups of a baboon population. Oecologia 87:467-472.
Nash, L. 1976 Fission in free ranging baboons in the Gombe Stream National Park, Tanzania. American Journal of Physical Anthropology 44:63-78.
Noë, R., and A.A. Sluijter 1990 Reproductive tactics of male savanna baboons. Behaviour 113:117-170.
1995 Which adult male savanna baboons form coalitions? International Journal of Primatology 16:77-105.
Packer, C. 1977 Reciprocal altruism in Papio anubis. Nature 265:441-443.
1979 Inter-troop transfer and inbreeding avoidance in Papio anubis. Animal Behavior 27:1-36.
Packer, C., D.A. Collins, A. Sindimwo, and J. Goodall 1995 Reproductive constraints on aggressive competition in female baboons. Nature 373:60-63.
Packer, C., D.A. Collins, and L.E. Eberly 2000 Problems with primate sex ratios. Philosophical Transactions of the Royal Society of London Series B-Biological Sciences 355:1627-1635.
Palombit, R.A. 2003 Male infanticide in savanna baboons: Adaptive significance and intraspecific variation. In Sexual Selection and Reproductive Competition in Primates: New Perspectives and Directions. C.B. Jones, ed. Special Topics in Primatology III American Society of Primatology (in press).
Pereira, M.E., and L.A. Fairbanks 1993 Juvenile Primates: Life History, Development and Behavior. Oxford: Oxford University Press.
Pfister, C.A. 1998 Patterns of variance in stage-structured populations: Evolutionary predictions and ecological implications. Proceedings of the National Academy of Sciences 95:213-218.
Post, D.G., G. Hausfater, and S.A. McCuskey 1980 Feeding behavior of yellow baboons (Papio cynocephalus): Relationship to age, gender and dominance rank. Folia Primatologia 34:170-195.
Potts, R. 1998 Environmental hypotheses of hominin evolution. Yearbook of Physical Anthropology 41:93-136.
Rhine, R.J., S.K. Wasser, and G.W. Norton 1988 Eight-year study of social and ecological correlates of mortality among immature baboons of Mikumi National Park, Tanzania. American Journal of Primatology 16:199-212.
Rhine, R.J., G.W. Norton, and S.K. Wasser 2000 Lifetime reproductive success, longevity, and reproductive life history of female yellow baboons (Papio cynocephalus) of Mikumi National Park, Tanzania. American Journal of Primatology 51:229-241.
Ron, T., S.P. Henzi, and U. Motro 1994 A new model of fission in primate troops. Animal Behaviour 47:223-226.
Ross, C., and A. MacLarnon 2000 The evolution of non-maternal care in anthropoid primates: A test of the hypotheses. Folio Primatologica 71:93-113.
Shopland, J.M., and J. Altmann 1987 Fatal intragroup kidnapping in yellow baboons. American Journal of Primatology 13:61-65.
Silk, J.B. 1987 Social behavior in evolutionary perspective. Pp. 318-329 in Primate Societies. B.B. Smuts, D.L. Cheney, R.M. Seyfarth, R.W. Wrangham, and T.T. Struhsaker, eds. Chicago: University of Chicago Press.
Smuts, B.B. 1985 Sex and Friendship in Baboons. New York: Aldine.
Strum, S.C. 1982 Agonistic dominance in male baboons: An alternative view. International Journal of Primatology 3:175-202.
Stearns, S.C. 1992 The Evolution of Life Histories. Oxford: Oxford University Press.
Stearns, S.C., and J.C. Koella 1986 The evolution of phenotypic plasticity in life-history traits: Predictions of reaction norms for age and size at maturity. Evolution 40:893-913.
Sterck, E.H.M., D.P. Watts, and C.P. van Schaik 1997 The evolution of female social relationships in nonhuman primates. Behavioral Ecology and Sociobiology 41:291-309.
van Schaik, C.P. 1983 Why are diurnal primates living in groups? Behaviour 87:120-144.
1989 The ecology of social relationships amongst female primates. Pp. 195-218 in Comparative Socioecology. V. Standen and R.A. Foley, eds. Oxford: Blackwell.
1996 Social evolution in primates: The role of ecological factors and male behaviour. Proceedings of the British Academy 88:9-31.
van Schaik, C.P. and M.A. van Noordwijk 1988 Scramble and contest in feeding competition among female long-tailed macaques (Macaca fascicularis). Behaviour 105:77-98.
van Schaik, C.P., and C.H. Janson 2000 Infanticide by Males and Its Implications. Cambridge: Cambridge University Press.
Walters, J. 1980 Interventions and the development of dominance relationships in female baboons. Folia Primatologica 34:61-89.
Wasser, S.K., and A.K. Starling 1988 Proximate and ultimate causes of reproductive suppression among female yellow baboons at Mikumi National Park, Tanzania. American Journal of Primatology 16:97-121.
Watts, D.P. 1996 Comparative socio-ecology of gorillas. Pp. 16-28 in Great Ape Societies. W.C. McGrew, L.F. Marchant, and T. Nishida, eds. Cambridge: Cambridge University Press.
Wrangham, R.W. 1980 An ecological model of female bonded primate groups. Behaviour 75:262-300.






























