
7
An Evolutionary and Ecological Analysis of Human Fertility, Mating Patterns, and Parental Investment
Hillard S. Kaplan and Jane B. Lancaster
This chapter considers the evolutionary biology of human fertility, parental investment, and mating and is designed to provide a broad overview of the topic. It focuses on three themes. The first is the timing of life events, including development, reproduction, and aging. Second is the regulation of reproductive rates and its relationship to parental investment. Sexual dimorphism and its relationship to mating systems together are the third theme. Each of these themes is addressed from two perspectives: first, in a comparative cross-species context, and second, in terms of variation within and among human groups. Our primary goal is to introduce a new ecological framework for understanding variations in each of those domains and then to apply the framework to understanding both the special characteristics of our species in a comparative perspective and variations within and among human groups. A secondary goal is to discuss how evolutionary biology can be integrated with more traditional approaches to human demography and the new research questions such integration would generate.
The first section of this chapter presents an introduction to life history theory and current thinking in evolutionary biology with respect to the three themes. Since the fitness consequences of alternative fertility and parental investment regimes depend on ecology and individual condition, both specialization and flexibility in life histories are considered. Building on this foundation, an ecological framework for understanding variation in each of those domains is then introduced. The second section discusses humans in a comparative context, with a particular emphasis on the hunter-and gatherer lifestyle because of its relevance to the vast majority of human
evolutionary history. The third section applies the framework developed in the first two parts to understanding major historical trends in human fertility, parental investment, and mating regimes. The transition from hunting and gathering to farming and pastoralism is considered first. Land- and power-based stratified societies are then discussed, followed by an analysis of wage-based competitive labor markets and demographic transition. The chapter concludes with a discussion of the new research questions and approaches to research design suggested by this framework.
THE THEORETICAL FRAMEWORK
Fundamental Trade-Offs in Life History Theory
Natural selection acts on variability in the traits of individual organisms within populations. Traits (and the genes that code for them) increase in frequency relative to other traits when their average effects on the individuals possessing those traits act to maximize their long-term production of descendents through time.1 Fertility is the most direct contributor to an organism’s fitness (i.e., the number of descendents it produces). In fact, all other fitness components, such as mortality, only affect fitness through their effects on fertility (e.g., mortality rates affect fitness by affecting the probability of living to the next reproductive event). All else constant, any increase in fertility increases an organism’s fitness. However, there are two trade-offs affecting natural selection on fertility.
The first is the trade-off between present and future reproduction. An organism can increase its energy capture rates in the future by growing and thus increasing its future fertility. For this reason, organisms typically have a juvenile phase in which fertility is zero until they reach a size at which some allocation to reproduction increases fitness more than growth. Similarly, among organisms that engage in repeated bouts of reproduction (humans included), some energy during the reproductive phase is diverted away from reproduction and allocated to maintenance so that it can live to reproduce again. The general expectation is that natural selection on age of first reproduction and on the adult reproductive rate will tend to maximize total allocations of energy to reproduction over the life course.
The second trade-off is between quantity and quality of offspring, where quality is a function of parental investment in offspring and reflects its ability to survive and reproduce. The general expectation is that natural selection on offspring number and investment per offspring will tend to maximize the long-term production of descendents; this may be estimated by the number of offspring that survive to reproduce themselves during an organism’s lifetime (Smith and Fretwell, 1974) or if fertility affects the production and survival of grandchildren, by more distant effects.
Sexual reproduction, which most probably evolved as a means of increasing variability among offspring through the sharing of parents’ genetic material, complicates the trade-off between quantity and quality of offspring. This is because offspring share roughly equal amounts of their parents’ genetic material, yet parents may contribute unequally to their viability. In this sense, offspring may be considered as “public goods,” with each parent profiting from the investments of the other and having an incentive to divert resources to the production of additional offspring. This public goods problem tends to create conflicts of interest between the sexes (see Gangestad, this volume, for a treatment of such conflicts).
In fact, an almost universal by-product of sexual reproduction is the divergent evolution of the two sexes. Sex is defined by gamete size, and the sex with the larger gametes is called female. Larger gametes represent greater initial energetic investment in offspring. With increased investment beyond energy in gametes, the divergence between the two sexes is often exaggerated but may also balance or even reverse. For example, females provide all investment to offspring in greater than 95 percent of mammalian species, but males provide similar amounts or more total investments among most altricial birds, male brooding fish, and some insects, such as katydids (see Clutton-Brock and Parker, 1992, for a review).
To the extent that one sex invests more in offspring than the other, the one that does more investing sex is in short supply resulting in operational sex ratios greater than unity and competition for mates among members of the sex that does less investing. This public goods problem generates the third major trade-off: that between mating and parental effort. Sexual reproduction involves two components: finding a mate and achieving a mating, on the one hand, and investing in the resulting offspring to increase its viability, on the other.
To the extent that there are gains from specialization in the two components, one sex will evolve to produce many small highly mobile gametes specialized to mating, and another will evolve to produce fewer larger gametes, specialized for energetic investments in offspring. Trivers (1972) recognized that these differences in relative parental investment affect the structure of mating markets and the characteristics of the more and less investing sexes. The more investing sex is selected to be choosy about when
and with whom to mate, and the less investing sex is selected to possess characteristics that increase its mating opportunities. This leads to what economists call negative externalities, since male resources are wasted on costly displays or handicaps (Grafen, 1991) or on fighting, rather than in offspring production. The general expectation is that natural selection acts on mating and parenting effort in populations of males and females so that individual fitness tends to maximize in a competitive equilibrium (i.e., it tends to generate distributions of mating and parenting effort among males and females that cannot be “invaded” by alternative distributions).
Ecology and Life History Evolution
Variations across taxa and across conditions in optimal energy allocations and optimal life histories are shaped by ecological factors, such as food supply, mortality hazards, and the effects of body size on both energy capture and mortality hazards (Charnov, 1993; Kozlowski and Weigert, 1987; Werner, 1986). It is generally recognized that there are species-level specializations that result in bundles of life history characteristics, which, in turn, can be arrayed on a fast-slow continuum (Promislow and Harvey, 1990). For example, among mammals, species on the fast end exhibit short gestation times, early reproduction, small body size, large litters, and high mortality rates, with species on the slow end having opposite characteristics.
It is also recognized that many, if not most, organisms are capable of slowing down or speeding up their life histories, depending on environmental conditions such as temperature, rainfall, food availability, density of conspecifics, and mortality hazards. Within-species variation in life history characteristics can operate over several different timescales. For example, there is abundant evidence that allocations to reproduction, as measured by fecundity and fertility, vary over the short term in relationship to food supply and energetic output among plants, birds, and humans (Hurtado and Hill, 1990; Lack, 1968). Extensive research on many bird species has shown that this phenotypic plasticity tracks fitness quite well (Godfray et al., 1991). Birds under variable conditions adjust clutch sizes in ways that tend to maximize the number of surviving young produced during the life course.
The impact of the environment may operate over longer time intervals through developmental effects (Lummaa and Clutton-Brock, 2002). For example, calorie restriction of rats at young ages tends to slow down growth rates and leads to short adult stature, even when food becomes abundant later in the juvenile period (Shanley and Kirkwood, 2000). Some intraspecific variation operates at even longer timescales, mediated through differential selection on genetic variants in different habitats. For example, rates of senescence vary across different populations of grass-
hoppers, with those at higher altitudes and earlier winters senescing faster than those at lower altitudes as a result of differential selection on genotypes (Tatar et al., 1997).
Similarly, there is a great deal of evidence suggesting that male and female parental investments vary in relation to local ecology over both the short run and the long run (see Clutton-Brock and Parker, 1992 for a review). For example, among katydids, males provide females with a “nuptial gift” (a bolus of condensed food energy) to support offspring production. Experimental manipulation of food density, affecting the foraging time necessary for males to produce the food package, produces shifts in male and female mating effort. When the food supply is low, male inputs into reproduction require more time than female inputs, males are in short supply, and females actively compete for males; as food density increases, this trend is reversed and males compete for access to females (Clutton-Brock, 1991; Gwyne, 1991; Gwyne and Simmons, 1990). This mix of specialization and flexibility is fundamental to understanding human life histories and mating systems. On the one hand, it is generally agreed that the large human brain supports the ability to respond flexibly to environmental variation and to learn culturally. This suggests that humans may be most capable of short-term flexibility in the timing of life events and investment strategies. On the other hand, the commitment to a large brain and the long period of development and exposure to environmental information necessary to make it fully functional place important constraints on the flexibility of the human life course and require specializations for a slow life history. In fact, consideration of brain- and learning-intensive human adaptation reveals shortcomings in existing biological theory and inspires the development of a more general approach to life history evolution, which is the focus next.
An Evolutionary Economic Framework
A general explanatory framework for understanding our species must be able to account for both its distinctive features when compared to other species and the enormous range of variation exhibited by humans under different conditions, in different societies, and at different points in time. To account for these evolutionary trends, we have expanded existing models of life history evolution by explicitly modeling the three trade-offs discussed above using capital investment theory (Becker, 1975; Kaplan, 1996; Kaplan and Robson, 2002; Robson and Kaplan, 2003). The processes of growth, development, and maintenance are treated as investments in stocks of somatic or embodied capital. In a physical sense, embodied capital is organized somatic tissue—muscles, digestive organs, brains, and so forth. In
a functional sense, embodied capital includes strength, speed, immune function, skill, knowledge, and other abilities. Since such stocks tend to depreciate with time, allocations to maintenance can also be seen as investments in embodied capital. Thus, the present-future reproductive trade-off can be understood in terms of optimal investments in own embodied capital versus reproduction, and the quantity-quality and mating-parenting trade-offs can be understood in terms of investments in the embodied capital of offspring versus their number.
The central thesis of this chapter is that there are four major factors affecting the timing of reproduction in the life course, reproductive rates, and parental investment for each sex: (1) the important resources consumed and utilized in reproduction and the production process by which those resources are obtained; (2) risks of mortality and the “technology” of mortality reduction; (3) the extent of complementarity between the sexes in the production of offspring; (4) the degree of variation in resource production and capital holdings among individuals and within individuals over time.
With respect to the first factor, the relative impacts of mass-based, brain-based, and extrasomatic physical capital on resource production are critical determinants (see Figure 7-1). What are the marginal effects of an increase in body size on acquisition and turnover rates for energy and other critical resources? What are the marginal effects of increases in brain size, brain complexity, knowledge, and skill on resource production? How does physical capital, such as land or a breeding territory, affect production? How do body mass, brain-based abilities, and extrasomatic physical capital combine in resource production? The general expectation is that, since investments in each of those forms of capital trade off against each other and against allocations to reproduction, natural selection will optimize those investments so as to maximize descendent production.
The brain is a special form of embodied capital. On the one hand, neural tissue monitors the organism’s internal and external environment and induces physiological and behavioral responses to stimuli (Jerison, 1973; 1976). On the other hand, the brain has the capacity to transform present experiences into future performance. This is particularly true of the cerebral cortex, which specializes in the storage, retrieval, and processing of experiences. To the extent that capital investments in the brain generate rewards that are realized over time (e.g., an increased reproductive rate during adulthood), the payoffs to those investments depend on mortality rates, since they affect the length of time over which the return will be realized. Dynamic models of this process show that investments in embodied capital coevolve with investments affecting mortality and longevity (Kaplan and Robson, 2002; Robson and Kaplan, 2003). The longer the time spent growing and learning prior to reproducing, the more natural
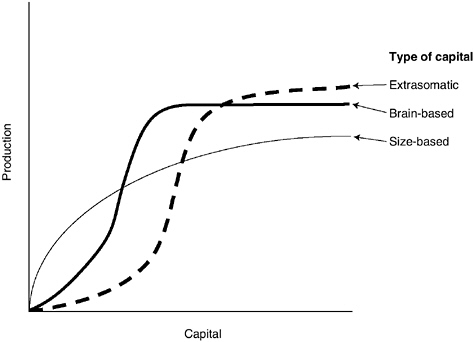
FIGURE 7-1 Production as a function of the capital stock.
NOTE: The relationship between production and each form of capital varies with ecology and the resources produced. Capital may be size-based, brain-based, or extrasomatic. More or less initial investment may be required before returns increase, and with further increases in investment, returns may diminish rapidly or slowly.
selection favors investments in staying alive to reap the benefits of those investments. Similarly, any investments that produce increased energy capture rates later in life, such as learning, select for additional investments to reach those older ages.
In addition to the production of energy, organisms can allocate energy and/or invest in forms of capital that reduce risks of mortality. While most biological models treat mortality as essentially exogenous, observed mortality is best understood in terms of an interaction between exogenous risks (environmental assaults) and endogenous responses designed to reduce mortality in the face of those risks. The technology of mortality reduction (the immune system, the ability to run, protective coverings such as shells, defensive weapons) also affects the likelihood of dying from environmental assaults. Models of embodied capital also show that ecological features or investments that increase the probability of survival to older ages also produce selection for greater investments in income-related embodied capi-
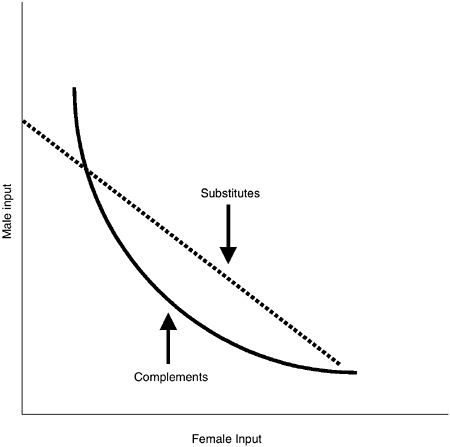
FIGURE 7-2 Offspring viability isoclines (indifference curves) as a function of male and female inputs.
tal. These coevolutionary effects appear to have been particularly important in human life history evolution.
With respect to the third factor, complementarity, each parent in sexually reproducing species contributes approximately half the offspring’s genes and some amount of parental investment. The fitness of offspring is likely to be some function of the genetic material and the investments received from each parent. Each of these inputs may act as substitutes or complements, as illustrated in Figure 7-2. Stated simply, complementarity occurs when the value of male investment in offspring depends positively on the amount given by females and vice versa (with fitness held constant).2 In
contrast, male and female inputs are substitutes when the relative values of the two inputs are independent of the amount provided by the other sex (again holding fitness constant). Thus, there are four axes of potential complementarity (e.g., between mother’s and father’s genes, between mother’s and father’s investment, and between each parent’s genes and their own and the other parent’s investments). Gains from specialization in parenting and/or mating effort and complementarity between genes and investment are forces favoring sexual dimorphism, with females typically specializing in parenting effort and males specializing in mating effort.
Complementarity between the investments of each sex is the force favoring decreased sexual dimorphism and increased male parental investment. This kind of complementarity can occur when both direct care and resources are important to offspring viability and when the provisioning of each conflicts with, or is incompatible with, the provisioning of the other. For example, protection and feeding of nestlings are incompatible among many flying bird species. Protection of the young by one parent complements provisioning by the other parent, since food is only valuable to offspring that have not been preyed on. This ecology favors biparental investment and taking turns in feeding and nest protection by males and females. Among grazing mammals, however, offspring follow their mothers, who are able to nurse and protect them simultaneously. Investments by males in this case are less complementary and would only substitute for the investments of females.
Mate choice criteria and mating “market” characteristics are expected to result from the variance among and within individuals over time with respect to the resources critical for reproduction. When females provide all the parental investment in response to the conditions discussed above, they are expected to exercise choice among males in terms of their genetic quality, and males are expected to compete with other males for access to fecund females, either through physical competition or appeals to female choice.
As support from males increases in value with a resultant increase in their contribution to reproduction, we expect female choice to respond to variation in male offers of investment and in their ability to acquire resources utilized in reproduction. Males, in turn, as their investments in offspring increase, are expected to exert choice with respect to variation in female quality and to compete with other males for access to the resources utilized in reproduction. Ecological variability affecting the variance among males and females in resource access or access to mates is expected to exert a significant influence on mating market dynamics and in male and female investments in parenting and mating effort. Intertemporal variation in productivity within individuals is also likely to affect their mate value because it increases the likelihood of shortfalls.
Our proposal is that human evolution has resulted in a specialized life history that is due to a particular constellation of the factors discussed above. This constellation derives from the hunter-gatherer way of life, which characterized the vast majority of human evolutionary history. While, as discussed in the next section, there are some universal features associated with this way of life, there is significant ecological variability across habitats. We also propose that as a result of exposure to such variation, human psychology and physiology have evolved to respond in systematic ways to variations in the four factors discussed above. Finally, the domestication of plants and animals and subsequent economic transformations produced new socioecological conditions to which people responded in radical shifts in parenting and mating practices.
HUMAN LIFE HISTORIES IN A COMPARATIVE CONTEXT
Relative to other mammalian orders, the primate order is slow growing, slow reproducing, long lived, and large brained. Humans are at the extreme of the primate continuum. Compared to other primates, there are at least four distinctive characteristics of human life histories: (1) an exceptionally long life span, (2) an extended period of juvenile dependence, resulting in families with multiple dependent children of different ages, (3) multigenerational resource flows and support of reproduction by older postreproductive individuals, and (4) male support of reproduction through the provisioning of females and their offspring. The brain and its attendant functional abilities are also extreme among humans.
Our theory (Kaplan et al., 2000; Kaplan and Robson, 2002; Robson andd Kaplan, 2003) is that these extreme values with respect to brain size and longevity are coevolved responses to an equally extreme commitment to learning-intensive foraging strategies and a dietary shift toward high-quality, nutrient-dense, difficult-to-acquire food resources. The following logic underlies our proposal. First, high levels of knowledge, skill, coordination, and strength are required to exploit the suite of high-quality, difficult-to-acquire resources that humans consume. The attainment of those abilities requires time and a significant commitment to development. This extended learning phase during which productivity is low is compensated for by higher productivity during the adult period, with an intergenerational flow of food from old to young. Since productivity increases with age, the time investment in skill acquisition and knowledge leads to selection for lowered mortality rates and greater longevity because the returns on the investments in development occur at older ages.
Second, the feeding niche specializing on large valuable food packages, and particularly hunting, promotes cooperation between men and women and high levels of male parental investment, because it favors sex-specific
specialization in embodied capital investments and generates a complementarity between male and female inputs. The economic and reproductive cooperation between men and women facilitates provisioning of juveniles, which both bankrolls their embodied capital investments and acts to lower mortality during the juvenile and early adult periods. Cooperation between males and females also allows women to allocate more time to child care and improves nutritional status, increasing both survival and reproductive rates. The nutritional dependence of multiple young of different ages favors sequential mating with the same individual, since it reduces conflicts between men and women over the allocation of food. Finally, large packages also appear to promote interfamilial food sharing. Food sharing assists recovery in times of illness and reduces the risk of food shortfalls due to both the vagaries of foraging luck and the variance in family size due to stochastic mortality and fertility. These buffers against mortality also favor a longer juvenile period and higher investment in other mechanisms to increase life span.
Thus, we propose that the long human life span, lengthening of the juvenile period, increased brain capacities for information processing and storage, intergenerational resource flows, and cooperative biparental investment in offspring coevolved in response to this dietary shift and the new production processes it entailed.
It is not yet possible to know many vital statistics and behavioral characteristics from paleontological and archeological remains. It must be recognized that modern hunter-gatherers are not living replicas of our Stone Age past, and global socioeconomic forces affect them all. Furthermore, many foragers today live in marginalized habitats that underreward male hunting efforts. Yet despite the variable historical, ecological, and political conditions affecting them, there is remarkable similarity among foraging peoples, and even the variation often makes adaptive sense. Comparisons between foraging peoples and other modern primates are an important source of information about the life histories of our ancestors and the selection pressures acting on them, the subject of the next sections.
Mortality and Production
The age-specific mortality profile among chimpanzees is relatively V-shaped, decreasing rapidly after infancy to its lowest point (about 3 percent per year) at about age 13, the age of first reproduction for females, and increasing sharply thereafter. In contrast, mortality among human foragers decreases to a much lower point (about 0.5 percent per year) and remains low with no increase between about 15 and 40 years of age. Mortality then increases slowly, until there is a very rapid rise in the 60s and 70s. The pattern is much more block U-shaped. The strong similarities in the mortal-
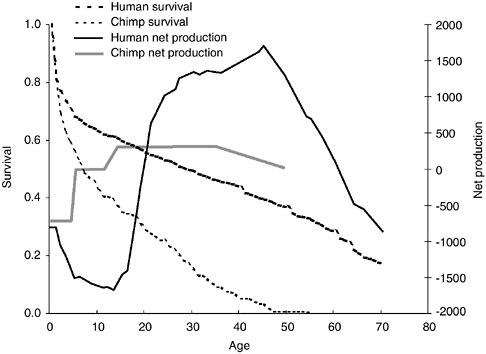
FIGURE 7-3 Net food production and survival: Human foragers and chimpanzees.
ity profiles of the foraging populations suggest that this pattern is an evolved life history characteristic of our species.
Figure 7-3 illustrates the differences between human foragers and wild-living chimpanzees. As a result of these differences in mortality pattern, hunter-gatherer children experience higher survival (60 vs. 35 percent) to age of first reproduction, at about ages 13 and 19 for chimpanzees and foragers, respectively. Chimpanzees have a much shorter adult life span than humans as well. At first reproduction, chimpanzee life expectancy is an additional 15 years, compared to 38 more years among human foragers. Importantly, women spend more than a third of their adult life in a postreproductive phase, whereas very few chimpanzee females survive to reach that phase. Fewer than 10 percent of chimpanzees survive to age 40, but more than 15 percent of hunter-gatherers survive to age 70.
Age profiles of net food production (food produced minus food consumed) also differ dramatically. Among chimpanzees, net production before age 5 is negative, representing complete, and then partial dependence on mother’s milk. The second phase is independent juvenile growth, lasting until adulthood, during which net production is zero. The third phase is reproductive, during which females produce a surplus of calories that they
allocate to nursing. Humans, in contrast, produce less than they consume for some 15 to 22 years, depending on the group. These same patterns appear to hold true even among subsistence agriculturalists (Kramer, 2002). Net production becomes increasingly negative until about age 14 and then begins to climb. Net production in adult humans is much higher than in chimpanzees and peaks at about 35 to 45 years of age. This peak is about five times as high as the chimpanzee peak. The human age profile of production could not exist if humans had the same mortality profile as chimpanzees (Kaplan and Robson, 2002; Robson and Kaplan, 2003). Only 30 percent of chimpanzees ever born reach the age when humans produce what they consume on average, and less than 10 percent reach the age when human production peaks.
Role of Men in Human Reproduction
Unlike most other mammals, men in foraging societies provide the majority of the energy necessary to support reproduction. After subtracting their own consumption from total production, women supply an average of 3 percent of the calories to offspring, with men providing the remaining 97 percent among the 10 foraging societies for which quantitative data on adult food production are available (Kaplan et al., 2001a). Hunting, as opposed to gathering of animal protein in small packets, is largely incompatible with the evolved commitment among primate females toward intensive mothering, carrying of infants, and lactation on demand in service of high infant survival rates. First, it is often risky, involving rapid travel and encounters with dangerous prey. Second, it is often most efficiently practiced over relatively long periods of time rather than in short stretches, due to search and travel costs. Third, it is extremely skill intensive, with improvements in return rate occurring over two decades of daily hunting. The first two qualities make hunting a high-cost activity for pregnant and lactating females. The third quality, in interaction with the first and second, generates life course effects such that gathering is a better option for females, even when they are not lactating, and hunting is a better option for males (Kaplan et al., 2001b). Since women spend about 75 percent of their time either nursing or more than 3 months’ pregnant during their reproductive lives, it would not pay to hunt; therefore they never get enough practice to make it worthwhile, even when they are not nursing or pregnant or are postreproductive.
The fact that humans are unique in raising multiple dependent offspring of different ages also reduces the payoffs to defection and increases the benefits for men and women to link their economic and reproductive lives over the long run. Men and women who divorce and remarry during the time they are raising offspring will face conflicts of interest with new
spouses over the division of resources. If they marry someone with children from previous marriages, they may disagree with their new spouses over the allocation of food and care to their joint children, relative to children from the previous marriage. Those conflicts increase the benefits of spouses staying together and having all or most of their children together.
Human females evidence physiological and behavioral adaptations that are consistent with an evolutionary history involving extensive male parental investment. They both decrease metabolic rates and store fat during pregnancy, suggesting that they lower work effort and are being provisioned (Ellison, 2001a; Lawrence and Whitehead, 1988; Pike, 1999; Poppitt et al., 1993). During lactation, women in foraging societies decrease work effort and focus on high-quality care (Hurtado et al., 1985; Lancaster et al., 2000). In contrast, nonhuman primate females do not store appreciable fat, and they increase work effort during lactation; as a result, they have increased risk of mortality (Lancaster et al., 2000). The human specialization could not have evolved if women did not depend on men for most of their food provisioning throughout human history.
Extensive cooperation among human men and women would only make sense if the reproductive performance of spouses were linked. When women reach menopause in their late 40s, men have the option to continue reproducing with younger women but generally do not do so. Among the Ache, for example, 83 percent of all last births to women also represent a last birth for the fathers of these children (Kaplan et al., 2001b).
The importance of male provisioning interacts with free access to the means of production, resulting in relatively low levels of polygyny. Although there are often territorial boundaries and disputes among neighboring groups, there is generally open access to foraging sites. Production is therefore determined by work effort and ability and is less variable as a result (although differences due to ability can be significant). The comparatively low variance in adult men’s food production, coupled with the importance of male provisioning, may be why monogamy is the predominant marriage form among foragers. Figure 7-4 (using cross-cultural data from Binford, 2001) shows that the modal percentage of marriages that are polygynous among foragers is about 3 percent, with the major outliers coming from Australia (see below).
Flexibility in Fertility, Parenting, and Mating
Traditionally, demographers have attempted to understand the onset and termination of reproduction and birth intervals in natural fertility regimes in terms of proximate determinants (Bongaarts and Porter, 1983; Wood, 1994). For the most part, those determinants are treated as givens, and there has been little consideration of the causal processes shaping them.
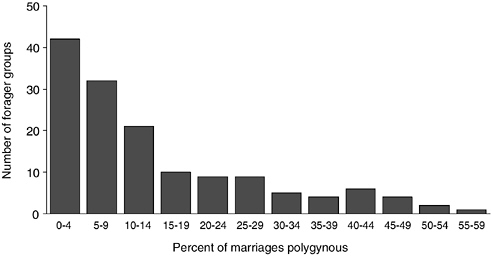
FIGURE 7-4 Frequency distribution of percentage of marriages that are polygynous among foragers.
In contrast, evolutionary demographers approach those determinants in terms of design and ask why the physiological, psychological, and cultural processes that regulate fecundity take the forms they do. The three trade-offs discussed above (current vs. future reproduction, quantity vs. quality of offspring, and mating vs. parenting effort) have organized research into those design features.
There is mounting evidence that human reproductive physiology is particularly specialized toward the production of high-quality, large-brained offspring. Two implications of this specialization are rigid control over embryo quality and a series of adaptations on the part of both mother and offspring designed to ensure an adequate energy supply to the nutrient-hungry, fast-growing brain. Given the massive investment in human offspring, this system ensures that investment is quickly terminated in an offspring of poor genetic quality. Fetal growth is more rapid in humans than in gorillas and chimpanzees, and both mother and offspring store exceptional amounts of fat (Kuzawa, 1998), probably to support an equally exceptional rate of extensive brain growth during the first five years of life (Ellison, 2001a, 2001b).
The physiological regulation of ovulation, fertilization, implantation, and maintenance of a pregnancy is highly responsive to energy stores in the form of fat, energy balance (calories consumed minus calories expended), and energy flux (rate of energy turnover per unit of time). Low body fat, weight loss due to negative energy balance, and extreme energy flux (either very low intake and very low expenditure, or very high intake and very high
expenditure) each lower monthly probabilities of conceiving a child that will survive to birth. Seasonal variations in workloads and diet have been shown to affect female fecundity and fertility (Bailey et al., 1992; Hurtado and Hill, 1990; Prentice and Whitehead, 1987). Variation across groups in both age of menarche and fecundity has been linked to differences in food intake and workloads.
Behavior and the underlying psychological processes that govern parental investment affect fertility indirectly via maternal physiology. One route is through breast-feeding. Patterns of breast-feeding and solid food supplementation vary both cross-culturally and among mother-infant pairs (McDade, 2001; Sellen and Smay, 2001).
The second route is due to the additional energetic constraints involved in provisioning children. One fundamental trade-off with respect to activity profiles is between present and future production (Bird and Bliege Bird, 2002; Bliege Bird and Bird, 2002; Bock, 2002; Kaplan, 1996; Kramer, 2002). If children engage in easy-to-perform foraging activities, such as fruit collection, this will provide immediate caloric benefits and lower the parental subsidy required. However, time allocation to those activities detracts from time spent learning more difficult activities, such as hunting and extractive foraging. During the learning phase, those activities produce very little food but generate high rates of production in the future. Another fundamental trade-off is between production and mortality risks, since foraging exposes people to predation, accidents, and getting lost. This suggests that parental psychology should be responsive to age-specific mortality rates and how they are affected by alternative activity profiles and on their short- and long-term consequences for production/productivity.
There is a growing body of evidence to support the view that people in foraging societies are sensitive to these trade-offs and adjust their behavior accordingly. A seminal series of papers by Blurton Jones and colleagues (1994a, 1994b; Hawkes et al., 1995) shows that !Kung hunter-gatherer parents consciously assess foraging risks to children and that differences between !Kung and Hadza children’s productivity are due to features of the local ecology related to ease of resource extraction and dangers associated with productive tasks. A study of foragers/agropastoralists in Botswana showed that the age profiles of children’s activity budgets reflected immediate skill and strength requirements as well as opportunities to learn (Bock, 1995). In all foraging societies for which data are available, people are aware of the many years of learning required to become a competent hunter, and young men are provisioned while they learn those skills (Kaplan et al., 2000).
These time allocation decisions affect fertility through maternal energetics. The connection between psychology and fertility is indirect in that psychology adjusts parental investment (through productive behavior, wealth flows, and breast-feeding) and reproductive physiology translates
parental investment decisions into fertility. The key to this system is that maximizing lifetime-expected resource production through optimal allocation of activities and wealth flows will tend also to maximize fitness when all wealth is in the form of food and extra food translates into higher fertility.
There have been several empirical applications of optimality models, designed to determine whether the onset and termination of reproduction and the size of interbirth intervals actually maximize fitness. The results of those analyses have been mixed. Blurton Jones et al. (1989) analyzed the relationship of the birth interval following a child who survived until the birth of its next sibling to the survival of either member of the sibling pair among the !Kung (Jo’hansi) in order to determine whether the observed mean interval of 48 months maximized the total number of surviving offspring. This analysis showed that 48-month birth intervals were, in fact, optimal (see Harpending, 1994; Pennington, 2001, for a critique).
In contrast, similar analyses among the Ache showed that observed birth intervals were longer than optimal (Hill and Hurtado, 1996). Phenotypic covariation, however, poses a major problem in those analyses. If healthier women have shorter birth intervals than less healthy women because they have larger effective energy resources, the estimated effect of birth intervals on offspring survival would be downwardly biased. In fact, studies examining natural variation among nonhuman organisms often show a positive, rather than a negative, relationship between fertility rate and offspring survival for the same reason (Partridge and Harvey, 1985). When fertility rate is experimentally manipulated among those same organisms, the relationship is reversed, as would be expected by a quantity-quality trade-off. Consistent with the self-selection hypothesis, Gambian women who had higher hemoglobin levels following the birth of a child exhibited both shorter interbirth intervals and higher child survival (Sear et al., 2001). Thus, it may be that women’s physiology tracks its own condition in such a way as to maximize their individual fitness.
Hill and Hurtado (1996) also examined age of first reproduction and menopause to test optimality models. They found that women began reproducing at the optimal weight to maximize their lifetime fitness. However, they did not find support for the proposition that menopause maximizes fitness, as expected by the “grandmother hypothesis” (Williams, 1957). The effects of grandmothers on the fertility of their children and on the survival of their grandchildren were not large enough to overcome the fitness costs of reproductive cessation. In a parallel “contrary-to-prediction” analysis, Rogers (1993) found that the expected reproductive success of older women would have to be implausibly low to favor menopause. However, as Hill and Hurtado (1991, 1996) point out, self-selection also complicates this analysis. Grandmother’s death is used to assess the impacts
TABLE 7-1 Regressions for Percent of Families That Are Polygynous Among Foragers
of older women on the fitness of their children and grandchildren, but in kin-based societies other relatives probably compensate for the death of a kinswoman, producing another downward bias in effect estimation.
Finally, there is evidence that mating behavior responds to ecological variability in subsistence ecology, associated with complementarity and variance in access. Using the same cross-cultural data set as in Figure 7-4, a regression of the percentage of marriages that are polygynous on the percentage of the diet derived from gathered foods and the size of mobile foraging groups suggests that increased reliance on female contributions is associated with higher levels of polygyny (see Table 7-1). The one exceptional area, where polygyny is much greater than average, is Australia. There high rates of polygyny are found in the northern territories and Queensland, with the most extreme case being the Tiwi (51 percent of marriages being polygynous). In a sense they are the exception that proves the rule because, according to ethnographic sources (Goodale, 1971; Hart and Pilling, 1960), Tiwi women are actively involved in hunting and fishing. In that coastal environment there are small game and fishing opportunities that are more like gathering than hunting (e.g., catching small, sleeping animals) and women can be more economically self-sufficient.
The next section treats major historical trends in reproduction, parenting, and mating practices associated with the domestication of plants and animals.
REPRODUCTIVE BEHAVIOR SINCE THE DOMESTICATION OF PLANTS AND ANIMALS
Sedentism and Tribal Horticulture
The Socioecological Context
Village sedentism and the domestication of plants had a profound, yet limited, impact on human socioecology. Subsistence based on horticulture
rests on land-extensive, slash-and-burn agriculture on prime resource patches, access to which was maintained by the social group and defended by males against outsiders. Within the group, access is on the basis of a usufruct system of land tenure that gives all group members direct rights to the means of production and reproduction (Boserup, 1970; Goody, 1976). People live in small villages, larger than hunter-gatherer bands but similarly scaled in terms of face-to-face, kinship-laden interactions.
There is evidence that sedentism brings a reduction in child mortality compared to hunter-gatherers, as well as higher female fertility, although it is unclear whether the strongest effects are in reduced birth spacing or higher rates of child survival (Bentley et al., 2001). The potential of deaths from chronic intergroup warfare and raiding increases, particularly affecting males in which the percentage mobilized often reaches 35 to 40 percent and male deaths from 10 to nearly 60 percent (Keeley, 1996). Complementarity in the male and female division of labor is complex because of its link to local ecology. Garden production by women using the dibble and hoe provides the carbohydrate and caloric base of the diet and is easily combined with child care (Boserup, 1970; Goody, 1976). Males contribute their labor in clearing fields, in animal protein through hunting and fishing, and in protection of the village resource base through defense.
The relative contribution and imperative of male help vary by ecological context. For example, female gardening of high-protein crops on riverine alluvial soils, such as millet and sorghum in much of village Africa (Colson, 1960; Lancaster, 1981), is very different from subsistence based on manioc in the thin soils of South America. There male hunting is critical to balanced macronutrients in the diet and frequent clearing is necessary. Similarly, the critical need for defense of the village resource base is supplied by males as an umbrella benefit rather than to specific wives. More recently, the benefit of such defense was much reduced under colonial suppression of warfare and raiding. However, since neither males nor females produce beyond subsistence needs and the means of production are held in common through usufruct, there is little opportunity for major variance in resource holding. However, variance in male reproductive success does arise on the basis of their success in raiding, which brings certain males both high status and more wives (Chagnon, 1979).
Mating Patterns, Parental Investment, and Reproduction
Reproduction in horticultural societies is associated with near-universal marriage for both sexes, with reproduction beginning at sexual maturity for women and extending through the entire period of fecundity. Reproduction for men is somewhat delayed due to the need to access wives through either bride service (local group) or bride capture (outside group),
the first being a personal cost in labor contributed to the bride’s family and the second a personal cost in terms of risk. However, polygyny extends the male reproductive period as new and younger wives are added through time.
The high frequency of polygynous marriages associated with horticulture may be because each wife is essentially able to support herself and her children through her own labor (Lancaster and Kaplan, 1992; Murdock, 1967). Males do not have to ponder whether they can afford additional wives and children, only how they can get and keep them. As White and Burton (1988) found, polygyny is most associated with fraternal interest groups, warfare for the capture of women, absence of constraints on expansion into new lands, especially for horticulturalists, environmental quality, and homogeneity. The practice of widow inheritance by husband’s kin also increases the frequency of polygyny (Kirwin, 1979). The form, sororal polygyny, is at its highest frequency due to the ease of sisters forming collaborative horticultural work groups (Irons, 1979b).
Parental investment in horticultural societies focuses on raising healthy children without concern for their marriage market endowments or inheritance of resources. Birth into a social group provides all the inheritance a child would need to access the means of production and reproduction. Such concepts as bastardy or disinheritance do not play a formal role in family dynamics. Child labor is valuable to families since horticulture provides a number of relatively low-skilled tasks that older children can perform. In fact, Kramer (2002) demonstrated that among Maya horticulturalists older children contribute at the level of “helpers-at-the-nest,” significantly increasing their parents’ fertility and without whose help their parents could not add further offspring to the family.
Variance in reproductive success is relatively low for women because marriage is universal, and female fertility and fecundity depend on their own productivity and work effort (Ellison, 2001a; Jasienska, 2000; Prentice and Whitehead, 1987). Greater variance among men was possible on the basis of raiding and bride capture, but the social system itself is not stratified and individual men cannot amass or control access to resources relative to other men or pass them on to their sons.
Extrasomatic Wealth and Tribal Pastoralism
The Socioecological Context
The domestication of animals, particularly large herd animals such as cattle, camels, and horses, proved to have a profound effect on human social and reproductive patterns. Large domesticated livestock have intrinsic qualities that affected human social relationships, marriage patterns,
and investment in children. For the first time in human history, men could control a form of extrasomatic wealth that could be held by individuals, thus increasing the variance in male quality based on the resources each could control.
Second, herds are the basis of the domestic economy through their products of meat, milk, and hides. There are advantages to dependence on such a resource supply: (1) improvements in diets rich in animal protein, (2) stability of diets since animals are stored hedges against fluctuations in annual or seasonal climatic effects, and (3) flexibility due to the divisibility of herds into smaller units that can be moved about the landscape on the basis of the richness and concentration of local resources. This improvement in diet may result in higher fertility and survivorship of women and children compared to foraging and horticulture but also higher mortality for males due to endemic conflict. Generally, however, pastoralists show a wide range in fertility levels due to the many different kinds of pastoralism with respect to agricultural supplementation, disease risk, mobility, and so forth (Borgerhoff Mulder, 1992; Sellen and Mace, 1997). Large-animal herding demands a high degree of complementarity between female processing and child care and male risk taking in herd management and defense. The products of herds require intensive processing of meat, milk and hides, labor provided by women. In contrast, the very existence of extrasomatic wealth in large stock in such a readily divisible and moveable form (as opposed to agricultural land) puts a high premium on males as defenders and raiders. We find the warrior complex full blown, with chronic internal warfare, blood feuds, the social segregation of a male warrior age class, fraternal interest groups, a geographic flow of women from subordinate to dominant groups through bride capture, and expansionist segmentary lineages based on the male line (DiVale and Harris, 1976; Low, 2000; White and Burton, 1988). Men with strong social alliances are more likely to find at least some of their wives from within their own social groups, whereas men from small or subordinate lineages are willing to take more risks in lieu of performing bride service (Chagnon, 1988, 2000; Lancaster, 1981).
The original distinction made by Orians (1969) between resource defense polygyny and harem defense polygyny is relevant here. The chronic warfare of pastoralists (Keeley, 1996; Manson and Wrangham, 1991; White and Burton, 1988) can be understood as resource defense polygyny, as opposed to the harem defense polygyny described earlier for horticultural societies. Both types of societies raid to capture women, but pastoralists also raid to capture resources that can be used to acquire and maintain new wives and their children. In a study of 75 traditional societies, the principal cause of warfare was either women (45 percent of cases) or material resources for obtaining a bride (39 percent of the cases), particularly in
pastoral societies where bride wealth must be paid (Manson and Wrangham, 1991).
Mating Patterns, Parental Investment, and Reproduction
Resource defense polygyny means that males will compete to control the resources that females must have for successful reproduction. A male’s ability to successfully control more resources translates directly into more wives and children (Borgerhoff Mulder, 1989, 1991, 1995). One extraordinary result of extrasomatic wealth, particularly readily partible wealth, is the institution of a new pawn on the marriage market table—bride wealth. Women and their families come to marriage negotiations with their traditional offers (youth, health, fecundity, and female labor). Men, however, now have to come up with a significant payment of resources in the form of bride wealth as a preferred substitute for bride service.
Bride wealth among pastoralists consists of horses, cattle, or camels, with sheep or goats as supplements or lower-valued substitutes. Among African pastoralists the close male kin of the groom helps him with his first bride wealth payment, but the acquisition of subsequent wives is his own responsibility. Livestock used for bride wealth has interesting attributes: (1) it creates conflicts of interest between fathers and sons and among brothers for its use to obtain a bride (Borgerhoff Mulder, 1988), (2) men from poor families will be more willing to take risks to obtain bride wealth or brides though capture (see Dunbar’s 1991 discussion of war chiefs and peace chiefs among the Cheyenne), and (3) livestock can be inherited.
Investment in children takes a novel form under a pastoralist system. The payment of bride wealth improves health and survivorship among young girls because their marriages bring in resources that can be used by their fathers and brothers to acquire more wives (Borgerhoff Mulder, 1998). Sub-Saharan Africa is notable for the fact that despite the patrilineal bias in so many societies, neither a survival nor a nutritional advantage is found for boys over girls (Svedberg, 1990). Furthermore, among the Kipsigis, who are agropastoralists, early-maturing (and presumably better fed and healthier) women have higher lifetime reproductive success than late-maturing women, command higher bride wealth, and hence constitute a higher return on parental investment in their upbringing (Borgerhoff Mulder, 1989). They also represent a better investment for a husband’s bride wealth payment because of a higher return in fertility.
Furthermore, children are able to provide child care of younger siblings as well as low-skilled labor in stock care and the processing of animal products, so they are able to substantially but not completely offset the costs of their rearing (Bock, 1995, 2002). However, this is countered by the fact that the parents of sons now have a new cost to meet, the balloon
payment (bride wealth) needed at the end of their sons’ development and perhaps equal to the summed previous investment to establish sons on the marriage market. The flow of stock through families that are both bride wealth receivers and givers helps maintain the system at the same time that it creates problems for families with unfavorable ratios of sons to daughters (Borgerhoff Mulder, 1998).
Finally and most significantly, there is suggestive evidence that for the first time humans began to reproduce at levels that may not maximize the number of descendents with the appearance of extrasomatic wealth and its inheritance. Men appear to marry fewer wives than they could afford in the interests of providing each child with a greater endowment. In other words, male pastoralists may pit quality against quantity of children to preserve a lineage status and resource base rather than simply maximizing the number of descendents (Borgerhoff Mulder, 2000; Mace, 2000).
Social Stratification in the PreModern Era: States and Despotism
The Socioecological Context
The rise of civilizations, beginning about 6,000 years ago in Mesopotamia and occurring at different times and places (e.g., Egypt in the Near East, the Aztec and Inca in the Americas, and India and China in Asia) marked a critical shift in how humans organized themselves in social systems and in relation to the environment (Betzig, 1993; Goody, 1976). These civilizations appear to have developed independently in response to local conditions without being the products of either conquest or diffusion. Despite this historical independence, they evidence significant similarities: (1) the presence of large stratified social groupings settled on particularly large and productive resource patches and (2) the appearance of social despots, men who use coercive political power to defend their wealth and reproduction and warfare to acquire more resource patches and slaves (Betzig, 1986). These two major effects flow from the nature of the resource patches.
The patches on which the first civilizations were settled had special qualities: (1) they were highly productive but set in environments where there was a rapid falloff to unproductive lands such as desert or forest and (2) these productive patches could not be intensively utilized without complex political organization as in regional irrigation systems. Political control and organization rested on the power of men. Although female nonhuman primates often form alliances with their female kin to protect and control access to the resources necessary for their reproduction (Isbell, 1991; Sterck et al., 1997), the reproductive benefits of resources are much greater for men than for women because of their impacts on polygyny. The
end result of these environmental conditions associated with early social stratification was that men competed for control of the resources necessary for reproduction and formed despotic hierarchies involving social alliances and stratification, with low-status men “agreeing” to live under political despotism because they could not readily move to another resource base.
The increased reliability of food resources, the costs of warfare, and the concentrations of large populations into small urban areas each had impacts on mortality and morbidity. A cross-cultural analysis of fertility and mode of subsistence found that, for a 10 percent increase in dependence on agriculture between “sister” cultures, there is a fertility increase of approximately 0.2 live births per woman (Sellen and Mace, 1997). Bentley et al. (2001), in reviewing the cross-cultural and archeological evidence, suggested a series of multidirectional effects: higher fertility due to more consistent food supply and earlier maturation; increased infectious diseases with regular visitations as well as endemic diseases, such as malaria and tuberculosis, due to long-distance trade and large urban populations; and a shift in peak mortality from infancy to middle childhood. Furthermore, warfare continues to reduce the numbers of young men in the mate pool.
With social stratification comes a complex division of labor with specialists in war, farming, crafts for the production of goods and services, and war captives and slaves for the hardest manual labor, as well as long-distance trade in luxury goods and slaves. The introduction of the plow in Eurasia and the need for food production beyond simple subsistence to service the needs of urban markets led to significant changes in the division of labor (Ember, 1983; Goody, 1976) and extremely high complementarity between male labor and resource acquisition and female labor and child care. There is evidence of increased workloads for women despite the fact that men assume more responsibility for farm labor, because of increased demands for women to process grains or secondary animal products such as milk, hides, and wool (Bentley et al., 2001). They cite evidence that women’s physical activity levels as measured by PALs (the multiple of resting basal metabolic rate required for activity) among hunter-gatherers and horticulturalists are very similar, with means of 1.72 and 1.79, respectively, but that with agriculture it increases substantially to 2.31.
Variance in male resource holding was probably the greatest it has ever been in human history (Betzig, 1986, 1992a, 1992b, 1993). The reason for this is that despotic males had enormous political and social control with the ability to eliminate rivals and their entire families through despotic edicts, to wage war to increase personal and state resource bases, to acquire slaves and war captives for labor and reproduction, and to determine political succession for favored sons. This extreme variance in male resource holding inevitably produces political instability due to the creation of too many potential heirs (sons of many wives) and too many males (slaves)
without access to the means of reproduction. The great wealth to be gained from domination also motivated expansion and intergroup conflict among would-be despots.
Finally, a notable characteristic of the premodern period in many parts of the world is evidence for a growing rural population resulting from higher fertility and associated growing concerns regarding saturation of the resource environment. This is often associated with urban growth, empire building and colonial expansionism, providing opportunities for migration by noninheriting or low-status children to areas of both higher mortality and risk but also with the potential for the acquisition of employment, land, or wealth and power. It also generated a great deal of concern about keeping the family estate intact and about the management of inheritance.
Mating Patterns, Parental Investment, and Reproduction
Despotic males are an extreme example of resource defense polygyny (Orians, 1969); that is, as individuals they control access to the resource base for reproduction that females require and, with few competitors, polygynous marriages to them become the only family formation strategy option for many women. The mating markets of despotic systems are characterized by historic extremes in male variance in resource holding and power. Table 7-2 presents data on the numbers of wives and children of despots in some of these early civilizations. As Betzig (1993) notes, the extreme sizes of royal harems ranging from 4,000 to 16,000 women in these examples are associated with smaller but still impressive numbers of wives and concubines for the royal supporters. In the case of the Inca, the size of a man’s harem was regulated by law and was in direct relationship to his social/political rank. Among the Inca there were nine levels of political rankings with polygyny ceilings for each except the top-most. These harems were exclusive holdings of large numbers of young fecund women with their children, and access to them was restricted to their mate and regulated
TABLE 7-2 The Wives of Despots
|
Country |
Century |
Royal Harems |
Reign |
|
India |
5th B.C. |
16,000 |
Udayama |
|
|
15th A.D. |
12,000 |
Deva Raya II |
|
|
16th A.D. |
4,000-12,000 |
|
|
China |
618-907 A.D. |
10,000 |
|
|
Inca Peru |
early 16th A.D. |
1,500 per province |
|
|
Aztec Mexico |
early 16th A.D. |
4,000 |
|
|
SOURCE: Betzig (1993). |
|||
with some sophistication to optimize female fertility. Many of these wives and concubines were collected as tribute or war booty, but others, as principal wives, probably represented important political alliances with their male relatives.
There are two clear outcomes of such variance. The first is that many men remain unmated or have only one wife, so male celibacy or at least nonmarital sex is prominent. In the words of Dickemann, polygyny in the context of extreme social stratification is “characterized not only by arbitrary sexual rights of lords and rulers but by large numbers of masculine floaters and promiscuous semi-floaters, beggars, bandits, outlaws, kidnappers, militia, and resentful slaves and serfs.” (Dickemann, 1981:427). Nevertheless, these early despotic states lasted for thousands of years. A second outcome of variance in male resource holding and male mating success is that there tends to be universal marriage for women, with only those most severely compromised by health or other personal qualities being unlikely to find a role as secondary wife or minor union. For access to the mating market, men must bring wealth, power, and land in order to be favorably placed or else get wives as high-risk booty in state warfare (Clarke and Low, 2001; Low, 2000).
Women, too, bring their traditional qualities of youth, health, and fecundity along with their labor. However, there is a historic shift in how women and their families approach the mating market that has been richly described in a series of papers on hypergyny, dowry, female infanticide, and paternity confidence (Dickemann, 1979a, 1979b, 1981). The extreme variance in male quality created by despotism and harem polygyny forces the families of women to put down more and more value on the mate market table to access a desirable groom or to move a daughter up in the social hierarchy. These extra payments form an area of female-female competition that includes actual wealth, in the form of dowry, and guarantees of paternity confidence (bridal virginity and wifely chastity; (Gaulin and Boster, 1990). Guarantees of a daughter’s virginity and chastity (a prerequisite for a bride destined to produce heirs to a male lineage holding a reproductive estate) are costly, involving female seclusion (special women’s quarters, harem guards, chaperones) and female incapacitation (e.g., foot binding and corseting) that bars their daughters from the outside world of productive labor.
Endowment of Children with Access to Reproductive Estates
Parental concern over the ability of their children to access reproductive estates transforms the nature of the marriage market. Parental investment in these systems varies in relation to the power and wealth of the male’s family. As is to be expected, under such conditions where male
access to and control of resources is the basis of social stratification, patrilineal descent and patrilocal residence are highly favored since males are the principal resource holders (Hartung, 1982). Resource-holding parents commit to a balloon payment, very large and late in the period of parental investment, to launch their children in marriage. This balloon payment takes the form of endowments, promised inheritance, and dowry as anticipatory inheritance for daughters (Dickemann, 1979a, 1979b; Goody, 1973, 1976). For resource-holding families, then, the marriage market formed by stratified social systems proves costly in terms of parental investment and forces a focus on endowments for both sexes at the age of marriage. Poor parents, on the other hand, attempt to balance labor demands with fertility, since in agricultural systems children can be productive at low-skill tasks or child care and thus add to the family economy. They might try to regulate birth spacing to optimize the productivity of already-born children before another mouth to feed is added to the family.
Historic Trends in the Reduction of Claimants to the Family Estate
With population growth and increased saturation of arable lands, parents adopt patterns of restricted and differential inheritance in order to keep the family estate intact and maintain the concentration of wealth or, in the case of the poor, to balance food supply with family size. Boswell (1990) and Hrdy (1994) provide evidence from classical Europe of attempts by parents to limit the burden of excess children through abandonment. However, well-to-do families also practice abandonment in an attempt to limit the number of potential heirs to political succession and inheritance, as evidenced by the placement of secret identifying markers so that a child might be retrieved later should its older same-sex sibling die.
This trend is particularly well documented in premodern Europe. Human evolutionary ecologists in collaboration with historical demographers have provided a unique record of the relationships between fertility, family formation strategies, and the socioecological context during the premodern and early modern periods of European history (see review in Voland, 2000)). Their studies, based on heraldic or parish records of births, marriages, deaths, and inheritance of estates, can be used to directly link reproductive strategies with resource holdings. This time period witnessed developments that had began centuries earlier but occurred without the benefit of quantifiable documentation. Boone, for example, traces the historical process of parental investment among Portuguese elites during the medieval/early modern periods of the 15th and 16th centuries (Boone, 1986, 1988). Saturation and resource stress are evident with a progressive narrowing of the numbers of claimants to an inheritance, first through monogamy and bastardy creating a single bloodline of inheritors (Goody, 1976, 1983), fol-
lowed by a preference for sons over daughters as inheritors, and finally by birth-order effects with preference for primogeniture within each sex for access to resources and the creation of celibate children to live as priests, nuns, bachelors, and spinsters (Hrdy and Judge, 1993). For the first time in human history, mating and reproduction are no longer universal for women. With survival through childhood and young adulthood still quite problematic, ancillary practices develop in which both sons and daughters would be held in reserve in monasteries and nunneries for inheritance should their older same-sex sibling die before reproduction (Boone, 1986, 1988; Goody, 1976, 1983). Within the scope of these restrictions that limit half-sibling and sibling competition, parents with wealth raise as many children as they can but endow a select number at adulthood.
During most of this historical period there is a strong correlation between wealth, probability of marriage, younger age at marriage, and completed fertility (Voland, 2000). However, restricted inheritance decreases the reproductive benefits of polygyny. The desire to concentrate wealth also limits the reproductive success of noninheriting sons and daughters. This may be another example in which reproductive and parental investment behavior in response to extrasomatic wealth results in outcomes that do not maximize parental fitness. In fact, toward the end of the period, as life expectancy improved and economic structures became saturated, resource holding groups delay marriage into the late 30s and early 40s for men and mid-20s for women (Szreter and Garrett, 2000; Voland, 2000).
Family reconstruction studies document very different reproductive strategies according to class.3 Generally, wealth brings higher probability of marriage at a younger age, to a younger spouse, and more children. However, as environments become more saturated, local resource competition among siblings differentially affects resource-holding families, as opposed to day laborers, and increases the likelihood of dispersal of later-born children (Clarke and Low, 1992; Towner, 1999, 2001; Voland and Dunbar, 1997). With saturation the benefits to resource holders of having an above-average number of children is offset by more and more intense sibling competition for access to inheritance (Voland, 2000). Parents without resources have no need to manipulate their offspring and are more likely to benefit from opportunistic strategies by their children (Voland and Dunbar, 1995). An extreme form of such parental manipulation of offspring marital
opportunities is polyandry in which a male sibship jointly marries a single woman in order to avoid division of property and labor among competing households of wives and children of brothers (Crook and Crook, 1988; Haddix, 2001). Wet-nursing during this period presents a fascinating example of how differentiation in parental investment strategies develops into extreme forms for both the highest and the lowest status groups of women. Throughout human history there has always existed a conflict between production and reproduction for women, a conflict that in fact troubles female mammals in general. Human women are especially caught in this conflict because they have multiple dependent young of differing ages and needs (Draper, 1992), which means that a true respite never occurs until all children are reared. Cross-culturally women’s work is organized by its compatibility with child care (Brown, 1970); however, this compatibility is never complete—only more or less so (Hurtado and Hill, 1990; Lancaster, 1997; Lancaster et al., 2000). During this period of social stratification some women, for the first time, are able to subvert the physiological capacity of lactation of other women to serve their own reproductive ends. Since intense breast-feeding lowers the likelihood of ovulation, wet-nurses, even if paid, are sacrificing their own fecundity to another’s (Hrdy, 1994, 1999).
Typically high-status women do not breast-feed their own children but use wet-nurses. This increases the fertility of high-status women, whose main function is to produce heirs, to a nearly annual birth rate (among the highest for any group of women in human history). In contrast, the birth spacing for their wet-nurses is closer to 4 years (Hrdy, 1994). A second group of women also used wet-nurses, especially towards the end of this historic period. These were single women working in urban centers or the wives of poor tradesman who found themselves in positions of servitude or trade where the incompatibility between breast-feeding and work was complete. To great detriment to their infant’s survival, these women placed their children with commercial wet-nurses. In these cases the demands of maternal work far outweighed the needs of infant growth, perhaps to further the interests of weaned older children.
Modern Labor Markets
The Ecological Context
Given rural reproductive and survival rates, the restricted inheritance system discussed above produces excess adults without access to land and the means of production. Colonization through conquest was one response by males, especially later-born sons (Boone, 1986; Curtin, 1989) to this situation. Another response by both men and women was to provide services for others and migration to cities in search of employment. This
supply of labor and of consumers helped fuel the growth of a mercantile economy that was to gradually supplant the power- and land-based hierarchies of the pre-modern period.
The directional change in the nature of labor markets toward greater wage premiums for skill- and education-based capital over the last two centuries is well documented (e.g., Burck, 1976; Herrnstein and Murray, 1994; Newcomer, 1955; Vinovskis, 1994). The relationship between education and income increased considerably during the second half of the 20th century. For example, in the United States, real wages actually dropped from 1958 to 1990 among men without high school degrees. In 1958, men with a graduate education earned about 2.3 times as much as men with an elementary school education; by 1990 they earned more than 3.5 times as much. Wage differentials among men with some college education, bachelor’s degrees, and graduate degrees also increased substantially. For women, wage differentials by education levels increased substantially in the 1980s (Kaplan et al., 2002).
As the extent of the labor and consumer markets grew, along with advances in production technologies, there was a concomitant increase in both private and public investments in education. In a sense, the relationship between embodied capital and production in modern skills-based labor markets is more similar to the foraging way of life than to its agricultural predecessor. Rather than generating wealth through control of land, people invest in learning to increase productivity, and individuals are free to move through the environment in search of economic opportunities.
These increases in educational capital investment and the nature of labor markets were accompanied by improvements in the “technology” of disease prevention and treatment and by increased public and private investments in health and mortality reduction. During the 19th century, there were large changes in the scientific understanding of disease (see Preston and Haines, 1991, for a review). This led to dramatic declines in infant, child, and adult mortality rates that continued for close to a century. As scientific advances enabled reductions in mortality rates, there was strong pressure to increase public investments in health and disease prevention, from protection of the water supply to development of vaccines and public access to medical care. As a result, infant and child mortality rates were reduced dramatically, greatly increasing the probability that investments in children would be realized in terms of productive adulthood. The length of the productive adult life span, especially when time lost to morbidity is taken into account, has also increased significantly. Together, the two shifts in production processes and mortality rates favor increased human capital investment in a way that is reminiscent of the initial dietary shift leading to the hominid specialization discussed above.
This historical process also resulted in much greater labor force partici-
pation by women. During the initial demographic transition in the developed world, the breadwinner-homemaker family structure was dominant. With increased demand for labor that requires skill as opposed to strength and with growth in the service sector of the economy, wage-earning opportunities for women increased. At the same time, the payoffs to “home” production decreased with labor-saving devices, such as washing machines and refrigerators, and with smaller family size reducing the number of years spent caring for small children. Over time, therefore, there was a trend from greater to lesser complementarity between men and women.
Although the shift toward an education-based wage structure has been largely monotonic, those changes occurred at different times in the developed and developing worlds, and the details of the supply and demand for labor of different levels of human capital have been both historically and regionally variable. Moreover, both within and among societies, there appears to be a great deal of variation in rates of return on investments in educational capital.
The production of human capital is human-capital intensive (Becker and Barro, 1988). To see this, it is useful to think of an “education production function.” In each year of a child’s life, the amount he or she learns and the changes in his or her knowledge, reading, writing, logic and mathematical skills will depend on many different inputs, such as time, prior abilities, parents’ time, and teacher’s time. The value of those inputs, in terms of the educational capital produced, depends on the quality of those inputs. First, consider parents’ time. There is significant evidence that the nature of parent-child interaction varies with the educational level of the parents (Hart and Risley, 1995; Hoff-Ginsberg and Tardiff, 1995). For example, Hart and Risley report that, by the age of 3, children heard six million words if their mothers were professionals, 3 million words if their mothers were “working class”, and only 1 million words if their mothers were on welfare. By the time children enter the public education system there are clear differences in school-related skills, and those differences are related to socioeconomic status.
Second, the rate at which a child learns may depend on the knowledge and skills he or she already possesses. Much of the education offered in schools is based on the premise that knowledge is cumulative (Cromer, 1993). Basic skills are acquired first, and those skills are used as a foundation for the acquisition of the next set of skills. This implies that the impact of a child’s time inputs would depend on the skills already in place. It also means that the net increase in embodied capital at each age is a function of both the quality of inputs and the capital acquired at younger ages.
Moreover, those qualities tend to be correlated across inputs. Children with more educated parents also attend better schools with better teachers and better fellow students. At the other extreme, children in developing
nations often come from families in which neither parent has had formal schooling and attend schools with very large class sizes, almost no library resources, and teachers with only a primary education themselves. Under those conditions, much less is learned per year spent in schooling. For example, in a study of a predominantly black school in Cape Town, South Africa, Anderson et al. (2001) found that, on average, it took children 15 years to complete 12 grades of schooling. By that age (20 to 21), only about 10 percent of students passed the final matriculation exam and earned a high school diploma. The variance in those inputs leads to an increasing differentiation in educational capital with age.
This within-population heterogeneity in the costs of embodying capital in children means that environment does not determine diminishing returns to parental investment as it would be in primary production economies but will be frequency dependent. First, consider the highest-skilled jobs in the economy. Those jobs would be filled by individuals with the lowest costs of skill acquisition in decreasing rank order until the point is reached when the next-cheapest worker is more expensive than the product he or she produces. His or her parents would therefore invest less in his or her than would be necessary to obtain the highest-skilled jobs, and he or she would find employment in the next skill tier. That tier would then be occupied by individuals in decreasing rank order, until the next-cheapest worker will not be paid enough to compensate for skill embodiment. This process would continue through to the lowest-skilled jobs in the economy.
Individuals with low levels of human capital are more likely to be unemployed as well as to have a lower income when they are employed. This is especially true in urban areas in the developing world. The massive rural to urban migration over the past 40 years has resulted in very large populations of people with low levels of education competing for a limited number of low- skilled jobs. In many places, male unemployment can be as high as 70 percent. This variability in educational capital, along with its impacts on income variation across individuals and within individuals over time, has profound effects on family formation and reproduction.
Reproduction and Parental Investment
There is considerable evidence that in response to these conditions people in modern societies do not maximize fitness through their fertility decisions (see Kaplan and Lancaster, 2000, for a review). Observed fertility behavior deviates from the predictions of fitness maximization in two ways. First, and most important, observed fertility is lower than would be predicted based on models of fitness maximization. For example, among men in Albuquerque, New Mexico, the number of third-generation descendants (i.e., grandchildren) is highest among those who produced the most (i.e.,
>12) children (Kaplan et al., 1995). This contrasts sharply with the observed modal fertility of two. Mueller (2001) presents similar evidence from several other data sets in Europe and the United States. Higher parental fertility in modern developed societies is associated with lower educational and economic status of offspring (Blake, 1989; Downey, 1995), but the lower earning capacity of children from large families does not decrease their fertility, and so there is no apparent fitness reduction associated with lowered parental investment per child (Kaplan et al., 1995).
The second way in which modern behavior deviates from the predictions of simple budget constraint models of quantity-quality trade-offs is that higher-earning adults produce no more children than their lesser-earning counterparts, even in well-controlled studies. Whereas available data on preindustrial societies consistently exhibit a positive relationship between resources or power and reproductive success (Betzig, 1986; Boone, 1986; Chagnon, 1988; Hughes, 1986; Irons, 1979a; Kaplan and Hill, 1985; Low, 1990; Voland, 1990), studies of postdemographic transition societies either find no relationship or a negative one (Kaplan et al., 1995; Vining, 1986).
There is some evidence that the transition to low fertility begins with a negative association between income and fertility and then proceeds to a phase in which there is no association between the two (Kaplan and Lancaster, 2000; Retherford, 1993; Szreter and Garrett, 2000). Perhaps, the reason why the transition begins with a negative association between wealth and fertility is that the payoffs to investment in oneself and one’s offspring were not the same across economic strata. First, infant and child mortality rates are much lower among the wealthy than the poor (Preston and Haines, 1991) in both Europe and America. Second, wealthy parents tend to be more educated and to have greater access to educational resources. Our hypothesis is that, as a result, wealthier parents were able to educate their offspring more efficiently than poorer parents could and had a greater return on investments in both their and their offspring’s education.
Public- and private-sector responses to these trends then begin to change the payoffs to such investments among the poorer and less educated sectors of society. Increased public investments in health and schooling decrease mortality rates and increase the efficiency of private investments in children’s schooling for all sectors of society. More effective and less costly forms of birth control technologies allow for a significant reduction in birth rates. A more educated populace expands the expertise in firms with respect to both management and the technology of production, further increasing the demand for skilled workers. Ultimately, those interactions produce equal fertility among economic strata. Wealthier, more educated people invest more per child than poorer, less educated people because they are still more efficient at embodying capital in their children, but their higher wealth allows them to have as many children (although more costly) as less wealthy individuals.
Thus, what appears to be a gradual decrease in fertility through time according to national-level statistics may actually be a process in which an increasing proportion of the population exhibits very low fertility.
The effect of education on fertility varied with the total fertility rate during the 20th century. For men and women born early in the century, who reproduced during the Depression and war years, education delayed fertility. There was little effect of education on fertility during the Baby Boom years, when all education groups showed a reduction in age of first reproduction. As education became an increasingly important determinant of wages in the last third of the 20th century and fertility decreased again, the effects of education on fertility were most pronounced (Kaplan et al., 2002).
Virtually all of the effects of education on fertility are due to postponement of marriage and a delay from marriage to reproduction. The delay, coupled with age effects on fecundity due to reproductive physiology, results in lower completed fertility. For example, data from the National Survey of Families and Households show that among women 35 to 44 years of age in 1990, high school dropouts have a mean of 2.77 children, which drops to 2.22 for those with high school degrees, 1.95 for those with bachelor’s degrees, and 1.43 for those with graduate degrees (Kaplan et al., 2002).
These shifts in the economy also appear to have had profound effects on mating and marriage. Two opposing forces act on marital stability. The decrease in complementarity associated with women’s entry into the labor force (especially when women began to reenter the labor force after short maternity leaves) renders divorce less costly, since women no longer rely solely on males for financial support. This change in the costs of divorce inspired the reform of divorce laws and, ultimately, increases in the divorce rate. On the other hand, the greater importance of education in wage determination increases educational homogamy in the mating market and parental assessments about the level of investment children require. Support for college is another form of balloon payment in the parental investment stream. Divorce has a large negative impact on both the probability that offspring will attend college and the likelihood of paternal financial support for college (Anderson et al., 1999b; Kaplan et al., 2002).
These two opposing forces affect people differentially, on the basis of their education and presumably their expected rate of return on investments in their children’s education. The effects of education on divorce when children are young are striking. Figure 7-5 shows the probability that a man will cease to live with a child before the age of 6 as a function of the child’s birth cohort and the father’s education level among men in Albuquerque, New Mexico. The increase in divorce rates is clearly evident. However, there is an interaction effect between a man’s education and the child’s birth cohort on the probability of separation between a child and its father. Among children whose father has less than a high school education,
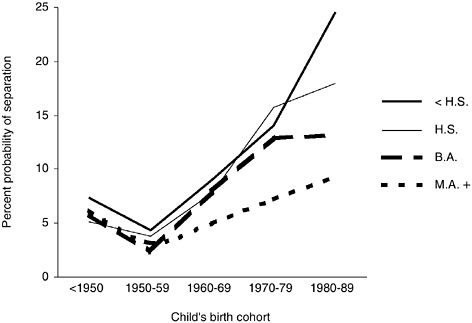
FIGURE 7-5 Father’s probability of ceasing to live with his child before the age of 6.
SOURCE: Kaplan et al. (2002). Reprinted with permission of Wiley-Liss, Inc., a subsidiary of John Wiley & Sons, Inc.
the percentage of children who cease to live with him before age 6 rises from about 5 percent for children born before 1960 to 23 percent for children born in the 1980 cohort. Among children whose father has a graduate degree, the percentage of children who cease to live with him before age 6 did not change significantly during this period and remains between 5 and 8 percent.
On the high end of the education continuum, both men and women delay marriage and delay the onset of childbearing after marriage until they are confident about their readiness to have children. After childbearing, their marriages are more stable, and their children are more likely to attend college and receive parental support to do so. The major cost of this strategy is that the delay can result in lower total fertility than desired because of age-related changes in fecundity (Kaplan et al., 2002).
We propose that a third factor comes into play at the low end of the education continuum. It has been pointed out that men who are less educated have higher mortality rates and greater rates of incarceration (Geronimus et al., 1999; Willis, 1987, 1994). This tends to make sex ratios female biased and promotes single parenthood. Additionally, periods of unemployment are likely to be interspersed with periods of employment,
making the income stream highly variable. This variability in male income streams may shift female strategies of mate choice and family formation. Rather than relying on a single man to provide resources over the long term, women derive support for themselves and their children from a number of different sources: (1) their own market labor; (2) assistance from female kin and other women (Geronimus, 1996; Geronimus and Korenman, 1992; Stack, 1974); or (3) temporary boyfriends, who are chosen in part because they are currently employed (Lancaster, 1989; Tucker and Mitchell-Kernan, 1995).
This pattern results in a system of serial and simultaneous polygynandry, where both women and men have multiple reproductive relationships (Lancaster, 1989; Lancaster and Kaplan, 1992). It also results in residential sibships composed of half-siblings with different fathers. Since it may be difficult for fathers to control the distribution of resources they provide to ex-partners so that they are preferentially directed toward their genetic offspring, men have less incentive to invest in them (Weiss and Willis, 1985; Willis and Haaga, 1996), placing further pressure on women to garner resources from other sources.
In the United States, many women, especially ethnic minorities, who face a mating market where male unemployment is great, choose to reproduce at younger ages while they are still living with their mothers, so that they can receive maternal or grand-maternal assistance in child rearing (Burton, 1990; Geronimus, 1996). The change in the family structure associated with teen childbearing over the second half of the 20th century was dramatic. For example, in 1960 only 15 percent of births to women under age 20 were to unmarried women; by 1994 the figure had risen to 76 percent (Child-Trends, 1995, 1996). Among ethnic groups with high levels of unemployed males, the proportion of all births to unmarried women has shown an equally dramatic increase; for example, by the late 1990s, more than 70 percent of all births to black and Native American women were nonmarital.
This trend is especially pronounced in the developing world, where male unemployment among migrants to the cities is exceptionally high. For example, Figure 7-6 shows the household living arrangements of the students in the Cape Town study discussed above. Less than a quarter of the students were living with both natural parents, about half were living either with their mother alone or with their mother and step-father, and more than a quarter were living with other relatives. It is interesting to note that the switch to the matrifocal family in South Africa from the traditional patrilineal patrilocal form was noted as early as the 1940s, as soon as there was significant migration to urban areas by women (Burman and Preston-Whyte, 1992; Burman and Reynolds, 1986). The global emergence of the matrifocal family in response to urbanization is another example of convergence among people with very different cultural backgrounds.
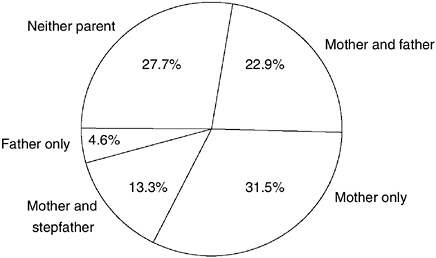
FIGURE 7-6 Living arrangements of I.D. Mkize High School students in Guguletu, South Africa.
SOURCE: Anderson et al. (2001).
Summary
In horticultural societies, access to the resources necessary for production and reproduction are reached through group membership. Groups are centered on productive resource patches that are used extensively and not possessed by individuals except through recognition of usufruct. Production by both males and females is for subsistence, and female labor underwrites much of the cost of supporting themselves and their children. This frees men to take as many wives as they can acquire through bride service or bride capture, creating conditions favoring endemic raiding for women. Parents focus their parental investment on raising as many healthy children as possible without concern for marriage market endowments or inheritance.
Pastoralism creates extrasomatic wealth in the form of highly mobile, easily divisible herds. This quality of livestock produces a reliable source of food but also a constant temptation to neighbors. Endemic warfare and raiding for women and livestock are the focus of the male life course, leading to expanded variance among men in resource holding and fertility based on the number of wives they can pay for or capture. At the same time that males must pay higher costs in bride wealth and in risk to access women, their families begin to help them by contributing a balloon payment at maturity to position them favorably on the mating market. Families of men now take responsibility not only to raise healthy children but also to
help sons find mates. Finally, owing to the fact that extrasomatic wealth creates a reproductive estate, families husband that estate for future generations by balancing quality of children (marriage endowments and inheritance) against quantity of children.
Early civilizations and premodern stratified social systems witness a complex of socioecological and reproductive patterns that are different from those of tribal societies. The first is an even stronger correlation between extrasomatic wealth and reproductive success than already found among pastoralists and brings into focus female-female competition in the marriage market. Since extrasomatic wealth has the potential of being controlled by powerful resource holders, social stratification and the ensuing variance among men restructure the mating market. The families of daughters compete for access to quality grooms by offering not only the youth and fertility of their daughters but also their virginity, chastity, and a dowry payment. Families that are resource holders focus on ways to maintain the integrity of the family estate through inheritance, at the same time that they promote both sons and daughters with an advantageous foothold in the marriage market. These families make reproductive decisions with an eye to long-term resource management considerations (Voland, 2000). Families without reproductive estates (laborers) live as reproductive opportunists in the here and now.
Over time, population growth and environmental saturation led to an ever-increasing emphasis on parental limitation of claimants to family reproductive estates through monogamy and bastardy, sex preference, sibling birth-order effects, and delayed or nonreproduction by selected sons and daughters. However, unlike the conditions of the early civilizations, population movement through urbanization, warfare, colonialism, or migration siphons off excess reproduction to less saturated areas.
The existence of extrasomatic wealth may be the critical environmental condition to which the evolved human physiology and psychology do not respond so as to maximize fitness. When there is heritable wealth, such as cattle or land, the breast-feeding/energy balance system may generate higher fertility than parents desire, in terms of their own wealth and the resources they wish to pass on to their children. Adjustments to this situation may involve primarily differential inheritance, such as primogeniture and illegitimacy, but may also include late age of marriage (Coale and Treadway, 1986) or even celibacy (Boone, 1986) and lowered rates of polygyny by wealthy men (Luttbeg et al., 2000). Thus, perhaps we should not be surprised to find deviations from fitness maximization as soon as there are forms of extrasomatic wealth.
The extremely low fertility in modern societies may reflect the extreme importance of extrasomatic wealth and the multiple ways in which it can be used. Changes in the economy and public health increase the rate of wage
returns on investments in education, albeit not equally both within and among societies. Moreover, in modern society, people face a tremendous array of consumption goods, including housing, clothing, electronic equipment, and vehicles. The medium of exchange for obtaining those goods is money. Money is highly fungible in that it can be translated into any one of those goods. As diminishing returns to consumption of one good are reached, money can then be allocated to other goods. With all the goods available, there is always some good of which little has been consumed and returns to purchasing some amount of it are still high. The same can be said for investment in offspring’s embodied capital, which appears to include not only investments in schooling but also in goods associated with social training and social status, such as hobbies and sports, clothing, and toys. Much of these investments are commitments to favorable placements of children in the mating market. The low fertility exhibited in modern societies may reflect both the payoffs to investments in offspring income and a perceived lack of diminishing returns to other forms of consumption (see Haaga, 2001, for a related discussion).
Changes in male-female complementarity affect people differently, on the basis of returns to investments in education. Those with the highest returns delay marriage and fertility, preferentially mating with one another and having fairly stable reproductive unions. Those with the lowest returns converge on the matrifocal family in response to insecurity in the income flow and the high relative costs of purchasing child care.
CONCLUSIONS
The final aim of this chapter is to discuss the implications of this evolutionary framework for new research and theory building. The above overview examined broad trends in fertility, parental investment, and mating behavior in response to large-scale shifts in socioecology. The utility of the framework, however, will depend on its ability to stimulate productive research designed to explain more fine-grained patterns of variation within and among populations over time.
Perhaps the most fundamental set of questions raised by this analysis concerns how people allocate resources among their own consumption, their children’s consumption, investments in their own and their children’s human capital, and reproduction. An understanding of the decision-making processes underlying those allocations remains elusive. For example, many men in our focus group discussions conducted in 1991 in Albuquerque, New Mexico, mentioned that the major factor limiting their fertility was the monetary and time costs necessary to raise a child. Several Hispanic men, who had come from large families, observed that when they were growing up it was considered acceptable to have several children share the
same bedroom, but today each child needs his or her own room. Very little is known about how individual decisions are made and how social standards evolve regarding investments in children.
The same can be said of allocations to a person’s own consumption. We do not know why members of the elite in all societies engage in and institutionalize elaborate displays of consumption (Boone and Kessler, 1999). We also have a very incomplete understanding of the preferences and decision-making processes underlying consumption patterns in modern societies. Given limited budgets, decisions about one’s own and one’s children’s consumption impinge on fertility decisions.
With respect to the second major theme in this chapter, mating systems and parental investment by sex, our understanding is even more incomplete. The factors affecting convergence and divergence between the sexes in each of those allocation decisions have not yet been systematically investigated. Clearly humans exhibit a great deal of flexibility in their family arrangements; the relationships between allocation decisions by sex, mating markets, and socioeconomic conditions are a new frontier for research.
These considerations suggest a “two-pronged” research program. One prong is to understand the mechanisms underlying those allocation processes, and the other is to understand how those mechanisms interact with socioeconomic conditions in generating behavioral and demographic outcomes. Several distinct lines of research may each contribute to such an understanding.
One approach is to examine individual variation in the weights or relative values that people attach to changes in their own and their children’s consumption, to relative social standing, to sexual gratification, and to number of children. It would be useful to know if some portion of that variation is associated with additive and nonadditive genetic variation. Rather than simply examining the heritability of fertility per se, a productive direction for genetic mapping and behavioral genetics research may be to examine psychological characteristics hypothesized to be directly or indirectly involved in fertility and mating decisions. For example, it has been suggested that natural selection has resulted in a desire for sexual gratification but not for children per se (Potts, 1997). However, this has not been examined empirically although there is some evidence that high male status in contemporary society correlates with a man’s number of sexual encounters but not reproductive success (Vining, 1986).
Developmental research on the ontogeny of decisions regarding consumption, fertility, sexual fidelity, and investment in children would also be useful. Do experiences taking care of infants and children during childhood and adolescence, investment of time and resources by parents, marital status of parents, exposure to media, and school composition affect desired levels of consumption and desire to have and/or rear children? To what
extent are differences in selectivity of partner choice, relationship fidelity, willingness to invest in children after marital separation, and willingness to invest in stepchildren associated with developmental experience?
In a similar vein, it would be useful to investigate how information is evaluated and how those processes of evaluation affect the diffusion of information and behavioral changes through networks and communities (see Bock, 1999, for a discussion). How do people evaluate messages about breast-feeding or the risks associated with teen pregnancy? For example, do they simply weight messages by their frequency of occurrence, according to the prestige of the source, or according to some perceived relevance to their own individual situation? Do people actively choose to participate in networks or expose themselves to media that increase or decrease the likelihood of receiving different messages?
In terms of the interaction of those decision-making processes with socioecological/socioeconomic conditions, the major research implication is that micro-level studies of budgetary allocations and fertility should be designed in terms of their contribution to an integrated and general theory. An integrated theory in this sense does not imply a unicausal model of transitions (such as mortality change, economic development, or access to birth control), but rather an understanding of how constellations of factors jointly determine outcomes through their impacts on an organized response system.
The framework introduced here is designed to provide guidance for such research. It will be critical to analyze and measure how resources are acquired, the impacts of investments on resource acquisition, mortality rates and the impacts of investments on mortality, complementarity of male and female inputs, and variation among and within individuals over time in access to resources.
There is still much to be learned from the study of traditional societies. While the impacts of breast-feeding frequency and intensity on reproductive physiology have received considerable attention, the factors determining their patterning have not been systematically investigated but rather relegated to unexplained cultural differences. What determines the timing of solid food introduction, the duration of breast-feeding, and its diurnal rhythm, both across and within cultures? To what extent do features of the ecology in terms of work, child care, and food resources explain the cross-cultural variation? To what extent does its patterning respond to the individual condition of mother or baby within communities? Is it the baby’s or the mother’s behavior that exerts the most influence over the process? Similar questions can be asked of the regulation of work and food sharing with children and adolescents. Such microecological studies should provide insights into the evolved psychology underlying parent-offspring interactions and how ecology translates into varying demographic outcomes.
Another important research area in traditional societies is the study of
social status and social capital. Since it is possible that concerns about relative social standing exert strong influence on modern consumption behavior and fertility decisions, an understanding of its role in contexts without material wealth should provide new insights. For example, anthropologists who have witnessed groups during the early phases of exposure to material goods, such as wrist watches, jewelry, and radios, are often impressed by how quickly those items are adopted in apparent displays of social status. What are the determinants of social standing when there is no stored wealth? Why are people motivated to pursue it? What are the costs and benefits of its pursuit? It is possible that behavior relevant to social standing had little direct impact on the regulation of fertility decisions in the past but now plays a greater role because the two trade off against one another with modern budgets.
There is still considerable debate about the relationships between men and women in traditional societies. According to one view (Hawkes and Bliege Bird, 2002), the provisioning of meat by men is actually generalized mating effort, not parental effort. Meat is acquired and shared with the whole band, and men gain access to women through the social prestige it confers. The view presented here is that men do provision their families, either directly or through return shares of food (Gurven et al., 2000), and complementarity in parental investment has led to pair bonding and specialized capital investment trajectories by men and women. Further research to resolve this debate should provide general insights into the factors affecting parental investment and mating systems.
The framework may also be applied to critical contrasts in historical and modern contexts. For example, it could be used to investigate why in the early 19th century couples in France had reduced marital fertility, whereas delays to marriage had larger impacts on fertility in England (Szreter and Garrett, 2000). One hypothesis is that differences in the labor markets between the two countries, with women in France juggling work and reproduction simultaneously and women in England working until marriage. The framework could also be applied to below-replacement fertility. Is very low fertility in Eastern and Southern Europe due to the same costs and benefits, or are there different factors leading to similar outcomes? For the most part, world population projections are based on statistical trends from the past and guesses about convergence on stable total fertility rates (Lutz et al., 2001). A deeper understanding of the relationship between conditions and fertility behavior would be useful for making population projections, especially in light of changing conditions.
The global growth of the matrifocal family is perhaps the most critical area for comparative research. Following the World Population Conference in Cairo (1994), the major policy emphasis has been on improving women’s education and autonomy in work, family, and reproductive decision mak-
ing. Such policies fail to consider the relationships between men’s economic opportunities, mating systems, and the investment that children receive. We need a better understanding of men’s and women’s criteria for choice of mates and how those criteria vary with socioeconomic conditions and how the dynamics of mating systems affect exchanges between men and women and between men and their genetic children and stepchildren. To what extent are men’s motivations and behavior regarding investments in children determined by their relationships with the child’s mother versus parental concerns, and how does that mix vary with socioeconomic conditions (see Anderson et al., 1999a, 1999b, for a discussion)?
The framework also has implications for research into the emergence of institutions and the interplay between institutions and individual behavior. Rather than viewing institutions (such as despotic systems of political control) as exogenous social determinants of individual behavior, the framework treats them as the emergent outcomes of interactions between individuals. In modern contexts, this suggests that increased research attention should be directed toward understanding the reciprocal influences of public and private investments and policies. For example, to what extent were increased public investments in health and education in the 20th century due to increased individual interest in such investments? Similarly, to what extent do government policies with regard to family planning reflect the individual behavior and interests of policy makers? How representative are policy makers of individuals in the general population? Since those institutional developments affect the costs and benefits of individual actions, there is clearly a route for reciprocal influence. We need better understanding of those reciprocal influences and of the relationship between micro- and macrolevel outcomes.
Evolutionary demography is best viewed not as an alternative to traditional approaches but as a general theoretical framework that can inform and enhance existing research endeavors. Economic modeling is fundamental to evolutionary analysis. Economists are primarily concerned with conscious, rational decision-making processes, but such processes are only a subset of the regulatory mechanisms of controlling fertility. In addition, there is no generally accepted causal theory of human utility in economics; natural selection is a causal process that constrains the types of utility functions that may evolve. Feminist demography details the conflicts of interest between men and women and how they vary with social context. A more basic evolutionary understanding of why men and women differ in their life histories and reproductive careers and how socioecology affects the extent to which the behavior and goals of the two sexes converge and diverge will enrich such insights. Most of human behavior is not instinctual, nor even the result of simple trial-and-error learning. Since information about the costs and benefits of alternative behavioral options is often so-
cially acquired, understanding cultural diffusion is critical. Evolutionary logic provides a framework for analysis of the active role that people play in determining which ideas they choose to adopt.
ACKNOWLEDGMENTS
This paper was written with support from the National Institute on Aging. The authors also wish to acknowledge the contributions of Kim Hill to the datasets and their analyses on the comparative diets and demography of chimpanzees and foragers published in a previous paper (Kaplan et al., 2000). We also thank Kim Hill and Magdalena Hurtado for their data on resource acquisition by age and sex among Hiwi and Ache. These datasets and analyses formed a critical base for part of this paper. We also wish to thank Monique Borgerhoff Mulder for her careful and extensive suggestions for revision of a draft of this paper.
REFERENCES
Anderson, K., H. Kaplan, D. Lam, and J. Lancaster 1999a Paternal care by genetic fathers and stepfathers II: Reports by Xhosa High School students. Evolution and Human Behavior 20:433-452.
Anderson K.G., H. Kaplan, and D. Lam 2001 Grade repetition, schooling attainment and family background in South Africa. Unpublished manuscript. Population Studies Center, University of Michigan, Ann Arbor.
Anderson, K.G., H. Kaplan, and J. Lancaster 1999b Paternal care by genetic fathers and stepfathers I: Reports from Albuquerque men. Evolution and Human Behavior 20:405-432.
Bailey, R.C., M.R. Jenike, P.T. Ellison, G.R. Bentley, A.M. Harrigan, and N.R. Peacock 1992 The ecology of birth seasonality among agriculturalists in Central Africa. Journal of Biosocial Science 24:393-412.
Becker, G.S. 1975 Human Capital. New York: Columbia University Press.
Becker, G.S., and R.J. Barro 1988 A reformulation of the economic theory of fertility. Quarterly Journal of Economics 103:1-25.
Bentley, G.R., R.R. Paine, and J.L. Boldsen 2001 Fertility changes with the prehistoric transition to agriculture. Pp. 203-232 in Reproductive Ecology and Human Evolution. P.T. Ellison, ed. Hawthorne, NY: Aldine de Gruyter.
Betzig, L. 1986 Despotism and Differential Reproduction: A Darwinian View of History. Hawthorne, NY: Aldine de Gruyter.
1992a Roman monogamy. Ethology and Sociobiology 13:351-383.
1992b Roman polygyny. Ethology and Sociobiology 13:309-349.
1993 Sex, succession and stratification in the first six civilizations: How powerful men reproduced, passed power on to their sons, and used their power to defend their wealth, women and children. Pp. 37-74 in Social Stratification and Socioeconomic Inequality. L. Ellis, ed. New York: Praeger.
Binford, L.R. 2001 Constructing Frames of Reference. Berkeley: University of California Press.
Bird, D.W., and R. Bliege Bird 2002 Children on the reef: Slow learning or strategic foraging. Human Nature 13:269-297.
Bliege Bird, R., and D.W. Bird 2002 Constraints of knowing or constraints of growing? Fishing and collecting among the children of Mer. Human Nature 13:00-00.
Blake, J. 1989 Family Size and Achievement. Los Angeles: University of California Press.
Blurton Jones, N.G., K. Hawkes, and J. O’Connell 1989 Modeling and measuring the costs of children in two foraging societies. Pp. 367-390 in Comparative Socioecology of Humans and Other Mammals. V. Standen and R.A. Foley, eds. London: Basil Blackwell.
Blurton Jones, N.G., K. Hawkes, and P. Draper 1994a Differences between Hadza and !Kung children’s work: Original affluence or practical reason. Pp. 189-215 in Key Issues in Hunter-Gatherer Research. E.S. Burch, ed. Oxford: Berg.
1994b Foraging returns of !Kung adults and children: Why didn’t !Kung children forage? Journal of Anthropological Research 50:217-248.
Bock, J. 1995 The Determinants of Variation in Children’s Activities in a Southern African Community. Ph.D. dissertation, University of New Mexico, Albuquerque.
1999 Evolutionary approaches to population: Implications for research and policy. Population and Environment 21:193-222.
2002 Learning, life history, and productivity: Children’s lives in the Okavango Delta, Botswana. Human Nature 13:161-198.
Bongaarts, J., and R.G. Porter 1983 Fertility, Biology and Behavior: An Analysis of Proximate Determinants. New York: Academic Press.
Boone, J. 1986 Parental investment and elite family structure in preindustrial states: A case study of late medieval-early modern Portuguese genealogies. American Anthropologist 88:259-278.
1988 Parental investment, social subordination and population processes among the 15th and 16th century Portuguese nobility. Pp. 201-220 in Human Reproductive Behavior: A Darwinian Perspective. L. Betzig, P. Turke, and M. Borgerhoff Mulder, eds. Cambridge: Cambridge University Press.
Boone, J.L., and K.L. Kessler 1999 More status or more children? Social status, fertility reduction, and long-term fitness. Evolution and Human Behavior 20:257-277.
Borgerhoff Mulder, M. 1988 Kipsigis bride wealth payments. In Human Reproductive Behavior. L. Betzig, M. Borgerhoff Mulder, and P. Turke, eds. Cambridge: Cambridge University Press.
1989 Early maturing Kipsigis women have higher reproductive success than late maturing women and cost more to marry. Behavioral Ecology and Sociobiology 24:145-153.
1991 Behavioural ecology of humans: Studies of foraging and reproduction. Pp. 69-98 in Behavioural Ecology. J.R. Krebs and N.B. Davies, eds. Oxford: Blackwell Scientific Publications.
1992 Demography of pastoralists: Preliminary data on the Datoga of Tanzania. Human Ecology 20:383-405.
1995 Bride wealth and its correlates: Quantifying changes over time. Current Anthropology 36:573-603.
1998 Brothers and sisters: How sibling interactions affect optimal parental investment. Human Nature 9:119-162.
2000 Optimizing offspring: The quantity-quality tradeoff in agropastoral Kipsigis. Evolution and Human Behavior 21:391-410.
Boserup, E. 1970 Women’s Role in Economic Development. London: Allen and Unwin.
Boswell, J. 1990 The Kindness of Strangers: The Abandonment of Children in Western Europe from Late Antiquity to the Renaissance. New York: Vintage Books.
Brown, J.K. 1970 A note on the division of labor by sex. American Anthropologist 72:1073-1078.
Burck, C.G. 1976 A group profile of the Fortune 500 child executives. Fortune (May Issue):173-177.
Burman, S., and E. Preston-Whyte, eds. 1992 Questionable Issue: Illegitimacy in South Africa. Cape Town: Oxford University Press.
Burman, S., and P. Reynolds, eds. 1986 Growing Up in a Divided Society: The Contexts of Childhood in South Africa. Johannesburg: Ravan Press.
Burton, L. 1990 Teenage childbearing as an alternative life-course strategy in multigenerational black families. Human Nature 1:123-144.
Chagnon, N.A. 1979 Is reproductive success equal in egalitarian societies? Pp. 374-401 in Evolutionary Biology and Human Social Behavior: An Anthropological Perspective. N.A. Chagnon and W. Irons, eds. North Scituate, MA: Duxbury Press.
1988 Life histories, blood revenge, and warfare in a tribal population. Science 239:985-992.
2000 Manipulating kinship rules: A form of male Yanomamo reproductive competition. Pp. 115-132 in Adaptation and Human Behavior: An Anthropological Perspective. L. Cronk, N. Chagnon, and W. Irons, eds. Hawthorne, NY: Aldine de Gruyter.
Charnov, E.L. 1993 Life History Invariants: Some Explanations of Symmetry in Evolutionary Ecology. Oxford: Oxford University Press.
Child Trends, Inc. 1995 Teen Births and Births to Unmarried Women: Facts at a Glance. Washington, DC: Author.
1996 New Data from Child Trends, Inc: Facts at a Glance. Washington, DC: Author.
Clarke, A.L. and B.S. Low 1992 Ecological correlates of human dispersal in 19th century Sweden. Animal Behavior 44:677-693.
2001 Testing evolutionary hypotheses with demographic data. Population and Development Review 27:633-660.
Clutton-Brock, T.H. 1991 The Evolution of Parental Care. Princeton, NJ: Princeton University Press.
Clutton-Brock, T.H., and G.A. Parker 1992 Potential reproductive rates and the operation of sexual selection. Quarterly Review of Biology 67:437-456.
Coale, A.J., and R. Treadway 1986 A summary of the changing distribution of overall fertility, marital fertility and the proportion married in the provinces of Europe. Pp. 31-181 in The Decline of Fertility in Europe. A.J. Coale and S.C. Watkins, eds. Princeton, NJ: Princeton University Press.
Colson, E. 1960 Social Organization of the Gwembe Tonga. Manchester: Rhodes-Livingstone Institute.
Cromer, A. 1993 Uncommon Sense: The Heretical Nature of Science. New York: Oxford University Press.
Crook, J.H., and S.J. Crook 1988 Tibetan polyandry: Problems of adaptation and fitness. Pp. 97-114 in Human Reproductive Behavior: A Darwinian Perspective. L. Betzig, M. Borgerhoff Mulder, and P. Turke, eds. Cambridge: Cambridge University Press.
Curtin, P.D. 1989 Death by Migration: Europe’s Encounter with the Tropical World in the Nineteenth Century. Cambridge: Cambridge University Press.
Dickemann, M. 1979a The ecology of mating systems in hypergynous dowry societies. Social Science Information 18:163-195.
1979b Female infanticide and the reproductive consequences of stratified human societies. In Evolutionary Societies and Human Social Behavior. N.A. Chagnon, and W. Irons, eds. North Scituate, MA: Duxbury Press.
1981 Paternal confidence and dowry competition: A biocultural analysis of purdah. In Natural Selection and Social Behavior. R.D. Alexander and D.W. Tinkle, eds. New York: Chiron Press.
DiVale, W., and M. Harris 1976 Population, warfare, and the male supremacist complex. American Anthropologist 80:21-41.
Downey, D.B. 1995 When bigger is not better: Family size, parental resources, and children’s education performance. American Sociological Review 60:746-761.
Draper, P. 1992 Room to maneuver: !Kung women cope with men. Pp. 43-63 in Sanctions and Sanctuary: Cultural Perspectives on the Beating of Wives. D. Counts, J.K. Brown, and J. Campbell, eds. Boulder, CO: Westview Press.
Dunbar, R.I.M. 1991 Sociobiological theory and the Cheyenne case. Current Anthropology 32:169-173.
Ellison, P.T. 2001a On Fertile Ground: A Natural History of Human Reproduction. Cambridge, MA: Harvard University Press.
Ellison, P.T., ed. 2001b Reproductive Ecology and Human Evolution. Hawthorne, NY: Aldine de Gruyter.
Ember, C.R. 1983 The relative decline in women’s contributions to agriculture with intensification. American Anthropologist 85:285-304.
Gaulin, S., and J. Boster 1990 Dowry as female competition. American Antrhopologist 92:994-1005.
Geronimus, A.T. 1996 What teen mothers know. Human Nature 7:323-352.
Geronimus, A.T., J. Bound, and T.A. Waidmann 1999 Health inequality and population variation in fertility timing. Social Science and Medicine 49:1623-1636.
Geronimus, A.T., and S. Korenman 1992 The socioeconomic consequences of teen childbearing reconsidered. The Quarterly Review of Economics 107:1187-1214.
Godfray, H.C.J., L. Partridge, and P.H. Harvey 1991 Clutch size. Annual Review of Ecology and Systematics 22:409-429.
Goodale, J.C. 1971 Tiwi Wives. Seattle: University of Washington Press.
Goody, J. 1973 Bride wealth and dowry in Africa and Eurasia. In Bride Wealth and Dowry. J.R. Goody and S.J. Tambiah, eds. Cambridge: Cambridge University Press.
1976 Production and Reproduction: A Comparative Study of the Domestic Domain. Cambridge: Cambridge University Press.
1983 The Development of the Family and Marriage in Europe. Cambridge: Cambridge University Press.
Grafen, A. 1991 Modeling a behavioural ecology. Pp. 5-31 in Behavioural Ecology: An Evolutionary Approach. J.R. Krebs and N.B. Davies, eds. Oxford: Blackwell Scientific.
Gurven, M., K. Hill, H. Kaplan, M. Hurtado, and R. Lyles 2000 Food transfers among Hiwi foragers of Venezuela: Tests of reciprocity. Human Ecology 28:171-218.
Gwyne, D.T. 1991 Sexual competition among females: What causes courtship role reversal. Trends in Evolution and Ecology 6:118-1222.
Gwyne, D.T., and L.W. Simmons 1990 Experimental reversal of courtship roles in an insect. Nature 346:172-174.
Haaga, J.G. 2001 Comment: The pace of fertility decline and the utility of evolutionary approaches. In Global Fertility Transition, R.A. Bulatao, and J.B. casterline, eds. Population and Development Review (suppl) 27:53-59.
Haddix, K. 2001 When polyandry falls apart: Leaving your wife and your brothers. Evolution and Human Behavior 22:47-60.
Harpending, H. 1994 Infertility and hunter-gatherer demography. American Journal of Physical Anthropology 93:385-390.
Hart, B., and T. Risley 1995 Meaningful Differences in the Everyday Experience of Young American Children. Baltimore, MD: Brookes.
Hart, C.W.M., and A.R. Pilling 1960 The Tiwi of North Australia. New York: Holt, Rinehart and Winston.
Hartung, J. 1982 Polygyny and the inheritance of wealth. Current Anthropology 23:1-12.
Hawkes, K., and R.Bliege Bird 2002 Showing off, handicap signaling, and the evolution of men’s work. Evolutionary Anthropology 11:58-67.
Hawkes, K., J.F. O’Connell, and H.G. Blurton Jones 1995 Hadza children’s foraging: Juvenile dependency, social arrangements and mobility among hunter-gatherers. Current Anthropology 36:688-700.
Herrnstein, R.J., and C. Murray 1994 The Bell Curve: Intelligence and Class Structure in American Life. New York: Free Press.
Hill, K., and A.M. Hurtado 1991 The evolution of reproductive senescence and menopause in human females. Human Nature 2:315-350.
1996 Ache Life History: The Ecology and Demography of a Foraging People. Hawthorne, NY: Aldine.
Hoff-Ginsberg, E., and T. Tardiff 1995 Socioeconomic status and parenting. In The Handbook of Parenting. M. Bornstein, ed. Hillsdate, NJ: Erlbaum.
Hrdy, S.B. 1994 Fitness tradeoffs in the history and evolution of delegated mothering with special reference to wet-nursing, abandonment, and infanticide. In Infanticide and Parental Care. S. Parmigiani and F.S. vom Saal, eds. Chur, Switzerland: Harwood Academic Publishers.
1999 Mother Nature: A History of Mothers, Infants, and Natural Selection. New York: Pantheon.
Hrdy, S.B., and D. Judge 1993 Darwin and the puzzle of primogeniture: An essay on biases in parental investment after death. Human Nature 4:1-45.
Hughes, A.L. 1986 Reproductive success and occupational class in eighteenth-century Lancashire, England. Social Biology 33:109-115.
Hurtado, A.M., K. Hawkes, K. Hill, and H. Kaplan 1985 Female subsistence strategies among Ache hunter-gatherers of Eastern Paraguay. Human Ecology 13:29-47.
Hurtado, A.M., and K. Hill 1990 Seasonality in a foraging society: Variation in diet, work effort, fertility, and the sexual division of labor among the Hiwi of Venezuela. Journal of Anthropological Research 46:293-345.
Irons, W. 1979a Cultural and biological success. Pp. 257-272 in Natural Selection and Social Behavior. N.A. Chagnon and W. Irons, eds. North Scituate, MA: Duxbury Press.
1979b Investment and primary social dyads. Pp. 181-213 in Evolutionary Biology and Human Social Behavior. N.A. Chagnon and W. Irons, eds. North Scituate, MA: Duxbury Press.
Isbell, L.A. 1991 Contest and scramble competition: Patterns of female aggression and ranging behavior among primates. Behavioral Ecology 2:143-155.
Jasienska, G. 2000 Why energy expenditure causes reproductive suppression in women. Pp. 59-84 in Reproductive Ecology and Human Evolution. P. Ellison, ed. Hawthorne, NY: Aldine de Gruyter.
Jerison, H. 1973 Evolution of the Brain and Intelligence. New York: Academic Press.
1976 Paleoneurology and the evolution of mind. Scientific American 234:90-101.
Kaplan, H.S. 1996 A theory of fertility and parental investment in traditional and modern human societies. Yearbook of Physical Anthropology 39:91-135.
Kaplan, H.S., and K. Hill 1985 Hunting ability and reproductive success among male Ache foragers. Current Anthropology 26:131-133.
Kaplan, H.S., and J.B. Lancaster 2000 The evolutionary economics and psychology of the demographic transition to low fertility. Pp. 238-322 in Human Behavior and Adaptation: An Anthropological Perspective. L. Cronk, W. Irons, and N. Chagnon, eds. Hawthorne, NY: Aldine de Gruyter.
Kaplan, H., K. Hill, A.M. Hurtado, J. Lancaster, and R. Robson 2001a Embodied Capital and the Evolutionary Economics of the Human Lifespan. Albuquerque: University of New Mexico.
Kaplan, H.S., K. Hill, A.M. Hurtado, and J.B. Lancaster 2001b The embodied capital theory of human evolution. Pp. 293-318 in Reproductive Ecology and Human Evolution. P.T. Ellison, ed. Hawthorne, NY: Aldine de Gruyter.
Kaplan, H., K. Hill, J.B. Lancaster, and A.M. Hurtado 2000 A theory of human life history evolution: Diet, intelligence, and longevity. Evolutionary Anthropology 9:156-185.
Kaplan, H.S., J. Lancaster, J. Bock, and S. Johnson 1995 Does observed fertility maximize fitness among New Mexican men? A test of an optimality model and a new theory of parental investment in the embodied capital of offspring. Human Nature 6:325-360.
Kaplan, H.S., J.B. Lancaster, W.T. Tucker, and K.G. Anderson 2002 An evolutionary approach to below replacement fertility. American Journal of Human Biology 14:1-24.
Kaplan, H.S., and A. Robson 2002 The emergence of humans: the coevolution of intelligence and longevity with intergenerational transfers. Proceedings of the National Academy of Sciences 99(15):10221-10226.
Keeley, L.H. 1996 War Before Civilization: The Myth of the Peaceful Savage. Oxford: Oxford University Press.
Kirwin, M. 1979 African Widows. Maryknoll: Orbis.
Kozlowski, J., and R.G. Weigert 1987 Optimal age and size at maturity in the annuals and perennials with determinant growth. Evolutionary Ecology 1:231-244.
Kramer, K. 2002 Variability in the duration of juvenile dependence: The benefits of Maya children’s work to parents. Human Nature 13:299-325.
Kuzawa, C.W. 1998 Adipose tissue in human infancy and childhood: An evolutionary perspective. Yearbook of Physical Anthropology 41:177-209.
Lack, D. 1968 Ecological Adaptations for Breeding in Birds. London: Methuen.
Lancaster, C.S. 1981 The Goba of the Zambezi: Sex Roles, Economics, and Change. Norman, OK: University of Oklahoma Press.
Lancaster, J.B. 1989 Evolutionary and cross-cultural perspectives on single parenthood. Pp. 63-72 in Sociobiology and the Social Sciences. R.W. Bell and N.J. Bell, eds. Lubbock: Texas Tech University Press.
1997 The evolutionary history of human parental investment in relation to population growth and social stratification. Pp. 466-489 in Feminism and Evolutionary Biology. P.A. Gowaty, ed. New York: Chapman and Hall.
Lancaster, J.B., and H. Kaplan 1992 Human mating and family formation strategies: The effects of variability among males in quality and the allocation of mating effort and parental investment. In Topics in Primatology: Human Origins. T. Nishida, W.C. McGrew, P. Marler, M. Pickford, and F. de Waal, eds. Tokyo: University of Tokyo Press.
Lancaster, J.B., H. Kaplan, K. Hill, and A.M. Hurtado 2000 The evolution of life history, intelligence and diet among chimpanzees and human foragers. Pp. 47-72 in Perspectives in Ethology: Evolution, Culture and Behavior. F. Tonneau and N.S. Thompson, eds. New York: Plenum.
Lawrence, M., and R.G. Whitehead 1988 Physical activity and total energy expenditure of child-bearing Gambian village women. European Journal of Clinical Nutrition 42:145-160.
Low, B.S. 1990 Occupational status, landownership, and reproductive behavior in 19th-century Sweden: Tuna parish. American Anthropologist 92:457-468.
1991 Reproductive life in nineteenth century Sweden: An evolutionary perspective on demographic phenomena. Ethology and Sociobiology 12:411-448.
1994 Men in the demographic transition. Human Nature 5:223-253.
2000 Why Sex Matters: A Darwinian Look at Human Behavior. Princeton, NJ: Princeton University Press.
Lummaa, V., and T. Clutton-Brock 2002 Early development, survival and reproduction in humans. Trends in Ecology and Evolution 17:141-147.
Luttbeg, B., M. Borgerhoff Mulder, and M.S. Mangel 2000 To marry again or not: A dynamic model for demographic transition. Pp. 345-368 in Adaptation and Human Behavior: An Anthropological Perspective. L. Cronk, N.A. Chagnon, and W. Irons, eds. Hawthorne, NY: Aldine de Gruyter.
Lutz, W., W. Sanderson, and S. Scherbov 2001 The end of world population growth. Nature 412:543-545.
Mace, R. 2000 An adaptive model of human reproductive rate where wealth is inherited: Why people have small families. Pp. 261-282 in Adaptation and Human Behavior: An Anthropological Perspective. L. Cronk, N.A. Chagnon, and W. Irons, eds. Hawthorne, NY: Aldine de Gruyter.
Manson, J., and R. Wrangham 1991 Intergroup aggression in chimpanzees and humans. Current Anthropology 32:369-390.
McDade, T.W. 2001 Parent-offspring conflict and the cultural ecology of breast feeding. Human Nature 12:9-25.
Mueller, U. 2001 Is there a stabilizing selection around average fertility in modern human populations? Population and Development Review 27:469-498.
Murdock, G.P. 1967 Ethnographic atlas: A summary. Ethnology 6:109-236.
Newcomer, M. 1955 The Big Business Executive: The Factors that Made Him, 1890-1950. New York: Columbia University Press.
Orians, G.H. 1969 On the evolution of mating systems in birds and mammals. American Naturalist 103:589-603.
Partridge, L., and P. Harvey 1985 Costs of reproduction. Nature 316:20-21.
Pennington, R. 2001 Hunter-gatherer demography. Pp. 170-204 in Hunter-Gatherers: An Interdisciplinary Perspective. C. Panter-Brick, R.H. Layton, and P. Rowley-Conwy, eds. Cambridge: Cambridge University Press.
Pike, I.L. 1999 Age, reproductive history, seasonality, and maternal body composition during pregnancy for nomadic Turkana of Kenya. American Journal of Human Biology 11:658-672.
Poppitt, S.D., A.M. Prentice, E. Jequier, Y. Schutz, and R.G. Whitehead 1993 Evidence of energy sparing in Gambian women during pregnancy: A longitudinal study using whole-body calorimetry. American Journal of Clinical Nutrition 57:353-364.
Potts, M. 1997 Sex and the birth rate. Population and Development Review 23:1-19.
Prentice, A.M., and R.G. Whitehead 1987 The energetics of human reproduction. Symposium of the Zoological Society of London 57:275-304.
Preston, S., and M. Haines 1991 Fatal Years: Child Mortality in Late Nineteenth Century America. Princeton, NJ: Princeton University Press.
Promislow, D.E.L., and P.H. Harvey 1990 Living fast and dying young: A comparative analysis of life history variation among mammals. Journal of Zoology 220:417-437.
Retherford, R.D. 1993 Demographic Transition and the Evolution of Intelligence: Theory and Evidence. Honolulu: East-West Center.
Robson, A., and H.S. Kaplan 2003 The coevolution of intelligence and life expectancy in hunter-gatherer economies. American Economics Review (in press).
Rogers, A. 1993 Why menopause? Evolutionary Ecology 7:406-420.
Scott, S., and C.J. Duncan 1999 Reproductive strategies and sex-biased investment. Human Nature 10:85-108.
Sear, R., R. Mace, and I.A. McGregor 2001 Determinants of fertility in rural Gambia: An evolutionary ecological approach. Paper presented at the General Population Conference, International Union for the Scientific Study of Population.
Sellen, D.W., and R. Mace 1997 Fertility and mode of subsistence: A phylogenetic analysis. Current Anthropology 38:878-889.
Sellen, D.W., and D.B. Smay 2001 Relationship between subsistence and age at weaning in “pre-industrial” societies. Human Nature 12:47-87.
Shanley, D.P., and T.B.L. Kirkwood 2000 Calorie restriction and aging: A life-history analysis. Evolution 54:740-750.
Smith, C.C., and S.D. Fretwell 1974 The optimal balance between size and number of offspring. American Naturalist 108:499-506.
Stack, C. 1974 All Our Kin: Strategies for Survival in a Black Community. New York: Harper & Row.
Sterck, E.H.M., D.P. Watts, and C.P. van Schaik 1997 The evolution of female social relationships in nonhuman primates. Behavioral Ecology and Sociobiology 41:291-309.
Svedberg, P. 1990 Undernutrition in Sub-Saharan Africa: Is there a gender bias? Journal for Development Studies 26:469-486.
Szreter, S., and E. Garrett 2000 Reproduction, compositional demography, and economic growth in early modern England. Population and Development Review 26:45-80.
Tatar, M., D.W. Grey, and J.R. Carey 1997 Altitudinal variation in senescence in a Melanoplus grasshopper species complex. Oecologia 111:357-364.
Towner, M. 1999 A dynamic model of human dispersal in a land-based economy. Behavioral Ecology and Sociobiology 46:82-94.
2001 Linking dispersal and resources in humans: Life history data from Oakham, Massachusetts (1750-1850). Human Nature 12:321-350.
Trivers, R.L. 1972 Parental investment and sexual selection. In Sexual Selection and the Descent of Man, 1871-1971. B. Campbell, ed. Chicago: Aldine.
Tucker, M.B., and C. Mitchell-Kernan 1995 The Decline in Marriage Among African Americans: Causes, Consequences, and Policy Implications. New York: Russell Sage Foundation.
Vining, D.R. 1986 Social versus reproductive success: The central theoretical problem of human sociobiology. Behavioral and Brain Sciences 9:167-216.
Vinovskis, M.A. 1994 Education and the economic transformation of nineteenth century America. Pp. 171-196 in Age and Structural Lag: Society’s Failure to Provide Meaningful Opportunities in Work, Family, and Leisure. M.W. Riley, R.L. Kahn, and A. Foner, eds. New York: John Wiley & Sons.
Voland, E. 1990 Differential reproductive success with the Krummhorn population (Germany, 18th and 19th centuries) . Behavioral Ecology and Sociobiology 26:65-72.
2000 Contributions of family reconstitution studies to evolutionary reproductive ecology. Evolutionary Anthropology 9:134-146.
Voland, E., and A. Chasiotis 1998 How female reproductive decisions cause social inequality in male reproductive fitness: Evidence from eighteenth- and nineteenth-century Germany. Pp. 220-238 in Human Biology and Social Inequality. S.S. Strickland and P.S. Shetty, eds. Cambridge: Cambridge University Press.
Voland, E., and R.I.M. Dunbar 1995 Resource competition and reproduction: The relationship between economic and parental strategies in the Krummhorn Population (1720-1874). Human Nature 6:33-49.
1997 The impact of social status and migration of female age at marriage in an historical population in north-west Germany. Journal of Biosocial Science 29:355-360.
Voland, E., R.I.M. Dunbar, C. Engel, and P. Stephan 1997 Population increase and sex-biased parental investment: Evidence from 18th and 19th Century Germany. Current Anthropology 38:129-135.
Voland, E., and C. Engel 1990 Female choice in humans: A conditional mate selection strategy of the Krummhorn women (Germany, 1720-1874). Ethology 84:144-154.
Voland, E., E. Suegrekjiwm, and C. Engel 1991 Cost/benefit oriented parental investment by high status families: The Krummhorn case. Ethology and Evolutionary Biology 12:105-118.
Weiss, Y., and R.J. Willis 1985 Children as collective goods and divorce settlements. Journal of Labor Economics 3:268-292.
Werner, E.E. 1986 Amphibian metamorphosis: Growth rate, predation risk and the optimal time to transform. American Naturalist 128:319-341.
White, D.R., and M.L. Burton 1988 Causes of polygyny: Ecology, economy, kinship, and warfare. American Anthropologist 90:871-887.
Williams, G.C. 1957 Pleitropy, natural selection and the evolution of senescence. Evolution 11:398-411.
Willis, R.J. 1987 Public and private intergenerational transfers, economic growth and demographic transition. Pp. 717-740 in Conference on Economic Development and Social Welfare in Taiwan. Taipei, Taiwan: Institute of Economics, Academica Sinica.
1994 Economic analysis of fertility: Micro foundations and aggregate implications. Pp. 139-171 in Population, Economic Development, and the Environment. K.L. Kiessling and H. Landberg, eds. New York: Oxford University Press.
Willis, R.J., and J.G. Haaga 1996 Economic approaches to understanding nonmarital fertility. In Fertility in the United States: New Patterns, New Theories. J.D. Casterline, R.D. Lee, and K.A. Foote, eds. New York: The Population Council.
Wood, J.W. 1994 Dynamics of Human Reproduction: Biology, Biometry and Demography. New York: Aldine de Gruyter.






















































