
3
Developing an Action Plan
The committee was charged with developing an action plan focused on preventing obesity in children and youth in the United States. The aim of the plan was to identify the most promising approaches for prevention, including policies and interventions for immediate action and in the longer term. The critical elements of the action plan’s development, described in this and subsequent chapters, were as follows:
-
Clarifying definitions related to key concepts
-
Developing a framework to guide the type and scope of data gathered
-
Articulating obesity prevention goals for children and youth
-
Identifying criteria for conducting an in-depth review of the available evidence
-
Translating the findings from the best available evidence into specific recommendations that comprise an integrated action plan.
DEFINITIONS AND TERMINOLOGY
Childhood and Adolescent Obesity
Body mass index (BMI) is an indirect measure of obesity based on the readily determined measures of height and weight. This report uses the term “obese” to refer to children and youth with BMIs equal to or greater than the 95th percentile of the age- and gender-specific BMI charts developed by
the Centers for Disease Control and Prevention (CDC) (Kuczmarski et al., 2000). In most children, values at this level are known to indicate excess body fat, which itself is difficult to measure accurately in either clinical or population-based settings.
What constitutes “excess” is an amount of body fat (often expressed as a percentage of body mass) that is sufficient to cause adverse health consequences. The exact percentage of body fat at which adverse consequences occur can vary widely across individuals and the consequences themselves—ranging from low self-esteem or mild glucose intolerance to major depression or nephropathy—show considerable variation as well.
BMI—calculated as weight in kilograms divided by the square of height measured in meters (kg/m2)—is the recommended indicator of obesity-related risks in both children and adults. For adults, overweight is defined as a BMI between 25 and 29.9 kg/m2 and obesity is defined as a BMI equal to or greater than 30 kg/m2 (NHLBI, 1998). The BMI cut-off points were based on epidemiological data that show increasing mortality above a BMI of 25 kg/m2, with greater increases above 30 kg/m2 (NHLBI, 1998).
Because children’s development varies with age, and because boys and girls develop at different rates, the use of BMI to assess body weight in children requires growth and gender considerations. Thus, BMI values for children and youth are specific to both age and gender (Barlow and Dietz, 1998; Dietz and Robinson, 1998).
The committee recognizes that it has been customary to use the term “overweight” instead of “obese” to refer to children with BMIs above the age- and gender-specific 95th percentiles (Himes and Dietz, 1994; Barlow and Dietz, 1998; DHHS, 2001a; Kuczmarski et al., 2002; AAP, 2003). Obese has often been considered to be a pejorative term, despite having a specific medical meaning. There have also been concerns about misclassification, as BMI is only a surrogate measure of body fatness in children as in adults. Furthermore, children may experience functional impairment (physical or emotional) at different levels of body fatness.
However, the term “obese” more effectively conveys the seriousness, urgency, and medical nature of this concern than does the term “overweight,” thereby reinforcing the importance of taking immediate action. Further, BMI in children correlates reasonably well to direct measures of body fatness (Mei et al., 2002), and high BMIs in children have been associated with many co-morbidities such as elevated blood pressure, insulin resistance, and increased lipids (Freedman et al., 2001). These are the same co-morbidities that often worsen in adult life and contribute to premature death from obesity.
The committee recognizes, however, that the term obese is probably not well suited for children younger than 2 years of age because the relationships among BMI, body fat, and morbidity are less clear at these ages.
Additionally, a high BMI in children younger than 2 years of age is less likely to persist than a high BMI in older children (Guo et al., 1994). BMI reference values are not established for children less than 2 years of age. Weight-for-length greater than the 95th percentile is used by CDC and the Special Supplemental Nutrition Program for Women, Infants, and Children to define overweight for children in this age group.
It is important that government agencies, researchers, health-care providers, insurers, and others agree on the same definition of childhood obesity. Although varying definitions have arisen from many uses of the term in public health, clinical medicine, insurance coverage, government programs and other settings, to the extent possible, there should be concurrence on definitions and terminology.
In this report, the term “obese” refers to children and youth between the ages of 2 and 18 years who have BMIs equal to or greater than the 95th percentile of the age- and gender-specific BMI charts developed by CDC.1
Prevention
To “prevent” means simply to take prior anticipatory action to hinder the occurrence of a course or event. Prevention efforts related to health traditionally have focused on preventing disease, particularly infectious disease. Conceptual frameworks have been developed that categorize health-related prevention efforts based on the segment of the population to which they are directed: the entire population (universal or population-based prevention); those who are at high risk of developing a disease (selective or high-risk prevention); or those who have a disease (targeted or indicated prevention) (Gordon, 1983; Rose, 1992; IOM, 1994; WHO, 2000).
Another traditional approach categorizes prevention according to disease progression: primary prevention involves avoiding the occurrence of a disease in a population; secondary prevention is aimed at early detection of the disease to limit its occurrence; and tertiary prevention is focused on limiting the consequences of the disease (DHHS, 2000).
A more recent framework conceptualizes a spectrum of prevention based on where—from the individual to the broader environment—the prevention actions are directed. Approaches include strengthening individual knowledge and skills, providing community education, educating
providers, fostering coalitions and networks, changing institutional practices, and influencing policy (Cohen and Swift, 1999).
The prevention frameworks discussed lend themselves relatively easily to infectious diseases in which there are clear endpoints and progressions. But the frameworks can be more complex to apply to health outcomes (e.g., childhood obesity) in which the progression is a continuum and the condition is both a risk factor for other chronic diseases and a health outcome in itself. The committee concluded that the well-established concept of primary prevention was most amenable to its assigned task of developing a broad-based action plan that addresses the social, cultural, and environmental factors associated with childhood obesity.
A primary prevention approach emphasizes efforts that can help the majority of children who are at a healthy weight to maintain that status and not become obese. Within this approach, the committee developed the majority of its recommendations as “population-based” actions—directed to the entire population instead of high-risk individuals. However, the committee acknowledges that obesity prevention will need to combine population-based efforts with targeted approaches for high-risk individuals and subgroups. Consequently, the report also contains specific actions aimed at high-risk populations affected by obesity, such as children and adolescents in particular ethnic groups with higher than average obesity-prevalence rates and communities in which there are recognizable social and economic disparities. Subpopulations of children warranting special consideration also include children with disabilities or special health-care needs. The complex medical, psychological, physical, and psychosocial difficulties that these children encounter may well put them at elevated risk for low physical activity levels and unhealthful dietary behaviors.
The committee acknowledges that although population-based prevention approaches may be theoretically or conceptually the most useful approaches for addressing a society-wide problem, the practical challenge is in determining how best to implement these interventions to achieve broad outreach and maximal coverage. These issues will be discussed further in the sections on local communities and evaluation of interventions (see Chapters 4 and 6).
The committee was not charged with, nor did it develop, recommendations directed specifically at obesity treatment or reducing excess weight in children and youth. However, it is likely that many of the suggested actions will also benefit children and youth who are already obese, even if the interventions are insufficient to produce enough short-term weight loss for achieving normal weight status. For example, obese children can benefit from healthful choices in the school cafeteria.
Prevention of obesity, particularly among those at high risk, may seem very similar to treatment in that screening is involved and individualized
intervention is often delivered in clinical settings. However, there are several important differences between prevention and treatment approaches (Kumanyika and Obarzanek, 2003). The targeted outcomes are different: prevention of weight gain is a satisfactory outcome for prevention approaches, whereas weight loss is the desired outcome for treatment. Motivations to maintain a healthful rate of weight gain for growing children may differ in nature and intensity from motivations to lose weight. Although treatment approaches may include relatively extreme behavioral changes over the short term, preventive strategies usually necessitate long-term continuation.
The committee’s approach to obesity prevention is similar to the range of prevention efforts that have been used to address many other public health problems. Some efforts directly change the physical environment but require no purposeful action on the part of the target population (e.g., fluoridation of community drinking water and food fortification); others directly require behavior change in targeted high-risk populations (e.g., immunization of children); and some require environmental change to facilitate behavioral change (e.g., zoning and land-use regulations to encourage physical activity). The majority of efforts require multiple approaches; for example, efforts to reduce underage drinking and tobacco control have involved legislation, media campaigns, counseling, and many other mechanisms (NRC and IOM, 2003; Mensah et al., 2004).
Appendix B provides a glossary of terms used throughout this report.
FRAMEWORK FOR ACTION
Using an ecological perspective, the committee developed a framework to depict the behavioral settings and leverage points that influence both sides of the energy balance2 equation—energy intake and energy expenditure. An ecological systems theory model postulates that changes in individual characteristics are affected not only by personal factors (e.g., age, gender, genetic profile) but also by interactions with the larger social, cultural, and environmental contexts in which they live (e.g., family, school, community) (Figure 3-1) (Davison and Birch, 2001; Lobstein et al., 2004).
Building on this ecological model and drawing upon concepts from several relevant frameworks (Swinburn et al., 1999; Booth et al., 2001; Kumanyika et al., 2002; Swinburn and Egger, 2004), the committee developed a framework that shows layers of ecologic factors as influences on energy imbalance, which is shown as the typical graphic in which energy
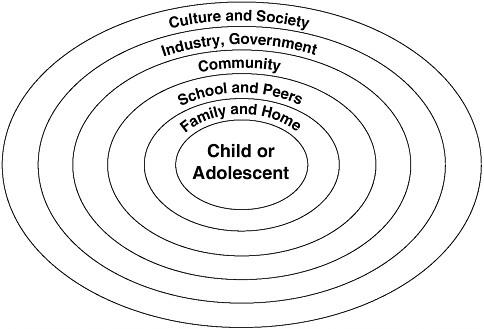
FIGURE 3-1 Simplified ecological systems theory model.
intake exceeds energy expenditure (Figure 3-2). Both aspects of energy imbalance (i.e., food and beverage intake and physical activity) interact with and are affected by multiple factors within each of the four ecological layers. The two innermost layers describe factors operating within the individual (including genetic factors, ethnic identity and culturally determined attitudes and beliefs, psychosocial factors, and current health status) and those operating within the physical and social locations and situations that define daily behavioral settings (Booth et al., 2001). The key behavioral settings for children and youth are the home, school, and community. As noted in the framework developed by the Partnership to Promote Healthy Eating and Active Living, behavioral settings are affected either directly or indirectly by a variety of other factors that potentially constitute primary and secondary leverage points for effecting changes (Booth et al., 2001). These leverage points include the major sectors that affect the food system, opportunities for physical activity or sedentary behavior, and information and education regarding dietary behaviors and physical activity. The outermost layer on the framework in Figure 3-2 reflects the critical concept of an overlay of social norms and values, that is, the social fabric that cuts across all the layers and processes below. Social norms and values both determine and respond to collective social and institutional processes within the con-
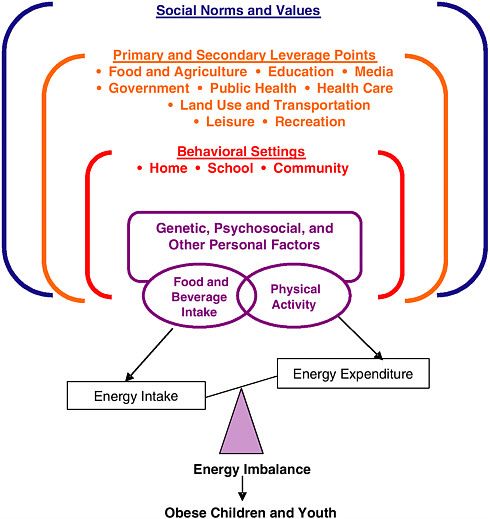
FIGURE 3-2 Framework for understanding obesity in children and youth.
NOTE: In this diagram energy intake is depicted as excessive when compared to energy expenditure, leading to a positive energy balance (or energy imbalance) resulting in obesity.
text of the larger U.S. culture. This framework, which emphasizes the need for obesity prevention efforts to leverage the interests and actions of a number of stakeholders working within and across multiple settings and sectors, guided the review of evidence and the development of recommendations in this report.
Clear specification of obesity prevention goals is essential in shaping an action plan and evaluating its success. Pertinent issues for setting obesity prevention goals for populations include concepts of optimum population BMI and healthy weight levels, potential effects on food intake and patterns of physical activity and inactivity (the primary modifiable determinants of obesity), as well as attitudes and social norms related to food and eating, physical activity and inactivity, body size, and dietary restrictions (WHO, 2000; Kumanyika et al., 2002). For children and youth, these considerations must be framed not only within the context of healthy physical, psychological, and cognitive development but in recognition that the increased prevalence of childhood obesity has broadened the emphasis of dietary guidance to address the overconsumption of energy-dense foods and beverages and physical activity patterns (ADA, 2003, 2004).
For individual children and youth, obesity prevention goals focus on maintaining energy balance (calories consumed versus calories expended). As discussed in greater detail later in the chapter, this involves engaging in healthful dietary behaviors and regular physical activity. Healthful dietary behaviors include choosing a balanced diet, eating moderate portion sizes, and heeding the body’s own satiety cues that indicate physiological fullness. It is currently recommended that children and adolescents accumulate a minimum of 60 minutes of moderate to vigorous physical activity each day (see section on physical activity).
Children’s food and beverage intake and their physical activity and sedentary behavior patterns can be influenced by a variety of environmental factors, including the availability and affordability of healthful foods, advertising messages, and opportunities to participate in physical activity within communities (Richter et al., 2000). Although individuals and families are embedded within broader social, economic, and political environments that influence their behaviors and may either promote or constrain the maintenance of health (IOM, 2001), such environments may also serve as contexts for change. These are the settings in which relationships are formed (e.g., home environment and support networks), and they represent a collection of formal and informal community institutions that monitor the behavior and safety of residents (Leventhal and Brooks-Gunn, 2001).
As will be noted throughout this report, changing the social, physical, and economic environments that contribute to the incidence and prevalence of childhood obesity—especially in populations in which the problem is longstanding and highly prevalent—may take many years to achieve. Therefore, the committee acknowledges that numerous intermediate goals, involving step-by-step improvements in diet patterns and physical activity levels of children and youth, are necessary for assessing progress. The ulti-
|
BOX 3-1 Goals of Obesity Prevention in Children and Youth The goal of obesity prevention in children and youth is to create—through directed social change—an environmental-behavioral synergy that promotes:
Because it may take a number of years to achieve and sustain these goals, intermediate goals are needed to assess progress toward reduction of obesity through policy and system changes. Examples include:
|
mate aim of obesity prevention in children and youth, however, is to create, through directed social change, an environmental-behavioral synergy that promotes positive outcomes both at the population and individual levels. Box 3-1 summarizes these long-term and intermediate goals, which will be discussed in greater detail throughout the report.
Optimum BMI and Healthy Weight
The concept of optimum BMI can be applied to populations. For countries such as the United States, where undernutrition is not as common as in developing countries,3 a BMI-distribution median of around 21 kg/m2 may be optimal (WHO, 2000). Population weight goals for obesity prevention in adults can also be stated in terms of decreasing the proportion that exceed the threshold of 30 kg/m2, although this goal includes both preventing new cases of obesity and reducing weight among those already over the threshold.
The same principles are appropriate for assessing the population of children in the United States in pursuit of the committee’s primary objective: to stop, and eventually reverse, current trends toward higher BMI levels. Also, as discussed in Chapter 2, there are particular concerns about the population of obese children becoming heavier. Achieving this objective would have the effects of reducing the mean BMI as well as decreasing the proportion of children and youth in the population that exceeds the threshold definition of obesity.
Available research does not currently allow the committee to define an optimum BMI for children and youth. It suggests, however, that future research toward this aim should be focused on defining the associations between BMI and objective measures of concurrent and future growth and between BMI and physiological and psychological morbidity, mortality, and health (Robinson, 1993; Robinson and Killen, 2001).
Analogous to the current practice for adults, the committee recommends the use of BMI for assessing individual and population changes in children and youth over time and in response to interventions. Population weight goals for childhood obesity prevention should be stated in terms of changes in the mean BMI and in the shape of the entire BMI distribution. Alternatively, goals can be stated in terms of decreasing the proportion of children or youth who exceed particular thresholds—e.g., 75th, 85th, 90th, 95th, or 97th percentiles of BMI for age and gender on the CDC BMI charts. In the absence of an appropriate evidence base, however, threshold goals are necessarily somewhat arbitrary and sacrifice substantial information about the rest of the distribution as well as substantial statistical power to detect differences between groups and over time (Robinson and Killen, 2001).
The current CDC guidelines for healthy weight in children and youth are in the range of the 5th to 85th percentiles of the age- and gender-specific BMI charts. Therefore, a child whose weight tracks in that range—that is, he or she does not cross to lower than the 5th or higher than the 85th percentiles—would be considered to be in the healthy weight range according to these definitions.
The CDC BMI charts are mathematically smoothed curves of the pooled growth parameters of children and adolescents sampled in cross-sectional national health surveys conducted from 1963 to 1994. An analogy would be to consider the curves as compiled from a series of “snapshots” of large national samples made at different times over three decades. But because the sample sizes at each age level get much smaller at the extremes of the distributions, the growth curves may be more prone to errors at the upper and lower ends.
Because of the increases in body weight that occurred in the 1980s and 1990s—after the second National Health and Nutrition Examination Survey (NHANES II) conducted in 1976-1980—a decision was made not to include the NHANES III (1988-1994) body-weight data in the revised 2000 BMI charts for children aged 6 years or older. The NHANES III data would have shifted the affected curves (weight-for-age and BMI-for-age) upward, which was considered to be biologically and medically undesirable. However, the fact that the CDC BMI charts were developed from data for a prior time period in which children were leaner, on average, leads to an occasionally confusing situation—for example, where more than 5 percent of the population is above the 95th percentile—but this is readily clarified in the context of the charts’ historical source.
The CDC BMI charts are derived from cross-sectional samples of children (data for different age groups are based on different children). That is, they do not directly represent the longitudinal growth trajectory for the same set of children who have been measured as they age.4 Therefore, it is not known whether an individual child’s height, weight, or BMI should be expected to follow along the same percentile curve over time in order to maintain health or whether there are health implications of variations throughout childhood (e.g., crossing percentiles by going from the 20th percentile at age 1 to the 60th percentile at age 5 to the 40th percentile at age 12). Mei and colleagues (2004) found that shifts in growth rates were
common during birth to 6 months and less common in children aged 2 to 5 years. More research is needed to determine whether there is an increased prevalence of “crossing” percentiles in different populations or during different age intervals and whether there are associations between crossing percentiles and health-related outcomes.
The problem is how to proceed despite this lack of certainty. The committee concluded that because the CDC BMI charts are based on large national samples of the U.S. population of children and youth, they are the best available tools for assessing growth in clinical and public health settings. Although there are many unknowns about how to apply this information to individual children, and clinicians face difficulties in making generalizations regarding normal growth trajectories, experience suggests that children who demonstrate rapid changes—that is, frequently crossing up or down percentiles—may require special health-care attention. Health-and medical-care professionals should be consulted regarding growth-related questions for individual children as they can assess a child’s own growth trajectory in context (see Chapter 6).
ENERGY BALANCE
Obesity prevention involves maintaining energy balance at a healthy weight while protecting overall health, growth and development, and nutritional status. Energy balance refers to the state in which energy intake is equivalent to energy expenditure, resulting in no net weight gain or weight loss. In adults, who have stopped growing, this relationship between energy intake and output must be equal and reach a zero net energy balance to prevent body storage of extra calories5 from food as fat and result in weight gain, which represents a positive energy balance. Strictly speaking, growing children, even those at a healthy body weight, must be in a slightly positive energy balance to satisfy the additional energy needs of tissue deposition for normal growth. However, for the purpose of simplicity in this report, the committee uses the term “energy balance” in children to indicate an equality between energy intake and energy expenditure that supports normal growth without promoting excess weight gain.
In children, energy expenditure constitutes the calories used for basal metabolism, processing of food, maintenance and repair of the body, and daily physical activity—in addition to the calories required for normal growth and development. Inappropriate weight gain (excess fat storage) results when energy expenditure is consistently exceeded by energy intake over time.
Energy intake is the calories ingested in the form of food and beverages. Children require a dietary pattern consisting of a variety of foods that provide all the necessary nutrients to support normal growth and development, as well as regular physical activity. Thus, a balanced diet refers to the consumption of appropriate amounts of a wide variety of nutrient-dense foods that provide adequate amounts and proportions of macronutrients (protein, fat, and carbohydrates) as well as sufficient essential micronutrients (vitamins, minerals) and dietary fiber, in addition to providing adequate energy to meet the needs of maintenance, growth, and development.
Although “energy intake = energy expenditure” looks like a fairly basic equation, in reality it is extraordinarily complex when considering the multitude of genetic, biological, psychological, sociocultural, and environmental factors that affect both sides of the energy balance equation and the interrelationships among these factors (Figure 3-2). For example, the amount, type, and intensity of physical activity influence body composition and physical fitness, which in turn influence the energy cost of physical activity (Hill et al., 2004).
There are several concepts regarding energy balance and weight gain in children and youth that the committee determined were important to clarify:
-
Genetics is a factor in excess weight but it is not the explanation for the recent epidemic of obesity (Koplan and Dietz, 1999). Although inherited tendencies toward weight gain may be a partial explanation for excess weight in children, as discussed below, there have been no measurable changes in the genetic composition of the population during the recent decades that could explain the significant increases in obesity.
-
Growth spurts do occur at several points throughout childhood and adolescence, but it cannot be assumed that a child will lose his or her excess weight at those times. Many experienced clinicians assess an individual child’s relative weight status by examining the consistency of that child’s weight or BMI percentiles over time. Thus, for example, after the age of about 4 years, normally growing children who are in the 20th or 50th or 65th percentile for weight would be expected to remain around these same percentiles for weight, during the remainder of their childhood. However, what can be considered normal variation to that pattern is not yet known, and is an important research question.
-
Physiological reasons for a child’s excess weight should be carefully explored by health-care professionals. However, the identifiable medical conditions that cause childhood obesity are rare and are not the principal underlying causes of the current obesity epidemic in the population.
-
The perceptions of what healthy children should “look like” differ among generations, cultures, and individuals. However, it is important that obesity not become the norm in society for children and youth as it poses
|
BOX 3-2 Balancing Food Intake and Physical Activity
SOURCE: DHHS, 2001b. |
-
serious health risks during childhood that can continue throughout adult life.
In the simplest terms, energy balance represents calories consumed versus calories expended, although as noted above, many individual variables can affect that balance. The discretionary variables under an individual’s control on a daily basis are dietary energy intake and the energy expended during physical activity.6 Daily energy intake is determined by the calorie content of the specific food and beverages consumed. Energy expenditure above resting metabolism is largely dependent on the nature and intensity of the activity and is often measured in calories per minute of activity (e.g., walking at a moderate or brisk pace of 3 to 4.5 miles per hour on a level surface expends between 3.5 and 7 calories per minute as measured in adults [CDC, 2004]). Knowing this, it is possible to determine the amount of physical activity that would be required to “burn off” the energy contained in a given food (Box 3-2). The relatively high amount of physical activity required to balance the calories in many preferred foods highlights the challenges of maintaining energy balance under conditions of a sedentary lifestyle and when surrounded by abundant food in large portions at relatively low cost. Much remains to be learned regarding the interactive effects of diet and physical activity—for example, the
extent to which increased physical activity or decreased dietary intake might improve the body’s own ability to regulate energy balance.
Furthermore, greater understanding is needed regarding the relative contribution of energy intake and energy expenditure to the energy imbalance that is driving the obesity epidemic. The increasing prevalence of obesity among children and youth in the United States could be the result of an upward shift in energy intake, a downward shift in energy expenditure, or the occurrence of both trends concurrently (Hill and Peters, 1998; Harnack et al., 2000; Hill et al., 2003). Some researchers have suggested that most of the effect is attributable to excessive energy intake (Sturm, 2005), while others have focused on the decline in regular physical activity and the increase in sedentary behaviors (Cutler et al., 2003).
It has been hypothesized that obesity can result from very small excesses in energy intake relative to expenditure and that the average weight gain in U.S. adults could be prevented if chronic energy expenditure exceeded intake by only 100 calories per day (Hill and Peters, 1998; Hill et al., 2003). However, estimates in a population of Hispanic children have shown greater potential energy gaps, ranging from approximately 200 to 500 calories per day (Butte and Ellis, 2003). This is an area requiring further research.
The following sections provide a brief overview of the context for energy balance and the complexities that researchers and policy makers face in these areas.
Genetic Variation and Biological Considerations
Obesity has long been recognized to occur in families, and having overweight or obese parents increases a child’s risk of being obese. After age 3, parental obesity is a stronger predictor of a child’s future obesity as an adult than is the child’s current weight (Whitaker et al., 1997).
Nonetheless, the familial clustering of obese individuals does not alone predict an individual’s weight characteristics, which reflect the combined effects of genetic variations, the common or shared environmental variations within family (which may include both intrauterine and infant feeding factors), and the environmental variations external to the family (Bouchard et al., 2003).
Quantifying with any precision the specific contributions of each of these factors to the development of obesity has been difficult, despite a variety of studies in nuclear families, in families with identical twins reared together or reared apart, and in families with adopted children. Bouchard and colleagues (2003) reviewed approximately 50 such studies and concluded that heritability accounts for about 25 to 40 percent of an individual’s expressed variation in weight and body fat mass. Specific ma-
ternal or paternal effects could not be identified. Using a new approach to twin studies, Segal and Allison (2002) concluded that common environmental effects might account for approximately 25 percent of the BMI variance in twins. It is important to note the difficulty in assigning proportionality to what is a gene-person-environment interaction.
Similarly, despite its intensity, the search for the specific genes responsible for an individual’s obese status has also been difficult. More than 400 genes, markers, and chromosomal regions have been linked to obesity phenotypes, 208 quantitative trait loci for human obesity have been identified, and 41 Mendelian disorders manifesting obesity have been genomically mapped (Snyder et al., 2004). However, only six single-gene defects resulting in obesity have been found, and in fewer than 150 individuals (Snyder et al., 2004). Thus, even though these monogenetic disorders have provided significant insight into the pathophysiology of obesity (Cummings and Schwartz, 2003; O’Rahilly et al., 2003), with few exceptions, human obesity appears to be a complex genetic trait. Nonetheless, genome-wide scans in widely varying populations have identified several genomic regions containing common quantitative trait loci for obesity phenotypes, suggesting that there may be shared genetic factors predisposing individuals of different ethnic origins to excessive storage of body fat (Bouchard et al., 2003). What is clear, however, is that the genetic characteristics of human populations have not changed in the last three decades, while the prevalence of obesity has approximately doubled. Thus, the recent population rise in body weight reflects the interaction of genotypes that predispose individuals to obesity with detrimental behavioral and environmental factors.
In animals, the evidence is strong for such gene-environment interactions affecting body weight and energy balance (Barsh et al., 2000), with the responsible genes orchestrating a complex system of biological feedback. In this system, central nervous system signals integrate messages about energy intake sent from the gastrointestinal tract with information about the current status of fuel reserves received from the energy-storing adipose tissue. The result is the direction of ingested food either into storage as fat or dissipation as energy, depending on the body’s status and needs at the time (Rosenbaum and Leibel, 1998; Havel, 2000, 2004; Druce and Bloom, 2003; Gale et al., 2004). What now seems clear is that this system evolved to defend the body from excessive energy deficit, a defense mechanism that has far less relevance today, when many humans are exposed to situations of food excess (Schwartz et al., 2003; Havel, 2004). Furthermore, although the system has now been characterized extensively in rodents and in adult humans, little is known about its development during the fetal period, infancy, or childhood (Box 3-3).
|
BOX 3-3 Food Intake Regulatory Systems In 1994, it was discovered that a peptide hormone—leptin—is manufactured and secreted by fat cells, travels through the circulatory system, crosses the blood-brain barrier, and acts on the brain’s hypothalamus to influence appetite (Zhang et al., 1994). This finding has led to the concept of a “fat-brain axis” (Elmquist and Flier, 2004), a pathway by which events in the periphery of the body are communicated to the brain. As a result, the brain may “monitor” the body’s energy or adipose stores and, when indicated, start a chain of events that either initiates or terminates feeding. There is now evidence that leptin affects both neuronal activity (Pinto et al., 2004) and synaptic plasticity (Bouret et al., 2004) in the arcuate nucleus of the hypothalamus, which is home to two distinct populations of neurons with opposing actions—one group that stimulates food intake and another that suppresses it (Elmquist and Flier, 2004). Furthermore, Bouret and colleagues (2004) suggest that leptin plays a neurotrophic role during the development of the hypothalamus that is restricted to a “neonatal critical period”—that is, the plasticity present early in life is apparently lost by adulthood. Although it is widely appreciated that good nutrition and a healthful lifestyle during the pregnancy period are important for producing healthy babies, these findings raise the possibility that the baby’s food-intake and body fat regulatory systems may be permanently shaped during this period. Future research undoubtedly will be directed to determining whether this communication system is indeed fundamental to the mechanisms of food-intake and body fat regulation in humans, and whether its timing is so narrowly focused. |
Psychosocial and Behavioral Considerations
Dietary Intake
Everyone needs to eat food and consume beverages for daily sustenance. But beyond the physical necessities are the complex social, cultural, and emotional nuances that involve food and permeate many facets of daily life. Children and adults alike consume food and beverages in part because they are hungry but also because eating and drinking are pleasurable and are an integral part of family life, celebrations, recreational events, and other social occasions. Food is also important in the psychosocial well-being, emotional expression, and coping responses of many people. It is, therefore, unrealistic to base recommended eating patterns solely on the chemical composition of foods without taking cultural, social, economic, and emotional drivers of food consumption into account. Furthermore, while few would dispute the negative aspects of individual substances such as tobacco, alcohol, or illegal drugs, there have been strong debates over
|
BOX 3-4 2000 Dietary Guidelines for Americans
SOURCE: USDA and DHHS, 2000. |
“good foods” versus “bad foods, even taking a purely nutritional perspective. Energy intake and dietary quality are determined by the total amounts and combination of foods consumed. A given food or beverage may have multiple nutritional quality dimensions and will have a differential impact on the overall eating pattern depending on what other foods are eaten. Nevertheless, the frequency of consuming certain types of foods is an indicator of the likelihood that the overall quantity and quality of foods will be appropriate, particularly in growing children for whom the nutrient density of diets (i.e., adequacy of vitamins and minerals per unit of energy intake) is important.
Based on current scientific evidence, the Dietary Guidelines for Americans provide nutritional advice to the American public on how to attain a balanced diet (defined in this report as an overall dietary pattern that provides all the essential nutrients in the appropriate amounts to meet nutritional needs and support life processes such as growth in children without promoting excess weight gain7) (Boxes 3-4 and 3-5; also see Chapter 5 and Appendix B).
Based on the Dietary Guidelines for Americans, the Healthy Eating
|
BOX 3-5 Benefits Associated with a Healthful Diet
SOURCES: IOM, 1997, 2002, 2004; USDA and DHHS, 2000. |
Index (HEI) is a tool developed by the U.S. Department of Agriculture to assess diet quality in order to provide a comprehensive assessment of diet in the U.S. population. A low HEI score suggests a poor diet and is also associated with overweight and obesity (Guo et al., 2004). Thus, the use of the HEI and the Dietary Guidelines for Americans as a way to improve health should be emphasized. However, the overall effectiveness of the Dietary Guidelines for Americans in disease prevention requires further research (Guo et al., 2004).
There are some indications of a small but significant increase in the average number of calories consumed daily by children over the last 15 to 20 years. The Continuing Survey of Food Intakes by Individuals, which examined changes between two time periods—1989-1991 and 1994-1996—in nationally representative samples of school-aged children, found an increase from 88 to 94 percent of the recommended energy allowance (Gleason and Suitor, 2001). Because no changes were seen in the energy intake from breakfast or lunch, the authors suggest that the increase was due to increased food consumption at dinner or in the form of snacks. Subsequent analyses of trends in energy intakes of children and youth have produced mixed findings (Enns et al., 2002; Nielsen et al., 2002; Sturm, 2005), and much remains to be learned about the dietary factors that contribute to the obesity epidemic in these groups.
Many challenges remain in conducting research on children’s dietary
intake. They include difficulties in children accurately recalling and quantifying foods consumed, the accuracy of third-party reports (usually parents or caregivers), and varying estimations of portion size. Use of the 24-hour recall method is common, but the need to collect information for multiple days to determine typical intake of foods or nutrients makes it a time- and labor-consuming process (Goran, 1998). Furthermore, the energy requirements for children vary, depending on the timing of growth and developmental spurts, and may be highly individualized.
Physical Activity
Physical activity, which has been defined as “any bodily movement produced by skeletal muscles that results in energy expenditure” (Caspersen et al., 1985), is in many respects synonymous with childhood. One of the joys and benefits of childhood is that being physically active is often a natural and fun part of playing and interacting with family and friends and does not generally involve a conscious decision to exercise. This play time is also developmentally important for children’s cognitive, motor-skill, and social development (NRC and IOM, 2000). Physical activity—not only in free play time, but in school, organized sports, and other activities—is an integral part of many children’s daily routines. However, as children grow, they generally become less physically active in adolescence and adulthood (Caspersen et al., 2000; Sallis, 2000). Additionally, children’s patterns of physical activity often differ from those of older adolescents and adults. Children often engage in intermittent activity mixed with brief periods of rest rather than in prolonged exercise (Goran et al., 1999).
Current recommendations are for children and adolescents to accumulate a minimum of 60 minutes of moderate to vigorous physical activity each day (Biddle et al., 1998; USDA and DHHS, 2000; Cavill et al., 2001; IOM, 2002; NASPE, 2004). The National Association for Sport and Physical Education recommends that children aged 5 through 12 years be involved in age-appropriate physical activity (including moderate to vigorous physical activity, most of it intermittent) that adds up to at least 60 minutes—and as much as several hours—per day on most days of the week (NASPE, 2004). Furthermore, long periods (two hours or more) of inactivity during the day time are discouraged in this age group. One of the strongest correlates of physical activity in children is time spent outside (Klesges et al., 1990; Baranowski et al., 1993; Sallis et al., 1993).
The health and quality-of-life benefits associated with regular moderate physical activity extend beyond the prevention of obesity (CDC, 1997) (Box 3-6). One of the major research challenges in this area is how to accurately measure physical activity, particularly in young children. Tools and techniques vary in terms of their intrusiveness into normal daily rou-
|
BOX 3-6 Benefits Associated with Physical Activity for Children and Adolescents Cardiovascular System
Musculoskeletal System
Mental Health, Psychological and Emotional Well-Being
Chronic Disease Prevention
SOURCES: DHHS, 1996; Sothern et al., 1999; Boreham and Riddock, 2001; Maziekas et al., 2003. |
tines (perhaps affecting activity level) and in the cost and time needed to collect and monitor the results. Questionnaires of parents and children are often confounded by recall problems and varying assessments of the type, intensity, and duration of the activity (Saris, 1986; Goran, 1998; Sirard and Pate, 2001). Measures of motion (e.g., pedometers and accelerometers) have come into wide use as research tools in recent years, but additional work is needed to ensure the validity of these methods in diverse groups of children and youth and in diverse settings. Additionally, research is needed to establish better methods of measurement of energy expenditure in children going through their normal daily activities in their home and school environments.
Sociocultural and Other Environmental Considerations
The specific types and levels of environmental factors to be considered as influences on food intake and physical activity are numerous. Tables 3-1 and 3-2 provide an illustrative listing of factors operating within different ecological layers (Swinburn et al., 1999). What is available with respect to food intake and physical activity opportunities (physical environment) is influenced by policies and financial inputs (political and economic environments) and is also targeted to the sociocultural milieu. Availability affects the range of possible individual choices, but personal choice is also mediated through a range of sociocultural variables that differ by age, gender, ethnicity, region, neighborhood characteristics, and socioeconomic status.
This matrix of environmental levels and types can also be developed to facilitate consideration of influences on obesity-related variables such as the availability of education and counseling and broader health promotion about weight gain prevention (physical environment), cost of preventive services (economic), and coverage of preventive services by third-party payers (policy environment). As discussed in the following sections, in the sociocultural domain, attitudes about body size and obesity are also critical contextual considerations when designing obesity prevention interventions.
Considerations Regarding Stigmatization
One of the concerns that arises in discussions regarding the prevention of childhood obesity is how to effectively focus on the behaviors that contribute to obesity without stigmatizing obese children and youth. As noted in Chapter 2, there is a body of research indicating that obese children and youth are stigmatized and experience negative stereotyping and discrimination by their peers, with adverse social and emotional consequences (Schwartz and Puhl, 2003).
Given that the stigmatization of obese children appears to have increased over a 40-year period from 1961 to 2001, there is a need to focus on the sensitivities regarding this issue and to explicitly reduce negative attitudes and behaviors such as teasing and discrimination directed toward obese children and youth (Latner and Stunkard, 2003; Schwartz and Puhl, 2003). This focus needs to be a consideration in the design of the range of interventions discussed throughout this report.
There is also the need to consider the adverse effects of normalization when discussing stigmatization. In many ways, American society has become more accepting of larger sizes in the products and portions we consume. Furthermore, our society often accommodates obesity as the social norm, for example, by resizing clothing, expanding the width of seating in public areas, and retrofitting ambulances to accommodate larger girth
TABLE 3-1 Examples of Environmental Influences on Food Intake, by Type of Environment
|
Size or Level of the Environment |
Type of Environment: Food-Related Influences |
|||
|
Physical |
Economic |
Policy/Political |
Sociocultural |
|
|
Microenvironments (e.g., behavioral settings such as homes, schools, and communities) |
|
|
|
|
|
Macroenvironments (e.g., societal sectorssuch as food and agriculture,education, medical, government,public health, or health care) |
|
|
|
|
|
SOURCE: Adapted from Swinburn et al., 1999. |
||||
TABLE 3-2 Examples of Environmental Influences on Physical Activity, by Type of Environment
|
Size or Level of the Environment |
Type of Environment |
|||
|
Physical |
Economic |
Policy/Political |
Sociocultural |
|
|
Microenvironments (e.g., behavioral settings such as homes, schools, and communities) |
|
|
|
|
|
Macroenvironments (e.g. societal sectors such as food and agriculture, education, medical, government, public health or health care) |
|
|
|
|
|
SOURCE: Adapted from Swinburn et al., 1999. |
||||
(Newman, 2004). Just as there are social and emotional consequences of stigmatization, there are also social and health consequences for obesity becoming the accepted social norm. This tension between stigmatization and normalization can be addressed, as it has been for other public health concerns, by focusing on the behaviors that can be changed to promote health rather than on the individual and his or her appearance.
It is important to note that the lessons learned from tobacco prevention and control efforts are not entirely applicable to obesity prevention. Bans against smoking in public buildings, on airplanes, and at other locations have encouraged some people to quit smoking due to the added inconvenience and public disapproval of this behavior. However, foods and beverages are necessary for sustenance and the issue is not “whether or not” to eat but rather what to eat, how much, and how often.
Areas of further research on this issue include how to encourage children to accept peers of all sizes and shapes and how to assist and support parents, teachers, children, and youth in addressing and coping with social stigma.
Body Image
A community’s norms, values, and expectations also affect the way that children in the normal or overweight (but not obese) range view their bodies. There is also concern that obesity prevention efforts will lead to inappropriate weight concern, dieting preoccupation, or unhealthful weight control practices among children and youth. Attitudes toward body size differ across cultures and especially affect females. Standards of attractiveness in males are less weight-dependent. Consistent with the stigma associated with being obese, the dominant attitudes in the United States and many similar societies favor a thin or lean body type in females, although as discussed below there is cultural variation in the degree of fatness or thinness that is acceptable as well as in preferred body shapes (Brown and Bentley-Condit, 1998). Attitudes about acceptable body size and shape also change over time and may apply differently to people of different ages.
The potential importance of this issue is underscored by reports of weight concerns in young children and in adolescents, in numerous ethnic groups, and in both low and high socioeconomic strata (see Chapter 2). Studies of children as young as the first grade have reported that a substantial proportion of children (about 50 percent of girls and 30 to 40 percent of boys), when given a choice of silhouettes will choose a thinner body size than their own as the “ideal” body size (Thompson et al., 1997). Robinson and colleagues (2001) studied a multiethnic and socioeconomically diverse sample of third graders (mean age was 8.5 years) in 13 northern California
elementary schools, and reported that concerns about being obese and dissatisfaction with body size were highly prevalent, increased with increasing BMI, and present—although to varying degrees—in all socioeconomic strata and ethnic groups. Furthermore, a study of 4,700 adolescents in Minnesota public schools (grades 7 through 12; mean age was 15 years) found high body satisfaction (versus low or moderate) in only 20 percent of girls and 34 percent of boys (Neumark-Sztainer et al., 2002).
Several studies have examined potential correlates of body image dissatisfaction and weight concerns or dieting practices, particularly gender, ethnicity, and socioeconomic status. Most of the studies that have examined ethnic differences consistently find less weight concern, less body size dissatisfaction, and a heavier ideal body size in African-American girls compared with white girls, but not necessarily boys, and sometimes demonstrate significant differences within African Americans across different socioeconomic levels (e.g., concern was greater at higher levels) (Thompson et al., 1997; Brown et al., 1998; Halpern et al., 1999; Adams et al., 2000; Neumark-Sztainer et al., 2002). These findings in children and adolescents are generally parallel to the numerous studies in adults indicating a relatively lower level of weight concern and higher level of body satisfaction in black women compared to white women; even considering the higher weight levels of the black women (Flynn and Fitzgibbon, 1998).
In contrast to the data for African Americans, available studies suggest that weight concerns in Hispanic and Asian girls are comparable to or exceed those in non-Hispanic white girls (Robinson et al., 2001; Neumark-Sztainer et al., 2002). The finding in Hispanic girls is consistent with data in adults (Serdula et al., 1999). Data for Native Americans in the Minnesota study (which were adjusted for grade level, socioeconomic status, and BMI) indicated a similar level of body satisfaction to that in white girls, but a significantly lower level of concern about controlling their weight (Neumark-Sztainer et al., 2002).
Socioeconomic Status
Socioeconomic status has generally been inversely associated with obesity prevalence (see Chapter 2) and children with obese mothers and low family income were found to have significantly elevated risks of becoming obese, independent of other demographic and socioeconomic factors (Strauss and Knight, 1999). When compared with food-insufficient households of higher income, low-income food-insufficient households had more obese children; however, food insufficiency by itself was not associated with self-reported measures of childhood obesity (Casey et al., 2001). Other studies have not been able to show a clear relationship between childhood
obesity and food insufficiency or food insecurity8 after adjusting for other confounding variables (Alaimo et al., 2001b; Kaiser et al., 2002; Matheson et al., 2002). However, food insecurity is associated with adverse health outcomes in infants and toddlers below 36 months of age (Cook et al., 2004) and with negative academic and psychosocial outcomes including depression in older children (Alaimo et al., 2001a, 2002).
Many of the variables in Tables 3-1 and 3-2 may be potential mediators of the relationship between socioeconomic inequities and childhood obesity. Both food and physical activity options are more likely to be periodically inadequate, unpredictable, or of lower quality for those with low personal incomes or those living in low-income neighborhoods (Travers, 1996; Morland et al., 2002a,b; Addy et al., 2004; Fitzgibbon and Stolley, 2004; Molnar et al., 2004). Poverty and living in low-income neighborhoods limit access to healthful foods. Some types of leisure-time physical activity are theoretically available at low or no cost, but these options may be less available to children in low-income neighborhoods because of neighborhood safety concerns, lack of adult supervision, or limited community recreational or other resources. Addressing childhood obesity in these contexts will require attention to root causes, and attempts to mitigate the underlying social and environmental adversity will be needed (Travers, 1997).
Racial and Ethnic Disparities
The substantially higher prevalence of obesity in adults, children, and youth in some African-American, Hispanic, American-Indian, and Pacific Islander populations (see Chapter 2) generates considerations across the entire ecologic framework (see Figure 3-2). A relatively high obesity prevalence in some Hispanic and American-Indian groups was noted prior to the obesity epidemic (Kumanyika, 1993); the pattern of excess weight gain and accelerated rates of obesity prevalence in African-American children and youth is a more recent development. It is now understood that issues of race are much more complex than the traditional U.S. Census Bureau racial and ethnic groupings often used in epidemiological research (Cooper, 2003; Cooper et al., 2003). However, the different historical and geographical
trajectories of these social and politically defined groups are associated with some differences in gene frequencies that may be linked with obesity development. Regardless, as discussed earlier in this chapter, the predominant factors responsible for the expression of obesity as a general population phenomenon are the linked behavioral and environmental factors outlined in the framework in Figure 3-2.
Many factors that potentially mediate racial and ethnic differences and predispose minority children and youth to high obesity risks can be postulated across physical, economic, sociocultural, and policy/political environments (Tables 3-1 and 3-2). Socioeconomic inequities are disproportionately common in minority populations and some of the excess risk may be mediated through economic and physical environmental factors related to low income or living in low-income communities. Other factors may affect individuals and communities on the basis of sociocultural factors that are not dependent upon socioeconomic status. Eating and physical activity patterns in some minority communities are less favorable to weight control than those in the general population, and these differences are observed within socioeconomic strata (Kumanyika and Krebs-Smith, 2001). For example, targeted marketing of high-calorie, low-nutrient-dense foods on black-oriented television has been reported (Tirodkar and Jain, 2003). Less access to supermarkets or to good quality food in supermarkets has been associated with black neighborhoods (Morland et al., 2002a) (see Chapter 6).
Sociocultural variables that need to be considered when approaching obesity prevention to reduce racial and ethnic disparities include traditional cuisines and any aspect of the attitudes, beliefs, and values (referred to in Tables 3-1 and Table 3-2 as the ethos or climate) that may facilitate or inhibit the promotion of healthful eating, physical activity, and weight control patterns in children and youth in these communities (Kumanyika and Morssink, 1997; Kumanyika, 2002, 2004). This ethos may include cultural values of responsiveness to or harmonization with the existing environmental context, as opposed to assumptions that the context can (or should) necessarily be changed. Included in the sociocultural environment are the high prevalence of obesity (e.g., the normative presence of the problem) as well as high levels of obesity-related health problems. In addition, to the extent that a history of discrimination or marginalization based on race or ethnicity becomes intertwined with other sociocultural factors, a certain level of skepticism or distrust relative to mainstream information and initiatives, including health information, may influence the receptivity to obesity prevention messages—particularly when these messages seem to conflict with pre-existing attitudes and beliefs.
REVIEW OF THE EVIDENCE
The committee identified a primary prevention, population-based approach to be the most viable long-term strategy for reducing obesity and its chronic disease burdens. Examples of the effectiveness of primary prevention interventions include smoking cessation to reduce lung cancer incidence, condom use to lower HIV transmission, and fruit and vegetable consumption to prevent cancer and cardiovascular diseases (CVDs) (Kroke et al., 2003; WHO, 2003).
There is no single acceptable standard, however, for assessing the entire range of prevention interventions and programs (Kellam and Langevin, 2003). Each phase of prevention research involves specific criteria for evidence and a variety of possible research designs. This is often a process whereby the preceding phase of research informs the subsequent generation of research—from efficacy to effectiveness, sustainability, going-to-scale, and, finally, sustaining system-wide9 (Figure 3-3). Numerous evidence-based prevention strategies are currently being used, though their focus—whether on individuals, institutions, or societal structures—can vary (Kellam and Langevin, 2003).
An Evidence-Based Medicine Approach
Evidence-based medicine is a valuable concept for informing clinical medicine that provides universally accepted standards for testing the scientific method and developing clinical practice guidelines (Harris et al., 2001; Heller and Page, 2002). This approach uses an accepted hierarchy of evidence—in accordance with its type, quality, and strength—to support recommendations (Table 3-3) (Harris et al., 2001; Kroke et al., 2003), and it establishes a cause-and-effect relationship guided by the principles of predictability, replicability, generalizability, and falsifiability. Predictability depends on a properly implemented intervention producing expected outcomes, a clear understanding of the intervention’s elements, and a cause-and-effect interaction among those elements (Tang et al., 2003). Replicability and generalizability rely on an intervention’s potential for
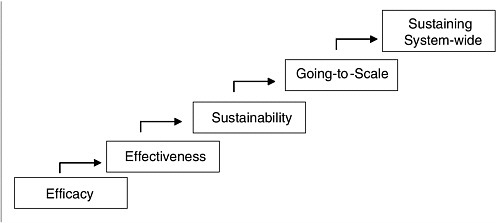
FIGURE 3-3 Five phases of prevention research.
SOURCE: Reprinted with permission, from Kellam and Langevin, 2003. Copyright 2003 by Prevention Science.
universal application that is independent of time, place, or context (Tang et al., 2003). Falsifiability refers to the possibility that an evaluation could determine, if relevant, that the intervention is ineffective (Tang et al., 2003).
An evidence-based medicine approach has been adopted by federal and scientific institutions to guide obesity treatment in adults (NHLBI, 1998). However, efforts to apply its principles to identifying effective interventions for other areas of disease prevention and health promotion have met with varying degrees of success (Osaka Declaration, 2001; McQueen, 2002; WHO, 2003; Victora et al., 2004). Indeed, it has been suggested that clinical decisions may have a relatively small impact on health outcomes compared to changes in the social environment, and that broadening evidence-based medicine beyond clinical policy decision-making—to public health decision-making—often has the potential to produce a larger beneficial impact on the health of populations (Heller and Page, 2002).
An Evidence-Based Public Health Approach
As the public health and health promotion disciplines have evolved, evidence-based public health has become the goal with a knowledge base that includes disease frequency and distribution; correlates, determinants and consequences of disease; and the safety, efficacy, effectiveness, and cost-effectiveness of a range of interventions (Victora et al., 2004). But given the complex environment in which multiple social, economic, cultural, and political elements interact to produce change in population-wide problems such as obesity, causality may not always be established for the
TABLE 3-3 Hierarchy of Research Design Used for Evidence-Based Clinical Medicine
|
Level of Evidence |
Type of Study |
|
I |
Evidence obtained from at least one properly designed randomized controlled trial (RCT) that provides a consistent pattern of findings in the population for which a recommendation is made. |
|
II-1 |
Evidence obtained from well-designed controlled trials without randomization. |
|
II-2 |
Evidence obtained from well-designed cohort or case-controlled analytical studies, preferably from more than one center or research group. |
|
II-3 |
Evidence obtained from multiple time-series or correlational studies with or without then intervention. |
|
III |
Evidence obtained from opinions of respected authorities, based on clinical experience, descriptive studies and case reports, or reports of expert committees. |
|
SOURCES: Harris et al., 2001; Kroke et al., 2003. |
|
relationships among the various interventions (McQueen, 2002; Tang et al., 2003).
Several factors complicate the task. The first is complexity in the causal sequences, including mediating factors, multiple causes acting simultaneously (some independently, others interactively), and the potential for unintended consequences from well-intended interventions. The second factor is that scientific uncertainty is associated with many or most of the causal links, which can vary across different social contexts and be constrained by current methods and ethical limitations (NRC, 1994). A third factor is that individuals and groups differ in the benefits and costs they attach to each of the causes, potential solutions, intended outcomes, and unintended consequences (Slovic, 1987, 2000). It has been suggested that there can be no purely scientific answer to the question of what should be done because the answer depends on social values (NRC, 1978). A fourth factor is that individuals and groups vary in how much uncertainty they are willing to tolerate before acting to address a problem (NRC, 1989).
The conclusion that results from these well-established principles is that while scientists can strive to clarify causal relations and reduce uncertainty, they are incapable of recommending specific actions (or inaction)
without making implicit value judgments (NRC, 1978, 1996). The solution to this dilemma ideally lies in the development and application of new approaches for integrating scientific considerations with social and normative considerations in a transparent, fair, and competent manner (Renn et al., 1995; NRC, 1996; Klinke and Renn, 2002).
Although randomized controlled trials (RCTs) are the gold standard for testing interventions in clinical and public health research, it is not always feasible, appropriate, or ethical to use that methodology in conducting population-based research; furthermore, RCTs may not always illuminate the complexity of some population-based prevention strategies (Robinson et al., 1998; Briss et al., 2000).
Therefore, the evidence base regarding public health prevention efforts often involves the integration of a range of research methodologies. Several health promotion and disease prevention initiatives have implemented comprehensive population health programs using a broader integrated approach to the evidence. For example, Table 3-4 illustrates the different approaches required for guiding the design of individual and community-based approaches to CVD prevention (Osaka Declaration, 2001).
TABLE 3-4 Comparison of Individual and Community Approaches for CVD Prevention
|
Clinical Practice Approaches for Individuals |
Community and Population-Based Approaches for Health Promotion |
|
The evidence standards are RCTs |
The evidence standards are RCTs and outcome and process evaluations that use both quantitative and qualitative methods |
|
The focus is on individual patients |
The focus is on the community |
|
Less than a therapeutic dose is unacceptable |
Preventive dose rarely applies |
|
Easier to treat an individual |
Difficult to scale up health promotion programs that reach the entire population |
|
Outcomes of interventions are individual change |
Outcomes are to change social norms, environments, and the behavior of entire populations |
|
Interventions can focus on most factors relevant to outcomes |
Interventions rarely take on social determinants external to the community |
|
SOURCE: Adapted from Osaka Declaration, 2001. |
|
Developing Recommendations Based on an Integrated Approach to the Evidence
The committee faced a significant challenge in deciding what types of evidence to use in formulating recommendations for obesity prevention in children and youth. A review of randomized controlled interventions for obesity prevention and treatment among children and adolescents identified only 35 such studies (Campbell et al., 2002). Due to the limited number of RCTs in obesity prevention efforts and methodological issues, including small sample sizes and high attrition rates of study participants, there is a paucity of RCT data from which to generalize results to broader populations (NHS Centre for Reviews and Dissemination, 2002).
The committee, therefore, developed guidelines for an integrated use of the available evidence to inform population-based obesity prevention interventions and on which to base its recommendations. This was deemed necessary to enhance the biological, psychosocial, and environmental plausibility of its inferences and identify consistency and congruency of information due to the paucity of causal research. Such an integrated-evidence approach has been used successfully to apply science-based principles to other public health efforts (Appendix D), such as in establishing a framework for evaluating the safety of dietary-supplement ingredients (IOM and NRC, 2004).
As childhood obesity is a serious public health problem calling for immediate reductions in obesity prevalence and in its health and social consequences, the committee strongly believed that actions should be based on the best available evidence—as opposed to waiting for the best possible evidence.
The different types of evidence that the committee used in developing the report’s recommendations are illustrated in Table 3-5, and the following principles guided the committee’s process:
-
Evidence is needed to inform and guide policy and programmatic decisions, justify a course of action, and evaluate the effectiveness of interventions that support obesity prevention.
-
Although the strength of the evidence is a basis for policy development, other considerations—including the fiscal and sociopolitical climate within which governments, institutions, and communities operate—must also be taken into account (Tang et al., 2003).
-
Absence of experimental evidence does not indicate a lack of causation or the ineffectiveness of an obesity prevention intervention. Given the methodological challenges, as well as the complexities in linkages between different elements and in their environments, certain interventions may prove effective even though their mechanisms for success are not known.
TABLE 3-5 Proposed Components of Evidence-Based Obesity Prevention
|
Objective |
Policy or Program Relevance |
Relevant Evidence and Information |
Types of Outputs |
|
Estimate the Health Burden Why should we do something about obesity? |
|
|
|
|
Identify the Determinants What are the causative and protective factors that could potentially be targeted by interventions? |
|
|
|
|
Describe the Framework for Action How and where should we intervene? |
|
|
|
|
Evaluate Potential Interventions What are the specific and potential interventions and their likely effectiveness? |
|
|
|
|
Select a Portfolio of Policies, Programs, and Actions What is a comprehensive portfolio of initiatives that is sufficient to prevent increases in obesity? |
|
|
|
|
SOURCE: Adapted from Swinburn et al., 2005. |
|||
-
This has been exemplified by programs that reduce television viewing time and decrease BMI in children (Robinson, 1999).
-
Given the significant shortage at present of experimental evidence to guide programs and policies, and the fact that many societal variables of interest have not been well addressed in controlled experimental studies as moderating or mediating factors, obesity prevention will require an evidence-based public health approach that continues to draw on RCTs, quasi-experiments, and observational studies as important sources of information (Victora et al., 2004).
-
Given that obesity is a serious health risk, preventive actions should be taken even if there is as-yet-incomplete scientific evidence on the interventions to address specific causes and correlates of obesity. However, there is an obligation to accumulate appropriate evidence not only to justify a course of action but to assess whether it has made a difference.
-
Finally, for interventions that have minimal potential risk and require few resources, formative and process evaluations may be sufficient to provide a “preponderance” of evidence (Robinson et al., 1998). As described in Appendix C, the committee conducted a thorough bibliographic search of the relevant scientific databases and benefited from the expertise of academic, industry, government, and nonprofit sector experts during its deliberations. In examining the literature, the committee focused on studies that examined weight and body composition outcomes, but it also broadened its scope to include studies that looked at changes in physical activity (or sedentary behavior) levels and in dietary intake patterns.
In examining the evidence on obesity-related prevention interventions, the committee considered the methodologies used by individual studies. Evaluating such studies involves characterizing the appropriateness of their designs for measuring target outcomes (e.g., increasing physical activity) as well as assessing the quality and generalizability of the study execution. The committee also considered the strength of the overall body of available evidence. Other factors considered by the committee included the feasibility of implementing the recommended actions, the opportunities for making changes, and the past success of parallel public health and social change efforts. Where trends of social, dietary, and other factors and health outcomes ran in parallel, the committee believes these trends merit further study and concern while acknowledging the possible occurrence of confounding.
It is also important to note that the committee focused on areas for improvement rather than on specific products, mechanisms for distribution, or industries. For example, the report emphasizes the nutritional evaluation of the contents of vending machines in schools rather than the re-
moval of vending machines (Chapter 7); considers the nutrient quality and energy density of foods and beverages rather than focusing on specific types of products (e.g., soft drinks, chips, candy); and highlights the improvements needed and actions that can promote energy balance rather than addressing any one industry (e.g., fast food restaurants).
SUMMARY
This report uses the term “obese” to refer to children and youth between the ages of 2 and 18 years who have BMIs equal to or greater than the 95th percentile of the age- and gender-specific BMI charts developed by CDC. For individuals, obesity prevention involves maintaining energy balance at a healthy weight while protecting overall health, growth and development, and nutritional status. Energy balance (calories consumed versus calories expended) is an extraordinarily complex concept when considering the multitude of genetic, biological, psychological, sociocultural, and environmental factors that affect both sides of the energy balance equation and the interrelationships among these factors.
Clear specification of obesity prevention goals is essential in shaping an action plan and evaluating its success. Relevant issues for setting obesity prevention goals for populations include concepts of optimum population BMI and healthy weight levels, potential effects on food intake and patterns of physical activity and inactivity, as well as attitudes and social norms related to food and eating, physical activity, inactivity, body size, and dietary restrictions. This chapter discusses a variety of influences on children’s diets and physical activity patterns including genetic variation and biological considerations, and sociocultural and other environmental factors.
Using an ecological systems theory model and a primary prevention evidence-based public health approach, this report focuses on how changes in the individual child’s behaviors are affected not only by individual factors but also through interactions with the larger social, cultural, and environmental contexts in which he or she lives (e.g., family, school, community, social and physical environments).
REFERENCES
AAP (American Academy of Pediatrics), Committee on Nutrition. 2003. Prevention of pediatric overweight and obesity. Pediatrics 112(2):424-430.
ADA (American Dietetic Association). 2003. Position of the American Dietetic Association: Child and adolescent food and nutrition programs. J Am Diet Assoc 103(7):887-893.
ADA. 2004. Position of the American Dietetic Association: Dietary guidance for healthy children ages 2 to 11 years. J Am Diet Assoc 104(4):660-677.
Adams K, Sargent RG, Thompson SH, Richter D, Corwin SJ, Rogan TJ. 2000. A study of body weight concerns and weight control practices of 4th and 7th grade adolescents. Ethn Health 5(1):79-94.
Addy CL, Wilson DK, Kirtland KA, Ainsworth BE, Sharpe P, Kimsey D. 2004. Associations of perceived social and physical environmental supports with physical activity and walking behavior. Am J Public Health 94(3):440-443.
Alaimo K, Olson CM, Frongillo EA Jr. 2001a. Food insufficiency and American school-aged children’s cognitive, academic, and psychosocial development. Pediatrics 108(1):44-53.
Alaimo K, Olson CM, Frongillo EA Jr. 2001b. Low family income and food insufficiency in relation to overweight in US children: Is there a paradox? Arch Pediatr Adolesc Med 155(10):1161-1167.
Alaimo K, Olson CM, Frongillo EA. 2002. Family food insufficiency, but not low family income, is positively associated with dysthymia and suicide symptoms in adolescents. J Nutr 132(4):719-725.
Ballew C, Bowman BA, Sowell AL, Gillespie C. 2001. Serum retinol distributions in residents of the United States: Third National Health and Nutrition Examination Survey, 1988-1994. Am J Clin Nutr 73(3):586-593.
Baranowski T, Thompson WO, DuRant RH, Baranowski J, Puhl J. 1993. Observations on physical activity in physical locations: Age, gender, ethnicity, and month effects. Res Q Exerc Sport 64(2):127-133.
Barlow SE, Dietz WH. 1998. Obesity evaluation and treatment: Expert committee recommendations. The Maternal and Child Health Bureau, Health Resources and Services Administration and the Department of Health and Human Services. Pediatrics 102(3):E29.
Barsh GS, Farooqi IS, O’Rahilly S. 2000. Genetics of body-weight regulation. Nature 404(6778):644-651.
Biddle S, Sallis JF, Cavill NA. 1998. Young and Active? Young People and Health Enhancing Physical Activity. Evidence and Implication. London: Health Education Authority.
Booth SL, Sallis JF, Ritenbaugh C, Hill JO, Birch LL, Frank LD, Glanz K, Himmelgreen DA, Mudd M, Popkin BM, Rickard KA, St Jeor S, Hays NP. 2001. Environmental and societal factors affect food choice and physical activity: Rationale, influences, and leverage points. Nutr Rev 59(3 Pt 2):S21-S39.
Boreham C, Riddock C. 2001. The physical activity, fitness and health of children. J Sports Sci 19(12):915-929.
Bouchard C, Pérusse L, Rice T, Rao DC. 2003. Genetics of human obesity. In: Bray GA, Bouchard C, eds. Handbook of Obesity. Etiology and Pathophysiology. 2nd ed. New York: Marcel Dekker.
Bouret SG, Draper SJ, Simerly RB. 2004. Trophic action of leptin on hypothalamic neurons that regulate feeding. Science 304(5667):108-110.
Briss PA, Zaza S, Pappaioanou M, Fielding J, Wright-De Aguero L, Truman BI, Hopkins DP, Mullen PD, Thompson RS, Woolf SH, Carande-Kulis VG, Anderson L, Hinman AR, McQueen DV, Teutsch SM, Harris JR, Task Force on Community Preventive Services. 2000. Developing an evidence-based guide to community preventive services: Methods. Am J Prev Med 18(1S):35-43.
Brown KM, McMahon RP, Biro FM, Crawford P, Schreiber GB, Similo SL, Waclawiw M, Striegel-Moore R. 1998. Changes in self-esteem in black and white girls between the ages of 9 and 14 years. The NHLBI Growth and Health Study. J Adolesc Health 23(1):7-19.
Brown PJ, Bentley-Condit VK. 1998. Culture, evolution, and obesity. In: Bray GA, Bouchard C, James WPT, eds. Handbook of Obesity. New York: Marcel Dekker. Pp. 143-156.
Butte NF, Ellis KJ. 2003. Comment on “Obesity and the environment: Where do we go from here?” Science 301(5633):598b.
Cameron N. 2002. British growth charts for height and weight with recommendations concerning their use in auxological assessment. Ann Hum Biol 29(1):1-10.
Campbell K, Waters E, O’Meara S, Kelly S, Summerbell C. 2002. Interventions for Preventing Obesity in Children. Oxford, U.K.: Cochrane Library.
Casey PH, Szeto K, Lensing S, Bogle M, Weber J. 2001. Children in food-insufficient, low-income families: Prevalence, health, and nutrition status. Arch Pediatr Adolesc Med 155(4):508-514.
Caspersen CJ, Powell KE, Christenson GM. 1985. Physical activity, exercise, and physical fitness: Definitions and distinctions for health-related research. Public Health Rep 100(2):126-131.
Caspersen CJ, Pereira MA, Curran KM. 2000. Changes in physical activity patterns in the United States, by sex and cross-sectional age. Med Sci Sports Exerc 32(9):1601-1609.
Cavill N, Biddle S, Sallis JF. 2001. Health enhancing physical activity for young people: Statement of the United Kingdom Expert Consensus Conference. Pediatr Exer Sci 13:12-25.
CDC (Centers for Disease Control and Prevention). 1997. Guidelines for school and community programs to promote lifelong physical activity among young people. MMWR Recomm Rep 46(RR-6):1-36.
CDC. 2004. Measuring Physical Activity. [Online]. Available: http://www.cdc.gov/nccdphp/dnpa/physical/measuring/index.htm [accessed August 18, 2004].
Cohen L, Swift S. 1999. The spectrum of prevention: Developing a comprehensive approach to injury prevention. Inj Prev 5(3):203-207.
Cook JT, Frank DA, Berkowitz C, Black MM, Casey PH, Cutts DB, Meyers AF, Zaldivar N, Skalicky A, Levenson S, Heeren T, Nord M. 2004. Food insecurity is associated with adverse health outcomes among human infants and toddlers. J Nutr 134(6):1432-1438.
Cooper RS. 2003. Race, genes, and health: New wine in old bottles? Int J Epidemiol 32:23-25.
Cooper RS, Kaufman JS, Ward R. 2003. Race and genomics. N Engl J Med 348(12):1166-1170.
Cummings DE, Schwartz MW. 2003. Genetics and pathophysiology of human obesity. Annu Rev Med 54(1):453-471.
Cutler D, Glaeser E, Shapiro J. 2003. Why Have Americans Become More Fat? NBER Working Paper No. W9446. Cambridge, MA: National Bureau of Economic Research.
Davison KK, Birch LL. 2001. Childhood overweight: A contextual model and recommendations for future research. Obes Rev 2(3):159-171.
DHHS (U.S. Department of Health and Human Services). 1996. Physical Activity and Health: A Report of the Surgeon General. Atlanta, GA: CDC.
DHHS. 2000. Healthy People 2010: Understanding and Improving Health. 2nd edition. Washington, DC: U.S. Government Printing Office. [Online]. Available: http://www.healthypeople.gov/document/tableofcontents.htm [accessed April 9, 2004].
DHHS. 2001a. The Surgeon General’s Call to Action to Prevent and Decrease Overweight and Obesity. Rockville, MD: Public Health Service, Office of the Surgeon General.
DHHS. 2001b. Overweight and Obesity: What You Can Do. Being Physically Active Can Help You Attain or Maintain a Healthy Weight. [Online]. Available: http://www.surgeongeneral.gov/topics/obesity/calltoaction/fact_whatcanyoudo.htm [accessed August 9, 2004].
Dietz WH, Robinson TN. 1998. Use of the body mass index (BMI) as a measure of overweight in children and adolescents. J Pediatr 132(2):191-193.
Druce M, Bloom SR. 2003. Central regulators of food intake. Curr Opin Clin Nutr Metab Care 6(4):361-367.
Elmquist JK, Flier JS. 2004. Neuroscience. The fat-brain axis enters a new dimension. Science 304(5667):63-64.
Enns CW, Mickle SJ, Goldman JD. 2002. Trends in food and nutrient intakes by children in the United States. Fam Econ Nutr Rev 14(2):56-68.
Fitzgibbon ML, Stolley MR. 2004. Environmental changes may be needed for prevention of overweight in minority children. Pediatr Ann 33(1):45-49.
Flegal KM, Ogden CL, Wei R, Kuczmarski RL, Johnson CL. 2001. Prevalence of overweight in US children: Comparison of US growth charts from the Centers for Disease Control and Prevention with other reference values for body mass index. Am J Clin Nutr 73(6):1086-1093.
Flynn KJ, Fitzgibbon M. 1998. Body images and obesity risk among black females: A review of the literature. Ann Behav Med 20(1):13-24.
Freedman DS, Khan LK, Dietz WH, Srinivasan SR, Berenson GS. 2001. Relationship of childhood obesity to coronary heart disease risk factors in adulthood: The Bogalusa Heart Study. Pediatrics 108(3):712-718.
Gale SM, Castracane VD, Mantzoros CS. 2004. Energy homeostasis, obesity and eating disorders: Recent advances in endocrinology. J Nutr 134(2):295-298.
Ganji V, Hampl JS, Betts NM. 2003. Race-, gender- and age-specific differences in dietary micronutrient intakes of US children. Int J Food Sci Nutr 54(6):485-490.
Gleason P, Suitor C. 2001. Children’s Diets in the Mid-1990’s: Dietary Intake and Its Relationship with School Meal Participation. Report No. CN-01-CD1. Alexandria, VA: USDA.
Goran MI. 1998. Measurement issues related to studies of childhood obesity: Assessment of body composition, body fat distribution, physical activity, and food intake. Pediatrics 101(3 Pt 2):505-518.
Goran MI, Reynolds KD, Lindquist CH. 1999. Role of physical activity in the prevention of obesity in children. Int J Obesity 23(S3):S18-S33.
Gordon R. 1983. An operational classification of disease prevention. Public Health Reports 98:107-109.
Guo SS, Roche AF, Chumlea WC, Gardner JD, Siervogel RM. 1994. The predictive value of childhood body mass index values for overweight at age 35 y. Am J Clin Nutr 59(4):810-819.
Guo X, Warden BA, Paeratakul S, Bray GA. 2004. Healthy Eating Index and obesity. Eur J Clin Nutr May 19 [Online]. Available: http://dx.doi.org/10.1038/sj.ejcn.1601989 [accessed September 8, 2004].
Halpern CT, Udry JR, Campbell B, Suchindran C. 1999. Effects of body fat on weight concerns, dating, and sexual activity: A longitudinal analysis of black and white adolescent girls. Dev Psychol 35(3):721-736.
Hampl JS, Taylor CA, Johnston CS. 2004. Vitamin C deficiency and depletion in the United States: The Third National Health and Nutrition Examination Survey, 1988 to 1994. Am J Public Health 94(5):870-875.
Harnack L, Snyder P, Story M, Holliday R, Lytle L, Neumark-Sztainer D. 2000. Availability of a la carte food items in junior and senior high schools: A needs assessment. J Am Diet Assoc 100(6):701-703.
Harris RP, Helfand M, Woolf SH, Lohr KN, Mulrow CD, Teutsch SM, Atkins D, Methods Work Group, Third US Preventive Services Task Force. 2001. Current methods of the U.S. Preventive Services Task Force: A review of the process. Am J Prev Med 20(3S):21-35.
Havel PJ. 2000. Role of adipose tissue in body-weight regulation: Mechanisms regulating leptin production and energy balance. Proc Nutr Soc 59(3):359-371.
Havel PJ. 2004. Update on adipocyte hormones: Regulation of energy balance and carbohydrate/lipid metabolism. Diabetes 53(S1):S143-S151.
Heller RF, Page J. 2002. A population perspective to evidence based medicine: “Evidence for population health.” J Epidemiol Community Health 56(1):45-47.
Hill JO, Peters JC. 1998. Environmental contributions to the obesity epidemic. Science 280(5368):1371-1374.
Hill JO, Wyatt HR, Reed GW, Peters JC. 2003. Obesity and the environment: Where do we go from here? Science 299(5608):853-855.
Hill JO, Saris WHM, Levine JA. 2004. Energy expenditure in physical activity. In: Bray GA, Bouchard C, eds. Handbook of Obesity: Etiology and Pathophysiology. 2nd ed. New York: Marcel Dekker.
Himes JH, Dietz WH. 1994. Guidelines for overweight in adolescent preventive services: Recommendations from an expert committee. The Expert Committee on Clinical Guidelines for Overweight in Adolescent Preventive Services. Am J Clin Nutr 59(2):307-316.
IOM (Institute of Medicine). 1994. Reducing Risks for Mental Disorders. Washington, DC: National Academy Press.
IOM. 1997. Dietary Reference Intakes for Calcium, Phosphorus, Magnesium, Vitamin D, and Fluoride. Washington, DC: National Academy Press.
IOM. 2001. Health and Behavior: The Interplay of Biological, Behavioral, and Societal Influences. Washington, DC: National Academy Press.
IOM. 2002. Dietary Reference Intakes for Energy, Carbohydrate, Fiber, Fat, Fatty Acids, Cholesterol, Protein, and Amino Acids. Washington, DC: The National Academies Press.
IOM. 2004. Dietary Reference Intakes for Water, Potassium, Sodium, Chloride, and Sulfate. Washington, DC: The National Academies Press.
IOM, NRC (National Research Council). 2004. Dietary Supplements: A Framework for Evaluating Safety. Washington, DC: The National Academies Press.
Kaiser LL, Melgar-Quinonez HR, Lamp CL, Johns MC, Sutherlin JM, Harwood JO. 2002. Food security and nutritional outcomes of preschool-age Mexican-American children. J Am Diet Assoc 102(7):924-929.
Kellam SG, Langevin DJ. 2003. A framework for understanding “evidence” in prevention research and programs. Prevention Science 4(3):137-153.
Klesges RC, Eck LH, Hanson CL, Haddock CK, Klesges LM. 1990. Effects of obesity, social interactions, and physical environment on physical activity in preschoolers. Health Psychol 9(4):435-449.
Klinke A, Renn O. 2002. A new approach to risk evaluation and management: Risk-based, precaution-based, and discourse-based strategies. Risk Anal 22(6):1071-1093.
Koplan JP, Dietz WH. 1999. Caloric imbalance and public health policy. J Am Med Assoc 282(16):1579-1582.
Kroke A, Boeing H, Rossnagel K, Willich SN. 2003. History of the concept of ‘levels of evidence’ and their current status in relation to primary prevention through lifestyle intervention. Public Health Nutr 7(2):279-284.
Kuczmarski RJ, Ogden CL, Grummer-Strawn LM, Flegal KM, Guo SS, Wei R, Mei Z, Curtin LR, Roche AF, Johnson CL. 2000. CDC growth charts: United States. Adv Data (314):1-27.
Kuczmarski RJ, Ogden CL, Guo SS, Grummer-Strawn LM, Flegal KM, Mei Z, Wei R, Curtin LR, Roche AF, Johnson CL. 2002. Data From the National Health Survey. 2000 CDC Growth Charts for the United States: Methods and Development. Hyattsville, MD: National Center for Health Statistics.
Kumanyika SK. 1993. Ethnicity and obesity development in children. Ann NY Acad Sci 699:81-92.
Kumanyika SK. 2002. Obesity treatment in minorities. In: Wadden TA, Stunkard AJ, eds. Handbook of Obesity Treatment. 3rd ed. New York: Guilford Publications. Pp. 416-446.
Kumanyika SK. 2004. Cultural differences as influences on approaches to obesity treatment. In: Bray GA, Bouchard C, eds. Handbook of Obesity: Clinical Applications. 2nd ed. New York: Marcel Dekker.
Kumanyika SK, Krebs-Smith SM. 2001. Preventive nutrition issues in ethnic and socioeconomic groups in the United States. In: Bendich A, Deckelbaum RJ, eds. Primary and Secondary Preventive Nutrition. Totowa, NJ: Humana Press. Pp. 325-356.
Kumanyika SK, Morssink CB. 1997. Cultural appropriateness of weight management programs. In: Dalton S, ed. Overweight and Weight Management. Gaithersburg, MD: Aspen. Pp. 69-106.
Kumanyika SK, Obarzanek E. 2003. Pathways to obesity prevention: Report of a National Institutes of Health Workshop. Obes Res 11(10):1263-1274.
Kumanyika S, Jeffery RW, Morabia A, Ritenbaugh C, Antipatis VJ, Public Health Approaches to the Prevention of Obesity (PHAPO) Working Group of the International Obesity Task Force (IOTF). 2002. Obesity prevention: The case for action. Int J Obes Relat Metab Disord 26(3):425-436.
Latner JD, Stunkard AJ. 2003. Getting worse: The stigmatization of obese children. Obes Res 11(3):452-456.
Leventhal T, Brooks-Gunn J. 2001. Changing neighborhoods and child well-being: Understanding how children may be affected in the coming century. In: Owens T, Hofferth S, eds. Children at the Millennium: Where Have We Come From, Where Are We Going? Advances in Life Course Research. New York: Elsevier Science.
Lobstein T, Baur L, Uauy R, IASO International Obesity TaskForce. 2004. Obesity in children and young people: A crisis in public health. Obes Rev 5(Suppl 1):4-85.
Matheson DM, Varady J, Varady A, Killen JD. 2002. Household food security and nutritional status of Hispanic children in the fifth grade. Am J Clin Nutr 76(1):210-217.
Maziekas MT, LeMura LM, Stoddard NM, Kaercher S, Martucci T. 2003. Follow up exercise studies in pediatric obesity: Implications for long term effectiveness. Br J Sports Med 37(5):425-429.
McQueen DV. 2002. Strengthening the evidence base for health promotion. Health Promotion International 16(3):261-268.
Mei Z, Grummer-Strawn LM, Pietrobelli A, Goulding A, Goran MI, Dietz WH. 2002. Validity of body mass index compared with other body-composition screening indexes for the assessment of body fatness in children and adolescents. Am J Clin Nutr 75(6):978-985.
Mei Z, Grummer-Strawn LM, Thompson D, Dietz WH. 2004. Shifts in percentiles of growth during early childhood: Analysis of longitudinal data from the California Child Health and Development Study. Pediatrics 113(6):e617-e627.
Mensah GA, Goodman RA, Zaza S, Moulton AD, Kocher PL, Dietz WH, Pechacek TF, Marks JS. 2004. (April). Law as a tool for preventing chronic diseases: Expanding the spectrum of effective public health strategies [Part 1 and 2]. Preventing Chronic Disease [Online]. Available: http://www.cdc.gov/pcd/issues/2004/apr/04_0009.htm [accessed April 20, 2004].
Molnar BE, Gortmaker SL, Bull FC, Buka SL. 2004. Unsafe to play? Neighborhood disorder and lack of safety predict reduced physical activity among urban children and adolescents. Am J Health Promot 18(5):378-386.
Morland K, Wing S, Diez Roux A, Poole C. 2002a. Neighborhood characteristics associated with the location of food stores and food service places. Am J Prev Med 22(1):23-29.
Morland K, Wing S, Diez Roux A. 2002b. The contextual effect of the local food environment on residents’ diets: The atherosclerosis risk in communities study. Am J Public Health 92(11):1761-1767.
NASPE (National Association for Sport and Physical Education). 2004. Physical Activity for Children: A Statement of Guidelines for Children Ages 5-12. 2nd edition. Reston, VA: NASPE.
Neumark-Sztainer D, Croll J, Story M, Hannan PJ, French SA, Perry C. 2002. Ethnic/racial differences in weight-related concerns and behaviors among adolescent girls and boys: Findings from Project EAT. J Psychosom Res 53(5):963-974.
Newman C. 2004. Why are we so fat? National Geographic (August):46-61.
NHLBI (National Heart, Lung, and Blood Institute). 1998. Clinical Guidelines on the Identification, Evaluation, and Treatment of Overweight and Obesity in Adults: The Evidence Report. NIH Publication No. 98-4083. Rockville, MD: NIH. [Online]. Available: http://www.nhlbi.nih.gov/guidelines/obesity/ob_gdlns.pdf [accessed November 21, 2003].
NHS Centre for Reviews and Dissemination. 2002. The prevention and treatment of childhood obesity. Eff Health Care 7(6):1-12.
Nielsen SJ, Siega-Riz AM, Popkin BM. 2002. Trends in energy intake in U.S. between 1977 and 1996: Similar shifts seen across age groups. Obes Res 10(5):370-378.
Nord M, Andrews M, Carlson S. 2003. Household Food Security in the United States, 2002. Alexandria, VA: USDA Economic Research Service. Food Assistance and Nutrition Research Report 35.
NRC (National Research Council). 1978. Knowledge and Policy: The Uncertain Connection. Washington, DC: National Academy Press.
NRC. 1989. Improving Risk Communication.Washington, DC: National Academy Press.
NRC. 1994. Science and Judgment in Risk Assessment. Washington, DC: National Academy Press.
NRC. 1996. Understanding Risk: Informing Decisions in a Democratic Society. Washington, DC: National Academy Press.
NRC, IOM. 2000. From Neurons to Neighborhoods: The Science of Early Childhood Development. Washington, DC: National Academy Press.
NRC, IOM. 2003. Reducing Underage Drinking: A Collective Responsibility. Washington, DC: The National Academies Press.
O’Rahilly S, Farooqi IS, Yeo GS, Challis BG. 2003. Minireview: Human obesity—Lessons from monogenic disorders. Endocrinology 144(9):3757-3764.
Osaka Declaration. 2001 (May). Health, Economics and Political Action: Stemming the Global Tide of Cardiovascular Disease. Declaration of the Fourth International Heart Health Conference, Osaka, Japan.
Pinto S, Roseberry AG, Liu H, Diano S, Shanabrough M, Cai X, Friedman JM, Horvath TL. 2004. Rapid rewiring of arcuate nucleus feeding circuits by leptin. Science 304(5667):110-115.
Renn O, Webler T, Wiedemann P, eds. 1995. Fairness and Competence in Citizen Participation: Evaluating Models for Environmental Discourse. Boston, MA: Kluwer Academic Publishers.
Richter KP, Harris KJ, Paine-Andrews A, Fawcett SB, Schmid TL, Lankenau BH, Johnston J. 2000. Measuring the health environment for physical activity and nutrition among youth: A review of the literature and applications for community initiatives. Prev Med 31(2 Part 2):S98-S111.
Robinson TN. 1993. Defining obesity in children and adolescents: Clinical approaches. Crit Rev Food Sci Nutr 33(4-5):313-320.
Robinson TN. 1999. Reducing children’s television viewing to prevent obesity: A randomized controlled trial. J Am Med Assoc 282(16):1561-1567.
Robinson TN, Killen JD. 2001. Obesity prevention for children and adolescents. In: Thompson JK, Smolak L, eds. Body Image, Eating Disorders, and Obesity in Youth: Assessment, Prevention, and Treatment. Washington, DC: American Psychological Association. Pp. 261-292.
Robinson TN, Patrick K, Eng TR, Gustafson D. 1998. An evidence-based approach to interactive health communication: A challenge to medicine in the information age. Science Panel on Interactive Communication and Health. J Am Med Assoc 280(14):1264-1269.
Robinson TN, Chang JY, Haydel KF, Killen JD. 2001. Overweight concerns and body dissatisfaction among third-grade children: The impacts of ethnicity and socioeconomic status. J Pediatr 138(2):181-187.
Rose G. 1992. The Strategy of Preventive Medicine. New York: Oxford University Press. Rosenbaum M, Leibel RL. 1998. The physiology of body weight regulation: Relevance to the etiology of obesity in children. Pediatrics 101(3 Part 2):525-539.
Sallis JF. 2000. Age-related decline in physical activity: A synthesis of human and animal studies. Med Sci Sports Exerc 32(9):1598-1600.
Sallis JF, Nader PR, Broyles SL, Berry CC, Elder JP, McKenzie TL, Nelson JA. 1993. Correlates of physical activity at home in Mexican-American and Anglo-American preschool children. Health Psychol 12(5):390-398.
Saris WH. 1986. Habitual physical activity in children: Methodology and findings in health and disease. Med Sci Sports Exerc 18(3):253-263.
Schwartz MB, Puhl R. 2003. Childhood obesity: A societal problem to solve. Obes Rev 4(1):57-71.
Schwartz MW, Woods SC, Seeley RJ, Barsh GS, Baskin DG, Leibel RL. 2003. Is the energy homeostasis system inherently biased toward weight gain? Diabetes 52(2):232-238.
Segal NL, Allison DB. 2002. Twins and virtual twins: Bases of relative body weight revisited. Int J Obes Relat Metab Disord 26(4):437-441.
Serdula MK, Mokdad AH, Williamson DF, Galuska DA, Mendlein JM, Heath GW. 1999. Prevalence of attempting weight loss and strategies for controlling weight. J Am Med Assoc 282(14):1353-1358.
Sirard JR, Pate RR. 2001. Physical activity assessment in children and adolescents. Sports Med 31(6):439-454.
Slovic P. 1987. Perception of risk. Science 236(4799):280-285.
Slovic P. 2000. Perceived risk, trust, and democracy. In: Connolly T, Arkes H, Hammond K, eds. Judgment and Decision Making: An Interdisciplinary Reader. New York: Cambridge University Press. Pp. 500-516.
Snyder EE, Walts B, Perusse L, Chagnon YC, Weisnagel SJ, Rankinen T, Bouchard C. 2004. The human obesity gene map: The 2003 update. Obes Res 12(3):369-439.
Sothern MS, Loftin M, Suskind RM, Udall JN, Blecker U. 1999. The health benefits of physical activity in children and adolescents and implications for chronic disease prevention. Eur J Pediatr 158(4):271-274.
Strauss RS, Knight J. 1999. Influence of the home environment on the development of obesity in children. Pediatrics 103(6):E85.
Sturm R. 2005 (in press). Childhood obesity – What can we learn from existing data on societal trends. Part 2. Preventing Chronic Disease [Online]. Available: http://www.cdc.gov/pcd/issues/2005/apr/04_0039.htm [access after March 15, 2005].
Swinburn BA, Egger GJ. 2004. Influence of obesity-producing environments. In: Bray GA, Bouchard C, eds. Handbook of Obesity. Clinical Applications. 2nd ed. New York: Marcel Dekker.
Swinburn BA, Egger G, Raza F. 1999. Dissecting obesogenic environments: The development and application of a framework for identifying and prioritizing environmental interventions for obesity. Prev Med 29(6 Pt 1):563-570.
Swinburn B, Gill T, Kumanyika S. 2005 (in press). Obesity prevention: A proposed framework for translating evidence into action. Obes Rev 6(1).
Tang KC, Ehsani JP, McQueen DV. 2003. Evidence-based health promotion: Recollections, reflections, and reconsiderations. J Epidemiol Community Health 57(11):841-843.
Tanner JM, Davies PS. 1985. Clinical longitudinal standards for height and height velocity for North American children. J Pediatr 107(3):317-329.
Thompson SH, Corwin SJ, Sargent RG. 1997. Ideal body size beliefs and weight concerns of fourth-grade children. Int J Eat Disord 21(3):279-284.
Tirodkar MA, Jain A. 2003. Food messages on African American television shows. Am J Public Health 93(3):439-441.
Travers KD. 1996. The social organization of nutritional inequities. Soc Sci Med 43(4):543-553.
Travers KD. 1997. Reducing inequities through participatory research and community empowerment. Health Educ Behav 24(3):344-356.
USDA (U.S. Department of Agriculture), DHHS (U.S. Department of Health and Human Services). 2000. Nutrition and Your Health: Dietary Guidelines for Americans. Home and Garden Bulletin No. 232, 5th ed. Washington, DC: Government Printing Office.
Victora CG, Habicht JP, Bryce J. 2004. Evidence-based public health: Moving beyond randomized trials. Am J Public Health 94(3):400-405.
Whitaker RC, Wright JA, Pepe MS, Seidel KD, Dietz WH. 1997. Predicting obesity in young adulthood from childhood and parental obesity. N Engl J Med 337(13):869-873.
WHO (World Health Organization). 2000. Obesity: Preventing and Managing the Global Epidemic. WHO Technical Report Series 894. Geneva: WHO.
WHO. 2003. Diet, Nutrition and the Prevention of Chronic Diseases. WHO Technical Report Series 916. Geneva: WHO.
Wright JD, Bialostosky K, Gunter EW, Carroll MD, Najjar MF, Bowman BA, Johnson CL. 1998. Blood folate and vitamin B12: United States, 1988-1994. Vital Health Stat 11(243):1-78.
Zhang Y, Proenca R, Maffei M, Barone M, Leopold L, Friedman JM. 1994. Positional cloning of the mouse obese gene and its human homologue. Nature 372(6505):425-432.














































