
3
How to Get from Acoustic Disturbance to Population Effects
The committee developed a conceptual model, named the Population Consequences of Acoustic Disturbance (PCAD) model as a framework to break the overwhelmingly difficult task of tracing acoustic stimuli to population effects into several manageable stages (Figure 3-1). The PCAD model was created as a first attempt to trace acoustic disturbance through the life history of a marine mammal and then to determine the consequences for the population. The model also serves as a framework for identifying existing data and data gaps. The model was distributed to workshop participants (Appendix D) before the workshop, discussed during the workshop, and, with the input of workshop participants, refined afterwards. The model involves five levels of variables that are related by four transfer functions. The first transfer function relates acoustic stimuli to behavioral responses. The second expresses behavioral disruption in terms of effects on critical life functions, such as feeding and breeding. The third integrates these functional outcomes of responses over daily and seasonal cycles, to link them to vital rates (see Figure 3-1) in life history. The fourth transfer function relates changes in the vital rates of individuals to population effects. Current data are insufficient to allow the PCAD model to serve as more than a conceptual model, so the listing of data at the first three levels—involving sound characteristics, behaviors, and life functions—is exemplary rather than all-inclusive. The relationship between vital rates and population effects is well defined, but the specification of relevant population effects involves policy decisions as well as scientific judgments.
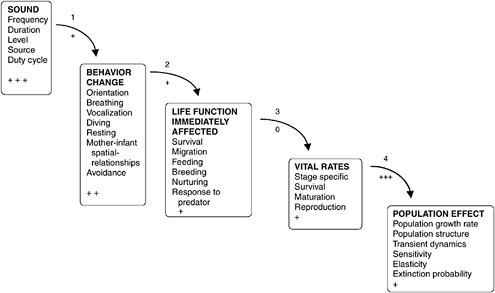
FIGURE 3-1 The conceptual Population Consequences of Acoustic Disturbance model describes several stages required to relate acoustic disturbance to effects on a marine mammal population. Five groups of variables are of interest, and transfer functions specify the relationships between the variables listed, for example, how sounds of a given frequency affect the vocalization rate of a given species of marine mammal under specified conditions. Each box lists variables with observable features (sound, behavior change, life function affected, vital rates, and population effect). In most cases, the causal mechanisms of responses are not known. For example, survival is included as one of the life functions that could be affected to account for such situations as the beaked whale strandings, in which it is generally agreed that exposure to sound leads to death. The causal steps between reception of sound and death are by no means known or agreed on, but the result is clear. The “+” signs at the bottoms of the boxes indicate how well the variables can be measured. The indicators between boxes show how well the “black box” nature of the transfer functions is understood; these indicators scale from “+++” (well known and easily observed) to “0” (unknown).
The bottom entry in each data level presents an indicator of how readily available or easily obtainable the critical data are.
Although it would be unrealistic to expect to acquire the data necessary to run such a model for all species of marine mammals, it will be important to model a representative sample of pinnipeds, baleen whales, and odontocetes with different hearing ranges and hearing anatomies (Ketten, 1994). The recently discovered particular sensitivity of beaked whales to mid-frequency tactical sonar (D’Amico, 1998; Evans and England, 2001; Jepson et al., 2003) demonstrates the necessity for both care and comprehensiveness in the selection of representative species. The 2000 National Research Council report provided a reasonable selection of species, sound types, and behavioral responses that could be used in the PCAD model (Box 3-1).
FINDING: Prior National Research Council reports (NRC, 1994, 2000, 2003b) identified high-priority subjects of research. The model proposed here requires the data and understanding that will become available on the fulfillment of the earlier National Research Council high-priority research recommendations.
RECOMMENDATION 1: The high-priority research identified by the National Research Council (1994, 2000, 2003b) should be completed. That research is essential for the model proposed in this report.
|
BOX 3-1 To move beyond the requirement for extensive study of each sound source and each area in which it may be operated (NRC [2000] recommended that), a coordinated plan should be developed to explore how sound characteristics affect the responses of a representative set of marine mammal species in several biological contexts (e.g., feeding, migrating, and breeding). Research should be focused on studies of representative species using standard signal types, measuring a standard set of biological parameters, based on hearing type (Ketten, 1994), taxonomic group, and behavioral ecology (at least one species per group). This could allow the development of mathematical models that predict the levels and types of noise that pose a risk of injury or behavioral disturbance to marine mammals. Such models could be used to predict in multidimensional space where temporary threshold shift (TTS) is likely (a “TTS potential region”) and TTS can be used as a threshold of potential risk of injury to the auditory systems. This coordinated plan can be used to set priorities for research required to determine measures of behavioral disruption for different species groups. Observations should include both trained (where possible) and wild animals (with attention to ecological validity). The results of such research could provide the necessary background for future environmental impact statements, regulations, and permitting processes. Groupings of Species Estimated to Have Similar Sensitivity to Sound. Research and observations should be conducted on at least one species in each of the following seven groups:
|
Signal Type. Standardized analytic signals should be developed for testing with individuals of the preceding seven species groups. These signals should emulate the signals used for human activities in the ocean, including impulse and continuous sources.
Biological Parameters to Measure. When testing representative species, several different biological parameters should be measured as a basis for future regulations and individual permitting decisions. These parameters include the following:
Modified from NRC (2000). |
All the transfer functions in the PCAD model may vary depending on the season and the species, location, age, and sex of the animal. Other external factors may also modulate the responses and effects. For example, behavioral responses that would be insignificant in a normal year may become biologically significant during an El Niño year. Behavioral responses, on individual and population scales, may differ between a stable population near environmental carrying capacity and a severely depleted population. Those types of modulations are considered in the model in two primary, but not exclusive, categories: time and energy budgets and homeostasis and allostasis.
The first stages in the PCAD model are relatively clear. In general, the characteristics of the sounds can be measured accurately. In some cases, the behavioral responses of the animals can be measured as well. Although mechanistic models that relate sounds to behavior are unavailable, such an understanding is not essential for management use of this model if the behavioral changes can be measured and predicted.
Dose-response studies are a good way to quantify the first transfer function, relationship of sounds to behavioral responses. For marine mammals, data are available on only a few sounds and a few behaviors in a few species. Observational and correlational studies can provide trend data, and expert-opinion modeling can provide at least a “lookup” table to serve as a surro-gate for this transfer function (Andelman et al., 2001). NOAA Fisheries has convened a panel of acoustic experts to survey the literature on mammalian hearing and the effects of noise and to draft noise-exposure criteria for five functional hearing groups of marine mammals (low-, middle-, and high-frequency cetaceans, pinnipeds in water, and pinnipeds in air) exposed to four sound types (single and multiple pulses and nonpulses). The criteria are based on individual sound-exposure events in which either the sound pressure (rms or peak) or the energy flux density exceeds one of two impact levels. The impact levels are tissue injury and behavioral disruption. Thus, the full matrix has 80 threshold criteria—the product of five animal groups, four sound types, two exposure metrics and two impact levels. The NOAA Fisheries panel has presented some portions of the criteria but has yet to complete a final draft. Some key elements of the criteria remain undetermined, particularly with regard to behavioral disturbance.
The second stage of the PCAD model attempts to evaluate how changes in behavior may affect life functions that are widely recognized as critical to population dynamics. With the exception of direct impact on life, the exact relationship of these functions to life-history characteristics is
largely unknown. Furthermore, the impacts of sound on these functions through behavior will be difficult to measure.
Time-scale integration is important in identifying impacts and determining relationships between changes in behavior patterns and resulting changes in life functions. Because most marine mammals have a diurnal cycle of activities, integration of short-term functional consequences over a duration of at least 24 hours may be appropriate and could be studied by using behavioral observations or electronic tagging methods. In addition, however, most marine mammals also have strong seasonal variations in behavior and physiology. As more data are accumulated, daily functional consequences might be summed over each season in relation to the expected duration of exposure to the specific sound of interest to evaluate daily and seasonal effects in a particular species.
The final stages of the model relate changes in life functions in individuals to impacts at the population level. There are at least two components to these final stages. The first is the most difficult—relating changes in life functions to changes in vital rates of individuals. If the link from life functions to age-specific vital rates is known, changes in population dynamics can be explored by using demographic analyses. Current demographic theory provides the capability to deal with vital rates not only on the basis of age but also in terms of biologically defined stages that reflect developmental, behavioral, or physiological properties of individuals.
A critical question is what population consequences should be identified as significant. The measure of population performance must integrate survival and reproduction across the lifespan. It should have implications for recovery, persistence, and extinction of populations. The most thoroughly investigated index is the population growth rate (modified by such adjectives as potential, intrinsic, and asymptotic). Demographic theory provides tools that explicitly link changes in the life cycle to changes in population growth rate. That makes demographic models a powerful tool for placing bounds on likely effects, for exploring the quantitative implications of hypothesized interactions, and for synthesizing what is currently known. Establishing acceptable population effects is a management question that has already received a good deal of attention. One example used for protecting marine mammals involves setting the potential number of marine mammals that can be removed from the population without endangering the population. The management criteria of this Potential Biological Removal (PBR) model (Taylor et al., 2000; also see Chapter 4) are:
-
Healthy populations will remain above the optimal sustainable population (OSP) numbers, as defined in the Marine Mammal Protection Act (MMPA), over the next 20 years.
-
Recovering populations will reach OSP numbers after 100 years.
-
The recovery of populations at high risk will not be delayed in reaching OSP numbers by more than 10% beyond the predicted time if there is no human-induced mortality.
The amount of information needed to map from sound to its population consequences is truly enormous. The PCAD diagram should be thought of not as the blueprint for an eventual universal model, but as a framework that clarifies where different kinds of information fit in and identifies processes that need study. Years of work will be required to accumulate data and develop models for the transfer functions between behavior and life functions, and between life functions and the vital rates. This report is essentially a status report and a roadmap for this critical long-term project of turning a conceptual model into a predictive model useful for science-based management of marine mammals and their exposure to sound. In the interim, techniques must be developed to use current information more effectively in making science-based management decisions. After discussion of the PCAD model, we propose (in Chapter 4) a means to achieve better management over a shorter timeline.
FINDING: A conceptual model, such as the PCAD model, is useful for clarifying the complex problem of acoustic-disturbance effects on marine mammal populations. Such a model can be used as a framework for identifying the cause-effect relationships necessary for determining consequences of disturbances. Data to complete this exercise are lacking and must be pursued from every available source.
RECOMMENDATION 2: A conceptual model, such as that described in this report, should be developed more fully to help to assess impacts of acoustic disturbance on marine mammal populations. Development of such a model will allow sensitivity analysis that can be used to focus, stimulate, and direct research on appropriate transfer functions.
In addition to research studies designed to evaluate reactions of marine mammals to noise, limited information is available from monitoring pro-
grams that are required of some activities that might “take” small numbers of marine mammals as defined in the MMPA. The incidental-harassment authorizations issued by the US government often contain the requirement for the operator to implement a program to monitor effects on marine mammals. For activities that produce intense noise, such as seismic surveys, the monitoring requirement often involves sighting animals from the vessel that is introducing the noise. Sighting surveys are also required by the United Kingdom and have been summarized in reports that identify avoidance reactions to seismic surveys (Stone, 2001, 2003). Few of those studies measured the acoustic stimulus from the activity as heard by the animal, and they typically scored easy-to-observe changes in behavior, such as respiration rate, time on the surface, duration of dive, change in swimming speed or direction, avoidance behavior, and aerial display. However, if those short-term measures are selected purely for ease of observation, it will often be difficult to link the responses to the functional categories described in the PCAD model, a link that is essential for extrapolating short-term measures to long-term effects that would alter some life function of an individual animal. Federal regulators for the last several decades have required monitoring programs instead of targeted research on the assumption that monitoring would detect developing problems. Monitoring programs, as implemented, have seldom provided the relevant data, suggesting that regulators and the regulated community should consider altering the balance of resources that they provide for monitoring versus research.
The impact of a behavioral reaction to sound depends on the number of animals affected in a population and on the duration and intensity of the reaction. The impact of avoidance reactions depends in the short term on the percentage of habitat reduction and on the ease with which animals can move to and use alternative habitat. Determining overall impact on the population requires estimation of
-
The range of the impact of individual sources in time and space.
-
The number of animals and the fraction of the population affected.
-
The total impact of all sources deployed.
-
The intensity of reaction of each animal.
-
The duration of the impact on each animal.
The presence of anthropogenic sound sources could have minimal effects on a healthy population that can relocate with minimal effort or
could be devastating to a small population that is living on the edge of its capabilities to survive where the sources affect its entire habitat (Box 3-2).
One of the few subjects of research that provide predictive models with connections from behavioral ecology through physiology to demography is how animals obtain and use energy. Behavioral ecologists have developed models to predict how animals maximize energy intake per unit of time as they forage (Stevens and Krebs, 1987). Physiologists and physiological ecologists have developed models to predict the baseline metabolic rates of animals and the metabolic costs of various activities. If a foraging animal takes in more energy than it uses for metabolism, it builds up an energy surplus that can be used for growth or reproduction. All large mammals have an initial period of sexual immaturity in which most surplus energy
|
BOX 3-2 The effects of seismic surveys on western gray whales (Eschrichtius robustus) off Sakhalin Island, Russia, illustrate the potential for anthropogenic sound to have a severe impact on a marine mammal population. The western gray whale is critically endangered, numbering about 100, and depends on the north-eastern Sakhalin Island feeding ground for most of its food intake. Weller et al. (2002) and Johnson (2002) report displacement of some whales during seismic surveys in 2001, and Johnson (2002) reports observations of gray whale behavior suggesting that they may have spent more time traveling and less time feeding during exposure to seismic signals, but aerial observations of feeding plumes were unable to detect any changes in feeding activity related to seismic activity. Disruption of feeding in preferred areas, especially in a small population in which many females (with and without calves) are already in poor condition and have long intervals between calf production (Brownell and Weller, 2002), could have major impacts on individual whales, their reproductive success, and even the survival of this critically endangered population (Weller et al., 2002). Observed changes in the distribution of individuals of this highly endangered population could be critical; deciphering their impact will require more detailed studies of prey distribution, foraging ecology, and energetics of these whales. |
reserves go to growth. The timing of the transition to sexual maturity is affected by the need to have grown to a particular point and the need to have amassed sufficient energy reserves to support the energy cost of the transition. Once a female is mature, the timing between her ovulations, the probability of successfully carrying a fetus to term, and the interval between offspring are all affected adversely by lack of energy resources. Those characteristics are all used directly in demographic models to estimate the reproductive rate of the population. During periods of starvation, the probability of survival may be affected if the animal’s metabolism exceeds energy intake for long periods. When foraging is not adequate, animals may abandon their young to conserve energy for their own survival. Limited energy resources may also make animals more vulnerable to other stressors (as discussed below in the section on allostasis). The various models that link foraging behavior, energy reserves, reproduction, and survival offer great promise for our proposed PCAD model, but more effort will be required to link the different submodels. The strength of research on energy budgets suggests that studies of effects of noise on foraging animals should focus on effects of disruption of time-energy budgets.
As noted earlier, repeated reports of unusual mass strandings of Blainville’s and Cuvier’s beaked whales show a correlation with naval maneuvers. The locations of whales with respect to the ships operating the sonars are unknown and cannot be reconstructed. However, the timing and spatial extent of the strandings suggest a possible risk of stranding for whales exposed to noise as low as 160 dB re 1 μPa. Current data on physiological or behavioral effects of well-studied marine mammals would not have suggested such a risk to poorly known beaked whales. The recent cases of the association of beaked whale strandings with naval sonar stimulated a review of prior records of beaked whale mass stranding events (Brownell et al., 2004; Taylor et al., 2004). This historical review indicated that mass strandings of beaked whales have occurred primarily subsequent to the introduction of mid-frequency tactical sonar in the early 1960s. However, caution must be exercised in these post hoc correlational studies. For example, when the radius of potential correlation extends to 500 km, as was the case with the strandings of Z. cavirostris and seismic in the Galapagos (Taylor et al., 2004), the potential for false positives increases proportionally. Therefore, there is a critical need for carefully designed and executed epidemiological studies to find potential problems as well as toxicological studies to evaluate precise dose-response relationships.
CURRENT DATA COLLECTION EFFORTS
In addition to basic research conducted primarily through universities and published in a host of peer-reviewed scientific journals, many data on marine mammals are gathered to fulfill regulatory requirements. For example, every permit application under the MMPA or the Endangered Species Act requires the applicant to provide a summary of the best available information on the status of the affected species or stock and on factors that affect the status. Permits for scientific research also contain many relevant data with respect to the habitat, behavior, physiology, or demography of the animals. A condition of many permits is the requirement to monitor the animals sighted, the time, location and oceanographic conditions of the observations, and the responses of the animals to the permitted activities. Federal agencies with responsibility for managing marine mammal populations conduct intramural research that often ends up as unpublished reports that contain valuable information. For example, NOAA Fisheries conducts surveys for assessing the status of marine mammal stocks. The agency publishes regular stock-assessment reports, but the sighting data would be extremely valuable for other purposes, such as predicting the species and number of animals that might be exposed to sound in a particular place and at a particular time.
Information from all these sources, with appropriate indicators of the sources, should be integrated into a common database. Peer-reviewed data and interpretation should be given the highest quality indicator. Other data sources should have appropriate quality indicators assigned. To facilitate the integration of data from many sources, federal agencies should establish standard data-reporting formats to be used in permit applications, permit reports, and research sponsored by other entities in fulfillment of permit requirements. Some federal support has been provided to begin the development of such integrated databases. Examples of such support are the Office of Naval Research Effects of Sound on the Marine Environment project and the Marine Resource Assessments by the Commander in Chiefs, U.S. Atlantic and Pacific Fleets.
FINDING: A wealth of data on marine mammals is collected in compliance with federal regulatory requirements. Such data are not collected in a manner that allows easy access or use beyond the original intent of their collection (such as permit issuance). A data-coordination effort could improve our knowledge of marine mammal distribution, behavior, and
population size; improve and standardize data used for regulatory processes; and greatly reduce the effort required of applicants for permits or authorization.
RECOMMENDATION 3: To assist in the development of the conceptual model, a centralized database of marine mammal sightings and their responses to anthropogenic sound in the ocean should be developed and should include
-
Published peer-reviewed papers in the scientific literature.
-
Government technical reports.
-
Data submitted to NOAA Fisheries and the US Fish and Wildlife Service in permit applications.
-
Data submitted by industry to the Minerals Management Service for regulating off-shore hydrocarbon exploration and production.
-
All relevant data accumulated by all federal agencies in the course of their research and operational activities, including monitoring.
To facilitate the integration of data from the various sources, federal agencies need to develop standardized data-reporting formats. Survey data should include locations where marine mammals were detected and the track lines when personnel were monitoring for marine mammals, regardless of whether any were sighted. All data entered into such an integrated database must be coded as to quality, and peer-reviewed data and interpretations should be rated highest.
DATA NEEDED TO DETERMINE PHYSIOLOGICAL RESPONSES TO ACOUSTIC STIMULI
Immediate behavioral responses are the easiest to observe, but the population consequences of sound will be modulated through physiological responses. The ear is the body structure most sensitive to acoustic input and is the site at which acoustic energy in the frequency range of hearing is most likely to have direct physiological effects. This report reiterates the recommendations of the 1994 and 2000 National Research Council reports to acquire more data on assessments of hearing characteristics such as
absolute sensitivity, masking, temporary threshold shifts, and temporal integration, and on the evaluation of behavior during exposure. However, the long-term effects of noise exposure on individuals can be best determined through the physiological integration that occurs and can be observed as indicators of cumulative stress.
Physiological Stress Effects
Anthropogenic sound is a potential source of stress in marine mammals, and it has been shown to increase blood pressure and catecholamine and cortisol concentrations in humans (Evans et al., 1995; Evans et al., 2001; Ising and Kruppa, 2004). Biomedical research on stress has provided a theoretical framework that can help scientists to conceptualize and ultimately measure the cumulative impact of multiple stressors on individual animals (McEwen and Stellar, 1993; Seeman et al., 2001). Application of the concepts, theories, and techniques to marine mammals could accelerate our understanding of the physiological effects of noise and other stressors on them.
Historically, the term stress has been used to refer to several concepts, including noxious stimuli, the physiological and behavioral coping responses of organisms to noxious stimuli, and the pathological states that result when the coping responses can no longer restore the body to a normal condition. Several attempts have been made to provide a less ambiguous terminology. For example, Romero (2004) refers to the three concepts listed above as stressors, the stress response, and chronic stress, respectively, and this terminology is used productively in the physiological and behavioral literature. An alternative terminology, which we will consider in some detail because of its conceptual integration with energy budgets and life-history events, has been offered by McEwen and Wingfield (2003). It focuses on the concept of allostatic load, which was adapted from the cardiovascular field and was introduced for more broad application and developed by McEwen and colleagues (McEwen and Stellar, 1993).
McEwen and Wingfield (2003) propose four terms—allostasis, allostatic state, allostatic load, and allostatic overload, that can be considered in relation to the life cycle and energy budget of any species (Figure 3-2). Although energy is a convenient currency to consider for illustrative purposes, it could be replaced in Figure 3-2 by any other resource vital to survival, such as a particular vitamin or mineral. Allostasis refers to the physiological and behavioral mechanisms used by an organism to support homeostasis (the
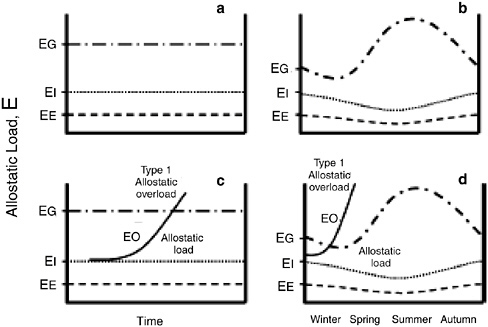
FIGURE 3-2 A framework for modeling energy requirements (E) of organisms during their life cycle. This energy requirement, E, includes all potential nutritional requirements, including energy itself. These separate and distinct requirements are represented more generally here for convenience, although essential components of nutrition could also be modeled separately. EE represents the energy required for basic homeostasis. EI represents the extra energy required for the organism to find, process, and assimilate food in ideal conditions. EG represents the amount of energy (in food) available in the environment (from Wingfield et al., 1998; Wingfield and Ramenofsky, 1999). (a) The three are represented as straight lines when environmental and physiological states do not vary over time. (b) The changes in the quantities have been adjusted to represent probable seasonal changes. EG would be expected to rise dramatically in spring and summer, when primary productivity is high, and then decline through autumn and winter, when primary productivity is low. In this scenario, EE would be lowest in summer, when ambient temperatures are highest. EI should be fairly constant (under ideal conditions) and should vary in parallel with EE. (c) EO represents additional costs incurred after a perturbation (such as a storm or anthropogenic disturbance) that increases costs above EE + EI. It represents the energy required to find food, process it, and assimilate nutrients in nonideal conditions. Allostatic load (see text) increases as EO persists. If EO exceeds EG, type 1 allostatic overload begins, resulting in an increase in plasma glucocorticosteroids. That usually triggers an emergency condition that results in altered physiology and behavior that reduces allostatic overload. (d) In more naturally fluctuating conditions, type 1 allostatic overload may occur first or more rapidly if a perturbation occurs during a season when the conditions are worse. If insults are permanent—such as those caused by even mild or moderate but persistent disturbances (e.g., Creel et al., 2002 for effects of snowmobile activity on wolves and elk), abnormally high densities of animals, increased pollutants (Porter et al., 1999), disadvantageous social status in some terrestrial species (Goymann and Wingfield, 2004; Sands and Creel, 2004), or individual differences in emotional or other vulnerable states (Sapolsky, 1994 for baboons; Cavigelli and McClintock, 2003 for rats)—overload will occur in most seasons and will be triggered readily even in seasons or conditions that are otherwise benign.
SOURCE: Adapted from McEwen and Wingfield, 2003.
stability of the physiological systems that maintain life) in the face of normal and relatively predictable life-history events, such as migration, mating, rearing young, and seasonal changes in resource availability; unpredictable events, such as decreases in oceanic productivity and increases in human disturbance; and more permanent handicaps, such as injuries, parasites, and contaminant loads. Examples of allostatic responses are the physiological changes that occur in lactating female mammals (Bauman, 2000) and the changes in metabolism, muscle morphology, and behavior that occur in migrating birds (Kuenzel et al., 1999).
An “allostatic state” is a sustained imbalance in the physiological mediators, such as various hormones, that integrate behavioral and physiological responses to changing environmental conditions. An allostatic state can be maintained for some time if environmental resources are sufficient. However, the cumulative result of an organism’s allostatic state is its “allostatic load.” The usual allostatic load results from the organism’s need to obtain enough food to survive plus any extra energy required for normal seasonal activities, such as migrating, molting, mating, and lactating. Animals can adapt to the extra demands within limits. However, if resources in the environment are insufficient (Figure 3-2d) or if other challenges—such as disease, human disturbance, or stressful social interactions—increase the allostatic load, the animal can no longer cope and will develop serious pathological conditions or die.
The concept of allostasis makes it clear that the effect of any given stressor will be contingent on multiple factors, including species, sex, nutritional and reproductive condition, and any other stressors currently affecting an animal. The closer an animal is to the condition of allostatic overload when subjected to an additional stressor, the more likely it is that
the additional stressor will have a deleterious impact. That is, the effect of a stressor often depends heavily on the context in which it occurs. The importance of context has also been shown by laboratory experiments that demonstrate that uncontrollable and unpredictable stimuli cause a greater stress response than controllable and predictable stimuli. For example, when two rats are given similar electrical shocks but only one can press a lever that terminates the shock for both, the rat that can terminate the shock has a dramatically lower hormonal response to the shock than the one that has no control over the length of the shock (Weiss, 1968).
Consideration of energy needs can also provide clues to the conditions in which marine mammals may be most likely to suffer allostatic overload. The following account of marine mammal energetics follows the recent review by Boyd (2002), who built on earlier reviews for pinnipeds (Lavigne et al., 1982; Costa, 1991, 1993) and cetaceans (Lockyer, 1981). Different species have different energy requirements and appear to balance their energy budgets by developing body sizes and life histories that match the distribution and abundance of their food. As body size increases, the period over which an animal must balance its energy budget lengthens. For example, the great baleen whales probably balance their energy budget on a 1-year cycle. They typically migrate to high latitudes during the summer to feed on krill or other seasonally abundant resources and store enough energy in the form of blubber for them to be able to fast for the rest of the year and reproduce in warmer but less productive tropical waters. Smaller species, such as most of the odontocetes (dolphins and porpoises), must balance their energy budgets on much shorter periods—months to days. Thus, energy considerations suggest that sound disturbance could severely affect the energy budget of baleen whales if it displaced them from their feeding grounds for a substantial fraction of the feeding season but would be less likely to have a serious effect on energy needs if it occurred in other circumstances, such as during migration, and merely displaced them temporarily from their normal migratory path.
The diverse lactation strategies of female pinnipeds provide a particularly good illustration of the relationships between body size, energetics, and behavior. Lactating pinnipeds nourish their pups from a food supply that may be near or very distant from the rookeries where they give birth. If sufficient food is available near the birth site for mothers to balance their own energy budget and provide for the pups, mothers make foraging trips during lactation. That strategy is followed by most of the otariids (fur seals and sea lions), which are relatively small for marine mammals (Costa,
1993). Larger species, such as most phocids (true seals), can forage over larger areas and use more dispersed prey resources. They can feed on lower-quality prey and need greater rates of prey consumption, but they can use a patchier prey distribution. For example, elephant seals feed thousands of kilometers from the sites where they give birth and, like the baleen whales, store enough energy in the form of fat to be able to fast while lactating (Costa et al., 1986; Boness and Bowen, 1996). Phocids appear to switch during lactation from foraging to fasting at a body mass of about 100 kg; harbor seals (80-100 kg) forage during lactation whereas gray seals (Halichoerus grypus, 130-180 kg) fast (Costa, 1991; Boyd, 2002).
The reproductive success of small pinnipeds that make repeated short-distance foraging trips during lactation is severely affected if they are unable to acquire normal amounts of prey. Evidence of that is provided by El Niño events, which occur at irregular intervals that tend to range between 2 and 7 years and result in greatly decreased productivity in the eastern tropical Pacific and greatly reduce the survival of pinniped pups (Trillmich and Ono, 1991). For example, during the 1982 El Niño, pup production was normal, but none of the pups survived the first 5 months after birth. In 1983, pup production was only 11% of normal, but survival of the pups returned to normal rates (Trillmich and Dellinger, 1991). Thus, energy considerations suggest that small otariid species could be affected rather quickly by anthropogenic noise close to their rookeries if it interrupted normal foraging whereas larger species that were not foraging during lactation would be more likely to meet their and their pups’ energy needs in the presence of a similar disturbance.
The physiological stress response is highly conserved and similar across vertebrate taxa (Wingfield and Romero, 2001). As an integrator of stresses, neuroendrocrinological responses include both direct and indirect effects of noise exposure. Physiological responses to stressors are initiated by activation of the hypothalamic-pituitary-adrenal axis, which results in the release of catecholamines and stress hormones, such as glucocorticoids, from the adrenal glands (McEwen, 2000). Because the extent of the stress response often correlates with the general health of an animal, measuring the response can serve as a general indicator of the current condition of an animal, reflecting its health, its energy allocation, and the effect of human disturbances on it. The promise of applying this approach in the field is illustrated by recent research on marine iguanas (Amblyrhynchus cristatus) in the Galapagos Islands (Romero and Wikelski, 2001, 2002). During El Niño years, iguanas had higher baseline corticosteroid concentrations dur-
ing famines. Handling of the iguanas also resulted in higher stress-induced corticosteroid concentrations than in normal years. Stress-induced corticosteroid concentrations in animals were good predictors of whether they would survive an El Niño event (Romero and Wikelski, 2001). Measurement of corticosteroid stress responses also showed that apparently low levels of oil contamination caused a strong hormonal stress response in iguanas. That response accurately predicted higher mortality over the next year among iguanas on oil-contaminated islands than on uncontaminated islands (Romero and Wikelski, 2002). A growing body of literature on terrestrial mammals has demonstrated sensitivity of glucocorticoids to sudden natural social stressors (e.g., Alberts et al., 1992 for wild baboons), to persistent natural stressors (e.g., Sapolsky, 1994), and to anthropogenic stressors (e.g., Creel et al., 2002 for wolves and elk).
Glucocorticoids may be part of the mechanisms by which behavioral effects are translated into altered rates of reproduction and mortality, and in other instances they will at least be indicators if not major players in the cascade of effects leading from behavior to survival and reproduction. As indicated above, it will be feasible in some cases to obtain fairly convincing evidence of the behavior-demography relationships with or without the physiological links between the two; but in most others, our greatest power will come from documenting behavior-glucocorticoid relationships in some studies and glucocorticoid-survival or glucocorticoid-reproduction relationships in others, as suggested by a number of studies already cited. Examples of an emerging picture of behavior-demography or behavior-glucocorticoid relationships from one of the best-studied wild large mammal species have been found in baboons (Box 3-3).
Physiological indicators of body condition and of pregnancy can be obtained from serum. Serum sampling of glucorticoid concentrations can also be used to obtain a physiological stress measure if the sample can be obtained before the stress of capture and sampling changes hormone concentrations in the blood. The maximal allowable time from capture to blood sampling is 2-3 min for small birds or rodents and 10-15 min for large monkeys. Determining the time for various marine mammals will identify the extent to which this technique can be applied usefully, at least in situations where capture for blood sampling is feasible.
In most cases, capture of marine mammals for blood sampling will be impossible. Instead, techniques will need to be developed to allow unrestrained blood sampling. Hill (1986) developed a package that could be attached to a freely diving Weddell seal and could take blood samples on
|
BOX 3-3 In baboons, a number of behavioral differences have been associated with altered demographics. Reduced travel time to foraging sites leads to a net positive increase in energy balance (Muruthi et al., 1991) and presumably thereby to the observed decreased age of maturation (Altmann et al., 1993), doubling of reproduction (halving of interbirth interval), and increasing offspring survival (J. Altmann, Department of Ecology and Evolutionary Biology, Princeton University, Princeton, NJ, unpublished data, 2004; S.C. Alberts, Department of Biology, Duke University, Durham, NC, unpublished data, 2004) despite increased rates of aggression (Altmann and Muruthi, 1988). Daughters and sons of low-status females mature later (Altmann et al., 1988; Alberts and Altmann, 1995). Larger baboon social groups are associated with decreased reproductive rates of lower-status females (Altmann and Alberts, 2003). Infants of females that are more social have higher survival (Silk et al., 2003). Effects of chronic or sudden behavioral differences on stress hormones have also been demonstrated in baboon studies. Among baboon males, either social status or degree of sociality affects glucocorticoid concentrations (Sapolsky et al., 1997), as does social style or recent winning or losing of fights (Sapolsky, 1994). Sudden social disruption by immigration of an aggressive male leads to high glucocorticoids in both sexes and in the aggressive immigrant itself (Alberts et al., 1992). Despite that body of data, however, studies linking small chronic differences in glucocorticoids to vital rates in such a large mammal are only now possible and are being conducted thanks to the recent techniques in fecal steroid analysis. |
a programmed schedule. More recently, sophisticated data logger tags have been attached to marine mammals to study their responses to anthropogenic sounds (Burgess, 2001; Johnson and Tyack, 2003). Data logging packages could be modified to incorporate blood sampling during controlled-exposure experiments (CEEs). Initial studies would likely need to be conducted on Weddell seals constrained to returning to an isolated hole to breathe. Eventually, the packages would benefit from the ability to take blood samples on a controlled basis and stabilize hormones for later
analysis or to conduct “on-board” blood-chemistry analysis to record responses of animals in situations less unusual than that of the Weddell seal.
Totally noninvasive, hands-off techniques of sampling glucocorticoids and other steroid hormone metabolites (such as estrogens, testosterone metabolites, and progestins) through collection of feces or urine are increasingly used for terrestrial mammals in situations or with species that make capture or any disruption to behavior intolerable (e.g., Wasser et al., 2000). The feasibility of feces collection from some marine mammals in the ocean has been demonstrated (Rolland et al., 2004); validation and calibration of the assays should have high priority (Buchanan and Goldsmith, 2004; Hunt et al., 2004). Preliminary studies measuring glucocorticoids in hair samples and up-regulation of stress-induced proteins in skin samples merit further development. Concentrations of fecal progestins are increasingly used in research and conservation for assessing pregnancy in terrestrial mammals. Application to marine mammals to evaluate pregnancy rates and fetal or early infant loss may be relatively straightforward (Larson et al., 2003) when the requisite samples can be obtained.
FINDING: Measurements of glucocorticoids and other steroid hormone metabolites in terrestrial vertebrates have proved to be good indicators of pregnancy, allostatic overload, and mortality risk posed by current and new stressors.
FINDING: Continued development of more-sophisticated data logger tags is necessary to advance the study of marine mammal responses to anthropogenic sounds. Data logging packages should be modified to incorporate blood sampling during controlled-exposure experiments (CEEs).
RECOMMENDATION 4: The use of glucocorticoid and other serum hormone concentrations to assess stress should be developed, validated, and calibrated for various marine mammal species and age-sex classes and conditions. Dose-Response curves for those indicators as a function of sound characteristics need to be established. Development of a sampling package that could take blood samples on a controlled basis and stabilize hormones for later analysis or process samples “on-board” for corticosteroids at various stages of a CEE would be invaluable for determining the stress that the sound is producing. The use of fecal sampling to measure condition or stress needs
to be investigated further and developed. Research efforts should seek to determine whether reliable long-term stress indicators exist and, if so, whether they can be used to differentiate between noise-induced stress and other sources of stress in representative marine mammal species (this recommendation was also made in NRC, 2003b).
Toxicology
The concept of allostasis provides a framework for understanding how anthropogenic noises that at first appear insignificant could, with repeated exposure or in combination with other stressors, compromise an animal’s survival and reproduction. Recent research in toxicology has provided cautionary examples of how the combined actions of apparently safe individual factors can have serious unforeseen consequences. For example, a mixture of several agrochemicals at concentrations commonly found in groundwater across the United States affected immune, endocrine, and nervous system function in wild deer mice (Peromyscus maniculatus) and outbred white mice when consumed for 14-103 days (Porter et al., 1999). In this 5-year study with a full factorial design, numerous deleterious changes occurred in response to mixtures of aldicarb (an insecticide), atrazine (a herbicide), and nitrate (a fertilizer) at low concentrations, but the changes were rarely seen when the compounds were tested individually at the same concentrations. In another study, a commercial herbicide containing a mixture of 2,4-dichlorophenoxyacetic acid (2,4-D), mecoprop, dicamba, and several inert ingredients led to a U-shaped dose-response curve for litter size in mice; the lowest dosages of the mixture caused the greatest decrease in the number of live pups born (Cavieres et al., 2002). Such studies demonstrate that multiple stressors can interact in complex and unforeseen ways to produce adverse effects on living organisms.
DATA NEEDED TO DESCRIBE MARINE MAMMAL POPULATIONS
To understand the behavioral effect that a sound may have in a given place and at a given time, it is necessary to be able to answer the following questions:
-
What species are present?
-
What is their distribution?
-
What are their grouping patterns?
-
What activities are they engaged in?
-
How is each activity disrupted by sound?
NOAA Fisheries has collected and analyzed data on sightings of marine mammals to assess the status of different populations, and extensive sighting data are available from other sources, but the data are not available in a form that allows the prediction of the number of animals likely to be exposed to a sound in a given place and at a given time. Grouping patterns are important because if animals live in groups an average density will not yield a correct probability of the number of animals exposed.
Even fewer data are available on how marine mammals use different areas. That data gap could be addressed by completing basic behavioral ecological studies of marine mammals in the wild. To understand the biological significance of behavioral disruption, a greatly accelerated program is needed for studying the behavior and ecology of marine mammals in the wild, with a focus on how variation in behavior may affect probabilities of survival, growth, and reproduction in different ecological settings. The first recommendations for research in the 1994 and 2000 National Research Council reports were to study the behavior of marine mammals in the wild. Ten years after the 1994 report, a major increase in support of research to fill this critical data gap is still needed. The urgency of a research program in this field is highlighted by the PCAD model.
INDIVIDUALS TO POPULATIONS: USING MODELS TO IMPROVE UNDERSTANDING
In the PCAD model, there are at least some data that link sounds to behavioral responses of individuals. The connection between individuals and the population is much more speculative. There are good reasons for this lack of data. Most effects on life functions are separated in time and space from the immediate behavioral responses to sounds. Thus, if later observations identified life-function activities outside the normal range, it would be difficult to relate them to prior exposure to sound. Furthermore, our current understanding of the behaviors associated with most life functions is incomplete. For example, we do not yet fully understand normal
ranges of the behaviors, so effects may not be detected even if they are observable. As noted previously, there is almost no understanding of how changes in any of the life functions lead to changes in vital rates.
The only way to build a bridge from the individual to a population is modeling of some kind. No single model will serve the purpose, but a number of modeling exercises could help to integrate what is known tactically (in the short term) and to structure strategic research in the longer term. We consider here the types of modeling that might prove helpful and the expectations for each.
Uses of Models: Prediction and Exploration
The use of models for prediction is most successful when a well-established understanding of the processes and a good database for parameterizing the model exist. With respect to linking individual to population effects in marine mammals, both understanding and data are lacking (Figure 3-3, Area 4). Predictive modeling to determine the population effects of noise on marine mammals is therefore not now an option.
The determination of an appropriate modeling technique depends on the information and understanding available (Starfield and Bleloch, 1991). A schematic representation can be used to describe possible approaches (Figure 3-3). Area 1 is the region of good data but little understanding; statistical tools are applicable and can be used to perform an exploratory data analysis (sensu Tukey, 1977) to search for patterns and relationships. Area 3 is the region of good data and good understanding where predictive modeling has the best chance of success; well-established paradigms and modeling approaches can be used with confidence and are backed by experience and theory.
If either data quantity or quality is poor, a modeler is restricted to Areas 2 and 4, referred to as “data-limited.” In Area 2, there is good understanding of the processes and structure of the problem; in Area 4, that understanding is weak. Marine mammal data are still sparse, so this report is concerned mainly with Areas 2 and 4. Issues in these two areas present the modeler with two daunting challenges:
-
Despite the lack of data and understanding, a management or policy decision must be made. How can modelers help to make the best scientific decisions under these circumstances?
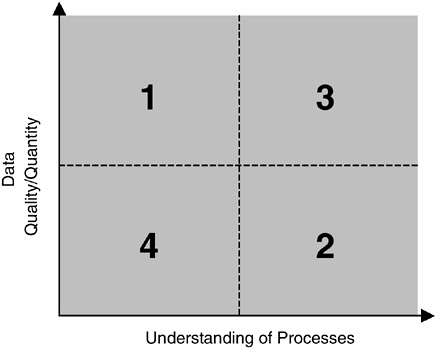
FIGURE 3-3 Classification of modeling problems.
SOURCE: Holling, 1978.
-
How can models be used to exploit available data to improve understanding and, in turn, identify data that are critically needed? In other words, how do we progress from Area 4 toward Area 3?
Scientists and modelers are often uncomfortable dealing with these issues. Some believe that modeling should be confined to Area 3; others push ahead and try to use predictive models in an area where prediction is, to say the least, risky. Starfield and Bleloch (1991) suggest that Areas 2 and 4 require a different paradigm in which models are used tentatively to explore alternative hypotheses, speculate on possible outcomes given whatever data are available, and then cautiously reach some conclusions—even if they are only conclusions about future research needs. The way that they propose using models in Areas 2 and 4 is philosophically akin to Tukey’s exploratory data analysis; that is why the term exploratory modeling is used.
Exploratory models can be used as tools for synthesizing what is known, explaining what may be happening, or perhaps guiding research or management. In all cases, if modeling is to serve a useful function, it is essential that the purpose or objective of each modeling exercise is clearly defined, the model is focused ruthlessly on the objectives, and all involved with the modeling exercises have a pragmatic appreciation of the power (or lack thereof) of whatever modeling paradigm is developed. These are some of the objectives for a suite of modeling exercises:
-
Objective 1: To bound the problem or look for significant thresholds. In the introduction, it was stated that it is not clear whether noise has a second-order effect on populations or whether what has so far been observed is only the tip of the iceberg. Models could help to categorize the likely effect of specified noise doses on different populations. That type of modeling exercise would be useful even if it produced the limited result that “Dose X is unlikely to have a measurable effect on a population with these characteristics, but it could have a measurable effect on a population with those characteristics.”
-
Objective 2: To speculate on the likely outcome of hypothesized interactions. The objective is to take a word model (such as “disruptions of courtship in species X will have a significant impact on the recovery of the species”) and tease out the implications quantitatively. The modeling would perforce be speculative, but there is value to exploring explicitly which assumptions and which sets of parameter values support the hypothesis. To quote Samuel Johnson, “That, sir, is the good of counting. It brings everything to a certainty which before floated in the mind indefinitely.”
-
Objective 3: To synthesize and organize what is currently known. For example, we know that responses can be situation-specific. It has already been noted that the responses of migrating gray whales depend on whether a low-frequency active source is in the migratory path or a few kilometers seaward of the migratory path even though the received levels were similar (Tyack and Clark, 2000) and that the responses of beluga whales in the high arctic to the initial seasonal exposure to an icebreaker are stronger and more prolonged (Cosens and Dueck, 1988; Finley et al., 1990) than the responses of beluga in the same region to the icebreaker a few days later (Finley et al., 1990) and the responses of Bristol Bay, Alaska, beluga to
-
direct harassment attempts (Fish and Vania, 1971). It is likely that more is known than appears to be known—synthesis will produce more than the sum of the parts. There are three related objectives:
-
To focus the attention of disciplinary scientists on how their knowledge and data can be combined or otherwise used to address the problem.
-
To identify gaps in data and knowledge and explore what one minimally needs to do to fill the gaps.
-
To provide guidelines for data collection and monitoring.
-
-
Objective 4: To develop a conceptual framework for management guidelines. Models can be used to organize and improve management guidelines, such as described for the Potential Biological Removal management regime described in Chapter 4.
A number of alternative modeling paradigms and constructs could fit with some of those objectives. For example, the age-structured demographic models (Caswell, 1989) usually used for predictive modeling could be used in an exploratory way to help to bound the problem and establish thresholds for different species. It is difficult to be specific about suitable paradigms or the design of a model until the precise objectives of an exploratory modeling exercise are spelled out, but two additional potential approaches are offered: individual-based models (IBMs) and categorical or qualitative models. In the next sections of this report we describe three modeling approaches and two additional tools that might prove helpful.
Demographic Models
The most well-developed and widely used approach to population modeling is that of age-structured demographic analysis. A demographic model is one that categorizes individuals into groups based on biological characteristics relevant to their survival and reproduction. In classical demography those groups were based on age (and implicitly on sex), but it is now known that other criteria, such as maturity, reproductive status, physiological condition, and spatial location may be more important (e.g., Caswell, 2001). Stage-structured models are most commonly expressed as population projection matrices, which may include environmental
stochasticity, demographic stochasticity, density-dependence, and spatial structure (Caswell, 2001).
Demographic models can be analyzed to obtain measures of population growth and structure, probabilities of extinction or quasi-extinction, and other measures of population performance. They employ a well-developed perturbation theory that permits calculation of the effect of changes in the vital rates on those measures of performance; this makes them particularly suitable for the exploration of thresholds and the effects of interactions. Matrix population models have a well-developed connection with statistical methods for parameter estimation, especially from observations of known individuals (e.g., Nichols et al., 1992; Fujiwara and Caswell, 2001, 2002a,b; Caswell and Fujiwara, 2004). These methods can incorporate measurements of individual animal condition into estimates of the vital rates. Recent research has explored Bayesian methods for parameter estimation in these models (Gross et al., 2002); such methods are particularly suitable for analysis of uncertainty.
Matrix models have been used for demographic analysis of killer whales (Brault and Caswell, 1993), humpback whales (Barlow and Clapham, 1997), right whales (Fujiwara and Caswell, 2001; Fujiwara, 2002), and harbor porpoises (Caswell et al., 1998) as well as various species of seals (e.g., Heide-Jorgensen et al., 1992; York, 1994; Kokko et al., 1997; Lalas and Bradshaw, 2003).
Although demographic models could be used to make predictions, their most common use is to explore the consequences of various biological processes in the face of unknown data. In two cases, the California condor (Mertz, 1971) and the Everglades kite (Nichols et al., 1980), only the most fragmentary data were available—both studies used demographic models to place bounds on population growth, to speculate on the outcome of hypothesized interactions, and to synthesize sparse data. More recent examples of exploratory use of demographic models include the exploration of management strategies for sea turtles (Crouse et al., 1987), the exploration of bycatch effects in harbor porpoise (Caswell et al., 1998), and exploration of research priorities for the sooty shearwater (Hunter et al., 2000).
Individual-Based Models
In an Individual-Based Model (IBM), the computer program is designed to simulate virtual individuals in a population, often from birth to death. Each individual carries a set of attributes or markers that describe
the state of the individual. They can include demographic factors, such as age and sex; energy factors, such as weight, stomach fullness, and diet composition; location descriptors, such as latitude and longitude; and behavioral descriptors, such as reproductive status, dive intensity, and dominance role. Such programs as Tagging of Pacific Pelagics (Block et al., 2003) provide data on movement patterns in relation to oceanographic features and seasonal patterns of movement essential for constructing a valid IBM for these species. IBMs have been constructed for species in a variety of habitats (Grimm, 1999).
For example, an IBM has been designed to compare the effects of alternative trophy-hunting strategies (Whitman et al., 2004). It describes each male lion (at any time step) in terms of his age, social status (cub, nomad, or pride lion), associates (like-aged cubs, fellow nomads in a nomadic group, and fellow males in a pride coalition), and spatial position (which pride a cub is born into, which territories a pride coalition controls and patrols, and which territories a nomadic group is temporarily visiting). Those attributes enable one to simulate such processes as competition between neighboring pride males, territorial battles between resident pride males and visiting nomads, and infanticide when pride coalitions are replaced—all essential to an understanding of how trophy hunting might affect the size and structure of a lion population. Some other examples are the modeling of deer and Florida panther (Puma concolor coryi) populations in the Florida Everglades (Abbott et al., 1997) and of walleye pollock (Theragra chalcogramma) in the western Gulf of Alaska (Hermann et al., 2001).
IBMs can be used for purposes similar to those of structured demographic models and can also directly address questions about the interaction between, for example, behavior of animals in relation to a source and the resulting acoustic exposure, behavior and reproduction, or behavior and growth. They offer a direct venue for considering the effects of noise on marine mammal individuals and populations. They can accommodate the kinds of data that are now becoming available on the relationships between behavior and acoustic exposure in a direct and comprehensible fashion. For example, the Acoustic Integration Model (AIM; Frankel et al., 2002) models the location and dive behavior of simulated marine mammals swimming near a modeled acoustic source. An acoustic-propagation model is used to predict the exposure of the simulated animals and can program different response strategies of the animals for the simulated source. It has been used to predict the exposure of animals with different response patterns to sources with different modes of operation, monitor-
ing, and mitigation; and it can help in selecting alternatives that minimize effects on marine mammals while maximizing operational effectiveness of the source.
Categorical or Qualitative Models
The links or transfer functions between changes in the behavior of individuals, effects on life functions, and effects on vital rates (survival and reproduction) of a population in the conceptual model (Figure 3-1) have been identified as ones on which there is little information. However, some progress might be made by combining whatever is known with an understanding of the behaviors and pressure points in different species to derive a qualitative ranking of the strength of a link. An example of behaviors and pressure points would be a marine mammal with an “income” breeding strategy (Costa, 1993) of intensively nursing newborn pups in bouts separated by extensive time at sea to replenish reserves. It can be argued that a reduction in the feeding success of mothers during that period will have a more severe effect on pup survival than an equivalent reduction in feeding success in a capital breeder (an animal that relies on stored energy to survive the breeding season).
A categorical or qualitative model would characterize effects in such terms as low, moderate, and severe. Such a model may separate the consequences of an effect from the probability that it will occur. It could be developed with a combination of available information on marine mammals, information on comparable nonmarine mammals selected on the basis of life-history scaling or body-size scaling, first principles, and expert opinion (Morgan and Henrion, 1990; Goodwin and Wright, 1991; Meyer and Booker, 1991; Anderson, 1998; Andelman et al., 2001).
The Scientific Committee on Antarctic Research (SCAR, 2004) created a series of risk-assessment matrices for different acoustic sources in Antarctic waters. The cells of a likelihood-consequences matrix indicated whether there was a potential risk to an individual or the population. One conclusion of this analysis was that the risks associated with the use of most scientific acoustic equipment in the Antarctic were less than or comparable with the risk associated simply with the passage of the research ship through Antarctic waters.
Categorical or qualitative models might serve two purposes: to create a structure for encouraging biologists to make the best determinations they can and to explore the feasibility of developing tactical management
strategies akin to the PBR model (see Chapter 4). Essential components of such a model would be estimates of the reliability of every categorization in the model and explanations of how each categorization was reached. The models would provide a structure for further refinement and, like the proposed IBM and demographic modeling exercises, help to identify gaps in knowledge. The key point to make is that modeling exercises like this can lead to robust management approaches, as the PBR model demonstrates, even when knowledge is incomplete.
Expert Opinion
Data on many links in the chain from acoustic stimuli to population effects on marine mammal populations are sparse or lacking. Therefore, regulators such as NOAA Fisheries and the US Fish and Wildlife Service (FWS) may often find it necessary to rely on expert opinion regarding the probable effects of specific activities until more data accumulate. Although the use of expert opinion does not necessarily produce an accurate result (experts can be wrong, especially when data are lacking), it does provide a structured, well-documented basis for decision-making that often with-stands legal scrutiny. Precedents for the use of expert opinion to evaluate risk in a conservation context are provided by the US Department of Agriculture Forest Service’s extensive reliance on expert opinion for population-viability assessments under the National Forest Management Act (Andelman et al., 2001) and FWS’s increasing use of expert opinion for making listing decisions under the Endangered Species Act (ESA; J. Cochrane, US Fish and Wildlife Service, personal communication, 2004). Because eliciting and using expert opinion are complex tasks beset with pitfalls for the inexperienced, any use of expert opinion should follow established procedures detailed in the substantial scientific literature on the subject (Morgan and Henrion, 1990; Goodwin and Wright, 1991; Meyer and Booker, 1991; Anderson, 1998; Andelman et al., 2001) to avoid bias and increase credibility.
Risk Assessment
Evaluating the effects of noise on marine mammal populations is a problem in risk assessment. Previous National Research Council reports have considered the general process of risk assessment by the federal government (NRC, 1983) and risk assessment in relation to contaminants and
human health (NRC, 1993). Uncertainty is always a prominent feature of risk assessment, and uncertainty regarding the probable effects of human activities on marine mammals is not limited to the effects of noise but rather is a pervasive problem, which can be addressed using population models (Caswell et al., 1998; Ralls and Taylor, 2000).
Risk assessment can be combined with decision analysis to make management decisions in the face of uncertainty (Harwood, 2000, 2002). The general approach is discussed in detail with respect to making decisions under the ESA in an earlier Research Council report (NRC, 1995). There are two main categories of errors in judging the effects of human activities on natural resources: we may conclude that a risk is great when it is not, which leads to overprotection and unnecessary economic loss, or we may conclude that a risk is small when it is not, which leads to underprotection and avoidable loss of a valued resource. It is impossible to minimize simultaneously the probability of making those two types of errors, and common statistical practices of hypothesis-testing may lead to a systematic bias against the welfare of species or populations that are in need of protective action (NRC, 1995, Chapter 8). Analyzing risks with the framework of decision analysis increases the probability that all types of errors and their consequences are adequately considered.
Advances in technology have enabled the use of computer-intensive methods in risk assessment (e.g., Slooten et al., 2000; Taylor et al., 2000). If relevant data on marine mammals are lacking, this kind of simulation approach can benefit from the use of data on other species selected on the basis of life history, ecology, or body size (e.g., Caswell et al., 1998). Bayesian decision theory, which allows choices among more than two decisions, offers many advantages and is increasingly recommended for use in risk assessment related to natural-resources management (Ludwig, 1996; Taylor et al., 1996; Wade, 2000).
FINDING: Focused effort is needed on a modeling exercise that should include demographic models, IBMs, and categorical modeling. Such an effort should start with, and calibrate against, expert opinion and should incorporate such characteristics as
-
An aim to pull together what is known—in different ways, from different disciplines—and to assess both the importance and the degrees of uncertainty associated with the information.
-
The use of tactical models, with the objective of probing how successfully current knowledge could be applied.
-
The use of structured models to test hypotheses.
-
The use of models to identify crucial gaps in knowledge. (A gap in knowledge is not just something we do not know; it is something we do not know and need to know if we are to meet our objectives.)
-
An aim to encourage interdisciplinary synthesis and provide a structure for it.
-
The requirement that all modeling efforts be explicit about uncertainty and its consequences.
-
A similar requirement that all models clearly state their limited purpose and that both their strengths and their shortcomings be evaluated.
-
A risk assessment for the species being modeled if the model is to be used for management decisions.
RECOMMENDATION 5: Several marine mammal species for which there are good long-term demographic and behavioral data on individuals should be selected as targets of an intensive exploratory modeling effort that would develop a series of individual-based models and stage- or age-structured demographic models for the species as appropriate. NOAA Fisheries should bring together an independent, interdisciplinary panel of modelers and relevant empirical scientists that would meet periodically to pursue the modeling effort collaboratively in an iterative and adaptive manner with the long-term goal of developing tools to support informed, practical decision-making.
Species should be chosen on the basis of how extensively they have been studied, and the models should concentrate on populations (or sub-populations) in which individual animals are known and have been tracked for some time. The different species should be chosen to span an array of life-history patterns (such as feeding and breeding strategies). The objectives of the modeling exercises should be to speculate on how harassment or acoustic injury of individuals might affect populations and to identify gaps in data and understanding. The exercises should also explore links between IBMs and demographic analyses for the same population; each should be
able to inform the other in important ways (see Caswell and John, 1992). Some candidate populations for such a study are the Puget Sound killer whales (Krahn et al., 2002), the North Atlantic right whales (Eubalaena glacialis; Waring et al., 2003), bottlenose dolphins in Sarasota Bay (Wells, 2003), the gray seals of Sable Island (Austin et al., 2004), and the northern elephant seals of Año Nuevo Island (LeBoeuf et al., 2000). All those have been studied extensively, and individual animals have been identified and resighted over multiple years. For most of the populations, the demographics are well defined; in some, the effects of major environmental stressors, such as an El Niño or the North Atlantic Oscillation, have been observed (Fujiwara and Caswell, 2001; Greene and Pershing, 2004). Such complex interdisciplinary modeling has been undertaken by the National Center for Ecological Analysis and Synthesis at the University of California, Santa Barbara.


































