
8
Evolutionary Animation: How Do Molecular Phylogenies Compare to Mayr’s Reconstruction of Speciation Patterns in the Sea?
STEPHEN R. PALUMBI* AND H. A. LESSIOS†
Ernst Mayr used the geography of closely related species in various stages of increasing divergence to “animate” the process of geographic, or allopatric, speciation. This approach was applied to a wide set of taxa, and a seminal paper by Mayr used it to explore speciation patterns in tropical sea urchins. Since then, taxonomic information in several of these genera has been augmented by detailed molecular phylogenies. We compare Mayr’s animation with the phylogenies of eight sea urchin genera placed by Mayr into four speciation groups. True to Mayr’s predictions, early-stage genera have on average lower species divergence and more polytypic species than genera in later stages. For six of these genera, we also have information about the evolution of the gamete recognition protein bindin, which is critical to reproductive isolation. These comparisons show that later-stage genera with many sympatric species tend to be those with rapid bindin evolution. By contrast, early-stage genera with few sympatric species are not necessarily earlier in the divergence process; they happen to be those with slow rates of bindin evolution. These results show that the rate of speciation in sea urchins does not only depend on the steady accumulation of genome divergence over time, but also on
the rate of evolution of gamete recognition proteins. The animation method used by Mayr is generally supported by molecular phylogenies. However, the existence of multiple rates in the acquisition of reproductive isolation complicates placement of different genera in an evolutionary series.
Ernst Mayr built an argument for the way speciation occurred based on the geographic patterns of variation among closely related species (Mayr, 1942, 1963). He showed that there was a hierarchy of species descriptions that could be ordered in a series of increasing complexity. Some descriptions pertained to recently diverged species, with morphologically identical populations inhabiting a continuous range. Other descriptions were of polytypic species, those with slightly differentiated populations inhabiting different parts of the range. Further along the speciation axis were superspecies, taxa with morphologically distinct, allopatric populations. Still later in the series, Mayr identified groups of related species in which some taxa were sympatric. The trajectory from homogeneous populations to overlapping sympatric species encompassed Mayr’s view of the process and pacing of geographic speciation. In addition to describing these separate elements, a major contribution by Mayr was to order these elements in a series. The elements thus served as separate frames in an evolutionary animation that sped up the slow process of speciation so that it could be viewed and studied by biologists.
The geographic distributions of species, subspecies, varieties, and slightly divergent populations constituted the database in Mayr’s analyses. He made the implicit assumption that the genetic and evolutionary divergence of these groups increased from population- to species-level distinction and used morphological differentiation as a proxy for evolutionary time. Mayr established sister-species relationships on the basis of morphological similarity and included a tacit phylogenetic framework for his animations based on the best information available at the time.
One difficulty faced by Mayr was that few concrete phylogenetic analyses were available during the development of these ideas. Since that era, molecular phylogenies have made it possible to obtain a statistically robust view of phylogenetic relationships, divergence order, and sister-species status (Hillis et al., 1996). Molecular phylogenies can also give an indication of the timing of divergence events through the application of molecular clock calibrations. Even without precise time calibrations, the record of the order of divergence of taxa permits an examination of the causes of each splitting event. Lastly, phylogenies can provide objective data on divergence levels to test predictions of Mayr’s evolutionary reconstructions.
For all these reasons, molecular phylogenies can contribute substantially to an updated view of evolutionary animation. To what extent do modern phylogenies confirm or reject the orderly animations suggested by Mayr’s analysis? Mayr’s ideas were shaped primarily from his studies of bird systematics, and, indeed, subsequent use of molecular phylogenies showed his insistence on the primacy of the allopatric mode of speciation to be correct for this group [Barraclough and Vogler, 2000; Edwards et al., “Speciation in Birds: Genes, Geography, and Sexual Selection” (Chapter 6)]. Mayr, however, was also interested in applying his view of speciation to all animals, both terrestrial and marine, and his evolutionary animations included more than bird species. Some of the nonavian genera that interested him have been examined extensively for phylogenetic relationships by using molecular tools.
In 1954, Mayr published a paper on the geographic speciation of marine taxa by focusing on the differentiation and geography of species in 20 genera of tropical sea urchins (Mayr, 1954). His goal was to examine the generality of his ideas about species formation by extending them to groups with ecology very different from that of birds. This goal was an important one for Mayr. In his view, even the most comprehensive recounting of speciation mechanisms was wanting if it applied to only a single taxon. Mayr’s conclusion at the end of this analysis was that geographic speciation applies equally well to marine species, such as sea urchins, as it does to birds, mammals, and insects.
Mayr took advantage of the publication of a complete monograph of sea urchin taxonomy by Mortensen (1928–1951) to examine the stages of speciation represented by each genus, restricting his attention to genera with species that inhabited the shallow seas in the West Indies. He divided the genera into four groups. Groups 1 and 2 included genera that had strictly allopatric species, the only difference between the groups being that group 2 genera included nontropical species. These two groups represented the earliest step in species formation, with a surfeit of polytypic species, and, presumably, low divergence among allopatric sister species. Group 3 represented the next step in speciation, in which congeneric species had sufficient time to develop genetic divergence, thereby allowing formerly allopatric sister species to invade each other’s ranges. Thus, these genera were presumed to show higher levels of divergence and the beginnings of sympatric overlap. Finally, group 4 genera were those in which the species were so old that current geographic ranges had nothing to do with speciation pattern. Whatever record of species formation there may have been among these ancient species was gone.
Although Mayr had the advantage of using the most comprehensive and up-to-date treatment of sea urchins provided by Mortensen’s monograph, this treatment was almost strictly taxonomic. Within genera, there
was little or no information of the relationships among species. This lack of systematics prevented Mayr from basing any of his group designations on sister-species relationships. However, in the past 10 years, detailed molecular phylogenetic studies have been completed for six genera in Mayr’s original list and two others that do not occur in the West Indies. In this paper, we collate these phylogenies and use them to test general predictions from Ernst Mayr’s reconstruction of sea urchin speciation patterns. General predictions across all genera in the study include familiar tenets of allopatric speciation. Testing these predictions with molecular data reveals a considerable concordance with Mayr’s animations but also shows some surprises about the way speciation proceeds in sea urchins.
MATERIALS AND METHODS
We compiled molecular data for variation in the mitochondrial cytochrome oxidase I (COI) genes for Mayr’s groups 1 and 2 genera Tripneustes (Lessios et al., 2003a), Eucidaris (Lessios et al., 1999), and Lytechinus (Zigler and Lessios, 2004); group 3 genera Echinometra (McCartney et al., 2000; Landry et al., 2003) and Diadema (Lessios et al., 2003b); and group 4 genera Strongylocentrotus (Biermann et al., 2003), Arbacia (Metz et al., 1998), and Heliocidaris (Zigler et al., 2003). Phylogenetic relationships are taken directly from the original analyses. Data for the gamete recognition molecule bindin are available for six of these eight genera: Tripneustes (Zigler and Lessios, 2003a), Lytechinus (Zigler and Lessios, 2004), Echinometra (Landry et al., 2003; Metz and Palumbi, 1996; McCartney and Lessios, 2004), Strongylocentrotus (Biermann, 1998), Arbacia (Metz et al., 1998), and Heliocidaris (Zigler et al., 2003). In general, phylogenetic relationships at COI and bindin are concordant. Major exceptions are the positions of Lytechinus williamsi and Echinometra insularis in their respective genera (Landry et al., 2003; Zigler and Lessios, 2004). In Strongylocentrotus, we have included the monotypic genera Allocentrotus and Hemicentrotus because phylogenetic analysis places the species in these genera firmly within the genus Strongylocentrotus (Biermann et al., 2003). We used Kimura two-parameter genetic distances based on COI comparisons. In Diadema, combined ATPase 8 and 6 and COI sequences were used. Distances were compiled for sister species (two or more species that split at the tip of a branch or a species that forms an outgroup to a clade of closely related species). Allopatric neighbors are defined as allopatric species that are not separated by an obvious, insurmountable geographic barrier, such as a land mass.
We characterize bindin evolution as “fast” if the nonsynonymous-to-synonymous substitution ratio in at least an ≈100-bp “hotspot” region of the gene is >1 and if there are several codons on which positive selection
has acted. By this definition, bindin is considered to be evolving quickly in the genera Echinometra, Strongylocentrotus, and Heliocidaris (Biermann, 1998; Landry et al., 2003; Metz and Palumbi, 1996; Zigler et al., 2003) and slowly in Lytechinus, Arbacia, and Tripneustes (Metz et al., 1998; Zigler and Lessios, 2003a, 2004). Although positive selection could not be statistically demonstrated for any comparison between species of Lytechinus, we consider the bindin of L. williamsi and Lytechinus variegatus variegatus as rapidly evolving, because its divergence is much higher than expected from comparisons of COI between these species. The COI haplotypes of L. williamsi and L. variegatus are intermingled, whereas bindin sequences are reciprocally monophyletic, suggesting the possibility of selection on the latter (Zigler and Lessios, 2004).
Estimation of time since species separation from COI divergence assumes a molecular clock, which is calibrated by the complete closure of the seaway between the eastern Pacific and the Atlantic by the isthmus of Panama at ≈3.1 million years ago (Coates and Obando, 1996). In all comparisons involving tropical genera, Atlantic and Pacific species are included. The divergence between clades presumably split by the isthmus provides a rough calibration of a molecular clock for each genus, allowing us to place a general time frame on species divergence patterns.
RESULTS
Groups 1 and 2
Polytypic species and low levels of genetic divergence between allopatric entities within an ocean basin are common in the genera Tripneustes, Lytechinus, and Eucidaris (Fig. 8.1). For example, in the genus Eucidaris, all Atlantic nominal species or subspecies, Eucidaris tribuloides, Eucidaris clavata, and Eucidaris tribuloides variegatus africana form one single genetic cluster with no distinction among them (Lessios et al., 1999). Together, this broad polytypic species is sister to the Eastern Pacific pair Eucidaris galapagensis and Eucidaris thouarsi. Based on a molecular rate estimate provided by the rise of the isthmus of Panama, the latter two sister species diverged ≈2 million years ago. An allopatric neighbor of these species, the widely distributed Pacific–Indian ocean species Eucidaris metularia is not a close relative of the eastern Pacific species. Rather, this species is an ancient offshoot in the genus, having diverged 5–8 million years ago.
The genus Tripneustes shows a pattern of dissimilar geographic distribution of genetic variation in two major oceanic regions (Lessios et al., 2003a). Tripneustes depressus from the eastern Pacific and Tripneustes gratilla from the rest of the Indo-Pacific are genetically indistinguishable, forming a single geographically widespread species complex that occupies most of the world’s tropical oceans. The Atlantic species Tripneustes
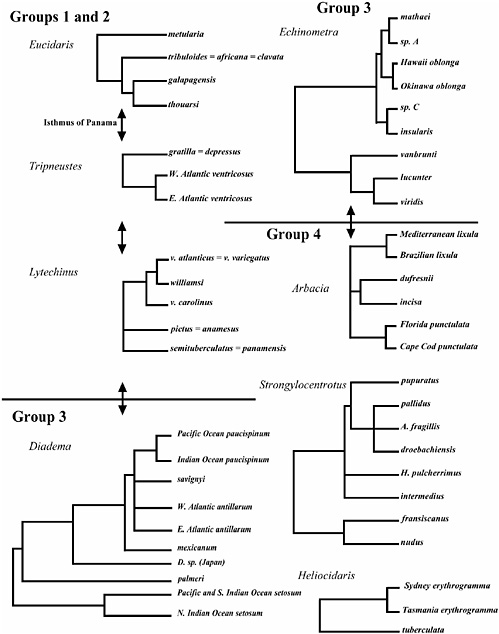
FIGURE 8.1 Molecular phylogenies based on COI (COI plus ATPase8/6 for Diadema) from eight sea urchin genera. Data are from Lessios et al. (1999, 2003a,b), Zigler and Lessios (2004), McCartney et al. (2000), Landry et al. (2003), Biermann et al. (2003), Metz et al. (1998), Zigler et al. (2003). The double arrows mark the completion of the Isthmus of Panama at ≈3.1 million years ago. This date is assumed to be approximate for the divergence of Caribbean and eastern Pacific species within the six genera marked by the arrows. Trees are rooted by species from closely related genera [see Biermann et al. (2003), Landry et al. (2003), Lessios et al. (1999, 2003a,b), McCartney et al. (2000), Metz et al. (1998), Zigler and Lessios (2004), Zigler et al. (2003) for details] and are drawn to the same temporal scale. See the text for group definitions.
ventricosus has considerable population structure, indicating a lack of gene flow between the American and African coasts. T. gratilla and T. ventricosus are assumed to have diverged at the Panamanian closure 3 million years ago.
The last genus in this cluster is more complex. Lytechinus has two sets of polytypic species: Lytechinus anamesus and Lytechinus pictus along the west coast of north America are indistinguishable genetically, as are the eastern Pacific Lytechinus semituberculatus and Lytechinus panamensis (Zigler and Lessios, 2004). These two Pacific species clusters diverged from each other 3–5 million years ago. In addition, there is a set of Atlantic species with questionable species status. Data from COI show that the subspecies L. variegatus variegatus and Lytechinus variegatus atlanticus cluster indistinguishably from one another but that L. williamsi, partially sympatric with L. variegatus variegatus, diverged at ≈500,000 years ago. An outgroup clade to this cluster is L. variegatus carolinus, which diverged 2–3 million years ago. The genealogy of bindin shows one discrepancy from that of COI: The phylogenetic positions of L. williamsi and L. variegatus carolinus are switched (Zigler and Lessios, 2004). In addition, there is evidence for acceleration of bindin evolution in the two sympatric species compared to COI, although maximum likelihood analysis fails to show positive selection, possibly because of low statistical power in these closely related sequences.
The summary of these studies of groups 1 and 2 genera is that molecular phylogenies support Mayr’s conclusions that widely distributed polytypic species are commonplace and that allopatric splitting events within ocean basins are sometimes very recent. However, some allopatric neighbors have been in existence for 2–8 million years without evidence that their ranges have begun to overlap.
Group 3
Molecular phylogenies also support Mayr’s classification of genera into group 3 because their species are in the initial stages of sympatry; but they also show that they are comprised of species groups with very different evolutionary patterns. In Diadema, the widely distributed species Diadema setosum and Diadema savignyi overlap throughout the western Pacific and Indian oceans (Lessios et al., 2003b). Based on ATPase and COI sequence differences, these two species are highly divergent, having split 7–14 million years ago (Lessios et al., 2003b). The widespread Diadema savignyi is also sympatric in Japan and the Marshal islands with an undescribed species, from which it diverged 6.5–13.5 million years ago. Isozymes and mitochondrial DNA have recently uncovered unsuspected cases of sympatry between Diadema paucispinum, a species originally
thought to be limited to Hawaii, and the other Indo-West Pacific species (Lessios and Pearse, 1996; Lessios et al., 2003b). The divergence time between the sympatric D. paucispinum and D. savignyi is <2 million years. By contrast, the eastern Pacific Diadema mexicanum has remained allopatric from the Indo-West Pacific species for 3 million years, with only a hint of range overlap with D. savignyi at the Clipperton Atoll, the closest point to the central Pacific (Lessios et al., 1996, 2003b). Within this genus, broadly distributed species tend to show divergence of supposedly conspecific allopatric populations. Diadema antillarum populations from the eastern and western Atlantic are as different from one another as are accepted species in this genus. Two clades of D. setosum in the Indian Ocean probably diverged 5 million years ago. Thus, the species in this genus show a generally higher degree of genetic divergence than genera at earlier stages, with moderately old allopatric populations within a morphospecies. Sympatry occurs between very old species pairs. Allopatric neighbors are old. One exception is the previously unsuspected sympatry of the relatively recently diverged species pair of D. savignyi and D. paucispinum.
By contrast, the genus Echinometra shows a large number of sympatric species with low divergence from one another. In the Pacific there is a cluster of very closely related sympatric species. Echinometra mathaei, Echinometra oblonga and a currently unnamed species “Echinometra sp. A” diverged 1–2 million years ago (Landry et al., 2003; Palumbi, 1996). COI data show that this cluster split from Echinometra sp. C and the Easter Island endemic E. insularis at about the same time. In the Caribbean, the sympatric species Echinometra lucunter and Echinometra viridis diverged ≈1.5 million years ago (McCartney et al., 2000). One ancient allopatric split persists in this genus: The eastern Pacific Echinometra vanbrunti differs from other Pacific species by ≈13% nucleotide differences in COI, corresponding to separation of ≈3.5 million years. However, the allopatry of E. vanbrunti from the other Pacific species may be in the process of being erased through infrequent larval influx from the central into the eastern Pacific: E. oblonga, although rare, is now present in the outer islands of the eastern Pacific (Lessios et al., 1996; McCartney et al., 2000). There is also one very recent allopatric split: E. oblonga appears to be at least two species (E. oblonga Okinawa and E. oblonga Hawaii). Distinguishable by sperm morphology and genetics, these species are allopatric and have diverged at most 250,000 to 500,000 years ago (Landry et al., 2003).
Data from the bindin locus show rapid evolution in this genus, and generally support the COI phylogeny (Landry et al., 2003; McCartney and Lessios, 2004; Metz and Palumbi, 1996). One major exception is that bindin alleles in E. sp. C differ dramatically from one region to another. In this genus, major differences in bindin gene sequence are associated with strong reproductive isolation among closely related species (Landry et al.,
2003; Metz and Palumbi, 1996; Palumbi and Metz, 1991). Where E. sp. C is in sympatry with E. oblonga (Okinawa) bindin alleles are highly divergent; in contrast, where they are allopatric, E. sp. C and E. oblonga (Hawaii) have very similar alleles (Geyer and Palumbi, 2003). In addition, bindin sequences in E. insularis are distinct and monophyletic and do not suggest a close relationship with E. sp C (Landry et al., 2003). Thus, the genus Echinometra has species with extensive sympatry, whereas the genus Diadema shows sympatry of just a few species pairs. However, contrary to predictions of Mayr’s animation, overall genetic divergence between species of Echinometra is smaller than between those of Diadema (Fig. 1). In particular, sympatric species of Echinometra show much less genetic divergence than sympatric species of Diadema.
Group 4
The genus Arbacia consists of purely allopatric species, but Mayr included it in group 4 because he doubted the validity of specific rank, even for species that were found in different oceans. He may actually have been right for the wrong reasons, because COI and bindin show that the original species designations correspond to divergent molecular clades and should not be considered as conspecific. However, neighboring allopatric clades are old and thus qualify the genus for inclusion in group 4. Based on COI sequences, Arbacia punctulata along the east coast of North America and Arbacia lixula from the eastern Atlantic and from Brazil are ≈3–5 million years old (Metz et al., 1998). The most recently derived species pair, Arbacia dufresnei and Arbacia incisa, are 2–4 million years old. Species also tend to be widespread: A. lixula occurs from the coast of Brazil to the Mediterranean, with an ≈500,000-year divergence between these genetically distinct populations. The western Atlantic species A. punctulata ranges from Cape Cod to Curacao, Trinidad and Tobago, and shows a 2% sequence divergence in COI between Florida and the northern end of its range.
Although Mayr mentioned studies of morphological variation in Strongylocentrotus droebachiensis and Strongylocentrotus pallidus, he did not place this genus in any of his groups, because it lacks tropical representatives. However, because molecular and morphological variation in Strongylocentrotus have been well studied, it can receive the same consideration as the other genera. There are many sympatric species in this genus. A cluster of species sympatric in the Northeast Pacific diverged from one another at ≈3–5 million years ago based on COI and fossil evidence (Biermann et al., 2003). Strongylocentrotus purpuratus, Strongylocentrotus droebachiensis, S. pallidus, and Allocentrotus fragilis all diverged from one another at about the same time. Although broadly overlapping
in geographic distribution, these species tend to live at different depths. A similar sympatric cluster in Japan is composed of several species that diverged slightly earlier. The two species clusters, one along North America and one along the coast of Japan, show no geographic overlap. A deep split in the genus at ≈10–15 million years ago gave rise to two allopatric sister species: Strongylocentrotus franciscanus in the east and Strongylocentrotus nudus in the west. The divergence time of this pair of species was ≈5 million years ago. Bindin evolution is rapid in this genus (Biermann, 1998).
An additional genus, also not included by Mayr because it does not occur in the West Indies, has received ample attention with regards to its molecular evolution, in part because of the remarkable divergence in mode of development between its two species. The genus Heliocidaris is restricted to Australia and consists of Heliocidaris tuberculata and Heliocidaris erythrogramma. These two species overlap in range along the southeast coast of Australia, and diverged at ≈5 million years ago (McMillan et al., 1992; Zigler et al., 2003). H. erythrogramma, which has an extremely large egg and has evolved direct development, has subpopulations in western and eastern Australia. There are no published sequence data from the western Australian subspecies, but populations from Sydney and Tasmania differ by ≈2% in COI. Bindin evolution has been rapid along the lineage leading to the direct developing H. erythrogramma.
General Tests of Predictions
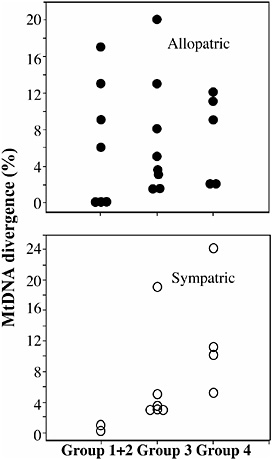
The prediction that genetic divergence of sister species increases from stage to stage of speciation is borne out by comparison of phylogenies and genetic distances among the eight genera of sea urchins (Fig. 8.2). Median COI genetic distance among 10 comparisons in groups 1 and 2 genera (the first stage of allopatric speciation) is ≈1%, whereas divergence is 3% and >10% for groups 3 and 4, respectively. Sister species in groups 1 and 2 tend to have adjoining ranges. In addition, polytypic species are most common in groups 1 and 2 genera. Five of 10 species comparisons in group 1 genera show no discernable genetic divergence, indicating genetic exchange among allopatric populations rather than species-level differentiation. By contrast, all sister-species comparisons in groups 3 and 4 genera are >2%, and there are no species with zero genetic divergence. Instead, in these genera, widely distributed species often contain allopatric populations with genetic divergences as high as that of many species pairs.
Simple predictions begin to falter, however, when comparisons are divided into those between sympatric species and those between allopatric species. Sympatric species are not generally more divergent than allo-

FIGURE 8.2 Genetic divergence among sister species in eight sea urchin genera separated into four groups, representing different stages of allopatric speciation. See the text for the definition of each group. Mitochondrial DNA divergence is based on Kimura two-parameter distances at the COI gene (COI plus ATPase8/6 for Diadema). Data are based on Fig. 8.1 and Lessios et al. (1999, 2003a,b), Zigler and Lessios (2004), McCartney et al. (2000), Landry et al. (2003), Biermann et al. (2003), Metz et al. (1998), Zigler et al. (2003). Medians are marked by arrows.
patric species. In fact, in groups 1 and 2 genera, allopatric species are more divergent than sympatric species (Fig. 8.3). This pattern is largely due to the existence of a large fraction of ancient allopatric neighbors, species that have diverged from sister taxa long ago and are separated by no insurmountable major geographic barrier, such as a land mass, even in early-stage genera. Dividing the species into allopatric and sympatric comparisons shows that divergence between sympatric species increases stage by stage but that divergence between allopatric species does not (Fig. 8.3).
Further comparisons show that most sympatric species are found in genera in which the sperm recognition protein bindin is evolving quickly
(Fig. 8.4). There is significant association between the frequency distribution of sympatric and allopatric species and the rate of their bindin evolution (Fisher’s exact test, P = 0.0033). Because the species are not phylogenetically independent, the statistical significance should not be interpreted to mean that the two quantities are related directly but rather that they tend to cooccur in the same genera. In the genera Echinometra and Strongylocentrotus, sympatric species are common, whereas in genera with slow rates of bindin evolution, sympatric species are rare. Among genera with rapid bindin evolution, 10 of 16 sister species comparisons are between sympatric species. In other genera, only two of 19 comparisons are between sympatric species (Fig. 8.4). The exception to this pattern is in the group 2 genus Lytechinus. Bindin evolution is generally slow in this ge-
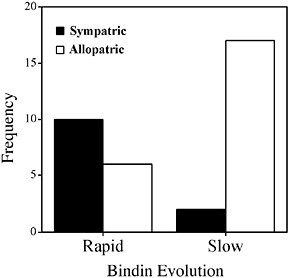
FIGURE 8.4 Frequency of sympatric and allopatric species in sea urchin genera with different rates of bindin evolution.
nus, and most species are allopatric. However, one sympatric species pair (L. williamsi and L. variegatus variegatus) can be found in the Caribbean. Divergence in bindin between these two species appears to be accelerated relative to COI divergence, although there is no statistically significant signal of positive selection.
DISCUSSION
Evolutionary series based on geography and taxonomy can be independently investigated by comparing molecular phylogenies of species groups at different stages of diversification. Phylogenies based on mitochondrial COI for eight genera of sea urchins, including 45 species, show general agreement with the evolutionary animation proposed by Mayr (1954). Groups 1 and 2 genera have lower genetic divergence and a higher incidence of polytypic species than genera at later stages. Many sister species pairs tend to be allopatric and closely related at the groups 1 and 2 stages, but, by groups 3 and 4, sister species are largely sympatric or are old allopatric neighbors.
However, ancient allopatric neighbors also occur in genera at early stages of speciation. Many of these ancient allopatric species exist across a deep water stretch of the Pacific Ocean termed the East Pacific Barrier (Ekman, 1953). Mayr (1954) writes “of the existing barriers, by far the
most potent has been that between Polynesia and the American coast.” Molecular phylogenies have revealed this statement to hold true for Eucidaris (Lessios et al., 1999), Diadema (Lessios et al., 2003b), and Echinometra (McCartney et al., 2000), although in the latter two genera there are indications that larvae are occasionally able to breach the barrier, which may represent the early stages leading toward secondary sympatry. Tripneustes in the Indo-Pacific, on the other hand, shows no evidence that its gene flow is in any way impeded by the 5,000 km of deep open water between Clipperton and the Marquesas (Lessios et al., 2003a), and a similar situation exists for Echinothrix, an Indo-Pacific genus of sea urchins that appears to have recently colonized the eastern Pacific (Lessios et al., 1996, 1998). It is unclear why some species are able to traverse the barrier so easily when the majority cannot, because there are no pronounced differences in the length of the competent larval stage of the genera.
Lessios and coworkers (Lessios and Cunningham, 1990; McCartney et al., 2000) suggest that rare immigrants into the range of an allopatric neighbor will most likely fail to reproduce or will hybridize with the resident species. Maintenance of a rare species within the range of a more numerous one demands some mechanism of reproductive isolation or assortative mating. Both mechanisms operate at the surfaces of gametes during sea urchin spawning (Palumbi, 1992). Sperm attachment and fusion is facilitated by interaction of bindin with a large protein receptor on the egg surface (Kamei and Glabe, 2003; Palumbi, 1999). Rapid evolution of bindin in sea urchins generates assortative mating (Palumbi, 1999) and egg–sperm incompatibility (Biermann, 1998; McCartney and Lessios, 2004; Metz and Palumbi, 1996). Other than the timing of gamete release (Lessios, 1984), few behavioral mechanisms of mate choice operate in these free spawning invertebrates; therefore, interactions of gametes assume a greater role in reproductive isolation. Positive selection in the bindin gene is associated with functional divergence of gamete recognition within and between species (McCartney and Lessios, 2004; Palumbi, 1999). In turn, this divergence may allow sympatry of species at an earlier stage of divergence. Genera with rapid bindin evolution include many closely related sympatric species. By contrast, in genera without rapid change in bindin, closely related or moderately related species are nearly exclusively allopatric (Fig. 8.4).
This difference among genera enhances our understanding of species formation and helps us interpret Mayr’s animation in more mechanistic ways. In Mayr’s original formulation, groups 1 and 2 genera eventually evolve into groups 3 and 4 genera. The major difference between these categories is the amount of time that species have had to diverge genetically: Reproductive isolation was thought to evolve as a consequence of
overall genetic divergence or adaptive differentiation (Mayr, 1942, 1963). The critical importance of a few gamete recognition loci to reproductive isolation can disrupt the steady pace of evolution of isolation (Dobzhansky, 1937, 1970; Orr, 1991; Wyckoff et al., 2001). In these cases, the way a few loci evolve may be more important than overall genetic divergence or adaptive differentiation (Dobzhansky, 1937, 1970).
Our phylogenetic summary shows that the distinction between sea urchin genera at different speciation stages is related to bindin evolutionary rate. Groups 1 and 2 genera have slow bindin evolution, have largely allopatric species, and are therefore earlier frames in the evolutionary animation. Groups 3 and 4 genera, with many more sympatric species, are classified later in the evolutionary series, but their species are not necessarily older. Instead, these can be the genera in which rapid bindin change generates reproductive isolation among even closely related species.
A similar hierarchy of speciation rate might be present in Mayr’s early work on tropical Pacific birds. Genera with bright male plumage and strong sexual selection were textbook cases of geographic variation within sister-species complexes (Mayr, 1942). Such genera might be akin to the sea urchin genera with fast bindin evolutionary rates. A key difference, however, is that evolution of plumage generates morphological diversity across a species that allows it to be divided into taxonomic units based on morphology (Mayr, 1942). By contrast, rapid evolution of bindin does not in and of itself generate strong morphological variation, and in genera with rapid bindin evolution, species designations had to await the ability of molecular tools to assay genetic differences. The most diverse set of closely related sea urchin species known, the five or six Indo-West Pacific species of Echinometra, had been classified as a single, large polymorphic species by Mortensen (1928–1951). Only after reproductive barriers and genetic differences became clear were subtle morphological and ecological distinctions discovered (Matsuoka and Hatanaka, 1991; Palumbi and Metz, 1991; Uehara and Shingaki, 1985).
Another potential difference between bindin and plumage evolution is the driver producing different evolutionary rates. Sexual selection is thought to drive the divergence of male coloration in birds through a runaway process based on female preference (Kirkpatrick, 1982; Lande, 1981). Because female preference and male traits coevolve differently in separate isolated areas, different populations can attain novel trait and preference combinations (Ryan and Wilczynski, 1988). For bindin and its receptor genes, an excess of amino acid replacement substitutions is a signal that evolution is driven by selection, but the source of this selection remains unclear. It is possible that selection is for increasingly better fertilization systems, with the bindin genes evolving to produce more optimal fertilization phenotypes. Positive selection in this case would be
driven by a change in fertilization environment, such as water motion or distance from conspecifics, and would not be an ongoing process of male–female coevolution.
Alternatively, several coevolutionary scenarios have been proposed for rapid evolution of gamete recognition. Selection for divergent bindin alleles in sympatric species by a process of reinforcement has been observed in the most recently diverged Echinometra. The Okinawa and Hawaiian populations of E. oblonga diverged 250,000–500,000 years ago, yet the Okinawa population is highly distinct in bindin sequence and sperm morphology (Landry et al., 2003). Rapid evolution in the Okinawa population is related to the presence of sympatric populations of E. sp. C (Geyer and Palumbi, 2003). and is probably responsible for high levels of conspecific sperm precedence and low hybridization (Geyer and Palumbi, 2005). McCartney and Lessios (2004) suggested that rapid evolution of bindin in Caribbean Echinometra was due to divergence of the fertilization systems of two sympatric species. In this case, rapid bindin evolution in one species was associated with evolution of egg specificity in the same species (Lessios and Cunningham, 1990; McCartney and Lessios, 2002). A third case of rapid bindin evolution associated with gamete morphological change has been reported by Zigler et al. (2003). Heliocidaris erythrogramma has a giant egg and has evolved strong developmental differences from typical sea urchins with planktotrphic larvae, including its congener H. tuberculata. Bindin has evolved rapidly along the branch leading to the developmentally novel species but not along the branch leading to H. tuberculata.
Yet reinforcement or rapid gamete differentiation between species are not the sole evolutionary pressures acting on gamete recognition. Sea urchin species with bindin that is rapidly diverging between species are typically also highly polymorphic intraspecifically, and males with different bindin alleles produce functionally different sperm (Palumbi, 1999). Recent experiments with sperm mixtures also show that heterozygote males are superior fertilizers but only for certain allele combinations (S.R.P., unpublished data). Thus, the maintenance of polymorphism within species is probably due, at least in part, to balancing selection among alleles.
Another plausible mechanism of within- and between-species evolution of bindin is a coevolutionary antagonism between males and females. Males are selected to produce sperm that fertilize quickly and indiscriminately, whereas females are selected to control which single sperm is to be used for fertilizing a single egg (Palumbi, 1998; Rice, 2000; Swanson and Vacquier, 2002a,b). If so, then we would expect the egg receptor gene to be highly polymorphic and evolution toward more and more choosy eggs to be an ongoing process. To date there are no data on the egg receptor to test these predictions.
Why do some sea urchin genera have higher rates of bindin evolution? Bindin evolves by a combination of amino acid substitution and insertion or deletion of repeated amino acid motifs (Zigler and Lessios, 2003b). Zigler and Lessios (2003b) compared the primary sequence of the bindin protein between genera showing different evolutionary rates in this molecule. They found no obvious differences among genera with fast versus slow rates. Levitan (2002) argued that fertilization rates are determined by population densities of conspecific sea urchins, but there is no clear relationship between ecological density and the rate of bindin evolution among the genera.
Overall, the clear relationship between bindin evolution and young, sympatric species remains a strong signal that gamete recognition and species formation are tightly linked. But the underlying mechanisms driving this pattern remain poorly explained. Understanding the coevolution of bindin and the recently described sperm-receptor gene (Kamei and Glabe, 2003) may help make these mechanisms clearer.
CONCLUSIONS
Ernst Mayr’s evolutionary animation is supported by recent molecular phylogenies of eight sea urchin genera at different stages of diversification. The details of these phylogenies reveal a tight association between sister-species status and geography at early stages. Molecular phylogenies also provide strong evidence for departures from Mayr’s strict series. In particular, genera in which gamete recognition loci are more quickly evolving tend to fall at later stages in Mayr’s evolutionary series. These genera are not necessarily comprised of older species. Instead, they can be composed of young species that have more rapidly evolved reproductive isolation and assortative mating. This result indicates that the assumption that reproductive isolation accumulates gradually with time does not hold when such isolation arises from changes in a single locus, instead of being the product of small changes in many loci. Nevertheless, the end result is basically identical to Mayr’s assertion that sympatric species tend to be found in genera with greater reproductive isolation. By placing genera at different stages of diversification in a temporal series, Mayr animated the process of speciation and made its mechanisms clearer. The same basic approach remains valuable today and is all the more accurate when phylogenetic data permit the determination of the order in which species were separated from each other.
REFERENCES
Barraclough, T. G. & Vogler, A. P. (2000) Detecting the geographical pattern of speciation from species-level phylogenies. Am. Nat. 155, 419–434.
Biermann, C. (1998) The molecular evolution of sperm bindin in six species of sea urchins (Echinoida: Strongylocentrotidae). Mol. Biol. Evol. 15, 1761–1771.
Biermann, C. H., Kessing, K. & Palumbi., S. R. (2003) Phylogeny and development of marine model species: Strongylocentrotid sea urchins. Evol. Dev. 5, 360–371.
Coates, A. G. & Obando, J. A. (1996) Geological evolution of the Central American isthmus. In Evolution and Environment in Tropical America, eds. Jackson, J. B. C., Coates, A. G. & Budd, A. (Univ. of Chicago Press, Chicago), pp. 21–56.
Dobzhansky, T. (1937) Genetics and the Origin of Species (Columbia Univ. Press, New York).
Dobzhansky, T. (1970) Genetics of the Evolutionary Process (Columbia Univ. Press, New York).
Ekman, S. (1953) Zoogeography of the Sea (Sidgwick & Jackson, London).
Geyer, L. B. & Palumbi, S. R. (2003) Reproductive character displacement and the genetics of gamete recognition in tropical sea urchins. Evolution 57, 1049–1060.
Geyer, L. B. & Palumbi, S. R. (2005) Conspecific sperm precedence in two species of tropical sea urchins. Evolution 59, 97–105.
Hillis, D. M., Moritz, C. & Mable, B. K. (1996) Molecular Systematics (Sinauer, Sunderland, MA), 2nd Ed.
Kamei, N. & Glabe, C. G. (2003) The species-specific egg receptor for sea urchin sperm adhesion is EBR1, a novel ADAMTS protein. Genes Dev. 17, 2505–2507.
Kirkpatrick, M. (1982) Sexual selection and the evolution of female choice. Evolution 36, 1–12.
Lande, R. (1981) Models of speciation by sexual selection on polygenic traits. Proc. Natl. Acad. Sci. USA 78, 3721–3725.
Landry, C., Geyer, L. B., Arakaki, Y., Uehara, T. & Palumbi, S. R. (2003) Recent speciation in the Indo-West Pacific: rapid evolution of gamete recognition and sperm morphology in cryptic species of sea urchin. Proc. R. Soc. London Ser. B 270, 1839–1847.
Lessios, H. A. (1984) Possible prezygotic reproductive isolation in sea urchins separated by the Isthmus of Panama. Evolution 38, 1144–1148.
Lessios, H. & Cunningham, C. W. (1990) Gametic incompatibility between species of the sea urchin Echinometra on the two sides of the Isthmus of Panama. Evolution 44, 933–941.
Lessios, H. A. & Pearse, J. S. (1996) Hybridization and introgression between Indo-Pacific species of Diadema. Mar. Biol. (Berlin) 126, 715–723.
Lessios, H. A., Kessing, B. D., Wellington, G. M. & Graybeal, A. (1996) Coral Reefs 15, 133–142.
Lessios, H. A., Kessing, B. D. & Robertson, D. R. (1998) Massive gene flow across the world’s most potent marine biogeographic barrier. Proc. R. Soc. London Ser. B 265, 583–588.
Lessios, H. A., Kessing, B., Robertson, D. R. & Paulay, G. (1999) Population structure and speciation in tropical seas: Global phylogeography of the sea urchin Diadema. Evolution 53, 806–817.
Lessios, H. A., Kane, J. & Robertson, D. R. (2003a) Phylogeography of the pantropical sea urchin Tripneustes: Contrasting patterns of population structure between oceans. Evolution 57, 2026–2036.
Lessios, H. A., Kessing, B. & Pearse, J. (2003b) Population structure and speciation in tropical seas: Global phylogeography of the sea urchin Diadema. Evolution 55, 955–975.
Levitan, D. R. (2002) The relationship between conspecific fertilization success and reproductive isolation among three congeneric sea urchins. Evolution 56, 1599–1609.
Matsuoka, N. & Hatanaka, T. (1991) Molecular evidence for the existence of four sibling species within the sea-urchin, Echinometra mathaei in Japanese waters and their evolutionary relationships. Zool. Sci. 8, 121–133.
Mayr, E. (1942) Systematics and the Origin of Species (Columbia Univ. Press, New York).
Mayr, E. (1954) Geographic speciation in tropical echinoids. Evolution 8, 1–18.
Mayr, E. (1963) Animal Species and Evolution (Harvard Univ. Press, Cambridge, U.K.).
McCartney, M. A. & Lessios, H. A. (2002) Quantitative analysis of gametic incompatibility between closely related species of neotropical sea urchins. Biol. Bull. (Woods Hole, Mass.) 202, 166–181.
McCartney, M. A. & Lessios, H. A. (2004) Adaptive evolution of sperm bindin tracks egg incompatibility in neotropical sea urchins of the genus Echinometra. Mol. Biol. Evol. 21, 732–745.
McCartney, M. A., Keller, G. & Lessios, H. A. (2000) Dispersal barriers in tropical oceans and speciation in Atlantic and Pacific sea urchins of the genus Echinometra . Mol. Ecol. 9, 1391–1400.
McMillan, W. O., Raff, R. A. & Palumbi, S. R. (1992) Population genetic consequences of developmental evolution and reduced dispersal in sea urchins (genus Heliocidaris). Evolution 46, 1299–1312.
Metz, E. C. & Palumbi, S. R. (1996) Positive selection and sequence rearrangements generate extensive polymorphism in the gamete recognition protein bindin. Mol. Biol. Evol. 13, 397–406.
Metz, E. C., Gomez, G. G. & Vacquier, V. D. (1998) Mitochondrial DNA and bindin gene sequence evolution among allopatric species of the sea urchin genus Arbacia. Mol. Biol. Evol. 15, 185–195.
Mortensen, T. (1928–1951) A Monograph of the Echinoidea (C. A. Reitzel, Copenhagen).
Orr, H. A. (1991) Is single-gene speciation possible? Evolution 45, 764–769.
Palumbi, S. R. (1992) Marine speciation on a small planet. Trends Ecol. Evol. 7, 114–118.
Palumbi, S. R. (1996) What can molecular genetics contribute to marine biogeography? An urchin’s tale. J. Exp. Mar. Biol. Ecol. 203, 75–92.
Palumbi, S. R. (1998) Species formation and the evolution of gamete recognition loci. In Endless Forms: Species and Speciation, eds. Howard, D. & Berlocher, S. (Oxford Univ. Press, New York), pp. 271–278.
Palumbi, S. R. (1999) All males are not created equal: Fertility differences depend on gamete recognition polymorphisms in sea urchins. Proc. Nat. Acad. Sci. USA 96, 12632–12637.
Palumbi, S. R. & Metz, E. (1991) Strong reproductive isolation between closely related tropical sea urchins (genus Echinometra). Mol. Biol. Evol. 8, 227–239.
Rice, W. R. (2000) Dangerous liaisons. Proc. Natl. Acad. Sci. USA 97, 12953–12955.
Ryan, M. J. & Wilczynski, W. (1988) Coevolution of sender and reciever: Effect on local mate preference in cricket frogs. Science 240, 1786–1789.
Swanson, W. J. and Vacquier, V. D. (2002a) Rapid evolution of reproductive proteins. Nat. Rev. Genet. 3, 137–144.
Swanson, W. J. and Vacquier, V. D. (2002b) Reproductive protein evolution. Ann. Rev. Ecol. Syst. 33, 161–179.
Uehara, T. & Shingaki, M. (1985) Taxonomic studies in the four types of the sea urchin, Echinometra mathaei, from Okinawa, Japan. Zool. Sci. 2, 1009.
Wyckoff, G. J., Wang, W. & Wu, C.-I. (2001) Rapid evolution of male reproductive genes in the descent of man. Nature 403, 304–309.
Zigler, K. S. & Lessios, H. A. (2003a) Evolution of bindin in the pantropical sea urchin Tripneustes: Comparisons to bindin of other genera. Mol. Biol. Evol. 20, 220–231.
Zigler, K. S. & Lessios, H. A. (2003b) 250 million years of bindin evolution. Biol. Bull. (Woods Hole, Mass.) 205, 8–15.
Zigler, K. S. & Lessios, H. A. (2004) Speciation in the coasts of the New World: Phylogenography and evolution of the bindin in the sea urchin gneus Lytechinus. Evolution 58, 1225–1241.
Zigler, K. S., Raff, E. C., Popodi, E., Raff, R. A. & Lessios, H. A. (2003) Adaptive evolution of bindin in the genus Heliocidaris is correlated with the shift to direct development. Evolution 57, 2293–2302.


















