
Promising Approaches to the Development of Immunomodulation for the Treatment of Infectious Diseases: Report of a Workshop
Committee on New Directions in the Study of Antimicrobial Therapeutics: Immunomodulation
INTRODUCTION
The human immune system is equipped to fight a wide array of potential pathogens. Infection leads to disease only when the interaction between the host and the microorganism results in damage sufficient to disrupt homeostasis. For the last 70 years, the effort to prevent or treat infectious diseases has relied heavily on targeting microorganisms themselves with antimicrobial drugs. That approach has been extraordinarily successful, especially against bacterial infections. As described in the previous report, it has also resulted in the generation of multiple-drug-resistant (MDR) microorganisms that are threatening to become untreatable. The success and low cost of broad-spectrum antibiotics have reduced the incentive to develop alternative antimicrobial strategies, such as augmenting host responses during infectious disease.
The National Institute of Allergy and Infectious Diseases asked the National Research Council to convene a committee of experts, the Committee on New Directions in the Study of Antimicrobial Therapeutics: Immunomodulation, to organize a brainstorming workshop to explore novel ways of modulating the host immune system to treat infectious disease (biographical sketches of the committee members are found in Appendix D). Therapeutic strategies based on modulating the immune response have several potential advantages over the use of traditional antimicrobials. First, because immunomodulators do not act on microorganisms directly,
they may circumvent the problem of rapid emergence of resistance. Second, immunomodulators may expand treatment options for immunocompromised patients, in whom traditional antimicrobials often work poorly. Third, they offer the potential of a broad spectrum of activity against viral and fungal, as well as bacterial, diseases and may provide nonspecific emergency-treatment options in the event of the emergence of a novel pathogen or a biowarfare attack.
The 2-day workshop, held on April 29-30, 2005, was attended by 33 invited participants, who had a wide array of expertise in molecular and cellular biology, chemistry, ecology, microbiology, immunology, and infectious disease. An agenda and a roster of participants and speakers can be found in Appendix E. Six speakers provided the participants with an overview of immune-system function, potential mechanisms of immunomodulation, and possible obstacles to the development of new immunomodulators. At the end of the first day, the participants formed four discussion groups. Each group identified a number of potential therapeutics and defined background research that would be necessary to make their development possible. The workshop concluded with a plenary discussion during which the breakout groups’ conclusions were presented and discussed. This report describes the most interesting insights that came out of the workshop and the most promising avenues for future research on and development of immunomodulators to treat infectious diseases.
An Informed Choice of Goals
The objective of developing a single immunomodulatory agent that is effective against all infectious agents in all patients was generally considered to be unrealistic or even foolhardy. A clear theme emerging from the workshop was that immunomodulatory interventions will probably be most effective in a tailored role: against particular agents, in subsets of patients, at critical points in the course of an infectious disease, or most promisingly as adjuncts to therapeutics—such as antibiotics and antivirals—that target the microorganism directly. Because so many of the potential immunomodulatory therapies discussed at the workshop would have narrow applications, an overarching conclusion of the discussions was the importance of developing rapid and specific diagnostics both to identify the disease-causing agent and to define the patient’s immune status and stage of infection. Improved ability to identify the patients most likely to require or respond to particular immunomodulators will greatly increase the likelihood of suc-
cess in clinical trials, another area identified during the workshop as a critical obstacle to the development of immunomodulators.
An infectious disease can occur only in a susceptible host, with susceptibility being a function of the effectiveness of the immune response. Damage to the host may be caused directly by microbial factors, can result from host factors—such as inflammatory responses—or both (Casadevall and Pirofski 2003). Some microbial diseases cause damage to the host because an overly vigorous immune response leads to excessive inflammation; others occur because immune responses are insufficient and require bolstering (Janeway et al. 2001). Host damage as a function of the immune response can be represented by a parabolic damage-response curve (Figure 1). The goal of therapeutic immunomodulation is to reduce host damage by shifting the damage-response curve to a point that is more beneficial to the host. This represents a shift from the more conventional therapeutic goal of killing the infectious organism.
Therefore, a fully stocked immunomodulation toolbox will require the development of both agents that stimulate and agents that suppress the immune response. Furthermore, optimal use of any immunomodulator will
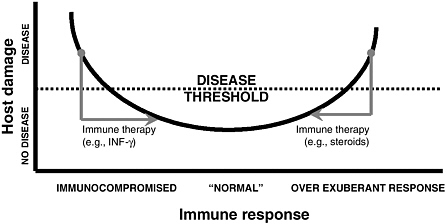
FIGURE 1 Host damage is a function of the quantitative and qualitative aspects of the host response to a microorganism. Damage can originate in microbial effects on host tissues, the immune response, or both. For most microorganism-host interactions, clinically apparent damage (disease) occurs at the extremes of the immune response. Conceptually, immunomodulator-based therapies can be considered interventions that attempt to reduce host damage by either enhancing or suppressing the immune response. For a more detailed discussion of the “damage-response framework,” see Casadevall and Pirofski (2003).
require new tools to measure host damage because the type of intervention needed could depend on where the host is on the curve of damage vs. immune response. The availability of a variety of agents with the ability to fine-tune the immune response would provide clinicians with many new options in the treatment of infectious diseases. The development of more sensitive tools to determine immune status and host response will allow greater predictability of the effects of immunomodulators on different groups of patients, such as the immunocompromised.
Mining the Immune System for New Therapies
The major advantage of developing interventions for infectious diseases based on immune modulation is that it recruits and engages a system that has evolved to protect against microorganism-related diseases. Experience with vaccination and with passive antibody and cytokine therapy has shown that immune modulation can be effective and safe. However, one difficulty in developing new strategies to augment the host immune system to prevent or treat infectious diseases lies in the nature of the immune system itself. It is a complex network of intricately related, overlapping subsystems that function together to maintain immune homeostasis. It must recognize a full array of microorganisms and react to those that cause damage in such a way that they are eliminated without undue damage to the host. In general, this network responds to microbial agents in a rapid, efficient, and self-limiting manner. Appropriate stimulation of immune pathways is crucial to successful resolution of infection. Turning the immune response off after an infection is resolved is equally important, in that overstimulation of the immune system can lead to uncontrolled inflammation.
A Web, Not a Firetruck
Current understanding of the immune system divides it into two systems: innate immunity, which reacts quickly and nonspecifically to any pathogen; and acquired immunity, which generates a specific response and remembers interactions with microbial agents. Increasingly, research suggests that this simple model is inadequate. For example, vaccines and passive antibody agents that target a specific microbial agent can stimulate innate immune mechanisms (Binder et al. 2005), and the interaction of particular microbial components with receptors of the innate immune sys-
tem can exhibit some specificity (Choe et al. 2005; O’Neill 2005). Coordination of the innate and acquired responses requires constant communication among a series of interlocking feedback loops, further blurring the distinction between the two systems (Dower and Qwarnstrom 2003; Smith and Bolouri 2005). Exact prediction of how a particular immunomodulator will affect this complex network is not yet feasible. Furthermore, it is increasingly clear that factors in addition to the innate and acquired immune systems affect a person’s total immune response. Some of those factors are shown in Figure 2.
We Are Not Alone
Biologists estimate that humans form integrated relationships with thousands of microbial species, collectively known as the resident microbiota (Wilson 2005; Nataro et al. 2005). These dynamic alliances begin to form immediately at birth, and undergo an ecological succession over the first few years of life that leads to the establishment of a mature set of communities at the age of about 4 years. The assemblages occur on the skin and along the mucosal surfaces of the body. They make a critical contribution to human health by providing nutrients and mediating normal development and function of host tissues, including those of the immune system (Noverr and Huffnagle 2004). The resident microbiota form the first line of defense against invasion of the human host by new pathogenic microorganisms (Figure 2, panel A). There is considerable evidence that the dynamics of the resident microbial communities are managed by their interactions with the host immune system and that a breakdown in the normal communication between the microbiota and the immune system can promote microbial disease (Kelly et al. 2005). However, the precise mechanisms that tune the interactions are poorly understood, and how the host distinguishes microbial friend from foe remains largely enigmatic.
Every Case Is Different
Developing immunomodulators is complicated by the fact that people have varied susceptibility and responses to microbial agents (Figure 2, panel E), and that the immune response changes during the course of a host-microorganism interaction. Infection with agents like HIV or immuno-suppressive treatments can result in varying degrees of immunocompromise. A person’s genetic background also affects his or her response
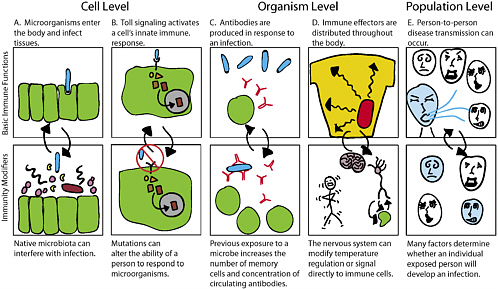
to any given agent; for example, some alleles of the CCR5 receptor confer resistance to infection with HIV (de Silva and Stumpf 2004), whereas variation along a particular innate immune pathway could dampen or stimulate an individual’s reaction to particular microorganisms (Figure 2, panel B). Furthermore, each person has a unique history of exposure to microbial agents and thus has varied susceptibility to newly encountered microorganisms (Figure 2, panel C). The immune response is constantly fine-tuned during the course of a host-microorganism interaction, rather than being fixed or static, so the same immunomodulator could have different activities at different times during this response. Finally, response to a microorganism depends on communication between the immune system and other systems that contribute to homeostasis, for example, the nervous system (Tracey et al. 2001) (Figure 2, panel D); thus, therapeutics that work through the nervous system could have immunomodulatory effects and be effective in treating infectious diseases.
Increasing awareness of the complexity of the human immune response may make the process of developing and testing immunomodulatory therapeutics daunting. However, basic research continues to reveal potential immunomodulatory compounds and intervention points. Furthermore, highly successful immunomodulatory strategies, such as vaccination and passive antibody therapy, have been developed and deployed without full understanding of their modes of action. Therefore, the committee, after considering the many ideas discussed during the workshop, recommends a number of approaches as most likely to result in the development of safe and effective immunomodulators for use in the treatment of infectious disease. The committee also included recommendations for research that could bring greater scientific rigor to innovation in already established immunomodulatory therapeutics like immunization. The committee organized its recommendations into four parts: the first two discuss the most promising interventions based on modulating innate and acquired immunity, the third presents ways in which better understanding of the resident microbiota could lead to novel immunomodulators and the fourth discusses cross-cutting research needed to advance the field of immunomodulation in general.
MODULATING INNATE IMMUNITY
Innate immune responses are highly conserved across evolution (Abreu and Arditi 2004; Beutler 2004). Until recently, the innate immune system
was viewed as rather primitive, ringing the same alarm bell for every attacker. Recent research, however, has revealed that the innate immune system encompasses a complex set of receptors, signaling peptides, and cytokines linked in a web of interlocking feedback loops (O’Neill 2005). Innate immunity shares with conventional antimicrobial therapy a rapid onset of action (within minutes to hours) and a relatively low specificity: once activated, it can act on diverse microorganisms. Modulators of innate immunity may be less likely than conventional antimicrobial therapy to elicit resistance because immune modulators do not disable a specific microbial target and their mechanisms of action involve multiple effector cells and mediators.
Some microorganisms damage the host through virulence factors that subvert or mute the innate immune response (Jarva et al. 2003; Fournier and Philpott 2005). In other cases, an excessive immune response or one that fails to turn off after the threat has passed is responsible for host damage (Casadevall and Pirofski 2003; Polderman and Girbes 2004). Therefore, therapeutics based on modulating innate immunity will require both positive and negative modulators and the ability to distinguish which will be beneficial at a particular point in a given infection. Many modulators of innate immunity may prove unable to cure disease on their own but will be of particular use as adjuncts to conventional antibiotic therapies. The timing of intervention is likely to be critical because an intervention that protects if given before infection may increase the likelihood of host damage if administered after infection has occurred. Indeed, it is possible that both positive and negative immunomodulators could be useful at different stages of the same disease. The committee considered separately the most promising approaches to boosting and suppressing innate immunity.
Boosting Innate Immunity
Research in the last 10 years has revealed that the human innate immune system includes a large array of pattern-recognition receptors (PRRs), including scavenger receptors, complement receptor 3, mannose receptor, Nucleotide-binding and Oligomerization Domain-containing (NOD) proteins, and at least 10 Toll-like receptors (TLRs; originally discovered in Drosophila) that react to different classes of microbial signals. Each PRR then stimulates overlapping but not identical signal-transduction pathways leading to the production of a cascade of cytokines. Many of the same cytokine pathways are triggered by different PRRs, but the pattern of stimu-
lation initiated by each PRR is somewhat tailored to the class of microorganism it detects. The discovery of PRRs has stimulated research into what triggers them (agonists), the resulting signaling cascade, and the complex interactions between the events set off by PRR activation and the rest of the immune system. The list of potential molecular targets for modulators of innate immunity is extensive (Beutler 2004; Germain 2004).
The Cytoscape image shown in Figure 3 includes some of the known genes that interconnect TLR4 (which detects lipopolysaccharides (LPS), a component of bacterial outer membranes) with genes that are regulated by the NFκB transcription factor. Clearly there are numerous potential intervention points even in this small part of the innate immune system, and hints that the process is highly orchestrated (regulated) are starting to be obtained.
The committee identified three potential boosters of innate immunity as having particular promise: TLR agonists and agents that modulate the TLR response pathway, cationic host-defense peptides, and direct expansion of the effector cells normally activated by the innate immune system.
TLR agonists have already shown promise as immunomodulators. For example, species-specific CpG oligonucleotides (representing a signature bacterial DNA sequence), acting through TLR9, can enhance host resistance to bacterial and viral infections without overtly causing adverse effects through overproduction of proinflammatory cytokines (Klinman 2004). CpGs are effective in experimental infection models against a variety of pathogens—including Francisella tularensis, Listeria monocytogenes, and Cryptococcus neoformans—and in models of polymicrobial intra-abdominal sepsis (Ito et al. 2005; Rice et al. 2005; Krieg 2002). Other modulators acting through PRRs include the peptidoglycan subunit muramyl dipeptide, LPS-derived monophosphyl lipid A, fungal cell wall β-glucans, and various synthetic agonists of TLRs (O’Neill 2003).
Alternatively, cationic-host defense (antimicrobial) peptides can modulate innate immunity and protect against infection without inducing inflammation or even while suppressing it (Bowdish et al. 2005; Finlay and Hancock 2004). Peptides in this diverse family, produced by phagocytes and epithelial cells and present in mucus and other fluid interfaces between the host and the environment, can act against microorganisms directly when they are present in high concentrations (for example, in the granules of phagocytes); in lower concentrations (for example, at mucosal surfaces), they fulfill a still largely uncharacterized regulatory role in the innate im-
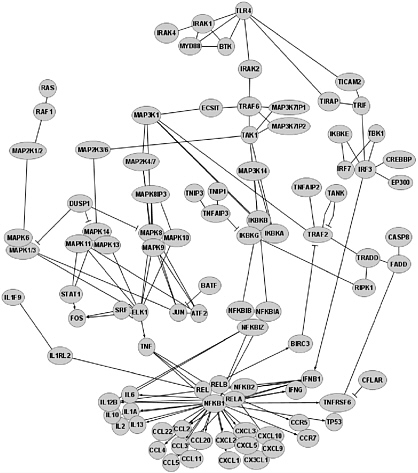
FIGURE 3 View of the TLR4 pathway with the Cytoscape program. Each circle represents a particular protein, and the lines connecting them reflect known interactions (signal transduction, gene regulation, or physical protein-protein interaction). The net effect of engagement of the TLR4 receptor by LPS is migration of transcription factor NFκB (indicated by its five known subunits which operate in pairs, NFκB1 (p50), RelA, RelB, C-Rel, and NFκB2) into the nucleus, which leads to upregulation of transcription of many genes (signified by arrows). Negative regulatory events are indicated by lines with perpendicular intersections (⊥). We believe that this network, although complicated, is much simpler than the actual case. Constructed and provided with permission by Fiona Roche, Simon Fraser University.
mune system. Derivatives without direct antimicrobial activity can also protect against infections (Leist and Jaattela 2002).
A downstream result of activation of innate immune sentinels, such as TLRs, is the expansion of various effector cells of both the acquired and innate immune systems. Therefore, another potential means of modulating innate immunity is to develop therapeutics that would increase the appropriate cell populations either indirectly by use of appropriate colony-stimulating factors (such as G-CSF and GM-CSF) or by improving technologies for white blood cell transfusions, boosting the lifespan of normal cells (for example, by controlling apoptosis), using ex vivo educated autologous effector cells (host effector cells that have been removed from the host and modified), or using synthetic effector cells (effector cells from a non-host source).
Compounds that modulate innate immunity could have unpredictable or even conflicting effects on the host response, depending on whether they counteract or augment the natural response induced by the host-microorganism interaction. The numerous interlocking regulatory mechanisms, which ensure that the immune response is sufficient to clear the disease-causing microorganism and is then promptly turned off, are not well enough understood to support prediction of the activity of innate immune modulators. Experience indicates that attempts to modulate innate immunity can have unwelcome results. Agonists of TLRs can trigger autoimmune diseases in mice that are genetically predisposed (Lang et al. 2005). Repetitive administration of CpG-containing oligonucleotide agonists of TLR9 can lead to massive lymphocyte depletion in mice (Heikenwalder et al. 2004). Administration of self-proteins such as antimicrobial proteins and CSFs, especially recombinant versions, occasionally elicits an antibody response (Reumaux et al. 2004). Therefore, the development of safe and effective innate immune modulators will require substantial basic research.
Research characterizing the intricacies of the innate immune system will facilitate the harnessing of these potential therapeutic approaches. In particular, the committee recommends basic research in the following areas to support the development of therapeutics to boost the innate immune system:
-
I-1.1 Characterization of PRR agonists, their downstream signaling pathways, and effector mechanisms.
-
I-1.2 Characterization of elements that regulate the balance between proinflammatory and anti-inflammatory pathways in innate immunity.
-
I-1.3 Characterization of the selective immunomodulatory effects of cationic host-defense peptides.
-
I-1.4 Identification of biomarkers of innate immune status.
Suppressing Inflammation
Excessive stimulation of innate immunity by some host-microorganism interactions can produce proinflammatory mediators, such as TNF-α, that promote host damage. Sepsis-like syndromes characterized by hemodynamic instability and metabolic abnormalities affect more than 750,000 people in North America annually and cause some 215,000 deaths (Angus et al. 2001). Therapies designed to treat sepsis by down-modulating immune responses have been largely unsuccessful (Opal 2003). The failures may be due, at least in part, to the heterogeneity of the syndrome and the late stage at which treatment is usually initiated. In addition, many of the candidate compounds that have been tested act specifically on a single step in the complex pathways that regulate the inflammatory process. Given the complex dynamics of the inflammatory response, the window of opportunity for such specific immunomodulators is likely to be narrow.
Many of the antisepsis agents that have had disappointing clinical results target individual mediators of inflammation. Agents acting at critical checkpoints may be more effective. Examples of promising targets that appear to amplify multiple inflammatory mediators are High Mobility Group Box-1 (HMGB1), Triggering Receptor Expressed on Myeloid Cells-1 (TREM-1), Macrophage Migration Inhibitory Factor (MIF) (Andersson and Tracey 2003; Bouchon et al. 2001; Bucala 1994) and TLR4 antagonists like eritoran and TAK-242 (Rossignol and Lynn 2005). Strategies to limit inflammation associated with infection include enhancing T-regulatory cell activity in vivo. These cells suppress a variety of inflammatory tissue disorders—such as inflammatory bowel disease, fibrosis, and asthma—and it is conceivable that harnessing their activity in a controlled manner during infection will be beneficial (Belkaid and Rouse 2005).
Control of neutrophil activation and trafficking may also be useful in this regard. Neutrophils are critically important in the innate immune response to infection, but neutrophil persistence at the site of infection can
lead to unmitigated local and systemic tissue damage. Dysregulated apoptosis, cytokine signaling, and delayed mononuclear cell infiltration to clear polymorphonuclear leukocytes contribute to the inflammatory process (McLoughlin et al. 2003); studies of this phenomenon are recommended with the goal of identifying therapeutics that target T-regulatory cell and neutrophil activation and trafficking. Alternative approaches would depend on a more profound understanding of the regulation of proinflammatory cytokines. For example, it is known that the human host-defense peptide LL-37 is one of the most effective antiendotoxins in mouse sepsis models (Klinman 2004) because of its ability to suppress the induction of the proinflammatory cytokine TNF-α by bacterial LPS; nevertheless, despite suppressing this component of innate immune responses, LL-37 can also protect against infections (Leist and Jaattela 2002). This led the committee to suggest that a better understanding of innate immunity will yield other approaches to suppressing sepsis selectively.
The disappointing results of interventions that target a single step in the inflammatory pathway may have been caused by the extremely narrow window of efficacy of such agents. The agents may be much more effective if given to the right patient at the right time. Therefore, the committee recommends prospective studies that allow for stratification of sepsis patients into distinct categories (Polderman and Girbes 2004). Advances in genomics and proteomics could provide opportunities for such stratification. In particular, gene-array technologies could lead to the discovery of novel biomarkers that can rapidly identify patients at the earliest stages of sepsis and group sepsis patients on the basis of their particular response patterns. The rapid advancement of proteomics technology and its application to sepsis could yield similar rewards. Prospective clinical studies of hospital inpatients would facilitate the identification of candidate biomarkers for assay development. Such studies should include collection and analysis of samples (for example, of blood and urine) and archiving of sample components such as serum and white cells with sophisticated database support. The development of a high-content database would allow correlation of the results of sample analysis with clinical course to identify candidate biomarkers. The archived samples could then be used for testing and validation. Eventually, the biomarkers would be used to stratify patients and guide treatment.
The committee therefore recommends two promising approaches to the development of agents to suppress innate immunity:
-
I-2.1 Development of therapeutics that can modulate regulation of the inflammatory network, especially those that harness T-regulatory cell function, the TLR signaling pathways, and neutrophil activity and trafficking.
-
I-2.2 Prospective studies with gene array and proteomic technology to allow the stratification of sepsis patients into distinct categories.
MODULATING ACQUIRED IMMUNITY
The exquisite specificity, long memory, and powerful effector mechanisms of the acquired immune system are its great strengths. Dramatic examples of the power of interventions targeting the acquired immune system are the eradication of epidemic smallpox by vaccination (Henderson 1976) and the development of passive antibody therapy as the first antimicrobial strategy for diverse infectious diseases from pneumococcal pneumonia to rabies (Buchwald and Pirofski 2003). Applying passive antibody therapy and vaccination to the treatment of infectious diseases is not a novel idea, but the committee identified several kinds of research that could allow these strategies to be used more widely and with greater effectiveness.
Passive Antibody Therapy
Passive antibody therapy is proven and immediately available for the treatment of infectious diseases. It involves the administration of an immunoglobulin molecule, or fraction thereof, to prevent or treat an infectious disease. Passive immunization by administration of preformed antibodies is generally well tolerated and may be particularly effective in immunocompromised people who are unable to mount a sufficient response to a vaccine. Several polyclonal antibody preparations and one monoclonal antibody, Respigam®, have been licensed for use against infectious diseases (Atkins et al. 2000). Despite the safety and efficacy of passive antibody therapy, further development is hampered by the specificity of antibody reagents, which limits each one’s use to relatively few patients and consequently affects the economics of development; the need for specific microbiological diagnosis before use; and the short half-lives of immunoglobulin fragments, which must have sufficient time to counteract the targeted microorganism before being cleared by the immune system.
Each of these limitations could be addressed in further research. Although specificity is a defining characteristic of antibodies, the recognition of new mechanisms of antibody action, including mediation of damage control and other immunomodulatory functions, suggests that broad-spectrum antibody reagents could be developed, which could act on diverse microorganisms (Casadevall et al. 2004). Availability of rapid and specific diagnostics for identification of microorganisms and of biomarkers of immune status would allow more effective deployment of passive antibody therapy.
The committee identified two potential applications of passive antibody therapy as particularly promising: the development of monoclonal antibodies for use as single reagents or cocktails to address the pressing problem of multiple-drug-resistant bacterial infections in hospitals and the development of antibody reagents that take advantage of normal antibody interaction with the innate immune system, for example, by inducing mediators of damage control (such as interleukin-10 and other cytokines) to serve as broad-spectrum stimulators or suppressors of immunity.
Three kinds of research are recommended that would contribute to the development and effective use of passive antibody therapy:
-
I-3.1 Understanding the relationships between antibody specificity, affinity, isotype, dose, and protective efficacy for pathogenic microorganisms would facilitate the development of antibodies that bind more effectively and are cleared more slowly. This would improve understanding of the optimal dose and timing of administration of passive antibodies.
-
I-3.2 Identifying the specific mechanisms by which antibodies interact with the innate and cellular immune systems to mediate damage control could allow the development of antibodies that are active against multiple agents. This research would probably also identify innate immune modulators that could act as vaccine adjuvants.
-
I-3.3 Rapid and specific diagnostics aimed at defining not only the infectious agent but also the patient’s immune status could allow rapid identification of patients in whom passive antibody therapy would be effective and could be used to determine the correct dose and timing of passive antibody treatment.
Therapeutic Vaccination
Another type of intervention that takes advantage of the acquired immune system is active immunization in the form of therapeutic vaccination. Active vaccination has been successful in preventing many viral and bacterial diseases, and it remains an attractive option. Most available vaccines are used to prevent infectious diseases. However, the rabies vaccine is given after infection and induces a protective immune response before the onset of disease. Another example of therapeutic vaccination is the prevention of herpes zoster in infected older individuals by administration of the varicella virus vaccine. The rabies and varicella vaccines illustrate the potential usefulness of therapeutic vaccination for infectious diseases.
Because of vaccination’s long history of safety and effectiveness, the committee feels that high priority should be given to research that is likely to lead to more-effective vaccines. In addition to improving the design of vaccines themselves, further research in these areas is likely to contribute to the development of more-effective adjuvants (compounds that are administered with vaccines to enhance the immune response). Given that only one adjuvant (aluminum salts such as hydroxide) is currently FDA-approved, the need for different adjuvants that can drive different types of immune responses (mucosal, Th1) is pressing. The demonstration that the Haemophilus influenzae type B-outer-membrane-protein-complex-glyco-conjugate vaccine depends on TLR2 engagement suggests that TLR agonists may be useful as adjuvants for some vaccine formulations (Latz et al. 2004).
The development of vaccination for use in chronic infectious diseases—such as those caused by protozoal parasites, mycobacteria, fungi, HIV, and other viruses—was identified as promising. In chronic disease, the natural immune response is insufficient to clear the infection, and damage is caused by microbial action on host tissues or by the inflammatory response to the persistent microorganism. The identification of antigens that elicit beneficial and harmful immune responses remains largely empirical. The availability of molecular information on the nature of B and T cell epitopes, combined with an enhanced understanding of antigen processing, presentation and the effect of co-stimulation, might allow the design of vaccines that would elicit antibodies and cell-mediated responses that are effective in clearing or controlling chronic infections.
Current vaccines generally elicit antibodies of the IgG isotype, but natural infection elicits a wide range of antibody isotypes and a long immu-
nological memory (Holmgren et al. 2005). One reason that most vaccines raise a limited range of isotypes is that the mode of administration usually bypasses the normal, mucosal site of entry of many infectious agents, where the production of other isotypes is stimulated. Similarly, because current vaccination techniques do not perfectly mimic natural infection, the protection they provide is often not as long-lasting. Dendritic cell vaccination and the development of antigens that target dendritic cells were viewed as promising approaches to enhance vaccine effectiveness because dendritic cells are critical components of innate immunity that also initiate acquired immune responses.
The relatively recent discovery of positive and negative signaling pathways involved in the activation of T cells presents the possibility of regulating T-cell function during immunization. By augmenting positive signaling pathways for T cells such as the CD40-CD40L and CD28-B7 pathways, it may be possible to enhance T-cell help and subsequent antibody production in response to some vaccines. Conversely, engagement of negative signaling pathways, such as the PD1-PDL1 pathway, to derepress T-cell function during immunization may also enhance antibody responses.
The following research was identified as most likely to contribute to improved vaccines:
-
I-4.1 Defining the molecular nature of B- and T-cell epitopes and using this information to design novel vaccines and to identify the characteristics of protective antibodies.
-
I-4.2 Determining how to elicit protective non-IgG responses and simulate the mucosal response.
-
I-4.3 Devising strategies that target dendritic cells and new approaches to optimize antigen delivery to dendritic cells.
-
I-4.4 Characterizing positive and negative signaling pathways between T cells and antigen-presenting cells.
TAKING ADVANTAGE OF THE RESIDENT MICROBIOTA
The growing recognition that most interactions between humans and bacteria are benign, or even cooperative, was a prominent theme of the workshop. Although study of the human-bacteria ecosystem is still in its early days, the committee identified it as an extremely promising avenue for the eventual development of therapeutic immunomodulators. The ma-
nipulation of normal processes, such as interactions of the immune system with the normal microbiota, to ameliorate the effects of pathogens may offer novel, noninvasive, and inexpensive therapeutic strategies.
The resident microbiota is essential to immune development in neonates and disturbances of the microbial community cause an imbalance in human health (Eckburg et al. 2005; Hooper and Gordon 2001). In collaboration with the immune system, microbial communities form the first line of defense against microorganisms that are potentially damaging. Bacteria and other constituents of the normal microbiota act in concert with the immune system in a variety of ways, including production of antimicrobials, which are harmless to the residents themselves but deter interactions with other bacteria; and modulation of the activity of both the innate and adaptive immune systems. The normal microbiota plays a role in inhibiting inflammation despite presenting high concentrations of specific microbial molecules—microorganism-associated molecular patterns (MAMPs), such as bacterial LPS and peptidoglycan—that interact with TLRs and induce inflammation when the same MAMPs are presented by microbial pathogens (Rakoff-Nahoum et al. 2004). How such seemingly opposite effects are reconciled to generate the healthy state remains to be determined, but understanding how the immune system distinguishes “good” from “bad” bacteria is an intriguing target for the development of therapeutics.
Perturbations in the development and maintenance of microbial communities are thought to render the host susceptible to disease. For example, accumulating data support the “hygiene hypothesis”—that antimicrobial hygiene, which has had an enormous beneficial effect on public health, may be contributing to the recent rise in allergy and autoimmune disorders, such as inflammatory bowel disease (Isolauri 2004). In addition, the use of antibiotics disrupts the balance of microbial communities. Imbalance can lead not only to the overgrowth of normal microbial residents to population densities that cause host damage (as in the case of Clostridium difficile in the intestine) but also to enhanced susceptibility to new, potentially damaging, microorganisms.
The workshop participants acknowledged that so little is known about our normal microbiota and how it affects the activity of the immune system that improved knowledge would be essential before these partnerships could to be used effectively in therapeutics. In the long term, therapeutic agents that target the resident microbiota may take many forms, but the only current interventions that capitalize on beneficial human-bacteria re-
lations are probiotics. Probiotic refers to the ingestion or application of live bacteria to promote human health; an example is the consumption of yogurt to reintroduce lactobacilli after antibiotic treatment. The committee identified the development of probiotics as an approach to treating infectious diseases that could yield results more quickly than other interventions based on manipulating the normal microbiota. It recommends the following three applications as particularly promising:
-
I-5.1 The displacement of pathogens from a niche, such as the skin or the oral cavity.
-
I-5.2 The replacement of disrupted bacterial communities after antibiotic treatment.
-
I-5.3 The engineering of probiotic bacteria that can signal the immune system to generate immunomodulatory cells or downregulate inflammatory pathways.
The committee recommends three kinds of basic research that would contribute to more rapid progress in developing therapeutics that are based on using the resident microbiota to prevent or treat infectious disease:1
-
I-6.1 Defining the composition of the resident microbiota on the skin and the mucosal surfaces.
Research would be directed at defining how microbial communities form and mature with the human body; how the microbial members of a community interact with one another, with the host, and with more pathogenic microorganisms; how community composition differs between individuals in association with age, sex, race, haplotype, ethnic background, health status, immunocompromise status or diet. Such research efforts would benefit greatly from incorporation of the expertise of engineers and computational biologists and from enhanced emphasis on methods to characterize non-culturable microorganisms.
-
I-6.2 Defining how the composition of the normal microbiota affects the establishment of the antibody and T cell repertoires.
Research would focus on how much of the antibody and T cell repertoires of a healthy person reflect interactions with the normal microbiota and how the repertoires are altered in people who experience autoimmune diseases or are immunocompromised.
-
I-6.3 Identifying the mechanisms by which the microbiota signal the host to maintain homeostasis and prevent damage from host-microorganism interactions.
Research would focus on how the host prevents intolerance and generates immunoregulatory cells such as regulatory T-cells, specialized dendritic-cell populations and γ/δ T cells. Efforts should also be directed at defining the nature of the host-microorganism interaction when a potentially pathogenic microorganism is present but there is no disease. For example, some pathogenic microorganisms such as Neisseria meningiditis, the causative agent of bacterial meningitis, can be recovered from healthy people who show no signs of disease; in some cases, this state may confer heightened resistance to contracting the disease by stimulating an immune response (Jordens et al. 2004; Goldschneider et al. 1969a,b). Learning how interactions between pathogenic microorganisms and hosts can persist without causing disease and identifying the conditions that promote disease are critical for an improved understanding of microbial pathogenesis.
Finally, the committee highlighted the importance of using a variety of animal models to study the resident microbiota. In animal-microorganism interactions, studies with germfree and gnotobiotic mice over the last several years have greatly expanded our understanding of the dynamics of these relationships. Support for the development and maintenance of additional germfree animal facilities would hasten progress in this field. However, a single model cannot provide the entire picture, so efforts should be directed at supporting the development and exploitation of other vertebrate and invertebrate animal models to complement the existing systems. Such models should be used to characterize not only the normal interactions of animals with microorganisms, but also pathogenic ones.
CROSS-CUTTING RESEARCH NEEDS FOR THE DEVELOPMENT OF IMMUNOTHERAPY
-
I-7.1 Encouraging multidisciplinary research among microbiologists, immunologists, ecologists, and clinicians.
The preceding sections of this report divided the potential novel therapeutics recommended by the committee into sections based on the part of the immune system from which they derive or that they would affect. However, workshop participants repeatedly emphasized that traditional descriptions of the immune system fail to take into account the interconnectedness of its subsystems and its complex relationship with the nervous system and the resident microbiota. Academic departments and study sections of funding agencies continue to be organized into traditional categories, so microbiologists, immunologists, ecologists, and clinicians find it difficult to interact. Many of this committee’s recommendations (for example, determining the effect of the resident microbiota on the normal antibody repertoire) will require collaboration among disciplines and may fit poorly into traditional study sections. Therefore, the committee recommends that thought be given to ensuring that such studies attract broad collaboration and receive multidisciplinary review.
-
I-7.2 Developing rapid diagnostics and determining markers of immune status.
Because the immune reaction is so complex, a fundamental problem in the development of immunomodulators is measuring their effect. Rapid diagnosis of the agent responsible for damage and rapid assessment of individual immune status will be critical in the development of immunotherapy. Therefore, the committee recommends that studies be undertaken to help to provide reliable measures of immune status and host damage. An additional value of the development of more sophisticated markers of immune status would be greater ability to predict the efficacy of immunomodulatory treatments in patients with various kinds of immunocompromise. Such studies could take the form of prospective sampling of patients entering hospitals to look for markers predictive of susceptibility to infectious diseases or response to treatment.
Another promising approach would be to look for patterns of immune markers that correlate with particular infectious agents, stages of infection,
or individual variations in immune response. The gene-array, proteomic, and metabolomic technologies that would make such studies successful are increasingly cost-effective. The ability to share the results of such studies in accessible, searchable databases, however, lags behind the ability to collect information.
-
I-7.3 Developing appropriate animal models.
Animal models play a vital role in biomedical research by paving the way for the development of novel therapeutic agents. However, the predictive value of these models for the clinical situation can be highly uncertain, especially for complex diseases. Although most animal models have their limitations, infectious disease models have been particularly useful in the discovery and development of agents that eradicate pathogenic microorganisms. The predictive value of the models rests on the notion that the in vivo efficacy of an antibiotic is relatively independent of host factors. In contrast, animal models of immunomodulatory therapies, and especially therapies that address the modulation of innate immunity, are inherently less predictive, and their reliability depends on the researcher’s ability to understand and manipulate the complex immune system of the host. This increases the likelihood of late-stage failure of novel immunomodulatory therapies compared with what has been observed with antibiotic therapies historically. For example, there have been many failures of immunomodulatory agents in clinical trials for multiple sclerosis despite the success of these therapies in animal models (Wiendl and Hohlfeld 2002).
Rodents, especially mice, are the most commonly used model species for studying immune responses and a variety of sophisticated tools are available for studies in these animals. However, although there are many similarities between the immune systems of mice and humans, there are also striking differences (Mestas and Hughes 2004) that make it difficult to extrapolate data from animal models into the clinical disease reliably. Non-human primates offer alternative models that are potentially more predictive, but experiments with them are expensive and are constrained by ethical considerations. The development of infectious disease models that use severe combined immunodeficiency (SCID) mice in which the immune system is reconstituted with human immune cells could offer an attractive alternative, although the mixing of species introduces new complexities.
Transgenic mouse models have been used extensively to dissect components of the immune system and have led to many important insights. In
addition, relatively simple model organisms with well-defined genetics and developmental pathways, such as Caenorhabditis elegans and Drosophila melanogaster, have effective innate immune responses and can be used to study conserved host-defense pathways. The successful development and testing of immunomodulatory therapeutics in general will require alternative animal models and therefore, the committee recommends support for research that uses novel model organisms.
THE NEAR AND FAR HORIZON
Treatment regimens that modulate the immune response have the potential to revolutionize how infectious diseases are treated. Such immunomodulatory therapies would be important additions to the current arsenal, especially because they would be expected to act synergistically with conventional antimicrobial therapies that target disease-causing organisms directly (Tzianabos and Kasper 2002). Some types of immune therapies, such as antibodies and cytokines, are already in clinical use and their application can be readily expanded to treat major infectious-disease problems. Other types are likely to require more preclinical development and their ultimate utility is more difficult to predict. This is why great emphasis has been placed on the need to develop biomarkers that accurately reflect immune status so that the effects of immunomodulators can be better predicted. However, it should be remembered that history shows that it is possible to develop immunomodulatory strategies without understanding the full complexity of immunology, as evidenced by early vaccines and serum therapy. Hence, the complexity of the host-microorganism interaction and the fact that we do not yet fully understand it need not prevent us from pursuing the development of immunomodulating therapeutics.
REFERENCES
Abreu, M.T., and Arditi, M. (2004) Innate immunity and toll-like receptors: clinical implications of basic science research. J. Pediatr. 144, 421-9.
Andersson, U., and Tracey, K.J. (2003) HMGB1 in sepsis. Scand. J. Infect. Dis. 35, 577-84.
Angus, D.C., Linde-Zwirble, W.T., Lidicker, J., Clermont, G., Carcillo, J., and Pinsky, M.R. (2001) Epidemiology of severe sepsis in the United States: analysis of incidence, outcome, and associated costs of care. Crit. Care Med. 29, 1303-10.
Atkins, J.T., Karimi, P., Morris, B.H., McDavid, G., and Shim, S. (2000) Prophylaxis for respiratory syncytial virus with respiratory syncytial virus-immunoglobulin intravenous among preterm infants of thirty-two weeks gestation and less: reduction in incidence, severity of illness and cost. Pediatr. Infect. Dis. J. 19, 138-43.
Belkaid, Y., and Rouse B.T. (2005) Natural regulatory T cells in infectious disease. Nat. Immunol. 6, 353-60.
Beutler, B. (2004) Innate immunity: an overview. Mol. Immunol. 40, 845-59.
Binder, C.J., Shaw, P.X., Chang, M.-K., Boullier, A., Hartvigsen, K., Hörkkö, S., Miller, Y.I., Woelkers, D.A., Corr, M., and Witztum, J.L. (2005) The role of natural antibodies in atherogenesis. J. Lipid Res. 46, 1353-63.
Bouchon, A., Facchetti, F., Weigand, M.A., and Colonna, M. (2001) TREM-1 amplifies inflammation and is a crucial mediator of septic shock. Nature 410, 1103-7.
Bowdish, D.M.E., Davidson, D.J., Lau, Y.E., Lee, K., Scott, M.G., and Hancock, R.E.W. (2005) Impact of LL-37 on anti-infective immunity. J. Leukoc. Biol. 77, 451-9.
Bucala, R. (1994) MIF, a previously unrecognized pituitary hormone and macrophage cytokine, is a pivotal mediator in endotoxic shock. Circ. Shock. 44, 35-9.
Buchwald, U.K., and Pirofski, L. (2003) Immune therapy for infectious diseases at the dawn of the 21st century: the past, present and future role of antibody therapy, therapeutic vaccination and biological response modifiers. Curr. Pharm. Des. 9, 945-68.
Casadevall, A., Dadachova, E., and Pirofski, L. (2004) Passive antibody therapy for infectious diseases. Nat. Rev. Microbiol. 2, 695-703.
Casadevall, A., and Pirofski, L. (2003) The damage-response framework of microbial pathogenesis. Nat. Rev. Microbiol. 1, 17-24.
Choe, J., Kelker, M.S., and Wilson, I.A. (2005) Crystal structure of human toll-like receptor 3 (TLR3) ectodomain. Science 309, 581-5.
de Silva, E., and Stumpf, M.P. (2004) HIV and the CCR5-Δ32 resistance allele. FEMS Microbiol. Lett. 241, 1-12.
Dower, S.K., and Qwarnstrom, E.E. (2003) Signalling networks, inflammation and innate immunity. Biochem. Soc. Trans. 31, 1462-71.
Eckburg, P.B., Bik, E.M., Bernstein, C.N., Purdom, E., Dethlefsen, L., Sargent, M., Gill, S.R., Nelson, K.E., and Relman, D.A. (2005) Diversity of the human intestinal microbial flora. Science 308, 1635-8.
Finlay, B.B., and Hancock, R.E. (2004) Can innate immunity be enhanced to treat microbial infections? Nat. Rev. Microbiol. 2, 497-504.
Fournier, B., and Philpott, D.J. (2005) Recognition of Staphylococcus aureus by the innate immune system. Clin. Microbiol. Rev. 18, 521-40.
Germain, R.N. (2004) An innately interesting decade of research in immunology. Nat. Med. 10, 1307-20.
Goldschneider, I., Gotschlich, E.C., and Artenstein, M.S. (1969a) Human immunity to the meningococcus. I. The role of humoral antibodies. J. Exp. Med. 129, 1307-26.
Goldschneider, I., Gotschlich, E.C., and Artenstein, M.S. (1969b) Human immunity to the meningococcus. II. Development of natural immunity. J. Exp. Med. 129, 1327-48.
Heikenwalder, M., Polymenidou, M., Junt, T., Sigurdson, C., Wagner, H., Akira, S., Zinkernagel, R., and Aguzzi, A. (2004) Lymphoid follicle destruction and immuno-suppression after repeated CpG oligodeoxynucleotide administration. Nat. Med. 10, 187-92.
Henderson, D.A. (1976) The eradication of smallpox. Sci. Am. 235, 25-33.
Holmgren, J., and Czerkinsky, C. (2005) Mucosal immunity and vaccines. Nat. Med. 11, S45-53.
Hooper, L.V., and Gordon, J.I. (2001) Commensal host-bacterial relationships in the gut. Science 292, 1115-8.
Isolauri, E. (2004) Dietary modification of atopic disease: Use of probiotics in the prevention of atopic dermatitis. Curr. Allergy Asthma Rep. 4, 270-5.
Ito, S., Ishii, K.J., Gursel, M., Shirotra, H., Ihata, A., and Klinman, D.M. (2005) CpG oligodeoxynucleotides enhance neonatal resistance to Listeria infection. J. Immunol. 174, 777-82.
Janeway, C., Travers, P., Walport, M., and Shlomchik, M. (2001) Immunobiology: The Immune System in Health and Disease, 5th edition. New York: Garland Publishing.
Jarva, H., Jokiranta, T.S.. Würzner, R., and Meri, S. (2003) Complement resistance mechanisms of streptococci. Mol. Immunol. 40, 95-107.
Jordens, J.Z., Williams, J.N., Jones, G.R., Christodoulides, M., and Heckels, J.E. (2004) Development of immunity to serogroup B meningococci during carriage of Neisseria meningitidis in a cohort of university students. Infect. Immun. 72, 6503-10.
Kelly, D., Conway, S., and Aminov, R. (2005) Commensal gut bacteria: mechanisms of immune modulation. Trends Immun. 26, 326-33.
Klinman, D.M. (2004) Use of CpG oligodeoxynucleotides as immunoprotective agents. Expert. Opin. Biol. Ther. 4, 937-46.
Krieg, A.M. (2002) CpG motifs in bacterial DNA and their immune effects. Annu. Rev. Immunol. 20, 709-60.
Lang, K.S., Recher, M., Junt, T., Navarini, A.A., Harris, N.L., Freigang, S., Odermatt, B., Conrad, C., Ittner, L.M., Bauer, S., Luther, S.A., Uematsu, S., Akira, S., Hengartner, H., and Zinkernagel, R.M. (2005) Toll-like receptor engagement converts T-cell autoreactivity into overt autoimmune disease. Nat. Med. 11, 138-45.
Latz, E., Franko, J., Golenbock, D.T., and Schreiber, J.R. (2004) Haemophilus influenzae type b-outer membrane protein complex glycoconjugate vaccine induces cytokine production by engaging human toll-like receptor 2 (TLR2) and requires the presence of TLR2 for optimal immunogenicity. J. Immunol. 172, 2413-8.
Leist, M., and Jaattela, M. (2002) Burning up TNF toxicity for cancer therapy. Nat. Med. 8, 667-8.
McLoughlin, R.M., Witowski, J., Robson, R.L., Wilkinson, T.S., Hurst, S.M., Williams, A.S., Williams, J.D., Rose-John, S., Jones, S.A., and Topley, N. (2003) Interplay between IFN-γ and IL-6 signaling governs neutrophil trafficking and apoptosis during acute inflammation. J. Clin. Invest. 112, 598-607.
Mestas, J., and Hughes, C.C. (2004) Of mice and not men: differences between mouse and human immunology. J. Immunol. 172, 2731-8.
Nataro, J.P., ed. (2005) Colonization of Mucosal Surfaces. Washington, DC: ASM Press.
Noverr, M.C., and Huffnagle, G.B. (2004) Does the microbiota regulate immune responses outside the gut? Trends Microbiol. 12, 562-8.
O’Neill, L.A.J. (2005) Immunity’s early-warning system. Sci. Am. 292(Jan), 38-45.
O’Neill, L.A. (2003) Therapeutic targeting of Toll-like receptors for inflammatory and infectious diseases. Curr. Opin. Pharmacol. 3, 396-403.
Opal, S.M. (2003) Clinical trial design and outcomes in patients with severe sepsis. Shock 20, 295-302.
Polderman, K.H., and Girbes, A.R. (2004) Drug intervention trials in sepsis: divergent results. Lancet 363, 1721-3.
Rakoff-Nahoum, S., Paglino, J., Eslami-Varzaneh, F., Edberg, S., and Medzhitov R. (2004) Recognition of commensal microflora by toll-like receptors is required for intestinal homeostasis. Cell 118, 229-41.
Reumaux, D., Duthilleul, P., and Roos, D. (2004) Pathogenesis of diseases associated with antineutrophil cytoplasm autoantibodies. Hum. Immunol. 65, 1-12.
Rice, L., Orlow, D., Ceonzo, K., Stahl, G.L., Tzianabos, A.O., Wada, H., Aird, W.C., and Buras, J.A. (2005) CpG oligodeoxynucleotide protection in polymicrobial sepsis is dependent on interleukin-17. J. Infect. Dis. 191, 1368-76.
Rossignol, D.P., and Lynn, M. (2005) TLR4 antagonists for endotoxemia and beyond. Curr. Opin. Investig. Drugs 6(5), 496-502.
Smith, K.D., and Bolouri, H. (2005) Dissecting innate immune responses with the tools of systems biology. Curr. Opin. Immunol. 17, 49-54.
Tracey, K.J., Czura, C.J., and Ivanova, S. (2001) Mind over immunity. FASEB J. 15, 1575-6.
Tzianabos, A.O., and Kasper, D.L. (2002) Role of T cells in abscess formation. Curr. Opin. Microbiol. 5, 92-6.
Wiendl, H., and Hohlfeld, R. (2002) Therapeutic approaches in multiple sclerosis: lessons from failed and interrupted treatment trials. BioDrugs 16, 183-200.
Wilson M. (2005) Microbial Inhabitants of Humans. Cambridge, UK: Cambridge University Press.


























