
2
Evidence for Ecosystem Effects of Fishing
Recent scientific literature has raised broad and multiple issues concerning changes to marine populations and food webs caused by fisheries removals. Vitousek et al. (1997) argue that no ecosystems on earth, including those in the ocean, are “free of pervasive human interference.” Here the concern is with the influence of fishing. Yields from ocean fisheries are approaching their upper limits (Botsford et al. 1997), and with the total global harvest of marine capture fisheries1 reaching 84.4 million metric tons in 2002, the likely maximum potential of conventional target species appears to have been reached (FAO 2005, Garcia and Grainger 2005). One can assume that removals of this magnitude must have some appreciable impact on ocean ecosystems; but to what extent has the species composition and biodiversity of the ocean changed as a result of fishing? And to what extent have these changes altered the current and potential benefits from the ocean as well as the functioning of ocean ecosystems?
Traditional fisheries management has been predicated on biomass reductions to 30 to 50 percent of unfished levels to maximize production (Mace 2004). Therefore, the fact that total biomass of fished species has decreased over time is not surprising. Yet, fishing is both size selective and species selective, meaning that the abundance and mean size of fished species are often reduced, and the genetic structures of populations are potentially altered. Furthermore, species interactions are often complex and fishing can modify elaborate connections in marine communities and food webs. These changes to populations and communi-
ties can and do alter species interactions and the functioning of ecosystems. The relations between “cause” and “effect” are often, perhaps always, non-linear and may include shifts in the state of the whole ecosystem.
In this chapter, the mechanisms, evidence, and magnitudes of fishing’s effects on marine ecosystems through modification of populations and food-web structure and function are reviewed and evaluated. Topics addressed are: (1) changing the abundance of fished stocks and species groups, altering biodiversity, and changing the genetic structure of populations; (2) altering food-web structure and function through the dynamics of trophic cascades; (3) fishing down and through food webs; and (4) inducing regime shifts through either physical or biological forcing. This chapter also presents a discussion of the reversibility of fishery-induced changes and the possible time frames for recovery.
CHANGES IN ABUNDANCE AND BIOMASS
Declining biomass is an expected effect of fishing on populations and is necessary for the density-dependent increase in production that is the basis for sustainable fisheries harvests, but in many cases overfishing has resulted in the collapse of populations and the fisheries that depended on them (e.g., northwest Atlantic cod). Numerous papers point to the decline in food fish biomass in various areas: the North Atlantic (Christensen et al. 2003), West Africa (Christensen et al. 2004), southeast Asia (Christensen 1998), the Gulf of California (Sala et al. 2004), and broadly around the world (Gulland 1988, Pauly and Maclean 2003, Garcia and Grainger 2005). The following point, drawn from Hilborn et al. (2003, p. 368), is replicated frequently in the fishery literature:
United Nations (UN) Food and Agriculture Organization’s (FAO) estimate that “75 percent of the world’s fisheries are fully or overexploited” has been widely quoted. Considering that being fully exploited is the objective of most national fishery agencies (and therefore not necessarily alarming), of more concern is the estimate that 33 percent of the U.S. fish stocks are overfished or depleted.
For U.S. fisheries, the pattern of overfishing among regions and stocks is characterized by heterogeneity (National Marine Fisheries Service [NMFS] 2005). In the same waters, some stocks are overfished while other stocks are not. The same species can be overfished in some areas while not in others. The proportion of stocks that are overfished or are experiencing overfishing varies greatly among U.S. management areas. The average for known stocks in 2004 was about 28 percent overfished, but proportions among regions based on at least 25 stocks ranged from 10 percent to 44 percent of the stocks. The lesson to be learned from this is that effects of fishing on exploited stocks vary greatly among and within regions. A danger of overgeneralization is always present. The depletion pattern is spatially heterogeneous, both in U.S. fisheries and worldwide.
Myers and Worm (2003) analyze the worldwide decline in longline catch per unit effort (CPUE) of predatory fish communities brought about by industrialized fisheries. Their analyses of 13 oceanic and coastal fisheries include the tropical, subtropical, and temperate Atlantic, Pacific, and Indian Oceans; the Gulf of Thailand; Saint Pierre Bank; the Antarctic Ocean off South Georgia; and the Southern Grand Banks. Typical reductions in catch per unit effort in the longline fishery were 80 percent of original catches within 15 years of the onset of industrialized fishing (Figure 2.1). Their results have been widely quoted: “Using a meta-analytic approach, we estimate that large predatory fish biomass today is only about 10 percent of pre-industrial levels” (Myers and Worm 2003, p. 280). Further, they report that most newly fished areas showed very high catch rates,
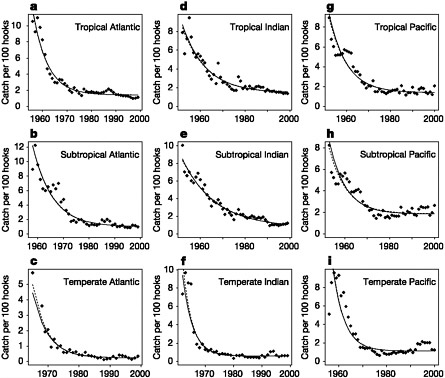
FIGURE 2.1 Time trends show decreasing catch per unit effort for nine different oceanic ecosystems. In these open-ocean communities, catch rates fell from 6–12 individuals per 100 hooks down to 0.5–2 during the first 10 years of exploitation. Relative biomass estimates from the beginning of industrialized fishing (solid points) are shown with superimposed fitted curves from individual maximum-likelihood fits (solid lines) and empirical Bayes predictions from a mixed-model fit (dashed lines).
SOURCE: Myers and Worm 2003; reprinted by permission from Macmillan Publishers, Ltd.
but declined to low levels after a few years, resulting in abandonment of once-productive areas.
Others have published less extreme estimates of biomass declines in the north central Pacific based on production models. For example, Cox et al. (2002a) estimate that between 1950 and 1998, biomass declined to 21 percent for blue marlin and 56 percent for swordfish. Conversely, estimated biomasses of juvenile bigeye (Thunnus obsesus) and yellowfin (T. albacares) tuna increased to 112 and 129 percent, respectively. Based on all their observations, Cox et al. conclude that the changes generally represent decreases in top predators and subsequent increases in small tunas, on which the top predators prey.
The changing abundance of tunas and billfish as a consequence of fishing is a subject of active analysis and the last word is likely not in. Some question the analyses and the magnitude of decline estimated by Myers and Worm (Walters 2003, Hampton et al. 2005, Polacheck in press), and many tuna and billfish managers note stable or increased catches of some species even after the alleged collapses. For example, Walters (2003) argues that Myers and Worm overestimate declines because they ignore unfished time and spatial cells in the database; he postulates that because the Japanese longline fishery was initially concentrated in a small area, the initial drops in catch per unit effort were not reflective of overall stock trends, an example of a phenomenon known as hyperdepletion (Hilborn and Walters 1992). In his alternative analyses, declines still occur, but were similar in one analysis and not as large in another. Hampton et al. (2005) argue that changes in catch per unit effort used by Myers and Worm are not a reliable indicator of abundance because fishermen target certain species in response to market demands and price and, additionally, the equatorial Pacific is not included in their analyses. When this area is included, the declines in CPUE in the western Pacific are 70 percent over 50 years for yellowfin tuna, but stable for bigeye tuna since about 1960, even though total catch has continuously increased. In their rebuttal, Myers and Worm (2005a) counter these criticisms and argue that their estimates of declines in large predators are conservative.
Polacheck (in press) makes a number of significant points in analyses of Indian Ocean data that suggest that the Myers and Worm estimates of declines in abundance are too high and that the apparent collapses are inconsistent with the increases observed in the pelagic fisheries over all years. Based on new analysis, Polacheck agrees that a rapid decline in longline catches occurred during the 1950s and 1960s, resulting in catch rates that were 20 percent of the early values by the early 1980s (Figure 2.2). But, in the 1980s, total catches, summed across all types of gears (longline, purse seine, and others), began to increase and by 2000 were two to six times greater than in the early 1970s. During this period of increased catches, the CPUE did not decline; in some species it even increased slightly. The growth in yield of these fisheries is due mostly to the introduction of new gear types—primarily purse seining, which targets smaller individuals nearer the surface than those caught by longlines—and to the introduction of existing
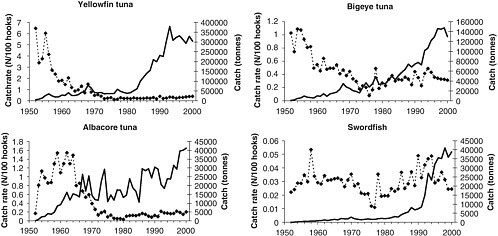
FIGURE 2.2 The annual nominal catch rates by Japanese longliners (dotted line) and estimates of total catch from all fisheries (solid line) are shown for the four principle species of tuna and billfish caught by longliners in the Indian Ocean. The increase in total catch is inconsistent with the hypothesized stock collapse indicated by the decreasing catch per unit effort.
SOURCE: Reprinted from Polacheck in press, with permission from Elsevier.
fisheries to new geographical areas. The observed increasing catch and stable catch per unit effort are inconsistent with collapsed stocks, even though the larger fishes captured in the longline fishery had been reduced in abundance. Apparently, the early declines in longline catches did not drive total population declines as these populations were later able to support fisheries targeting smaller individuals with different gears. As Cox et al. (2002a) pointed out for the Pacific, the declines of the largest fish were more than compensated by the increased production and capture of the prey of these large individuals. Production likely increased because cannibalism by and competition with the larger individuals was reduced, and the fishery was then able to take a higher proportion of individuals from slightly lower trophic levels.
Polacheck (in press) also was concerned that the Myers and Worm analyses, where they summed across all species in the Indian Ocean, had some biases as well. Myers and Worm did not point out that the species composition of the catch had changed over the 50 years, largely as a result of targeting different species for different markets. Finally, the spatial pattern of decline analyzed by Polacheck does not support the idea of serial depletion following the first years of exploitation in an area. Catch rates in newly fished grid cells were not greater than those in previously fished cells.
One can deduce from the subsequent analyses that the contentious findings in the Myers and Worm paper center mostly on large pelagic species; few argue that their conclusions do not represent the current situation for demersal fish included in the study. Also, as with all large-scale syntheses, their conclusions fail to realize the diversity of possible abundance declines when specific species and specific locations are examined. It seems likely, when these subsequent papers are taken into account, that the 90 percent decline reported by Myers and Worm (2003) is an overestimation. Yet even these newer papers report declines in the 65 to 80 percent range. Although the estimated magnitude of the decline may vary, all of these analyses indicate that abundances of large predators have been reduced. Polacheck (in press) points out that catches are, in fact, high and that the ecosystem effects of removing so many large predators have not been evaluated nor are the data being collected to evaluate such effects. He also warns that as the debate continues on the meaning of the sharp declines in catch per unit effort in the early years of these fisheries, it would be unwise not to respond to clear evidence of overfishing of some of these tuna stocks in recent years.
Biomass Recovery
As a response to reported declines, some analyses of worldwide data have been used to examine the ability of depleted stocks to recover to former abundance levels. After analyzing trends for 90 stocks, Hutchings (2000) finds little evidence for rapid recovery from prolonged declines, with the exception of early-maturing herring and related species. He concludes that although the effects of
overfishing on single species may, in general, be reversible (Myers et al. 1995), the actual time required for recovery appears to be considerable and species-specific. However, as Mace (2004) points out, Hutchings’s analysis does not consider whether or not there had been a reduction in fishing pressure after the stock declined. Indeed, stock recoveries were more likely to occur when exploitation was reduced (Hutchings and Reynolds 2004) or when stocks had not been severely depleted. But even in cases of reduction in fishing mortality, populations that have been driven to extremely low levels exhibited little or no recovery after 15 years. More recent data not included in this analysis may be more encouraging, as some stocks in New England show recent increases in biomass (Mace 2004). However, even if fishing mortality is reduced, recovery may be slow or may fail. The most well-known case is that of the northern Canadian cod, which has not rebounded after 15 years of low or no fishing and is unlikely to do so in the near future (Bundy 2005, Bundy and Fanning 2005).
Biodiversity and the Extinctions of Species
Biodiversity is intricately related to both community productivity and stability (Tilman 1996, Worm and Duffy 2003). Reductions in biodiversity could mean that the ecosystem is more vulnerable to changes that previously could be absorbed. Changes in species composition, species richness, or functional type also affect the efficiency with which resources are processed within an ecosystem, meaning that the biogeochemical functioning of an ecosystem might also be impaired by a loss of species richness (Naeem et al. 1999). At the very least, evidence reveals that the likelihood of fishery-induced regime shifts (discussed later in the chapter) may increase when humans reduce ecosystem resilience by decreasing stock size and altering size structure through fishing activities.
Worm et al. (2005) report quantitative declines in diversity for heavily fished areas as measured by species richness (expected number of species) and species density (species per unit of fishing effort) (Figure 2.3). For longline catches in the Atlantic and Indian Oceans from 1950 to 1999, richness declined by about 10 percent. For the Pacific, there was an initial decline, but levels in the 1980s and 1990s returned to those previously seen in the 1950s. Declines in species density were greater, about 50 percent in the Atlantic and Indian Oceans and about 25 percent in the Pacific Ocean. Such declines in diversity are significant, but they are less dramatic than the proportional declines in abundance reported by Myers and Worm (2003).
High diversity areas were mostly in the subtropical seas. In the Worm et al. (2005) analysis, the two measures of diversity were related to sea surface temperature (greater at intermediate latitudes), sea surface temperature gradient (greater in high gradient, frontal areas), and dissolved oxygen (lower with low dissolved oxygen). An El Niño signal was also documented in the interannual changes in diversity. Furthermore, the slopes of temporal decline in the two
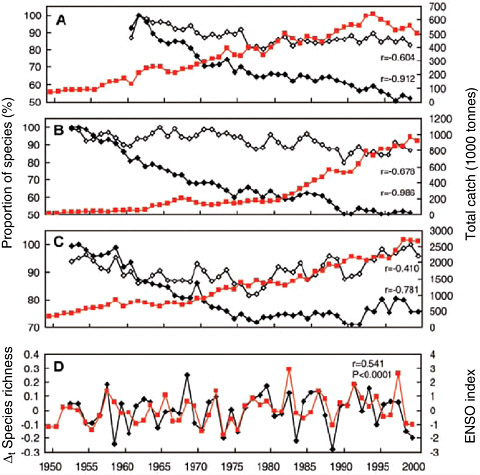
FIGURE 2.3 Proportion of species (in percent of maximum) per 50 individuals (species richness, open diamonds), per 1000 hooks (species density, closed diamonds), and total catch (squares) of tuna and billfish across the (A) Atlantic, (B) Indian, and (C) Pacific Oceans. Species richness (D) shows pronounced year-to-year fluctuations and decadal declines of 10 to 20 percent in all oceans (black line shows species richness in the Pacific; gray line shows El Niño Southern Oscillation [ENSO] index), a trend that reverses in the Pacific in the 1970s. Species density shows gradual declines, ~50 percent in the Atlantic and Indian Oceans and ~25 percent in the Pacific. These declines were most pronounced in intensely fished tropical areas, particularly in the Indian and Atlantic Oceans.
SOURCE: Reprinted from Worm et al. 2005, with permission of the American Association for the Advancement of Science. © 2005 AAAS.
measures of diversity were statistically related to a five- to tenfold increase in fishing effort, and the intensity of fishing was concluded to be a significant cause.
Of the five threats to marine biodiversity, Jackson et al. (2001) place fishing effects first both in magnitude and in time. In most coastal ecosystems they evaluated, fishing impacts occurred first and then were followed in time by pollution, habitat destruction, invasive species, and finally climate change. But as they rightly note, the fact that many overfished species still exist, even if they are commercially extinct, offers the potential for restoration of ecosystem structure and function if fishing and other factors can be controlled. This potential provides opportunities forgone in many terrestrial systems where the megafauna have been forever lost or reduced to an irrecoverable remnant. But can reductions in abundance from fishing lead to irrecoverable extinctions? Further, what kinds of species are more likely to go extinct and are there specific areas more susceptible to extinction owing to fishing?
Jackson et al. (2001) note that few species of marine megafauna are globally extinct. Only 12 global extinctions of marine species have been documented, including 3 mammals, 5 birds, and 4 invertebrates (Carlton et al. 1999); however, many species are now reduced to such low population levels that they are ecologically or commercially extinct. For example, a number of species of elasmobranchs (14 in the Gulf of Lions in the Mediterranean and 9 in the Bay of Biscay) have disappeared completely in trawl surveys (Aldebert 1997, Quero 1998). Examining the relationship between species extinctions and fishing, Dulvy et al. (2003) published a compilation of 133 known local, regional, or global extinctions of marine populations and reported that exploitation was the cause in 55 percent of these cases. Examples from their list, representing either local or regional extinctions due to exploitation, include the sea otter (Carlton et al. 1999, Day 1989), European sturgeon (Jukic-Peladic et al. 2001), angel shark (Quero 1998), and bumphead parrot fish (a decline that they state was due to the effects of an artisanal fishery). But even the authors note that the quality of evidence supporting some of the entries on their list is variable.
Often the bycatch species of the target fishery are those most in danger of precipitous declines since fishing effort is not influenced by their abundance. Bycatch increases the extinction probabilities for a growing number of non-target species. For example, net entanglement poses a major threat to the seriously endangered northern right whale, accounting for a quarter or more of annual mortalities (Kraus et al. 2005). Pacific populations of leatherback turtles and many albatross are also in steep decline owing to unsustainable mortality from longline fisheries (Lewison and Crowder 2003, Lewison et al. 2004b).
Using a simple population model, Worm et al. (1999) and Myers and Worm (2005b) estimate the sensitivity of species to extinction and the proportions of populations going extinct under various rates of fishing mortality. Not surprisingly, in their model, sensitive species with lower reproductive rates (fewer young and later age of maturity)—such as sharks—are twice as likely to go extinct as
bony fishes. But even bony fishes have high extinction probabilities at high rates of fishing. Modeled extinction rates are also greater when recruitment into the fishery occurs prior to maturity. These modeled results match general expectations and suggest that future extinctions may result from high rates of fishing. Hutchings and Reynolds (2004) argue that IUCN (The World Conservation Union) marine-fish extinction criteria (IUCN 2001) should not likely differ from terrestrial plants and animals and freshwater fishes. They back up this statement using analyses of empirical data on recovery probabilities for severely depleted populations. Dulvy et al. (2003) also conclude that high fecundity does not necessarily protect fish species from extinction risks, as has been previously believed.
While there have been few global extinctions as a result of fishing, the potential exists that future extinctions—including ecological extinctions—are possible, contrary to past thinking. The data presented for the declines in biodiversity and new examinations into the extinction potential for marine fish indicate that these issues deserve consideration in fisheries management decisions, especially as managers try to incorporate larger ecosystem concerns (the multi-species nature of fisheries management, accounting for effects of predation and competition).
GENETIC CHANGES IN POPULATIONS
Law and Stokes (2005) suggest that fisheries are an enormous uncontrolled selection experiment—removing some classes of individuals in preference to others. This “experiment” is also continually revised in some fisheries as managers set new regulations such as size limits, catch quotas, and closed areas. These regulations influence the behavior of fishermen and lead to mortality of marine organisms of particular sizes, life histories, and behaviors. As long as there is an appropriate genetic component to variation in traits under directional selection, there can be no question that marine populations evolve as a result of exploitation.
New experimental evidence suggests that strongly size-selective fisheries may reduce the growth potential of individuals, leading to less productive populations. Conover and Munch (2002) show that fishing can drive phenotypic evolution of life-history traits using experiments with the Atlantic silverside (Menidia menidia). They applied selective removal of small or large individuals in replicate populations and found that within four generations large genetic differences had developed between the populations. One such difference was a faster rate of body growth when small individuals were removed; the total biomass yield from these populations was nearly twice that of populations from which large individuals were fished. Within a small number of generations, size-selective fishing could bring about observable genetic change in life history, placing evolutionary pressure on the potential production of a population.
Since the 1970s, many major exploited fish stocks have undergone large changes in their life histories. Cod, haddock (Melanogrammus aeglefinus), and
pollack (Pollachius virens) stocks in the Northwest Atlantic, for example, have declined in age and length at maturity (Trippel et al. 1997). Since the early 1990s, these stocks have shown substantial declines in both age and size at maturity of approximately 20 percent. However, it is unclear whether these changes have a genetic basis. On the other hand, Olsen et al. (2004) show that fishing pressure did result in genetic changes for northern cod. Using probabilistic maturation reaction norms, they conclude that early-maturing genotypes are favored relative to late-maturing genotypes prior to the collapse of the stock in the early 1990s. They also suggest that this trend has begun to reverse due to decreased fishing pressure caused by the moratoria imposed in 1992.
Data has also been collected on Pacific salmon that indicate a decrease in mean size at age following years of selective fishing (Ricker 1981). For species such as salmon, where harvest often occurs during their spawning period, fishing pressure can select for such life history traits as time of spawning. The selection can be for either late or early spawners depending on the time of fishing. Fishing during the spawning period for other marine species can produce interesting results. In the case of the northeast Arctic cod, calculations suggest that the ancestral fishery on the spawning grounds was beneficial to the long-term production of the fishery since it selected for late maturation (Law and Grey 1989).
Studies like those cited above make it clear that fishing has the potential to alter life-history characteristics of populations; some of these changes are likely to be genetically based. Of course, fluctuations in the physical environment can have direct effects on life-history traits and the overall production of the fishery. These factors confound attempts to disentangle the contribution of genetic changes. However, new methods designed to distinguish temporal change in reaction norms from proximal effects of the environment support the theory that genetic changes are also taking place; results on size and age at maturation of northeast Arctic cod suggest a strong signal of change still remains in place (Heino et al. 2002). In general, it is proving increasingly difficult to account for large changes in the life histories of exploited fish on the basis of environmental factors alone.
THE PHENOMENA OF SHIFTING BASELINES
In marine conservation, there is often a lack of reliable baselines—measures of the pristine state or knowledge of what existed in the past—upon which to base future effort levels and fishery restrictions (Pauly 1995, Dayton et al. 1998, Myers and Worm 2003). Recent publications based on paleoecological, archeological, and historical data suggest populations of marine organisms were once much larger than currently observed in kelp forests, coral reefs, and estuaries (Jackson et al. 2001). Many methods for assessing fish stocks use historical records to estimate average stock sizes prior to exploitation and current levels of stock depletion relative to those levels. However, a real problem with long data sets is
often that both fishing technology and assessment methodologies change with time, breaking the time series into stanzas that tend to become “incompatible.” This leads to analyses being undertaken on only part of the data. In many instances, stock assessments only begin when catch-at-age data are available (typically in the 1970s or 1980s) because the particular stock assessment methods used require these types of data. As a result, analytical historical reconstructions often do not cover the entire period of exploitation, even if catch and survey data go back several more decades (NRC 1998). The problem can be severe in fisheries that already were intensively exploited before the systematic collection of fisheries statistics began, as has been shown for North Atlantic cod (Rosenberg et al. 2005).
An analysis of the pre-European densities of green sea turtles (Chelonia mydas) and hawksbill sea turtles (Eretmochelys imbricata) in the Caribbean relative to current population sizes concludes that these endangered turtles once grazed seagrasses and sponges in numbers ranging from tens to hundreds of millions (Bjorndal and Jackson 2003). These numbers are difficult for contemporary sea turtle biologists and marine ecologists to fathom. Another possible case of a shifting baseline is that of the northeast Pacific kelp forests (Dayton et al. 1998). The distribution and abundance of the major structure-forming species, the giant kelp (Macrocystis pyrifera), are highly influenced by changes in ocean productivity at a variety of scales from storm events to El Niño/Southern Oscillation fluctuations to other decades-long regimes of the Pacific Decadal Oscillation. Thus, the kelp “baseline” is a dynamic one. Sufficient historical data to quantify the roles for the major consumers in these communities (e.g., sea otters, large fishes, urchins, abalones) before exploitation are absent. But human exploitation began in this system some 10,000 years ago and likely changed the abundance of these consumers. Jackson (1997) makes a similar case for coral reefs; most modern reef ecology focuses on the last 50 years, but reef systems in the Caribbean had undergone centuries of change and loss to the megafauna before scientists began careful observations or experiments.
In the absence of reliable data about historical trends in abundance, it is not uncommon that diverging interpretations of the available data or different methodologies lead to widely different estimates of historical population levels. Estimates from catch records2 can vary from estimates based on fishermen’s logbook data, survey information, or other systemically collected data that can be more qualitative. One such example is the historical reconstructions of the great whales. Fishermen targeted great whales around the world for centuries; some are now extinct, many are endangered or threatened. It is known from catch records of depleted whale populations that a large proportion of their biomass was removed by targeted fishing throughout the world’s oceans (see Hilborn et al. 2003). Using
genetic analysis of neutral genetic variation, Roman and Palumbi (2003) estimate that the number of whales in the North Atlantic prior to whaling was an order of magnitude higher than historical estimates based on whalers’ logbook data. Although they explored various assumptions that might cause the genetic analysis to overestimate the baseline numbers of whales in the North Atlantic, their most conservative estimates are still three to five times higher than historical estimates based on catch records. However, both the demographic and the genetic models used to reconstruct historical abundances depend on uncertain model inputs, and a unified analytical framework may be needed to try to reconcile both approaches (Baker and Clapham 2004).
Only a decade ago, when Pauly (1995) introduced the concept of shifting baselines, the absence of early data was only part of the phenomenon. He argued that each generation of scientists and managers accepts as a baseline the conditions in marine systems and particular fisheries stocks that they observe early in their careers, and they then document changes over their careers. The next generation does the same, resulting in a gradual shift of the baseline in terms of species observed and abundances. A sense of history is often absent over periods longer than a career. This hypothesis was recently evaluated by surveying over 100 people representing three generations of fishermen from Mexico’s Gulf of California (Saenz-Arroyo et al. 2005). Fish populations have declined steeply in this region over the past 60 years and researchers probed the fishermen’s perceptions of these changes. Compared to young fishermen, older fishermen named five times as many species and four times as many fishing locations as having once been abundant but now depleted. The older fishermen recalled catch rates on their best fishing days as much as 25 times higher than those reported by the younger fishermen. These changes have occurred within living memory, but young fishermen’s perceptions of the state of the system differed dramatically from those of older fishermen.
A long-term view is not employed by scientists and managers either because early information is lacking or because they operate only on recent information. This precludes a vision to rehabilitate ocean ecosystems to earlier and perhaps more ecologically and economically valuable states. Taking a long-term view is an approach that can improve our perception of the world and how it works or might work (Magnuson 1990).
ALTERED FOOD WEBS
Fisheries not only affect populations but also alter the energy flows and species interactions in marine food webs and communities simply because all fished species are components of food webs and interact with other species through predation and competition. Thus, any alteration of a stock biomass or size and age structure also alters food-web structure, energy flow, and species interactions as well as the strength of these interactions in marine ecosystems.
Some responses can be compensatory in nature. The following section defines and explains what food webs are and how they are viewed and analyzed, reviews information on trophic cascades in food webs, and considers top-down (consumer-control) and bottom-up (resource-control) effects on species.
Food Webs and Trophic Interactions
Food webs are the road maps to known or even imagined species interactions and therefore display ecological connections. Their use as a descriptive tool extends back to the 1800s. Elton (1927), who termed them “food-cycles” and initially identified their central place in community ecology, used as his first example the North Sea web featuring diatoms, zooplankton, and copepods assembled by Hardy (1924). An unambiguous vocabulary has since evolved to describe their general features. “Links” connect individual predators to their prey; these are direct interactions. “Trophic level” has developed as a very general collective term, describing groups of species that are a similar distance in terms of energy transfers from the photosynthetic base.
A very simplified version of a food web is the ubiquitous food chain where smaller organisms are successively consumed by larger ones. Thus one reads of primary producers (organisms that are able to create biological energy through photosynthesis), herbivores, and primary and secondary carnivores.
Species feeding at more than one trophic level as adults (i.e., omnivores) are not easily categorized; assignment of a species to fractional trophic levels is increasingly common. Also, all fish feed at different trophic levels at different life stages. Due to their smaller size, juveniles are limited to feeding on small organisms, creating a situation where juveniles may be competing against species that will become their prey in later life stages. In more dynamical presentations of webs, a system of arrows scored by signs (+,-) can connect the entries. In this fashion it becomes possible to illustrate how a species can be “indirectly benefited,” either through a reduction in predation or through reduced competition for resources. Analyses suggest that 40 to 50 percent of observed ecological impacts are transmitted through these indirect linkages (Schoener 1983, Wootton 1994, Menge 1995).
The notion of “strength of interaction” also can be added as a measure of how changes in prey and predator abundance affect the mortality rate of the prey, and how such changes affect mortality, growth, and recruitment of the predator. These can be “strong” or “weak” individual interactions (MacArthur 1972, Paine 1980) and can be presented on a per capita basis. Beyond characterizing species interactions as weak or strong, there are concerns about the functional and numerical responses of predators to changes in prey abundance. Are predators able to maintain their food intake rate at low prey densities, thus increasing predation mortality on depleted preys in a depensatory way? Or is prey vulnerability limited by refuges or by the predator switching to alternative prey resources?
Characterizations of food webs fall into three general categories, each with its own strengths and weaknesses (Paine 1980). Descriptive or classical webs can display the enormous trophic complexity of natural systems, but they treat all linkages as being equivalent (Figure 2.4). This visually appealing format can be replaced by a predator–prey matrix that quantifies specific dietary details including cannibalism. However, this format remains unable to incorporate indirect effects, although the potential pathways are illustrated.
A second food-web category emphasizes the flow of energy along connected links: Species can be identified as important or not by the magnitude of their linkages as conduits of mass or energy transfer (Figure 2.5). A limitation of this approach is that the magnitude of energy flow is not always a useful predictor of population responses and, therefore, the importance of a species may be underestimated. For example, quantitatively “weak” trophic flows from juvenile fishes (typically of small biomass) to their predators can represent large mortality rates for the juveniles with potentially large impacts on recruitment rates (Walters and Martell 2004). This is relevant to the choice of species to represent in food-web models, so that potentially important interactions are not ignored. Ecosim/Ecopath models (Polovina 1984, Christensen and Pauly 1992, Walters et al. 1997) widely employed in fisheries science fall into this category. These models are dynamical and assume that all energy entering the system must be accounted for (i.e., input is equated to output).
The final category, now called interaction webs (Menge 1995), allows critically significant or keystone species (Paine 1969), trophic cascades (Paine 1980, Carpenter et al. 1985), and the dynamically related swarms of indirect effects to be highlighted (Figure 2.6). Interaction webs are best used to distill an assemblage down to its known, most important ecological features and to identify indirect effects that develop as a consequence of apex predator manipulation. Their robustness depends on both substantial biological intuition and some form of system perturbation sufficiently intense to reveal before and after conditions. Therefore, identifying a baseline (and its natural fluctuations) can be of fundamental importance.
Although the entirely descriptive portrayals were historically important, and have been the focus of numerous attempts to distill general patterns from the static details, the two other approaches (energy transfer and interaction webs) are more realistic because they acknowledge that species interact with direct consequences for both participants. Tradeoffs in their respective utilities involve the consideration of space as a resource, the role of indirect effects, and the consequences of requiring that the models have a mass balance. In the energy/mass balance webs and models, both predatory and potential competitive interactions are effectively displayed. These are popular in fisheries ecology (e.g., Trites et al. 1999, Schindler et al. 2002) but have not yet been incorporated into scientific advice for fishery management.
Ecosystems are biologically complex, and food webs seem the most appropriate vehicle to display the implied interactions and their potential consequences
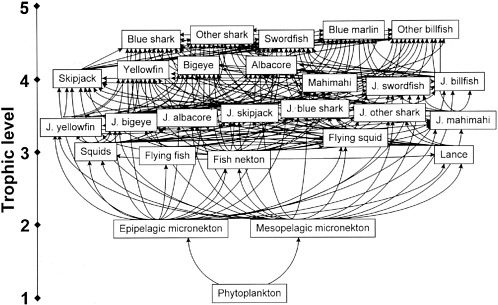
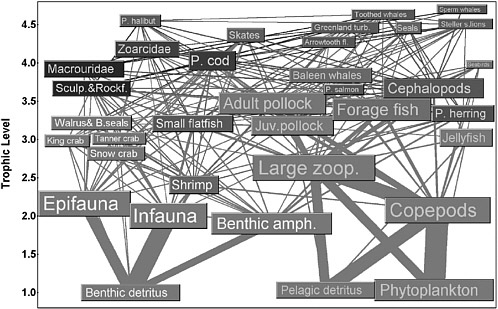
FIGURE 2.5 An example of a mass balance food web for the western Bering Sea. Unlike the previous example, relative biomass of each species is indicated by the size of the box, and the strength of each interaction is represented by the width of the connecting line.
SOURCE: Reprinted from Aydin and Livingston 2003, courtesy of the National Oceanic and Atmospheric Administration.
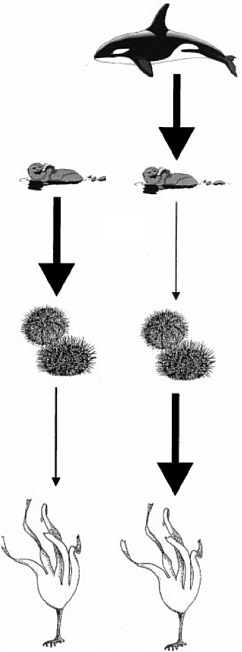
FIGURE 2.6 Interaction webs show the strength of species interactions, where the per capita interaction strength is indicated by the width of the arrow, and how these interactions change due to perturbation. For this example, the perturbation is the introduction of orcas into a food web previously dominated by otters.
SOURCE: Modified from Estes et al. 1998, with permission of the American Association for the Advancement of Science. © 1998 AAAS.
in a fisheries context. Regardless of how they are visualized, food webs involve a necessary simplification by aggregating species, particularly those at lower trophic levels, into single trophic categories. No web displays all the known feeding relationships; usually, only those considered meaningful or common are included. Further, a problem with trophic models, particularly in data-poor areas, is that interactions are often developed using parameters estimated for different times and different places (e.g., from databases), and sometimes for different species, assuming that the trophic flow is constant in space and time—a very unlikely assumption.
Despite these and other constraints, webs based on energy flux have been broadly applied to exploited ecosystems (Walters et al. 1999; Pauly et al. 2000; Christensen et al. 2002; Cox et al. 2002a, 2002b; Martell et al. 2002; Olson and Watters 2003; Hinke et al. 2004; Shannon et al. 2004). Interaction webs, by featuring strong interactors and indirect effects, generate insights on predator-controlled ecosystems, especially in those experimentally tractable systems where space utilization must be considered. Can the approaches of interaction webs and energy flow webs be melded into useful fishery management protocols? This goal is highly desirable, especially when there are hints of significant benthic-pelagic coupling, such as on Georges Bank and in the Gulf of Maine (Witman and Sebens 1992), on the eastern Bering shelf (NRC 1996a), or across the entire North Atlantic (Worm and Myers 2003).
TROPHIC CASCADES
Although humans have been fishing for millennia, the rapid expansion of industrialized fishing over the past 50 years has increased the impacts on marine food webs through both the magnitude of the removals and the exploitation of species in waters far beyond traditional fishing grounds. Removals of both target and non-target species (bycatch) can alter the abundance and productivity of components both higher and lower in the food web. But to what extent is the fishing effect limited to the population of the exploited species, and what additional effects cascade to the structure and function of the entire marine food web? It has been long accepted that the transfer of energy and thus organic matter upward through food webs influences community structure and overall productivity (Steele 1974). More recently, the importance of trophic cascades or the selective influence of consumers on the abundance and productivity of food-web components at lower trophic levels has emerged and is now recognized as possibly of equal importance in determining the dynamics of marine food webs (Estes et al. 1998).
Power’s (1992) paper states the obvious: Both top-down and bottom-up processes must characterize all ecosystems. The interesting questions revolve around their relative influence. The notion that the removal of top predators could have dramatic effects on food-web structure and function actually began with the
Hrbáček et al. study (1961) of riparian fish ponds in Prague and with Paine’s (1966, 1969) classic species-removal experiments in the rocky intertidal of the northeast Pacific. Data increasingly point to dramatic impacts on marine ecosystems caused by removal of large predators (Jackson et al. 2001).
Analyses of the statistical relations between different trophic levels in marine ecosystems provide evidence for both strong consumer effects (top-down) and strong influence of productivity at lower levels in the food web (bottom-up effects). Bottom-up productivity effects are apparent in comparisons among locations in the northeast Pacific (Ware and Thomson 2005). Strong cascading effects of removing top predators have been shown in the Scotian Shelf ecosystem off Nova Scotia using time-series data (Frank et al. 2005). (Both of these examples are described in Box 2.1.) In a comparison using freshwater lakes, Carpenter et al. (1991) estimate that the influence of consumer and nutrient effects on chlorophyll abundance are similar in magnitude; the error in the estimates of chlorophyll is lower when both top-down and bottom-up variables are included in the analysis. The joint influence of both consumer-driven and productivity-driven dynamics are not contradictory even if both are acting in the same ecosystem. The importance of each set of influences would be expected to differ among systems with respect to the magnitude of variability in production and the magnitude of variability in consumers over the time or space scales analyzed. Both could be equally important, or one or the other may dominate the dynamics.
Many other examples of predator-controlled cascades exist and they are more important to the discussion here than bottom-up influences since humans can be viewed as the top predator in many systems. These studies show that top-down control can affect every part of the system, with several of these examples ultimately affecting the foundational species, or habitat, of the ecosystem. In the Caribbean coral reef/sea grass system examined by Jackson et al. (2001), increased fishing leads to reductions in sharks and crocodiles and the extinction of Caribbean monk seals (Monachus tropicalis). Predatory fishes and invertebrates also decline due to fishing, as do grazers including manatees (Trichecus manatus) and sea turtles. The cascading effects propagate to the sea grass, sponges, and macroalgae (which increase) and to the corals (which decline). The current ecological role of grazing urchins, fish, sea turtles, and manatees is severely diminished and the primary producers are flourishing.
Silliman and Bertness (2002) use an interaction web to illustrate how a chain of coupled interactions potentially contributes to salt marsh grass (Spartina) destruction in the southeastern United States. Blue crabs (Callinectes sapidus) indirectly benefit the marsh grass by controlling the density of a marsh snail. However, these crabs are commercially valuable and their populations are declining. When snail populations were experimentally augmented, they reduced healthy Spartina stands to mud flats within eight months.
Perhaps the best known example of the unintended consequences of a trophic cascade linked to marine fishing/hunting—one that is relatively uncontaminated
|
BOX 2.1 Northeast Pacific—productivity control Differences in productivity, measured using surface chlorophyll, in various locations in the northeast Pacific are reflected in the abundance of zooplankton and resident-fish yields (Ware and Thomson 2005). For areas off of British Columbia, the correlation between chlorophyll and zooplankton was +0.92 and the correlation between zooplankton and resident-fish yields was +0.87. This study provides strong quantitative, empirical evidence for bottom-up trophic linkages between phytoplankton, zooplankton, and resident-fish yields among areas at different spatial scales. Evidence of climatic decadal changes in the North Pacific influencing productivity throughout the ecosystem also comes from analyses of time series (Francis et al. 1998, Hare and Mantua 2000). Scotian Shelf—predator control The trophic cascade on the Scotian shelf was initiated by the virtual elimination of the ecosystem-structuring role of the large predators (Frank et al. 2005). In a trophic cascade where the consumers are driving the dynamics, adjacent trophic levels should be negatively correlated and those separated by an intermediate trophic level should be positively correlated. Alternatively stated, the prey of the large predators (i.e., small pelagic fishes and bottom-living invertebrates) should increase and show a negative correlation, which they did (r = −0.61 to −0.76), and the prey of the prey (i.e., herbivorous zooplankton) should then decrease. As expected, the correlation between the large predators and the herbivorous zooplankton, which are two levels apart, was positive, r = 0.45, and that between the herbivorous zooplankton and the phytoplankton (the next level down) was negative, r = −0.72. The final step in this cascade resulted in a decrease of nitrogen concentration that resulted from an increase in abundance of phytoplankton. This example provides strong quantitative, empirical evidence for a trophic cascade initiated by fishing down the large predators. The ecosystem response was nonlinear and resulted from complex interactions in a food web that included humans at the top. |
by alternative explanations such as climate change, pollution, and habitat loss—is the ecosystem along the Aleutian Islands chain and North Pacific coast. The cascading relationship between sea otters (Enhydra lutris), kelps, sea urchins, other marine mammals, and fishing and hunting is a textbook example in marine ecology (Box 2.2).
Certain species exert strong controlling influences on marine food webs and these special species have an unexpectedly large impact on food-web structure and function. In other words, they have a large per capita effect. Such species are often found in top predator roles, which explains why systems are so highly altered when top predators are lost or severely reduced (Jackson et al. 2001).
|
BOX 2.2 Within the North Pacific food web, there is a direct correlation between sea otter and urchin populations—when sea otter populations decline, sea urchin populations increase owing to a lack of predation. When this happens, increasingly abundant sea urchins are able to virtually eliminate kelp populations from particular habitats. The alternate states of this community, kelps or urchin barrens, can persist for long periods of time as determined from the exploration of Aleut middens in Alaska (Simenstad et al. 1978). Because kelps are structure-forming species, they create habitat for many fishes and invertebrates; when kelps are lost, their associates are lost as well. The balance between a sea otter- or sea urchin-dominated system changes spatially as well as temporally. After being protected from overhunting, the recovering populations of sea otters changed nearshore ecosystems by reducing the abundance of urchins and thus promoting kelp forest expansion (Estes and Duggins 1995). However, in the late 1990s, sea otter populations started to decline precipitously over large regions of western Alaska. The best explanation for these declines seems to be increased predation by killer whales (Orca orcinus) (Estes et al. 1998), the influence of still another food-web component. In an orca-dominated system, sea otters are suppressed, urchins recover, and kelp forests decline. Populations of large pinnipeds, including Steller sea lions (Eumetopias jubatus), northern fur seals (Callorhinus ursinus), and harbor seals (Phoca vitulina) also collapsed in the western North Pacific beginning in the late 1970s. One interesting hypothesis suggests that the decline of marine mammals, including otters, in the North Pacific may be consistent with increased mortality, possibly from orcas, rather than from reduced food or any other bottom-up effect (Springer et al. 2003). Springer et al. (2003) attribute the sequential declines in this suite of marine mammals (Steller sea lions, northern fur seals, harbor seals, and sea otters) to whaling in the North Pacific ecosystem. Killer whales likely consumed great whales (in fact, they were first dubbed “whale killers” by the early whalers [Scammon 1874]), and when the great whales were suppressed due to hunting, killer whales expanded their diet to include harbor seals, fur seals, sea lions, and, finally, sea otters. |
Structure-forming species like kelps, sea grasses, and reef-forming corals are also strong interactors because they create a habitat that supports a high diversity of associated species.
This perspective of interaction in food webs is important to management. Humans have now become perhaps the strongest interactor in marine food webs. The previous examples and analyses point out that human policy decisions related to increasing or decreasing fishing (or hunting) pressures do occur and reverberate through trophic cascades in complex food webs and ecosystems. Sequences of events are set in motion and reveal themselves over decades. These changes have
sometimes resulted in higher allowable catches of certain species that are released from predation, but in other cases the results are less favorable. Many of these cascades have been surprises and certainly were unintended consequences of management actions. However, methods are being tested to see if future cascades can be predicted. In three out of four simulations for the central Pacific Ocean, Cox et al. (2002b) were able to predict changes in small tunas and other major prey as the abundance of the larger Thunnids was reduced.
FISHING DOWN AND THROUGH THE FOOD WEB
Long-term reductions in the mean trophic levels of fisheries landings have been measured worldwide. Explanations as to why trophic level changes in fisheries landings have occurred include (1) serial depletion of high-trophic-level species and subsequent replacement by lower-trophic-level species (i.e., the commonly held interpretation of fishing down the food web) (Pauly et al. 1998a), (2) sequential additions of lower-trophic-level fisheries while maintaining catches of higher predators (Essington et al. 2006), and (3) environmentally induced changes and natural cycles.
Pauly et al. (1998a) use FAO global fisheries landings statistics and estimates of trophic levels for fish and invertebrates determined by diet data and mass-balance trophic models. They conclude that the mean trophic level of fisheries landings worldwide have declined since 1950 (Figure 2.7). Globally, the overall mean decline in trophic level of fisheries landings was 0.05 to 0.10 per decade without the landings themselves increasing substantially. Further, most ocean areas were characterized as having declines in mean trophic level. The interpretation of the general nature of the steady declines was that they reflected a transition of the fisheries from long-lived, high-trophic-level piscivorous bottom fish toward short-lived, low-trophic-level invertebrates and planktivorous pelagic fish. Pauly’s group also points out that as fisheries moved from high-trophic-level fish to lower-trophic-level fish, the catches initially increased as expected, but then subsequently declined. These patterns of decline were complex and resulted from several different causes. In addition, the group reported that a few FAO ocean areas either showed no clear trend or perhaps even an increase in trophic level (e.g., the IndoPacific); these were interpreted as inadequacies in the statistics.
Fishing down the food web has been argued to cause a reduction in the number and length of the pathways in the food web, making the ecosystems less resilient to environmental fluctuations (Pauly and Maclean 2003). Other possible implications include reduction of apex predator guilds, restructuring of marine ecosystems, and a loss of biodiversity. Yet it is unclear whether the changes impact overall productivity or simply divert the system productivity to other species. Fishing down the food web is often considered indicative of unsustainable fishing and has been viewed as the summary index of negative effects of fishing on marine ecosystems (Barange et al. 2004). According to Steneck (1998,
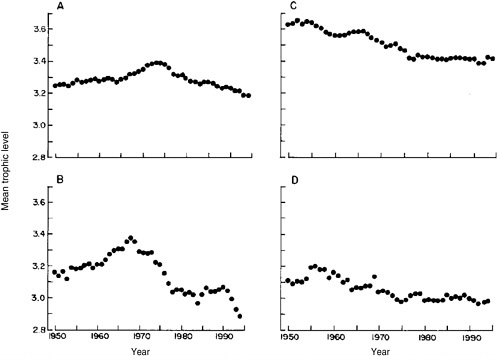
FIGURE 2.7 Trends of mean trophic level of fisheries landings in northern temperate areas from 1950 to 1994 for the (A) North Pacific, (B) Northwest and Western Central Atlantic, (C) Northeast Atlantic, and (D) Mediterranean.
SOURCE: Reprinted from Pauly et al. 1998a, with permission of the American Association for the Advancement of Science. © 1998 AAAS.
p. 430), “Fishing down trophic cascades is the antithesis of sustainable harvests and is clear evidence of ineffective management.” However, the cause of the reductions in mean trophic levels of fisheries is an active area of inquiry by the fishery and ecological science communities.
Caddy et al. (1998) criticize the analyses in Pauly et al. (1998a) based on four considerations: inadequate taxonomic resolution, the use of landings data as ecosystem indicators, aquaculture development, and eutrophication of coastal areas. These scientists do not disagree that a general decline in mean trophic level of marine landings was likely in many regions, but they are not convinced that the explanation is only “fishing down the food web.” The concerns expressed by Caddy et al. (1998) are addressed by Pauly et al. (1998b) and later by Pauly and Palomares (2005). These responses include clarification of several misunderstandings, and in several cases they reanalyze the data or remove certain types of questionable data from the analyses. These changes do not alter their original conclusions, but the debate is likely to continue.
Given the potential utility of monitoring mean trophic levels of fishery landings in the eastern tropical Pacific, the Inter-American Tropical Tuna Commission (IATTC) estimates trophic levels for time series of annual catches and discards from 1993 to the present for purse seine fisheries that target tunas at high trophic levels (IATTC 2004) but also catch small quantities of sharks, billfishes, dorado (Coryphaena spp.), wahoo (Acanthocybium solandri), and other species. High mean trophic levels occurred when the annual bycatch of large apex predators (e.g., billfishes and sharks) increased. On the other hand, anomalies that resulted in lower trophic levels in the time series were caused by increased bycatch of manta rays and other species that feed on plankton and other small, low-trophic-level species. Variation in mean trophic levels through the time series thus reflects the relative composition of the bycatch, either higher catches of large apex predators or higher catches of lower-trophic-level species. This analysis again portrays the complexity of the causes in any change in the mean trophic level of a fishery over time.
A useful extension of the continuing analyses of the mean trophic levels of fisheries is developed by Essington and colleagues (2006), who determine the frequency of two alternative causes to explain the decline in mean trophic level of catches. The two patterns, already recognized by Sprague and Arnold (1972) and Pauly et al. (1998a), were distinguished as (1) fishing down the food web and (2) fishing through the food web (Figure 2.8). The first pattern, fishing down the food web, involves serial depletion of fished species from the top of the food web. The second, fishing through the food web, involves serial additions of lower-trophic-level species to the catches. Declines in mean trophic level by fishing through the food web can be caused by initial depletion of predators in the early stages of a fishery, followed by release of prey from predation, thereby increasing their abundance and their contribution to the fishery. An example of fishing through the food web provided by Essington et al. (2006) is from the
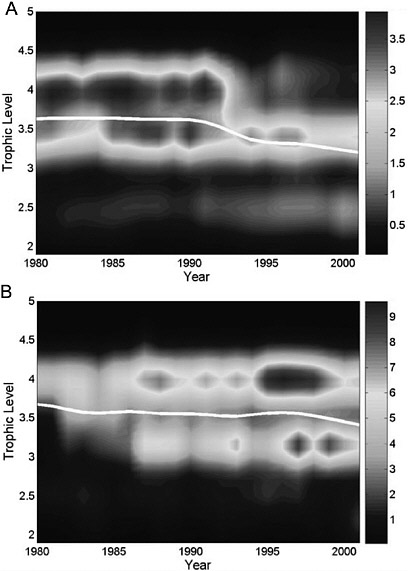
FIGURE 2.8 Total yearly catch is shown for each 0.1 trophic level increment, indicated by the color bar on the right (104 kg yr-1). The white line shows the mean trophic level. The Scotian Shelf (A) provides a typical example of the sequential collapse and replacement mode. The mean trophic level in fisheries landings exhibited a sharp decline from 1990 to 2001, owing to the collapse of the cod fishery, followed by a decline in the herring fishery and then the growth of the northern prawn fishery. In contrast, the Patagonian Shelf (B) exhibited a similar decline in mean trophic level over the same time period, but landings of high-trophic level species generally increased over this period, while new fisheries developed for lower trophic species.
SOURCE: Essington et al. 2006. © 2006 National Academy of Sciences, U.S.A.
Patagonian Shelf where the mean trophic level of the catch declined during the 1990s, but landings of a high-trophic-level species (Argentine hake, Merluccius hubbsi) increased during this period. The decline in trophic level of the catches from this region resulted from the addition of a new fishery for the shortfin squid (Ilex argentinus).
Essington et al. (2006) use annual catches by species and estimate the trophic level for each species to analyze trophic level of catches in 48 large marine ecosystems around the world as defined by Sherman (1991). Of the 48 ecosystems, 30 showed evidence of declining mean trophic levels, with an average decline greater than that described by Pauly et al. (1998a), who use more spatially aggregated data. Examples of both patterns were found, but sequential addition was more common than sequential depletion; in 23 of the 30 cases (77 percent), fishing through the food web prevailed. The sequential collapse and replacement mode was generally restricted to ecosystems in the North Atlantic, where management measures have been ineffectively implemented and many stocks have become grossly overfished.
One limitation that should be noted of both the Pauly et al. and the Essington et al. approaches derives from the use of catch data as an indication of stock status. In particular, the maintenance of high catches while a stock is being fished down may obscure a pattern of stock depletion. The example from the Patagonian Shelf is a case in point; while catches of the Argentine hake increased in the 1990s, the stock biomass decreased, reaching a critical status in 2000. These issues arise because most available data are fishery dependent, yet catch statistics are dependent on many factors including changing gear, effort, or area fished. This creates uncertainty about the extent to which the composition and trends in the catch reflect ecosystem changes.
Because both occur, the management implications of fishing down the food web (serial depletion of high-trophic-level species) and fishing through the food web (serial additions of low-trophic-level species) need to be addressed on a case-by-case basis. In particular, because fishing down and fishing through the food web affect multiple trophic levels simultaneously, nonlinear responses in the ecosystem may be more likely and managers may have to be sensitive to these possible changes. However, the overall message derived from examining both analyses is that it is not possible to determine the underlying force of change using only a trophic index. Therefore, while this index can be used to indicate that food-web level changes may be occurring, more data will be needed to judge the significance of the change to sustainable fishing.
Trophic Efficiency of Food Webs
Irrespective of what causes the change in the mean trophic levels of landings, a reduction in mean trophic level of an ecosystem would be expected to result in an increase in productivity available for fisheries harvest. Transfer efficiency
from one level of the food chain to the next averaged 10.13 percent (standard deviation 5.81) from analysis of 140 estimates of transfer efficiencies from 48 trophic models of aquatic ecosystems (Pauly and Christensen 1995). This is consistent with the usual 10 percent efficiency assumed over the last several decades, dating back at least to the work of Ryther (1969). This transfer efficiency should result in a tenfold increase in the available biomass for harvest at the next lower trophic level.
Few studies examine the relationship between fishing and ocean ecosystem productivity, but, in theory, an upper limit exists to the amount of production that can be removed by fisheries. Pauly and Christensen (1995) report that the primary production required to sustain reported catches plus discarded bycatch amounted to 8 percent of global aquatic primary production; however, this study includes freshwater fisheries as well. By ecosystem type, the requirements were only 2 percent for open-ocean systems, but ranged from 24 to 35 percent in freshwater, upwelling, and shelf systems (Pauly and Christensen 1995). Christensen (1998) further argues that only about 33 percent of primary productivity can be expected to be used for fisheries, meaning that for some areas, no growth in overall fisheries production can be expected.
However, if more productivity is available at lower trophic levels, one could argue that intentionally fishing down the higher trophic levels would be in the best interest of increasing long-term productivity from the ecosystem (Sprague and Arnold 1972). However, this may result in the harvest of species that were not previously viewed as desirable or those of lower economic value, and it might also require improved technology to catch and transform these new species into acceptable products (Garcia and Grainger 2005). The alternative scenario in which depletion of a top predator results in increased yields from valuable species is also possible. For example, shrimp and crab populations in the Northwest Atlantic increased greatly following overfishing of cod stocks (Worm and Myers 2003), and with high shellfish prices, the total value of the fishery is now greater than before the cod collapse.
A trophic index, called the FIB (fishing-in-balance) index, was developed by Pauly et al. (2000) to help judge whether additional yields expected by fishing lower in the trophic structure were actually occurring. The index is negative and decreases when the yield benefits are not realized by fishing lower trophic levels. In the Northwest Atlantic, this index initially increased as trophic levels and catches both increased and then declined after about 1970 and went strongly negative after 1990 without any yield benefits from fishing lower trophic levels. In a contrasting example for global tuna and billfishes as a group, while mean trophic level declined from 1950 to 2000, the FIB index steadily increased, suggesting the possibility of some yield benefits owing to fishing at lower trophic levels. Pauly and Palomares (2005) suggest that this resulted from a steady geographic extension of the fishery, but this is inconsistent with the fact that the mean trophic level was steadily declining during these years of increasing catches.
Are lower yields of the high predators more desirable in terms of consumption and/or profits than higher yields of lower trophic species? And what are the potential effects of manipulating fishery productivity on the overall ecosystem productivity? Certainly these issues demand more examination and the interactions between fisheries yield, ecosystem productivity, species composition, and fisheries market demands will need to be better understood and managed. Answering these types of questions will depend on the fishery and region in question, but the decision-making framework discussed in Chapter 4 may allow managers and stakeholders to consider these food-web interactions more explicitly in management decisions.
RESPONDING TO REGIME SHIFTS
If ecosystem effects of fishing are to be identified, and perhaps more importantly, if ecosystem-based management is to be possible, the effects of decadal-scale climate variability on the inherent productivity of ecosystems and the fish stocks they contain will require explicit recognition and consideration. Toward this end, the effects of climate regime shifts on fish population dynamics have received increasing attention in recent decades. A climate regime can be defined as a persistent state in climate, ocean, and biological systems, with a regime shift being an abrupt, nonrandom change from one state or baseline to another (Beamish et al. 2004). Polovina (2005) reports that changes in biological baselines in response to regime shifts exhibit several characteristics. These are evident when abundances of various species covering a range of trophic levels that persist around long-term baselines suddenly and coherently shift to a new baseline level where they again persist. These shifts occur both for exploited and unexploited species. Additionally, shifts in ecosystems spanning broad spatial scales, including ocean basins, are temporally coherent (Polovina 2005). Interannual variation can and does occur within a regime, but the climate conditions indicative of the regime are relatively consistent and persistent compared to the magnitude of change that occurs between regimes (King 2005).
In the Pacific Ocean, and more recently in the Atlantic Ocean, decadal-scale climate regimes have been documented (Hurrell and van Loon 1997, Miller and Schneider 2000). These regimes are driven largely by changes in the relative position and strength of dominant atmospheric high and low pressure systems and are identified as the Pacific Decadal Oscillation (PDO) and the North Atlantic Oscillation (NAO), respectively. Scientific research has focused on both descriptions of how physical and environmental conditions vary among regimes and how these conditions propagate through food webs (Benson and Trites 2002).
The fisheries and ecological literature provides many examples of environmentally induced shifts in ecological communities large enough to be considered a regime shift. As Barange et al. (2004) point out, historical, fishery-independent records show large natural cycles of exploited fish populations in the absence of
industrial fishing. For example, paleorecords of fish scales in anoxic sediments suggest large natural fluctuations in anchovy abundance off California during the last 1,700 years (Baumgartner et al. 1992) and in sockeye salmon abundance in the North Pacific over the last 2,200 years (Finney et al. 2002).
Changes in the population dynamics of fish species have been observed in phase with oceanic climate regime shifts across geographic regions (Francis et al. 1998, Benson and Trites 2002, Beamish et al. 2004). Kawasaki (1992) finds simultaneous variation in annual catches of Japanese, Californian, and Chilean sardine populations between 1910 and 1990. Strengths of year classes in several groundfish species, ranging from the California Current to the Bering Sea, show a dependence on the temperature regime; warm conditions were necessary but not sufficient for recruitment of a strong year class (Hollowed and Wooster 1992, 1995). Beamish (1995) reports other examples of fish population responses to climate regime shifts in the North Pacific.
Biological responses to regime shifts, especially in upper trophic levels, can lag or be masked by other processes and events (Miller and Schneider 2000). While direct, readily detectable responses to a regime shift have been observed in many cases, indirect food-web responses, variability in life history strategies, and longevity of some fish species may obscure or delay the onset of the effects of a regime shift on other fish populations (Benson and Trites 2002, Beamish et al. 2004). In different geographic areas, regime shifts could have opposite effects on the same species and, within an area, different species could respond in opposite ways (Benson and Trites 2002, Polovina 2005).
An interesting question is whether fishing, free from physical forcing, induces regime shifts. The discussion on trophic cascades above provides clear examples of ecosystem shifts induced by fishing at the top of the food web. This includes the northwest Atlantic example in Box 2.1, where the cascade contributed to an alternative dominant state, with a switch from groundfish to invertebrates. Fishing has clearly played a role in a number of radical shifts in marine ecosystems, but whether these changes constitute a regime shift is only determined by the longevity and the resiliency of fished species. Confounding the issues further, it is often difficult to tie changes in community composition to a single cause (Breitburg and Reidel 2005). Yet, Mangel and Levin (2005) make the case that many regime shifts could be driven in large part by fisheries. In addition, Collie et al. (2004) provide evidence that a discontinuous regime shift happened for Georges Bank haddock in 1965. The most likely trigger for this shift was the high fishing mortality during this time (Fogarty and Murawski 1998), but they were unable to rule out possible environmental contributions.
Clearly, intensive fishing can have drastic and long-lived effects on fished ecosystems. However, it is the combination of the two—climate change and fishing—that can result in the most severe effects. The response of fishery management can be critical in these situations, especially in a transition from a more productive to a less productive ocean ecosystem. And while a shift is sometimes
only definable several years after it has occurred, it may be possible and desirable to account for more regular oscillations in management decisions if a stock is known to be heavily fished and also sensitive to climatic regimes.
RECOVERY, STABILITY, AND MULTIPLE STABLE STATES
Stability and resilience are emergent traits of all ecosystems in the sense that they are properties of the integrated whole. They attain great importance for all, but especially for exploited or managed systems if a tipping point or threshold is exceeded under perturbation, and the system collapses to a different structure and species composition that may be an alternative stable state. The stressors can be biotic or abiotic, intrinsic or extrinsic, and can vary greatly in magnitude and impact. Holling (1973) defines “stability” as the rate of return to some prior state after a perturbation. More stable systems would recover more rapidly with reduced fluctuations. “Resilience,” on the other hand, depends on the maintenance of important relationships, thereby measuring the system’s capacity to absorb stresses imposed on its original organization. To the extent that resilience implies ecological continuity or persistence and stability implies the capacity to recover from a perturbation, both terms are useful and commonly employed in the basic ecological and fisheries literature. However, managers must also be concerned about abrupt system collapses and the formation of an alternative state, a feature anticipated in the behavior of nonlinear systems and one with potentially severe ecological consequences. The primary issue is whether these states are stable and resilient.
The reality of and environmental threats to anthropogenically altered ecosystems are extensively discussed in the literature (e.g., Sheffer et al. 2001, Beisner et al. 2003, Carpenter 2003). The crux of the matter is the rate of return—if possible—to prior conditions. The sudden die-off of a Caribbean grazer, the urchin Diadema (Lessios 1988, Knowlton 1992), permitted benthic algae to smother the usual extensive coral cover (Hughes 1994). However, Diadema appears to be slowly recovering (Edmunds and Carpenter 2001), and it will be interesting to see if the ecosystem is able to switch back to its previous state. Plagues of carnivorous starfish continue to devour the Australian Great Barrier Reef. Anthropogenic influences, including fisheries, have driven the plague (Birkeland 1997); reduction of these forces could permit reef recovery.
Once a stock has been depleted by fishing, complex competition and predator-prey interactions may prevent reversal after fishing has ceased. For example, one hypothesis proposed to explain the lack of recovery of Newfoundland cod involves increased predation on cod juveniles by a predator that, prior to the cod collapse, was kept under control by cod predation (Walters and Kitchell 2001). Changes in fisheries and desired harvest species can also impede recovery. For example, off of northwest Africa, large resources of sea breams were intensely exploited in the 1960s and 1970s. After their collapse, they were progressively
replaced by octopus stocks, and large fisheries for cephalopods developed as a consequence of increased market demand. However, sea breams were still taken as bycatch of the cephalopod fishery and the population never rebuilt to earlier levels despite not being targeted (Gulland and Garcia 1984).
The early debate about what was, or was not, a legitimate alternative and stable state has been extensively discussed in the primary ecological literature (e.g., Connell and Sousa 1983, Peterson 1984, Sutherland 1990). The debate identified two essential hallmarks: For community states to be comparable before and after perturbation, a common physical space must be involved, and the stressor must have vanished. That is, comparison of similar assemblages existing at spatially separated sites is inadmissible evidence, and the stressing force (e.g., persistent pollution or overfishing) must have been reduced to preperturbation intensity. The issue then becomes does the new state persist and, if so, for how long? Arbitrary judgments based on persistence, rate and degree of recovery, and magnitude of disruption are all required for analysis. For exploited populations, these are at the core of management decisions.
One can argue that anthropogenically influenced marine states are alternatives at both the population and community level. Whether they are stable may be difficult to distinguish in practice because a hysteresis from a single equilibrium value with a very long response time may appear stable. A single-equilibrium model should respond to a small intervention, albeit slowly. In contrast the multi-equilibrium model may not respond at all to a small intervention and may require a large intervention to initiate recovery.
Differences in recovery times of some stocks relative to others are not unexpected, however, and may be attributable in part to differences in the magnitude of compensatory reserve among interacting species. This fact alone could have a great deal of bearing on the way an ecosystem responds to the application or reduction of fishing pressure. In a relevant review, Rose et al. (2001) infer that life history and compensatory reserve are correlated and report that most of the world’s fisheries target periodic life history strategists—fish that are relatively long-lived, highly fecund, broadcast spawners. Periodic strategists depend upon occasional large year-classes to persist over time scales relevant to the life expectancy of populations rather than that of individuals. Therefore, these species often are resistant to overfishing (i.e., have high compensatory reserve) because effects of removals are distributed over many year classes. But they generally do not recovery quickly (i.e., they are not resilient), even after fishing ceases, because of long population generation times.
On the other hand, species like the early maturing herring are opportunistic strategists (Winemiller and Rose 1992), that is, they are short-lived, colonizing type species that often exhibit boom-bust cycles in the face of fishing. These species often can and do respond quickly to changes in exploitation. In special
cases, population levels can become so low that depensatory mechanisms may preclude population recovery no matter what curative measures are taken to restore the population (Shelton and Healey 1999).
From an ecosystem perspective, the mix of life-history strategies within communities that comprise the system and, hence, the mix in capacity of strong interactors to compensate for losses due to directed fishing or bycatch, will undoubtedly govern recovery times in ways that will be nonlinear and hard to predict. The economics of directed fishing usually deter fishing a stock to the point where depensation becomes a real danger. But for many of the most dramatic stock collapses, high economic value actually increases this risk, particularly for species that command extremely high prices at the market, such as large tuna or several invertebrate species. Bycatch species are also at risk since they are poorly monitored and assessed. At the very least, compensatory responses may be more difficult to assimilate and interpret from the ecosystem perspective, even though they have been the tenet of sustainable fishing for much of the history of fisheries science.
Key to this report is whether fishery management approaches can promote recovery from a fishery-induced regime shift. In the example presented in Box 2.2, the ecosystem did move from one state to another and back again when fishing pressures changed. Most of these changes were neither anticipated nor planned, but they demonstrate that shifting from one state to another is possible when fishing practices are changed. One could argue that these switches were not managed and that the favored state had not been established as a goal of management in a broad sense. Another example is the previously mentioned switch from cod to invertebrates in the northeast U.S. fishery. This shift was not planned and has not switched back even though fishing pressure has been reduced. But this unintended change can be seen by some as a positive outcome due to the high value of the subsequent shrimp fishery and therefore recovery to the previous state is not necessarily desirable.
If recovery of populations and food webs to earlier states is determined to be desirable, it will usually be a slow process, especially for fishes with older ages of maturation. In some cases recovery may not occur. Another lesson from the Hutchings (2000) study is that prevention of declines and collapses may be easier than designing for recovery. A prudent approach might be to prevent additional declines and collapses by more conservative population management, to develop and test alternative scenarios for food-web and community recovery, and to initiate long-term programs for recovery to serve as adaptive management experiments in the real world. Further, sustainability and the avoidance of persistent and unwanted alternatives critically depend on better data on the abiotic and biotic factors increasingly impacting all natural resources.
MAJOR FINDINGS AND CONCLUSIONS FOR CHAPTER 2
Ecosystem-level effects of fishing are evident. Due to their extractive nature, fishing changes fish populations and therefore ecosystems. Reduced biomass and abundance, changes in size structure, genetic changes, and changes in trophic structure of ecosystems (including alternative states and altered productivity, species interactions, trophic cascades, and food webs) have all been documented as consequences of fishing. While some changes are the intended outcome of management actions, for many cases the measured effects are quantitatively and qualitatively more severe than management intended.
Large declines in overall abundance of many stocks are evident even though the scientific debate continues regarding the magnitudes and implications of the declines. Differences in reported declines of the same stocks have resulted from different assumptions, methods, and data used for the analyses; these issues are being worked out in the scientific literature. However, few deny there have been declines in numerous stocks and, in many cases, these declines have resulted in the loss of sustainable yields of particular species and species groups. Furthermore, the magnitudes of these declines vary spatially. Spatial variation is considerable in the condition of fisheries, different species, and geographically distinct stocks.
Effects of fisheries removals can cascade through marine ecosystems. Direct predator-prey interactions and indirect effects with lower or higher trophic levels are certain. Such cascading effects are often unpredictable and management actions frequently have surprising results that add an element of risk in predicting management outcomes. Further, some of the greatest long-term impacts of fishing have been observed in non-targeted ecosystem components. Many species, including marine mammals, seabirds, sea turtles, sharks, corals, kelps, and sea grasses have been affected negatively by fisheries either directly through bycatch or habitat damage, or indirectly through altered food-web interactions.
Both fishing down the food web (sequential depletion) and fishing through the food web (sequential addition) are occurring. Several lines of evidence suggest that the trophic structure of marine ecosystems is changing. However, studies based on landings data alone can be misleading because these data do not directly translate into accurate assessment of abundances in nature and may not reveal the underlying causes of trophic change. A single index of the mean trophic level of landings can be used as an indicator that one of these effects may be occurring. But because the management response will differ based on whether the cause is fishing down or fishing through, management decisions cannot be made from the index alone.
Regime shifts caused by physical forcing, fishing, or both do occur. Regime shifts caused by physical forcing are important to recognize for management purposes, especially when the shift results in lower ecosystem productivity. In addition, for systems that have changed primarily due to fishing activities, reversing ecosystem effects is not guaranteed because multiple stable states in communities may resist returns to an earlier state. The combination of the two creates conditions where recovery is even more uncertain.
The shifting baselines phenomenon does occur. When management actions and goals are likely to be influenced by shifting baselines, possible earlier conditions become a significant consideration. The policy implications are important with respect to setting rebuilding targets and strategies. For example, potential rebuilding targets for stocks may be set lower than what actually could be achieved for stocks released from fishing pressure.
Realizing that there is a theoretical limit to the productivity that can be taken from the oceans and that we may currently be at or approaching that limit, food-web interactions will become increasingly important in future fisheries management decisions. With the inability to change one ecosystem component without affecting numerous others, society will need to determine which ecosystem components are the most desirable for harvest, and then managers will need to implement policies designed to maximize this desired production while recognizing that this will affect other species. Future management advice should include ways to examine such interactions if the desired mix of fisheries and other ecosystem services are to be realized and achieved and if undesired or irreversible states are to be avoided.




































