
Summary and Assessment
ENDING THE WAR METAPHOR: THE CHANGING AGENDA FOR UNRAVELING THE HOST-MICROBE RELATIONSHIP
The History of Medicine
2000 B.C.—Here, eat this root.
1000 A.D.—That root is heathen. Here, say this prayer.
1850 A.D.—That prayer is superstition. Here, drink this potion.
1920 A.D.—That potion is snake oil. Here, swallow this pill.
1945 A.D.—That pill is ineffective. Here, take this penicillin.
1955 A.D.—Oops…bugs mutated. Here, take this tetracycline.
1960–1999—39 more “oops.” Here, take this more powerful antibiotic.
2000 A.D.—The bugs have won! Here, eat this root.
—Anonymous (WHO, 2000)
In 1967, U.S. Surgeon General William H. Stewart told a White House gathering of health officers that “it was time to close the book on infectious diseases and shift all national attention (and dollars) to what he termed ‘the New Dimensions’ of health: chronic diseases” (Garrett, 1994; Stewart, 1967). In the ensuing years, Americans became intimately acquainted with a range of emerging infections including Legionnaire’s disease, toxic shock syndrome, AIDS, Lyme disease, West Nile encephalitis, and SARS. Complacency has given way to concern regarding a spectrum of microbial threats—including antimicrobial-resistant pathogens, emergent and reemergent diseases with pandemic potential, and outbreaks of exotic viruses such as monkeypox—propelled by a seemingly inevitable convergence of biological, environmental, ecological, and socioeconomic factors (IOM, 2003a,b). At the same time, the association of various chronic dis-
eases with microbial infection (e.g, peptic ulcer with Helicobacter pylori, liver cancer with hepatitis B and C viruses, and Lyme arthritis with Borrelia burgdorferi) has deepened respect for the destructive potential of infectious agents (IOM, 2004).
Infectious diseases continue to cause high morbidity and mortality throughout the world, particularly in developing countries. In 2001, infectious diseases accounted for an estimated 26 percent of deaths worldwide (Kindhauser, 2003). Moreover, there are indications that the tide of human conquest over microbial pathogens is turning. Over the last 30 years, 37 new pathogens have been identified as human disease threats, and an estimated 12 percent of known human pathogens have been recognized as either emerging or reemerging (Merell and Falkow, 2004). Having fallen steadily since the turn of the century, the number of deaths attributable to infection in the United States began to increase in the early 1980s, due in large part to the HIV/AIDS pandemic (Armstrong et al., 1999; Lederberg, 2000).
In the face of these challenges, the metaphor of “war” on infectious diseases—characterized by the systematic search for the microbial “cause” of each disease, followed by the development of antimicrobial therapies—can no longer guide biomedical science or clinical medicine. A new paradigm is needed that incorporates a more realistic and detailed picture of the dynamic interactions among and between host organisms and their diverse populations of microbes, only a fraction of which act as pathogens. To explore the crafting of a new metaphor for host-microbe relationships, and to consider how such a new perspective might inform and prioritize biomedical research, the Forum on Microbial Threats of the Institute of Medicine (IOM) convened the workshop, Ending the War Metaphor: The Changing Agenda for Unraveling the Host-Microbe Relationship on March 16 and 17, 2005.
Workshop participants reviewed current knowledge and approaches to studying the best-known host-microbe system—the bacterial inhabitants of the human gut—as well as key findings from studies of microbial communities associated with other mammals, fish, plants, soil, and insects. Participants and discussants also considered the evolutionary and environmental origins of pathogenesis and reviewed recent findings describing how hosts recognize and respond to pathogens. Additional presentations and discussions addressed the complexity of microbial communities and ecological relationships among pathogens, such as zoonoses, that infect multiple hosts. Finally, participants examined the prospects for manipulating host-microbe relationships to promote health and mitigate disease.
The workshop’s primary goal of replacing the war metaphor for infectious disease intervention represents an expansion of the Forum’s focus on microbial threats to health. The perspective adopted herein is one that recognizes the breadth and diversity of host-microbe relationships beyond those relative few that result in overt disease.
ORGANIZATION OF WORKSHOP SUMMARY
This workshop summary report is prepared for the Forum membership in the name of the editors as a collection of individually authored papers and commentary. Sections of the workshop summary not specifically attributed to an individual reflect the views of the editors and not those of the Forum on Microbial Threats, its sponsors, or the Institute of Medicine (IOM). The contents of the unattributed sections are based on the presentations and discussions that took place during the workshop.
The workshop summary is organized within chapters as a topic-by-topic description of the presentations and discussions. Its purpose is to present lessons from relevant experience, delineate a range of pivotal issues and their respective problems, and put forth some potential responses as described by the workshop participants.
Although this workshop summary provides an account of the individual presentations, it also reflects an important aspect of the Forum philosophy. The workshop functions as a dialogue among representatives from different sectors and presents their beliefs on which areas may merit further attention. However, the reader should be aware that the material presented here expresses the views and opinions of the individuals participating in the workshop and not the deliberations of a formally constituted IOM study committee. These proceedings summarize only what participants stated in the workshop and are not intended to be an exhaustive exploration of the subject matter or a representation of consensus evaluation.
THE RISE AND FALL OF THE WAR METAPHOR
More than a century of research, sparked by the germ theory of disease and rooted in historic notions of contagion that long precede Pasteur and Koch’s 19th-century research and intellectual synthesis, underlies current knowledge of microbe-host interactions (Lederberg, 2000). This pathogen-centered understanding attributed disease entirely to the actions of “invading” microorganisms, thereby drawing the lines of battle between “them” and “us,” the injured hosts (Casadevall and Pirofski, 1999). Although it was recognized in Koch’s time that some microbes did not cause disease in previously exposed hosts (e.g., milkmaids who had been exposed to cowpox did not become infected with smallpox), the fact that his postulates1 could not account for microbes that did not cause
disease in all hosts was not generally appreciated until the advent of vaccines and the subsequent introduction of immunosuppressive therapies in the 20th century (Casadevall and Pirofski, 1999; Isenberg, 1988). By then, the paradigm of the systematized search for the microbial causes of disease, followed by the development of antimicrobial and other therapies to eradicate them, had been firmly established in clinical practice.
The considerable impact of this approach, further enabled by improvements in sanitation, diet, and living conditions in the industrialized world, served to cement the belief that humanity was engaged in a war against pathogenic microbes, and that we were winning (Lederberg, 2000). By the mid-1960s, experts opined that, since infectious disease was all but controlled, researchers should focus their attention on other difficult medical challenges, such as heart disease, cancer, and psychiatric disorders. This optimism and complacency was shaken with the appearance of HIV/AIDS in the early 1980s, and was dealt a further blow with the emergence and spread of multidrug-resistant bacteria. As these experiences began to lead researchers to reexamine the host-microbe relationship, additional reasons to do so began to accumulate: pandemic threats from newly emergent (e.g., SARS) and reemergent (e.g., influenza) infectious diseases; lethal outbreaks of Ebola, hantavirus, and other such exotic viruses; and a new appreciation for the associations of various chronic diseases with prior microbial infections, as noted above.
Forum member Joshua Lederberg has envisioned the future of humanity and microbes as “episodes of a suspense thriller that could be entitled, Our Wits Versus Their Genes” (Lederberg, 2000). Our wits have so far afforded us increased longevity and reduced mortality from infectious disease, but the defenses we have mounted to make these gains are no match, over the long run, for the rapidly changing and adaptable genomes of microbial pathogens. We are vulnerable not only to emerging infectious diseases, but also to less treatable strains of pathogens (e.g., Staphylococcus aureus, Streptococcus pneumoniae) once seemingly conquered. The global health threat and economic burden posed by microbial resistance to therapeutics was highlighted in a recent Forum workshop, in which participants concluded that the management of microbial resistance over the long term would require “a sea of change…in how we view the ecology and evolution of infection” and the recognition of resistance “as an integral part—not an aberrant part—of the ecology of microbial life” (IOM, 2003b). However, doing so will require a far greater understanding of the evolutionary processes that underlie the development of resistance.
Changes in global ecology, climate, and weather are also increasing human vulnerability and exposure to microbial threats, as are more localized factors such as economic development, land use, travel, poverty, and war. A convergence of biological, environmental, sociopolitical, and ecological factors, depicted in Figure S-1, can be seen to influence the host-microbe relationships that lie at the core of disease emergence.
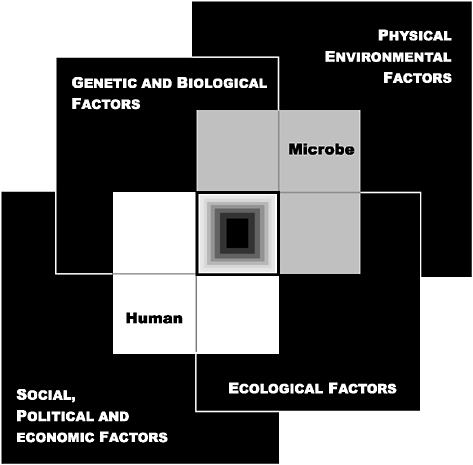
FIGURE S-1 The Convergence Model. At the center of the model is a box representing the convergence of factors leading to the emergence of an infectious disease. The interior of the box is a gradient flowing from white to black; the white outer edges represent what is known about the factors in emergence, and the black center represents the unknown (similar to the theoretical construct of the “black box” with its unknown constituents and means of operation). Interlocking with the center box are the two focal players in a microbial threat to health—the human and the microbe. The microbe-host interaction is influenced by the interlocking domains of the determinants of the emergence of infection: genetic and biological factors; physical environmental factors; ecological factors; and social, political, and economic factors.
SOURCE: IOM (2003a).
PATHOGENESIS REVISITED
The vast majority of microbes do not produce overt illness in their hosts, but instead establish themselves as persistent colonists that can be described as either low-impact parasites (e.g., causes of asymptomatic infection), commensals (or-
ganisms that “eat from the same table,” deriving benefit without harming their hosts), or symbionts (establishing a mutually beneficial relationship with the host) (Blaser, 1997; Merrell and Falkow, 2004). These states, while separate, represent part of a continuum—one that extends to pathogenesis and disease—that may be occupied at any point by a specific microbial species through the influence of environmental and genetic factors (Casadevall and Pirofski, 2000, 2002, 2003). Persistent colonization of a host by a microbe is rarely a random event; such coexistence depends upon a relationship between host and microbe that can be characterized as a stable equilibrium (Blaser, 1997). In the case of microbes that cause persistent, asymptomatic infection, physiological or genetic changes in either host or microbe may disrupt this equilibrium and shift the relationship toward pathogenesis, resulting in illness and possibly death for the host (Merrell and Falkow, 2004).
Early views of pathogenesis and virulence were based on the assumption that these characteristics were intrinsic properties of microorganisms, although it was recognized that pathogenesis was neither invariant nor absolute (Casadevall and Pirofski, 1999). Over the course of the last century, the identification of increasing numbers of microbial pathogens and the characterization of the diseases they cause has begun to reveal the extraordinary complexity and individuality of host-pathogen relationships. As a result, it has become exceedingly difficult to identify what makes a microbe a pathogen. One response to this dilemma has been to define pathogenesis from the perspective of the host, who experiences disease only when the presence of a microbe (whether protozoan, bacterial, or viral) results in damage—whether that damage is actually mediated by the pathogen itself, or by the host’s immune response to it (Casadevall and Pirofski, 1999, 2003).
A broader view, reflected in many workshop presentations and discussions, considers how pathogens coexist within host-microbial communities and places infectious disease within an ecological context. This perspective acknowledges the ecological and evolutionary impact of advancing civilization—and particularly the “war on disease”—on host-microbe systems, and promotes a more realistic, deeper, and nuanced understanding of the relationships upon which these systems depend (Lederberg, 2000). The time has come to abandon notions that put host against microbe in favor of an ecological view that recognizes the interdependence of hosts with their microbial flora and fauna and the importance of each for the other’s survival. Such a paradigm shift would advance efforts to domesticate and subvert potential pathogens and to explore and exploit the vast potential of nonpathogenic microbial communities to improve health.2
|
2 |
Refer to p. 27 first paragraph under the section “Raising Awareness of the Host-Microbe Relationship” for further information on the paradigm shift. |
IT’S A SMALL WORLD: MICROBIAL COMMUNITIES OF THE GUT
The gastrointestinal (GI) tract represents an important and challenging system for exploring how microbial communities become established within their hosts, how their members maintain stable ecological niches, and how these dynamics relate to host health and disease (see Chapter 1). The complex, dynamic, and spatially diversified microbial community, or microbiota, of the human gut is believed to be comprised of at least 1013 microorganisms (Xu and Gordon, 2003). This estimation includes more than 800 species of bacteria (Bäckhed et al., 2005; Gordon, 2005) (most of which have not yet been successfully cultured in the laboratory), fungi, numerous viral species including bacteriophages, (Breitbart et al., 2003) archaea (e.g., methanogens), and eukaryotes (e.g., helminths and protozoa) (Dominguez-Bello, 2005; Fagarasan et al., 2002; Hylemon and Harder, 1998; Xu and Gordon, 2003). The collective genomes of the microbiota in the human gut, known as the microbiome, is approximately one hundred-fold larger than that of its host (Bäckhed et al., 2004; Savage, 1977; Xu and Gordon, 2003). Therefore, as Bäckhed et al. have recently argued, “It seems appropriate to view ourselves as a composite of many species and our genetic landscape as an amalgam” of the human genome and the microbiome (2005).
The complexity of the human gut microbiota has been studied using culture-based assays and, more recently, a variety of molecular methods. These include fluorescent in situ hybridization, terminal restriction fragment length polymorphisms, microarrays, and direct sequencing of 16S libraries (Breitbart et al., 2003). The latter has revealed the presence of only 8 of 55 known bacterial divisions in the human gut, but great diversity among them at the strain and subspecies level exists (Bäckhed et al., 2005; Winter et al, 2004). This pattern suggests strong selection pressure on these microbes and coevolution between them and their host—a conclusion supported by findings that reveal how Bacteroides thetaiotaomicron, a prominent intestinal anaerobe, maintains a stable ecological niche despite dietary shifts and attacks by bacteriophage and the human immune system (see Gordon in Chapter 1). Knowledge of the viral components of the gut microbiota is comparatively limited; however, a recent study of viral genotypes in human feces (Breitbart et al., 2003) suggests that bacteriophages play an important role in shaping the gut microbiome (Bäckhed et al., 2005). Methanogenic archaea in the gut appear to orchestrate the final step in processing plant polysaccharides, but their role in determining the structural and functional stability and diversity of the gut microbiota is largely unexplored (Bäckhed et al., 2005; Gordon, 2005).
The Molecular Basis of Mutualism
The gut microbiota acts as an exquisitely tuned metabolic “organ” within the host, according to presenter Jeffrey Gordon (see Chapter 1) (Bäckhed et al., 2004;
Casadevall and Pirofski, 2000; Xu and Gordon, 2003). Microbes and mammals have coevolved mutually beneficial (symbiotic) relationships, typically based on nutrient sharing, in which the microbiota perform functions that their hosts have not evolved. For example, mammals are inherently limited in their ability to break down polysaccharides, which represent an abundant energy source in a plant-based diet (Hooper et al., 2002). Instead of producing the enzymes for carbohydrate hydrolysis, mammals recruit a diverse community of microorganisms that allow them to make efficient use of a broad range of foodstuffs; the microbes, in turn, gain access to abundant, readily fermentable carbon sources. An especially complex version of this exchange occurs among ruminants, who obtain nutrients solely by digesting the bacteria that feed upon the grass and other fodder swallowed by their mammalian hosts, as described by presenter Maria Dominguez-Bello (see Chapter 3). Thus over the course of evolution, symbiotic gut bacteria have become master physiological chemists, employing a broad range of strategies to manipulate host genomes, Gordon observed.
Identifying the host genes targeted by gut microbes and the mechanisms by which they manipulate host gene expression could lead to novel approaches for preventing and controlling a variety of diseases and promoting human health. To explore such host-microbe interactions at a molecular level, researchers have introduced genetically mutable components of the human intestinal microbiota into germ-free animals (Bäckhed et al., 2004; Hooper et al., 1998, 2001, 2002; Rawls et al., 2004; Xu and Gordon, 2003); several such studies are described in Chapter 1. Examination of the transcriptional response of germ-free mice to colonization with B. thetaiotaomicron reveal that the bacterium modulates the expression of host genes known to influence a wide range of intestinal functions in addition to nutrient absorption, including mucosal barrier formation, xenobiotic metabolism, angiogenesis, and postnatal intestinal maturation (Hooper et al., 2001; Xu and Gordon, 2003). Related studies show that in addition to mediating energy harvest from the diet, the gut microbiota also influences energy storage by the host, and thereby, individual predisposition toward obesity (see Gordon in Chapter 1) (Bäckhed et al., 2004).
Additional research being performed in germ-free animals involves the initial establishment of microbiota in the gut (in humans, during the early days of infancy), its influence on host development (e.g., immunity), and the mechanisms by which hosts perceive and respond to the presence of colonizing microbes. Presenter Karen Guillemin (see Chapter 1) pursues these fundamental questions in germ-free zebrafish, an experimental system that simplifies analyses of microbial influence on host development while closely approximating GI tract and immune system maturation, as well as gut microbiota diversity, in mammals. Although this approach has demonstrated the pervasive influence of the microbiota over a variety of events in the maturation of the GI tract, it raises further questions regarding the potential for individual developmental variation arising from differences in microbiota from one member of a species to another. In humans, such
variation could accrue among contemporaries who live in different environments, as well as over the course of history.
Microbe-Microbe Interactions
The complex web of interactions that must occur among the denizens of the gut is even less well studied than those that take place between microbe and host, but undoubtedly no less important to gut function and development. Microbe-microbe relationships include nutritional interactions—such as the previously described metabolism of the end products of bacterial fermentation by archaea—and genetic exchanges that occur through transformation, phage transduction, and conjugation (see Salyers in Chapter 1). For example, Bacteroides species recently have been shown to acquire and transfer antibiotic resistance genes among distantly related bacteria (e.g., Escherichia coli) that colonize the same ecological niche (Whittle et al., 2002; Wilson and Salyers, 2003). While important as a mechanism in the spread of antibiotic resistance, with its attendant impact on public health, Salyers emphasized it as merely an indicator, the “tip of the iceberg” of pervasive genetic exchange among members of endogenous microbial communities.
In light of this discovery, further study of the gut as a “cauldron of microevolution” is clearly warranted (Gordon, 2005; Salyers, 2005; Wilson and Salyers, 2003). Key questions to be investigated include how and where microbial gene transfers occur, the extent to which such transfers have contributed to the evolution of pathogens, and the potential for such transfers to influence phenomena other than antibiotic resistance, such as host metabolism and microbial virulence.
IT’S A SMALL UNIVERSE: INSIGHTS FROM OTHER HOST-MICROBE SYSTEMS
The host-microbe environment of the human gut is complex, compelling, and likely to yield important scientific and medical insights, but the same can be said for microbial communities in plants, insects, and the soil (dubbed “nature’s GI tract” by presenter Jeffrey Gordon) that have received considerably less attention. One workshop contributor highlighted recent findings in these systems that suggest the importance of inter-microbe communication (see Handelsman in Chapter 2); another noted similarity between the strategies used by plant and animal pathogens and compared the defenses mounted against them by their disparate hosts (see Staskewicz in Chapter 2). Moreover, the gut is but one site of microbial colonization in mammals; recent studies of oral microbial communities in humans (Smoot et al., 2005) are discussed later in this summary and in Chapter 5 (see Relman), and new data are emerging rapidly on the microbial communities of the human female genital tract.
Camouflage and Communication in Microbial Communities
Many momentous discoveries in microbiology—the germ theory of disease, the discovery and characterization of viruses, the techniques of cell culture, and the analysis of cell differentiation—were first achieved by plant biologists, but were not recognized at the time by their peers in other disciplines (Handelsman, 2005). Thus, while it is not surprising that studies of plants and their associated microbial communities have added considerably to knowledge of host-microbe relationships, these findings have not been widely appreciated nor have they been well integrated with current understanding of the human gut microbiota. There is a need to take a broader view to include perspectives from other biological systems. Many significant biological control processes were originally recognized in plants—such as RNA silencing—only to be demonstrated decades later to be conserved and operational in mammalian systems as well.
Research by presenter Jo Handelsman offers a new opportunity to apply insights derived from host-microbe studies in plants, in this case, toward a new understanding of the importance of microbial signaling to host health. Handelsman and coworkers demonstrated that inter-microbe communications that lead to disease could be disrupted, and that beneficial lines of communication could be protected against pathogenic saboteurs (see Chapter 2). For example, they observed that plant diseases can be suppressed by treatments that modify the microbial community of the root to make it more like the community in the soil, a conclusion which they have dubbed the “camouflage hypothesis” (Gilbert et al., 1994).
These studies have led to further examinations of the interactions between endogenous microbes and disease outcomes in other host-microbe systems, with the most recent example being the gut of the gypsy moth caterpillar (Broderick et al., 2004). In the relatively simple gypsy moth system, the Handelsman lab has begun to explore how signaling within microbial communities influences their ability to protect their hosts from disease and other perturbations (Handelsman, 2005). Among their discoveries in examining the gypsy moth “metagenome”—a representative collection of genomic clones derived from its gut microflora—is the presence of at least one gene associated with quorum sensing (also known as autoinduction), a bacterial system that monitors population density and coordinates gene expression with population growth (Bassler 1999; Dunn and Handelsman, 2002; Greenberg 1997; Handelsman, 2005; Hastings and Greenberg, 1999). The researchers are currently pursuing experiments to gauge the impact of this gene on gut community structure and robustness.
Microbial Disease in Plants and Animals
Research presented by Brian Staskawicz (see Chapter 2) suggests that microbial pathogens that colonize in animals share common strategies with those
that infect plants (Staskawicz et al., 2001). Both sorts of pathogens can deliver proteins into host cells that mimic, suppress, or modulate host defense signaling pathways and enhance pathogen fitness, and both are recognized by similarly sophisticated host surveillance systems. Striking architectural similarities between surface appendages of plant and animal pathogenic bacteria suggest common mechanisms of infection, while structural differences reflect the profound differences between plant and animal cells, most notably the presence or absence of a cell wall.
Plants lack the mobilized immune surveillance system and capacity for adaptive immunity present in animals. However, the form of innate immunity evident in plants, which responds to the presence of pathogen effector proteins, is in many ways comparable to innate mammalian mechanisms that recognize conserved molecular patterns on microbial surfaces (Staskawicz, 2005). Host surveillance proteins in plants, encoded by resistance (R) genes, are thought to mediate pathogen recognition by functioning as receptors for specific phytopathogen effector proteins (Baker et al., 1997). Interaction between these components triggers a rapid defensive reaction, known as the hypersensitive response, characterized by tissue death at the site of infection (Baker et al., 1997). This localized reaction limits the spread of infection and often precedes the development of nonspecific resistance throughout the plant, a phenomenon known as systemic acquired resistance. Striking structural similarities have been noted among R genes derived from several plant species that confer resistance to diverse bacterial, fungal, viral, and nematode pathogens. This suggests common patterns of defensive signaling among plants (Baker et al., 1997).
Conserved cellular defense responses in plants may also be analogous to certain innate immune responses to pathogens in vertebrates and insects, suggesting that these defense pathways are highly conserved and may be inherited from a common ancestor (Baker et al., 1997). Animals, plants, and yeast have been found to share structural (and in the case of plants and animals, functional) homology in a key enzyme (caspase) that regulates programmed cell death upon infection with a pathogen (Rojo et al., 2004). A prevalent protein class involved in plant disease resistance, the nucleotide-binding/leucine-rich repeat (NB/LRR) proteins, contains significant homology with Toll-like receptor (TLR) proteins associated with innate immunity in insects and mammals; more specifically, plant NB/LRR disease resistance proteins share homology with mammalian intracellular protein receptors NOD13 and NOD2, which function as intracellular receptors of bacterial peptidoglycan and which participate in the inflammatory cascade that causes Crohn’s disease (Staskawicz, 2005; Staskawicz et al., 2001). The prepon-
derance of conserved motifs and, presumably, mechanisms among plant and animal proteins involved with innate immunity has encouraged communication and even collaboration among the scientists who study these systems in widely different species—an unfortunately rare occurrence that may yield significant insights on the structure, function, and evolution of innate immunity.
THE ECOLOGY OF PATHOGENESIS
An ecological view of pathogenesis recognizes it as a strategy for microbial survival that reflects ongoing evolution between a microbe and its host (Falkow, 2005; Merrell and Falkow, 2004). Pathogenic microbes acquire genes that enable them to exploit their hosts through relationships that, under conditions of adaptation, tend toward persistence. Deadly disease may result if this delicate equilibrium is upset by genetic or physiological changes (in either the host or the microbe), or by ecological changes that result in infection of a host that is not part of a microbe’s established transmission cycle (e.g., Lyme disease in humans) (Falkow, 2005; IOM, 2003a). As contributor Stanley Falkow (see Chapter 3) and others—including Jared Diamond in his Pulitzer Prize-winning book Guns, Germs, and Steel (Diamond, 1999)—have observed, the advent of infectious disease adapted to humans is a relatively recent phenomenon, made possible by the existence of large host populations in close contact with one another and abetted by poor hygiene, malnutrition, and the opportunity for zoonotic transmission afforded by animal domestication (Falkow, 2005; McNeill, 1976).
Applying the ecological model to the distinction between commensal and pathogen, Falkow proposed that true pathogens—microbes that depend upon a pathogenic relationship with their hosts for survival—use their invasive properties, such as toxins and virulence factors, to establish a niche that is devoid of competition from other (noninvasive, and therefore nonpathogenic) microbes (Falkow, 2005). He compared the diverse and elaborate means by which true pathogens cross anatomical barriers and breach defenses in their hosts with the claws and fangs of eukaryotic predators. In addition to these obligate invaders, Falkow also ascribed significant human morbidity and mortality to both “accidental” pathogens, which can infect us but are not dependent upon our species for survival, and “opportunistic” microbes, which cause disease only in patients with immune system defects. He noted that while overt clinical disease frequently results from these encounters, it is not a necessary outcome of the host-pathogen interaction. Thus, he concluded, “the focus on clinical disease may distract us from understanding the actual mechanisms of the host-pathogen relationship.”
Pathogenesis in Context: The Dual Nature of Helicobacter pylori
The human stomach bacterium Helicobacter pylori is an exemplar of those microbes that confer both costs and benefits upon their hosts (Blaser, 1997, 2005).
While strongly associated with increased risk for peptic ulcer disease and gastric cancer, recent evidence suggests that H. pylori reduces the risk of esophageal diseases (e.g., severe gastric reflux and esophageal cancer), and possibly also of obesity and childhood diarrheal diseases (Blaser, 2005; Blaser and Atherton, 2004; Domínguez-Bello, 2005). In the model of this host-microbe interaction presented by Martin Blaser (see Chapter 3), the persistence of H. pylori in the stomach depends upon a precisely balanced equilibrium between bacterial effectors and host responses. Rather than causing acute harm, colonization gradually and subtly increases the host’s vulnerability to serious health risks (Blaser, 2005; Blaser and Atherton, 2004).
Considerable evidence shows that H. pylori has inhabited the human stomach for at least 60,000 years and that the course of coevolution between microbe and host has spanned millions of years (Blaser, 2005; Blaser and Atherton, 2004). Blaser described the basis for this prototypical mammalian-microbe partnership as an elaborate, bidirectional interplay of signals: microbes communicating with their host via metabolites and toxins, and hosts communicating with their microbes via metabolites and defense molecules. However, recent changes in human ecology (e.g., reduced transmission resulting from improved hygiene and smaller family size; decreased host advantage from colonization; increased use of antibiotics) have apparently led to a decline in prevalence of H. pylori among people in developed countries—a trend that suggests the possible eventual disappearance of the microbe from the human microbiota.
What consequences will result from this loss? Blaser presented a range of evidence suggesting that one might expect a decline in cases of gastric cancer and peptic ulcer, as well as an increase in cases of esophageal cancer—and, in fact, these opposing trends appear to be under way in populations in developed countries (Blaser, 2005). He also characterized the disappearance of H. pylori as an indicator of the probable fate of other members of the human microbiota, each of which occupies a distinct niche by exchanging signals with its host. Thus, in addition to monitoring the specific effects of the retreat of H. pylori from human populations, workshop participants echoed the conclusions of Blaser and Atherton (2004) in urging investigation of the general phenomenon of extinction among members of our microflora (Blaser, 2005). Such losses, which are likely to alter physiological signaling, may contribute to the increasing prevalence of diseases such as gastroesophageal reflux, obesity, diabetes, asthma, and various malignancies.
Further support for this notion was provided by presenter Maria Domínguez-Bello (see Chapter 3), whose studies of indigenous American Indians determined that the presence of multiple intestinal parasites was not correlated with reduced lean body mass in this population. Indeed, Domínguez-Bello found that the presence of H. pylori in the GI tracts of American Indian children was associated with higher lean body mass and better nutritional status. To understand the importance of microbiome diversity to human health, and therefore, the consequences of los-
ing that diversity, Dominguez-Bello argues that we must study host-microbe relationships in primitive societies as well as in industrialized populations.
Microbial and Host Factors in Disease
Additional workshop presentations and discussions considered intricate host-microbe interactions associated with a variety of diseases, most notably inflammatory bowel diseases (IBDs) such as Crohn’s disease and ulcerative colitis (Sartor, 2005). Such conditions highlight the inadequacy of the war metaphor, with its “us versus them” paradigm for pathogenesis. Workshop presenters also support Falkow’s observation that the innate immune system (which provides nonspecific protection against microbes, in contrast to the agent-specific defenses mounted by the adaptive immune system) has largely evolved “to keep the commensals in their place,” rather than to defend against primary pathogens (Falkow, 2005).
An apt description of the interplay between the innate immune system and commensal flora in the human gut appears in a recent review by MacDonald and Monteleone, who describe this relationship as “precarious,” given the mounting evidence that perturbations in this finely balanced ecosystem can lead to inflammation (MacDonald and Monteleone, 2005). When that balance is upset, the authors observe, “the commensal flora appears to act as a surrogate bacterial pathogen, and it is thought that lifelong inflammation ensues because the host response is incapable of eliminating the flora.” The cause of this imbalance remains to be determined, but a prevailing explanation implicates improvements in hygiene and readily available antibiotics in the developed world, where IBD and related inflammatory diseases, as well as allergies and asthma, are on the rise (MacDonald and Monteleone, 2005). These developments have drastically reduced human exposure to overt infection, and perhaps more importantly, to harmless gut microbes that somehow influence the maturation and regulation of host immunity.
Multiple factors apparently contribute to the host-microbe disequilibrium that underlies IBDs, according to Balfour Sartor (see Chapter 3). Genetic susceptibility predisposes some hosts to mount an overly hostile immune response to luminal commensal bacteria, but it also appears that the relative balance of injurious versus protective bacterial species is altered in affected individuals (Sartor, 2004, 2005.) Promising modes of therapy for IBD may therefore employ selective antibiotics to reduce the populations of injurious species, and conversely, introduce beneficial bacterial species (probiotics; see subsequent discussion and Chapter 6) and/or dietary supplements, known as prebiotics, that stimulate the growth or activity of beneficial species. However, Sartor observed, this promising picture is complicated by striking host and bacterial species specificity in the production of colitis. Since the definition of a “bad” bacterium depends upon the context, antibiotic, probiotic, or prebiotic therapeutic regimens for IBD need to be tailored to patients’ genetic profiles and microbiota.
THE HOST RESPONSE
Recent insights on the complex etiologies of IBDs and related inflammatory diseases reveal limitations in our ability, as host organisms, to appropriately react to the microbes we encounter. Thus, it is clear that a complete understanding of pathogenesis must consider not only how microorganisms inflict damage—the primary focus of research under the war metaphor—but also the mechanisms by which hosts discriminate among microbes and convert that information into an immune response. This expanded field of inquiry is yielding intriguing results, as described in workshop presentations and discussions on the following topics: the identification in mammalian epithelial cells of pattern recognition receptors (PRRs) and their previously mentioned counterparts in plants (MacDonald and Monteleone, 2005; Querishi and Medzhitov, 2003; Staskawicz, 2005; Staskawicz et al., 2001); the characterization of host-microbe interactions that establish mucosal immunity in the gut (Neutra, 2005; Neutra et al., 2001); and the analysis of patterns of global host gene expression in response to infection (Manger and Relman, 2000; Relman, 2005).
Host Recognition of Microbe-Associated Molecular Patterns
PRRs are a broad class of host molecules that activate innate immune responses upon encountering certain conserved molecules and structures4 common to pathogenic bacteria, viruses, and fungi (but not, apparently, to multicellular parasites) (Kopp and Medzhitov, 2003; Querishi and Medzhitov, 2003). Among the features recognized by PRRs are peptidoglycans and lipoproteins found in most bacteria, CpG sequence strings in DNA, lipopolysaccharides associated with gram-negative bacteria, teichoic acids associated with gram-positive bacteria, and double-stranded RNA replication intermediates of viruses (Didierlaurent et al., 2002). PRRs have been implicated in triggering a variety of innate immune responses, including the opsonization of foreign antigens and the induction of host complement, coagulation, phagocytic, and proinflammatory signaling cascades. Cytokines, interferons, chemokines, and other proinflammatory effectors induced as part of the innate immune response to microbial invasion in turn help to stimulate and shape the adaptive immune response (mediated primarily through B and T lymphocytes) to specific antigens on invading microbes.
PRRs include cytosolic proteins such as NOD2 and RIG-1, proteins such as TLRs expressed on the cell surface or within cytoplasmic vesicles, and extracel-
lular proteins such as C-reactive protein (Querishi and Medzhitov, 2003). Recent findings associate three major polymorphisms in the NOD2 protein of gut epithelial cells with increased risk for Crohn’s disease (see Sartor in Chapter 3) (Hugot et al., 2001; Kobayashi et al., 2005; Maeda et al., 2005; Ogura et al., 2001). Individuals bearing these mutant forms of NOD2 are impaired in their ability to clear bacteria from the intestinal mucosal surface or from infected epithelial cells.
Knowledge of the diverse microbial signatures recognized by PRRs has expanded rapidly, but recent findings indicate that there is much more to be learned about the various means by which hosts detect the microbes in their midst. For example, presenter David Relman raised the question of whether mammalian hosts recognize endogenous archaea, since these organisms do not display any of the known molecular patterns recognized by PRRs (Relman, 2005). As he and coworkers have recently discovered, methanogenic archaea play a central role in some cases of gingivitis by removing molecular hydrogen from their environment, thereby benefiting the bacteria that actually damage gum tissue. In other patients, microbes other than archaea have been found to serve as hydrogen scavengers; thus, the “pathogen” that causes gingivitis is not a single microbe, but a microbial community. It remains to be determined whether archaea are detected by their mammalian hosts, and if so, what sort of disease response their presence might arouse.
The Intestinal Epithelium: An Interactive Barrier
Epithelial cells raise daunting barriers to microbial invasion. Tight junctions, apical surface coats, the secretion of protective substances (e.g., mucins), and antimicrobial agents (e.g., defensins, cecropins, lactoferrin, and lysozyme) provide broad protection against microorganisms to which hosts have not been previously exposed (Boman, 2000; Corfield et al., 2000; Didierlaurrent et al., 2001; Frey et al., 1996; Madara et al., 1990). Much of the intestinal epithelium features a combination of these innate defenses, but they are reduced in cells that lie between lymphoid follicles and the intestinal lumen (Neutra, 2005). The follicle-associated epithelium serves as a key interchange in signaling between luminal microbes (both pathogens and commensals) on one side of the epithelial barrier, and host immune and inflammatory cells on the other (see Neutra in Chapter 4) (Kagnoff and Eckmann, 1997). These specialized sites permit selective sampling of the microbial contents of the lumen and delivery of microbes and antigens to the mucosa-associated lymphoid tissues. This occurs without provoking the sort of massive intestinal inflammation typified by IBDs, which harms both host and beneficial microbes.
Lymphoid follicles are not widely distributed along the path of the gut, but are collected in relatively rare locations known as immune induction sites (Neutra, 2005). In humans, immune induction sites such as Peyer’s patches are most numerous in the colon and rectum, and are also found in the oral cavities, tonsils,
and adenoids. According to workshop presenter Marian Neutra, this makes sense, as these locations represent areas of greatest exposure to microbes. Also, because sampling microbes exposes the host to the risk of infection, it is best confined to a fraction of the otherwise well-defended epithelium. The cells responsible for transporting pathogens and antigens from the lumen to the lymphoid tissue, known as epithelial M cells, are unique to immune induction sites (Neutra, 2005; Neutra et al., 2001). These immunological “gatekeepers” collaborate closely with memory (and probably naive) B and T cells, as well as dendritic cells.
Neutra described a variety of recent discoveries regarding the workings of M cells (see Chapter 4). Understanding the molecular means by which M cells fulfill their gatekeeping role could inform the design of mucosal vaccines to prevent infection by pathogens such as HIV and provide insights on the prevention and treatment of IBDs and associated immune disorders (Kozlowski and Neutra, 2003; Neutra, 2005). Similarly, knowing how certain pathogens exploit immune induction sites in order to gain entry to a protected niche within their hosts could lead to novel preventive strategies against a variety of microbial threats, including Salmonella, Shigella, poliovirus, and reoviruses (Neutra and Kraehenbuhl, 1994).
Variability in Host Responses to Pathogens
Using molecular techniques such as complete genome sequences, DNA microarrays, and advanced proteomics to monitor microbial and host cell gene expression, researchers have greatly expanded our understanding of the complexities of host-microbe interactions (Chang et al., 1994; Cummings and Relman, 2000; Dalwadi et al., 2001; Gao and Moore, 1996; Hemmer et al., 1999; Kroes et al., 1999; Relman 2002; Relman et al., 1990, 1992; Sutton et al., 2000). David Relman described a variety of studies he and coworkers have conducted using DNA microarrays to examine transcript abundance in white blood cells following exposure to known bacterial and viral pathogens (see Chapter 4). Their findings illustrate the range of host transcriptional response to infection, identify recurring patterns, and suggest sources of variability.
One factor that has not been found to produce significant differences in host gene expression is the distinction between pathogen- versus commensal-associated stimuli (Relman, 2005). A considerable literature of in vitro studies of cultured human cells exposed to various microbes and microbial components, as well as in vivo experiments conducted by Relman and coworkers in nonhuman primates infected with viral or bacterial pathogens (see Chapter 4) suggests that the host transcriptional response to such stimuli is to a large extent stereotypical. Several genetic loci involved in this response have been identified; many control common mechanisms by which hosts recognize and respond to microbes, such as genes encoding PRRs (e.g., TLRs, NOD) and elements of immune response systems (e.g., the NF-kappa-B regulon).
But if the host cannot distinguish between pathogenic and commensal mi-
crobes, what initiates the disease response? Relman suggests that hosts react when they detect microbial stimuli in a location, or at a time, or in a concentration different from the normal parameters of their microbiota. In this scenario, lifelong stimulation of the host by commensal microbes establishes a dynamic homeostasis which, when disturbed by the unusual behavior of pathogens, alerts the host to mount a disease response.
Detailed comparisons of host transcriptional responses to various pathogens over time are, however, revealing subtle transcriptional signatures upon the host response. Such variability—which is thought to result from pathogen-specific mechanisms that co-opt, subvert, or modify the stereotypical host response to microbial stimuli—could be exploited for diagnostic purposes. Some promising initial efforts toward this goal were described, including the use of host transcriptional patterns to identify infants with Kawasaki syndrome, which in its symptomatology resembles several other pediatric infectious diseases (Relman, 2005), and also to distinguish among patients with dengue, malaria, and typhoid (Falkow, 2005). More generally, workshop participants reflected on the promise of using the tools of molecular genetics (which, it was noted, will continue to improve with time) to better understand host-microbe relationships—both individually and in the context of microbial communities—by examining gene expression patterns in both hosts and microbes over time.
ADDRESSING COMPLEXITY IN MICROBIAL AND HOST COMMUNITIES
The task of placing host-microbe relationships within an ecological context was undertaken by workshop presentations on two major sources of complexity in host-microbe ecosystems: the biodiversity of endogenous microbial communities, and the networks of host-microbe relationships among pathogens that typically infect multiple hosts (which are in turn infected by multiple pathogens).
Microbial ecologist David Stahl (see Chapter 5) set the stage for this session by asserting that a better understanding of the natural history of microbes—their diversity, as well as their phylogenetic and ecological relationships to each other and to other living things—will improve our ability to develop questions and hypotheses that probe host-microbe relationships at the molecular level. Unlike other biological disciplines concerned with higher organisms (e.g., botany, herpetology, primatology), microbiology did not experience a comparable period of natural history, due in large part to the difficulty of culturing most microbes. With the advent of genomic technologies, however, researchers are no longer limited to surveying the mere fraction of microbial diversity represented by culturable organisms, and are beginning to discover, in Stahl’s words, our “planet of the microbes.”
Diversity and Dynamics in Microbial Ecosystems
The same technologies that enable researchers to recover molecular information directly from the environment also permit them to deduce phylogenetic relationships and assess the dynamics of complex microbial systems over time (Stahl, 2005). Stahl described how he and coworkers have employed these methods to investigate a variety of host-microbe systems, including microbial fermentation in the cow, the community structure of human oral microbiota and its relationship to local and systemic disease, and the relationship between gut microflora composition and fiber (cellulose) digestion in pigs.
The pig study was intended to shed light on the larger question of whether gut function in mammals—and especially humans—could be modified by altering the structure of the endogenous microbial community (Stahl, 2005). After initial sampling and characterization of the microflora via 16S ribosomal RNA fingerprinting in both experimental and control animals, the researchers increased the cellulose content in the experimental animals’ feed over an 8-week period. During this time, they sampled the microbial community in both control and experimental animals on a weekly basis. Initial analyses of these samples revealed a dramatic increase in one bacterial species—Prevotella—in the microbiota of pigs with the highest fiber content in their diet. A similar shift in community composition occurred across two separate groups of experimental animals, suggesting that the microbial response to this dietary change is reproducible.
Population Biology of Multiple Hosts and Multiple Pathogens
Veterinary biologist Mark Woolhouse (see Chapter 5) enlarged the ecological perspective beyond microbial ecosystems within individual hosts to address the relationships among all hosts colonized by the same microbial species. In particular, he addressed the web of relationships surrounding the transmission and survival of zoonotic pathogens in novel hosts. These organisms make up more than half of all known pathogens and, when compared with nonzoonotic pathogens, are twice as likely to be associated with emerging diseases (Cleaveland et al., 2001; Woolhouse, 2002, 2005).
The population biology approach to examining multihost-pathogen relationships taken by Woolhouse and colleagues has afforded several important insights regarding pathogen emergence and evolution. For example:
-
Pathogens “jump” between species quite rapidly—on an ecological, rather than evolutionary, time scale—and frequently jump into unrelated host species.
-
Single-stranded RNA viruses jump between hosts more frequently than other pathogenic microbes in part because they are more genetically unstable and prone to mutation, and therefore more adaptable.
-
Transmission efficiency for many zoonoses—including H5N1 influenza,
-
sleeping sickness, and verocytotoxigenic E. coli (VTEC)5—is such that a relatively small ecological change (in the host) or a genetic change (in the pathogen) could spark an epidemic.
-
Newly emerged pathogens are not subject to historical evolutionary constraints; therefore, the best evolutionary strategy for a pathogen population coexisting in two or more host species may be to become more virulent in one host species.
-
Constraints on susceptibility and pathogenicity imposed by coevolution may no longer hold when new host-pathogen associations emerge or ancient associations are disrupted, affecting both the magnitude and severity of disease outbreaks (Woolhouse and Dye, 2001; Woolhouse et al., 2002).
These findings suggest that coevolution between hosts and pathogens plays an important role in disease emergence and reemergence. Demonstrating this influence requires the integration of research on the population biology of microbes and hosts with molecular biological and genetic analyses of host-microbe interactions (Woolhouse and Dye, 2001; Woolhouse et al., 2002). Workshop participants noted that an important first step toward achieving such a synthesis would be to forge connections among diverse scientific disciplines. Collaborations among researchers in the medical, veterinary, plant science, and microbiology research communities, who currently study host-microbe interactions in relative isolation, represent a promising engine of advancement for this crucial field of scientific inquiry.
MANIPULATING HOST-MICROBE INTERACTIONS: PROBIOTIC RESEARCH AND REGULATION
The concept that ingesting certain microbes could prevent or even treat intestinal disease was introduced more than a century ago (Metchnikoff, 1901; Nowak-Wegryzn, 2005). However, despite far-reaching health claims associated with daily consumption of foods containing live bacterial cultures, the biological effects of such products have been demonstrated only under extremely limited
|
5 |
Verocytotoxigenic E. coli (VTEC) cause serious diarrheal disease. In 10 to 15 percent of cases, hemolytic uremic syndrome (which can result in kidney failure or even death) develops as a complication. The most frequent serotype isolated is O157, but other serotypes, such as O139, have been reported. VTEC is a common commensal organism in cattle, sheep, and other farm animals, The infectious dose is very low, about 10–100 organisms, and thus often causes waterborne outbreaks when animal feces-contaminated material gains access to water supplies post-treatment or where treatment is inadequate. Source: Lightfoot NF. 2003. Bacteria of potential health concern. In Bartram J, Cotruvo J, Exner M, Fricker C, Glasmacher A, eds. Heterotrophic Plate Counts and Drinking-Water Safety. London, UK: IWA Publishing. Available: www.who.int/entity/water_sanitation_health/dwq/HPC5.pdf. |
conditions. Indeed, as several workshop participants pointed out, even the general term for such ingested microbial products, probiotics, is subject to broad interpretation.
As defined by an expert panel convened in 2002 by the Food and Agriculture Organization (FAO) of the United Nations and the World Health Organization (WHO), probiotics are “live microorganisms administered in adequate amounts which confer a beneficial health effect on the host” (Joint FAO/WHO Working Group, 2002). Currently, the main sources of probiotics are dairy foods containing live cultures of Lactobacillus and/or Bifidobacterium; however, more expansive definitions of probiotics have included microbial components and nonviable or killed microorganisms (Vaillancourt, 2005). Prebiotics, a related concept, are generally considered to be nondigestible substances (e.g., oligosaccharides) added to the diet with the intent of selectively stimulating growth or activity of beneficial indigenous intestinal bacteria (ISAPP, 2005). Synbiotics, according to presenter Lorenzo Morelli, are defined as “mixtures of probiotics and prebiotics that beneficially affect the host by improving the survival and implantation of live microbial dietary supplements in the GI tract of the host” (Andersson et al., 2001).
The central question of what is meant by “beneficial health effects” in this context, and more importantly, how such overtly qualitative notions can be replaced by quantifiable variables, dominated workshop discussion on this topic. The most convincing evidence of probiotic efficacy concerns the treatment of acute infectious diarrhea and the prevention of antibiotic-associated diarrhea (Cremonini et al., 2002; D’Souza et al., 2002; Szajewska and Mrukowicz, 2001; Van Niel et al., 2002). Other conditions potentially treatable by microbes include chronic diarrhea (Xiao et al., 2003), IBD (Schultz et al., 2004), irritable bowel syndrome (Saggioro, 2004), food allergy (Majamaa and Isolauri, 1997). and cancer (Agrawal et al., 2004; Bettegowda et al., 2003; Dang et al., 2001, 2004; Peyromaure and Zerbib, 2004; Wiemann and Starnes, 1994). Conditions that could potentially be prevented with probiotics include travelers’ diarrhea (Hilton et al., 1997), necrotizing enterocolitis (Dani et al., 2002), urogenital infections (Cadieux et al., 2002), atopic diseases (e.g., asthma, atopic dermatitis, allergic rhinitis) (Kalliomaki et al., 2001), and dental caries (Nase et al., 2001). Some researchers have also envisioned the use of probiotics to prevent infectious complications of cystic fibrosis, rheumatoid arthritis, and cancer (Vanderhoof, 2001). However, very few double blind, placebo-controlled clinical trials have been conducted to monitor any of these potential effects.
In addition to exploring the potential of probiotic bacteria to promote a variety of specific beneficial changes in their human hosts and how such benefits might be measured, workshop participants considered the regulation of probiotic products. Current FAO/WHO guidelines for the evaluation of probiotics in food and relevant United States Food and Drug Administration (FDA) regulations were presented, along with several unresolved issues that need to be addressed in order to keep pace with this fast-moving and promising field.
Genomic Assessment of Probiotic Activity
Recently developed genomic technologies provide a promising route to evaluating the effects of probiotics on the host-microbe relationship, according to presenter Michiel Kleerebezem of the Wageningen Centre for Food Sciences in the Netherlands. He described the use of bioinformatics techniques to identify, in a single probiotic strain (WCFS1) of the bacterium Lactobacillus plantarum, genes that are transcriptionally activated upon introduction into the host, to monitor host gene expression following colonization with L. plantarum, and to reveal candidate genes associated with specific beneficial probiotic phenotypes (see Chapter 6).
Kleerebezem and coworkers discovered that of the 72 genes in L. plantarum WCFS1 they found were induced upon introduction to a mouse GI tract model, 30 had previously been identified as having been induced in pathogenic bacteria introduced into the same model. These commonly induced genes would, therefore, seem to be associated with microbial survival, rather than virulence—an idea that was further explored by introducing individual deletion mutants of L. plantarum WCFS1, each of which lacked a candidate “survival” gene, into the mouse model. These experiments identified a subset of genes essential to microbial persistence in the host, although their actual function remains unknown. Taking this strategy to a human model, the researchers collaborated with colleagues at Lund University in Sweden to examine the activity of a closely related strain, L. plantarum 299V, in the human colon. Despite limitations in the detection of bacterial gene expression in this host system, they found increased activity in several genes known to be associated with metabolic and biosynthetic pathways.
Kleerebezem and colleagues have also established in vitro and in vivo human models to monitor the interaction with probiotic bacteria from the host’s perspective, revealing distinct changes in transcriptional activity upon stimulation with the L. plantarum WCFS1 probiotic strain. The responding host genes were found to encode fatty acid metabolism and lipid transfer factors, as well as cytoskeletal factors that suggest an increased turnover of epithelial cells.
In a further attempt to find clues to identify specific probiotic functions at the molecular level, Kleerebezem’s group is using gene chips to compare L. plantarum strains. By correlating phenotypic and genotypic data, they have identified candidate genes associated with probiotic properties that include reduced risk of VTEC infection and protection against inflammatory disease. If these genotype-phenotype links are confirmed, it may be possible to engineer the overexpression of such protective loci in second-generation probiotic bacterial strains.
Immune Regulation by Probiotics: Evidence and Application
Do probiotic bacteria actually regulate the mucosal immune response? This central question was approached through a series of related queries posed by presenter Suzanne Cunningham-Rundles. She reviewed progress to date toward
providing answers, describing studies that support the following conclusions (see Chapter 6):
-
Probiotic microbes have short-term regulatory effects on immune response.
-
Specific immune mechanisms are selectively affected by commensal (probiotic) microbes.
-
Although host immune response to commensals is to a great extent stereotyped (as is the host response to pathogens, as previously discussed), there is enough specificity to permit manipulation of the immune response by probiotics.
-
Commensal bacteria influence the priming of the immune response, probably through the action of T regulatory cells.
-
Oral consumption of probiotic bacteria can have a therapeutic effect by improving gut flora richness and barrier function and by down-regulating inflammation.
Research in Cunningham-Rundles’ lab, concerning the possible role of commensal bacteria in the priming of the immune response, has led to a series of studies that explore the possibility that microbial colonization could be manipulated. The primary goal of this effort is to encourage colonization by commensal strains known to reduce inflammation, and its associated risks for neonatal development, in vulnerable preterm and other low birth-weight infants. In collaboration with the aforementioned Swedish developers of L. plantarum 299V, Cunningham-Rundles and coworkers have also conducted a trial in which HIV positive children with failure to thrive, a condition linked to chronic inflammation, were treated with juice fortified with the probiotic bacterium. Although the results were mixed, those children who gained height and weight also showed a dramatically improved immune response. Based on the results of this study, the group is preparing an investigational new drug (IND) application (see subsequent discussion of regulatory issues) for probiotic-fortified formula intended for use in low birth-weight infants.
Regulatory Approaches and Future Challenges
Both FAO/WHO joint guidelines on the evaluation of probiotics in food (see Morelli in Chapter 6) and U.S. FDA regulations (see Vaillancourt in Chapter 6) distinguish between microbes delivered as food and those that are used for a specific therapeutic purpose, such as to mitigate a pathological condition (Morelli, 2005; Vaillancourt, 2005). However, FDA regulations do not include or define the terms probiotic or prebiotic. The FDA’s Office of Vaccine Research and Review in the Center for Biologics Evaluation and Research (OVRR/CBER) uses the term live biotherapeutics to describe the probiotics it regulates; this category encompasses bacteria, yeast, or live virus used in prevention or treatment.
Because intended use determines how a probiotic is regulated, another FDA entity, the Center for Food Safety and Applied Nutrition (CFSAN), regulates probiotics and prebiotics marketed as dietary supplements or food ingredients.
Most prebiotics fit the FDA definition of a dietary supplement (“a product taken by mouth that contains a dietary ingredient intended to supplement the diet”), and to date, so do all probiotic products on the market. Although this situation is expected to change, workshop participants noted, there is little incentive for manufacturers of probiotics currently marketed as dietary supplements to develop them as biotherapeutics, given the rigors and expense of the associated review and regulation process. This situation confuses many consumers, who struggle to understand the vague health claims associated with probiotics and other dietary supplements, and who may (especially if they are ill) misinterpret such claims as proof of therapeutic efficacy. But unless serious adverse events can be shown to result from the use of a dietary supplement, the FDA cannot remove the product from the market. It was noted that a similarly confusing situation currently exists for European consumers, but that many countries in Europe are currently considering legislation to require proof for all health claims.
Although the dietary supplement/biotherapeutic dichotomy may remain a part of U.S. regulation of probiotics for some time, presenter Julienne Vaillancourt of OVRR/CBER expects the regulatory process for biotherapeutics to expand and change to reflect new knowledge. In fact, she identified several issues that need to be addressed in revisions to current regulations; these include the need to define and set guidelines for the evaluation of colonization and potency as they relate to biotherapeutics, and also to establish protocols for investigating the potential pathogenicity of probiotic strains. In addition, it was observed that although the most promising populations of beneficial microbes adhere to mucosal surfaces, most probiotics currently on the market have been isolated from stool samples that contain very few mucosal-adherent bacteria. Moreover, the vast majority of probiotic efficacy trials are conducted on the basis of the analysis of stool samples.
Thus, it is not only clear that guidelines and regulations governing probiotics must be revised to reflect recent research findings, but also that this goal is a fast-moving target. Lorenzo Morelli predicted the advent, within two to three years, of new products such as targeted probiotics or biotherapeutics that enhance production of specific cytokine or suppress specific pathogens, as well as new genotype-based methods of surveying microbial populations and assessing host-microbe interactions. By their very nature, such innovations will demand adjustments to current regulatory practices for probiotics.
PURSUING A NEW PARADIGM
With the development of genomic and bioinformatic tools, and with the expectation that the future will bring even more powerful technologies for resolving
the vast diversity of microbial communities, researchers can at last begin to study host-microbe relationships in their complexity. There is much to be discovered about the composition of microbial communities, how they assemble and self-regulate, and the means by which their members communicate with each other and with their hosts. Even in the relatively familiar and well-studied territory of the human gut, many basic questions remain unanswered. The following list of such queries, posed by David Relman and elaborated upon by several workshop participants, could equally be applied to a variety of other endogenous microbial communities and host-microbe ecosystems including, but not limited to:
-
How variable is the composition of the gut microbiota among human populations?
-
What is the role of timing in determining the acquisition and composition of the gut microbiota, and how does initial exposure and host genetics influence this process?
-
What drives the development of the immune response with respect to the temporal exposure to different pathogens?
-
How variable is the gut microbiota across space? Is the gut an assembly of microhabitats? Is it continuously variable?
-
What is the role of the individual host or host species in dictating the nature of the commensal microflora? Will this specificity permit the manipulation of either host or microbial community to benefit both?
-
How do microbial communities self-regulate?
-
What mechanisms enable endogenous communities to exchange information with their hosts, and vice versa?
-
How can the presence of phages, viruses, and archaea in the gut microbiota be characterized in terms of diversity and population sizes? What ecological roles do these organisms play in this and other microbial communities?
-
What is the role of polymicrobial interactions, biofilms, and other communities of indigenous microbes (e.g., skin)?
-
What do microbial community members do? What do pathogens do when they are not being pathogens (e.g., do their toxins have an ecological role)?
Interdisciplinary Research on Host-Microbe Interactions
Participants noted that the understanding of host-microbe relationships could be greatly advanced by the expansion and implementation of key recommendations of the IOM report, Microbial Threats to Health (2003a), that encourage an integrated and cooperative research effort by human and animal health communities on infectious disease threats. It was recognized that these goals would be furthered by engaging the plant research community and that the collaborative research agenda on infectious disease should incorporate host-microbe ecology. Interdisciplinary infectious disease centers such as those proposed in the Micro-
bial Threats to Health report could support research on such topics as the ecology of microbial communities across species and among multiple hosts and the response of microbial communities to novel ecological pressures and opportunities for host colonization. Much more needs to be learned about the roles of eukaryotic viruses in these processes—discussion of the possible roles played by DNA or RNA viruses were virtually absent in this workshop, reflecting a major gap in our understanding.
Connection among such centers on an international scale would further advance research goals by providing a “critical mass” of researchers to address the extreme complexity of scientific inquiry at the community and ecosystem level. A logical partner for international collaboration (as well as an example upon which to base U.S. programs) is the European network for research on the prevention and control of zoonoses, Med-Vet-Net (Med-Vet-Net, 2005). Founded in 2004, Med-Vet-Net comprises 300 scientists from 8 veterinary and 7 public health institutes, along with the Society for Applied Microbiology (UK), who are linked by a variety of structures intended to improve scientific collaboration and the dissemination of knowledge.
Opportunities for Global Survey of the Gut Microbiome
Workshop presentations and discussions clearly demonstrated both the feasibility and promise of conducting a microbial survey of human and animal microbiota. As noted by Bäckhed et al. (2005), “experimental and computational tools are now in hand to comprehensively characterize the nature of microbial diversity in the gut, the genomic features of its keystone members, the operating principles that underlie the nutrient foraging and sharing behaviors of these organisms, the mechanisms that ensure the adaptability and robustness of this systems, and the physiological benefits we accrue from this mutualistic relationship.” The technical feasibility of microbial genomic surveillance now makes it possible to conduct global surveys of gut microbiota and also to monitor how these microbial communities respond (in terms of structure and composition) to environmental change. Indeed, it was observed, the collection of this data could be viewed as an extension of the human genome project to encompass the “organismal metagenome.” In addition to advancing understanding of the etiology and epidemiology of infectious disease, this project may shed light on microbial influence on a host of chronic disorders, including GI conditions, allergy, asthma, diabetes, and obesity.
Participants also considered the collection, organization, and analysis of survey data on the gut microbiome. To assemble an encyclopedic representation, samples must be obtained from humans and animals across a broad range of geographic, nutritional, and health environments, as well as from several anatomical microenvironments. Standards for sampling methods would need to be established, and it was agreed that attempts should be made to identify and obtain
appropriate samples that may already exist in clinical and research communities—an effort that, in addition to reducing the time and expense of data collection, could also strengthen ties among potential collaborators in studies of host-microbe interactions. Finally, participants emphasized the importance of archiving sample material so that trends can be followed over time, and also to permit future analyses based on improved technologies.
RAISING AWARENESS OF THE HOST-MICROBE RELATIONSHIP
Our “war” on infectious microbes has restricted the spread of several pathogens and drastically reduced the burden of human disease, but the metaphor appears to be reaching the end of its usefulness. Recent findings on host-microbe interactions in a variety of settings, which highlight the many benefits of some microbes—as well as the potential for exploiting those benefits to further advantage—reveal the limitations of pure antagonism toward the microbes among us. At best, the war metaphor is a limiting mental shortcut that distracts from abundant opportunities to improve human and animal health. At worst, it represents a dangerous influence on disease control practices that have accelerated the development of antimicrobial resistance among human and animal pathogens, and perhaps also increased virulence in some pathogens. Put simply, the war metaphor must be replaced or, as comically (yet ominously) predicted in the epigraph to this summary, the bugs will win. We hosts are far better served by recognizing microbes as the allies they (mostly) are, and by making the best of our intimate alliances with them.
Such a message, which does not invoke the threat of catastrophe, will be difficult to send. The notion of microbes as the “enemy” will not fade quickly, especially given the relative complexity of the ecological perspective that would supplant the “us vs. them” paradigm. The most optimistic scenario for changing this opinion may be to begin within the infectious disease research community, where scientists who tend to focus on interactions between individual microbes and hosts could be encouraged to better understand and incorporate the concepts of community and ecosystem dynamics in their studies. A better-informed research community could then help to influence governmental and other funding agencies to recognize the importance of studying and funding proposals to examine host-microbe relationships to human health. Recognition of the commercial potential of probiotics could also encourage federal support for research, regulation, and the development of strain collections, reagents, and good manufacturing practices.
A similar “sea change” could occur if medical professionals encourage their patients to appreciate the benefits associated with the microbial flora and fauna that exist on and in us, and indeed to recognize that without these microbes, life as we know it would not exist. Many physicians are exercising new caution in prescribing antibiotics and some are able to explain their reasons for doing so to
their patients, but far more must be done—and said—to promote public understanding and support of the largely beneficial role that microbes play in their lives. If advances in the understanding of the specific actions of microbes in the development of immunity and protection from chronic disease can be translated into clinical practice, people may be able to declare a truce in the war on germs.
REFERENCES
Aderem A, Ulevitch RJ. 2000. Toll-like receptors in the induction of the innate immune response. Nature 406(6797):782–787.
Agrawal N, Bettegowda C, Cheong I, Geschwind JF, Drake CG, Hipkiss EL, Tatsumi M, Dang LH, Diaz LA Jr, Pomper M, Abusedera M, Wahl RL, Kinzer KW, Zhou S, Huso DL, Vogelstein B. 2004. Bacteriolytic therapy can generate a potent immune response against experimental tumors. Proceedings of the National Academy of Sciences 101(42):15172–15177.
Andersson H, Asp NG, Bruce A, Roos S, Wadstrom T, Wold AE. 2001. Health affects of probiotics and prebiotics: A literature review on human studies. Scandanavian Journal of Nutrition 45: 58–75.
Armstrong GL, Conn LA, Pinner RW. 1999. Trends in infectious disease mortality in the United States during the 20th century. Journal of the American Medical Association 281(1):61–66.
Bäckhed F, Ding H, Wang T, Hooper LV, Koh GY, Nagy A, Semenkovich CF, Gordon JI. 2004. The gut microbiota as an environmental factor that regulates fat storage. Proceedings of the National Academy of Sciences 101(44):15718-15723.
Bäckhed F, Ley RE, Sonnenburg JL, Peterson DA, Gordon JI. 2005. Host-bacterial mutualism in the human intestine. Science 307(5717):1915–1920.
Baker B, Zambryski P, Staskawicz B, Dinesh-Kumar SP. 1997. Signaling in plant-microbe interactions. Science 276(5313):726–733.
Bassler BL. 1999. How bacteria talk to each other: Regulation of gene expression by quorum sensing. Current Opinion in Microbiology 2(6):582–587.
Bettegowda C, Dang LH, Abrams R, Huso DL, Dillehay L, Cheong I, Agrawal N, Borzillary S, McCaffery JM, Watson EL, Lin KS, Bunz F, Baidoo K, Pomper MG, Kinzler KW, Vogelstein B, Zhou S. 2003. Overcoming the hypoxic barrier to radiation therapy with anaerobic bacteria. Proceedings of the National Academy of Sciences USA 100(25):15083–15088.
Blaser M. 1997. Ecology of Helicobacter pylori in the human stomach. Journal of Clinical Investigation 100(4):759–762.
Blaser MJ. 2005 (March 16). Session I: Host-Pathogen Interactions: Defining the Concepts of Pathogenicity, Virulence, Colonization, Commensalism, and Symbiosis. Presentation at the Forum on Microbial Threats Workshop Ending the War Metaphor: The Changing Agenda for Unraveling the Host-Microbe Relationship, Washington, D.C., Institute of Medicine, Forum on Microbial Threats.
Blaser MJ, Atherton JC. 2004. Helicobacter pylori persistence: Biology and disease. Journal of Clinical Investigation 113(3):321–333.
Boman HG. 2000. Innate immunity and the normal microflora. Immunology Review 173:5–16.
Breitbart M, Hewson I, Mahaffy JM, Nulton J, Salamon P, Rohwer F. 2003. Metagenomic analysis of an uncultured viral community from human feces. Journal of Bacteriology 185(20):6220–6223.
Broderick NA, Raffa KF, Goodman RM, Handelsman J. 2004. Census of the bacterial community of the gypsy moth larval midgut by using culturing and culture-independent methods. Applied Environmental Microbiology 70(1):293–300.
Cadieux P, Burton J, Gardiner G, Braunstein L, Bruce AW, Kang CY, Reid G. 2002. Lactobacillus strains and vaginal ecology. Journal of the American Medical Association 287(15):1940–1941.
Casadevall A, Pirofski LA. 1999. Host-pathogen interactions: Redefining the basic concepts of virulence and pathogenicity. Infection and Immunity 67(8):3703–3713.
Casadevell A, Pirofski LA. 2000. Host-pathogen interactions: Basic concepts of microbial commensalism, colonization, infection, and disease. Infection and Immunity 68(12):6511–6518.
Casadevell A, Pirofski LA. 2002. The meaning of microbial exposure, infection, colonization, and disease in clinical practice. Lancet Infectious Diseases 2(10):628–635.
Casadevell A, Pirofski LA. 2003. The damage-response framework of microbial pathogenesis. Nature Reviews Microbiology 1(1):17–24.
Chang Y, Cesarman E, Pessin MS, Lee F, Culpepper J, Knowles DM, Moore PS. 1994. Identification of herpesvirus-like DNA sequences in AIDS-associated Kaposi’s sarcoma. Science 266(5192): 1865–1869.
Cleaveland S, Laurenson MK, Taylor LH. 2001. Diseases of humans and their domestic mammals: Pathogen characteristics, host range and the risk of emergence. Philosophical Transactions of the Royal Society of London Series B 365(1411):991–999.
Corfield AP, Myerscough N, Longman R, Sylvester P, Arul S, Pignatelli M. 2000. Mucins and mucosal protection in the gastrointestinal tract: New prospects for mucins in the pathology of gastrointestinal disease. Gut 47(4):589–594.
Cremonini F, Di Caro S, Nista EC, Bartolozzi F, Capelli G, Gasbarrini G, Gasbarrini A. 2002. Meta-analysis: The effect of probiotic administration on antibiotic-associated diarrhoea. Alimentary Pharmacology and Therapeutics 16(8):1461–1467.
Cummings CA, Relman DA. 2000. Using DNA microarrays to study host-microbe interactions. Emerging Infectious Diseases 6(5):513–525.
Dalwadi H, Wei B, Kronenberg M, Sutton CL, Braun J. 2001. The Crohn’s disease-associated bacterial protein I2 is a novel enteric T cell superantigen. Immunity 15(1):149–158.
Dang LH, Bettegowda C, Huso DL, Kinzler KW, Vogelstein B. 2001. Combination bacteriolytic therapy for the treatment of experimental tumors. Proceedings of the National Academy of Sciences USA 98(26):15155–15160.
Dang LH, Bettegowda C, Agrawal N, Cheong I, Huso DL, Frost P, Loganzo F, Greenberger L, Barkoczy J, Pettit GR, Smith AB 3rd, Gurulingappa H, Khan S, Parmigiani G, Kinzler KW, Zhou S, Vogelstein B. 2004. Targeting vascular and avascular compartments of tumors with C. novyi-NT and anti-microtubule agents. Cancer Biology and Therapy 3(3):326–337.
Dani C, Biadaioli R, Bertini G, Martelli E, Rubaltelli FF. 2002. Probiotics feeding in prevention of urinary tract infection, bacterial sepsis and necrotizing enterocolitis in preterm infants. A prospective double-blind study. Biology of the Neonate 82(2):103–108.
Diamond, J. 1999. Guns, Germs, and Steel: The Fates of Human Societies. New York: W.W. Norton & Company.
Didierlaurent A, Sirard JC, Kraehenbuhl JP, Neutra MR. 2001. How the gut senses its content. Cellular Microbiology 4(2):61–72.
Didierlaurent A, Sirard JC, Kraehenbuhl JP, Neutra MR. 2002 (February). How the gut senses its content. Cellular Microbiology 4(2):61–72.
Dominguez-Bello. 2005 (March 16). Session I: Host-Pathogen Interactions: Defining the Concepts of Pathogenicity, Virulence, Colonization, Commensalism, and Symbiosis. Presentation at the Forum on Microbial Threats Workshop Ending the War Metaphor: The Changing Agenda for Unraveling the Host-Microbe Relationship, Washington, D.C., Institute of Medicine, Forum on Microbial Threats.
D’Souza AL, Rajkumar C, Cooke J, Bulpitt CJ. 2002. Probiotics in prevention of antibiotic associated diarrhoea: Meta-analysis. British Medical Journal 324(7350):1361.
Dunn AK, Handelsman J. 2002. Toward an understanding of microbial communities through analysis of communication networks. Antonie Van Leeuwenhoek 81(1–4):565–574.
Fagarasan S, Muramatsu M, Suzuki K, Nagaoka H, Hiai H, Honjo T. 2002. Critical roles of activation-induced cytidine deaminase in the homeostasis of gut flora. Science 298(5597):1424–1427.
Falkow S. 2005 (March 17). Session III: Understanding the Dynamic Relationships of Host-Microbe Interactions. Presentation at the Forum on Microbial Threats Workshop Ending the War Metaphor: The Changing for Unraveling the Host-Microbe Relationship, Washington, D.C., Institute of Medicine, Forum on Microbial Threats.
Frey A, Giannasca KT, Weltzin R, Giannasca PJ, Reggio H, Lencer WI, Neutra MR. 1996. Role of the glycocalyx in regulating access of microparticles to apical plasma membranes of intestinal epithelial cells—implications for microbial attachment and oral vaccine targeting. Journal of Experimental Medicine 184(3):1045–1059.
Gao SJ, Moore PS. 1996. Molecular approaches to the identification of unculturable infectious agents. Emerging Infectious Diseases 2(3):159–167.
Gilbert GS, Handelsman J, Parke JL. 1994. Root camouflage and disease control. Phytopathology 84(3):222–225.
Gordon J. 2005 (March 16). Session I: Host-Pathogen Interactions: Defining the Concepts of Pathogenicity, Virulence, Colonization, Commensalism, and Symbiosis. Presentation at the Forum on Microbial Threats Workshop Ending the War Metaphor: The Changing Agenda for Unraveling the Host-Microbe Relationship, Washington, D.C., Institute of Medicine, Forum on Microbial Threats.
Greenberg EP. 1997. Quorum sensing in gram-negative bacteria: Cell density-dependent gene expression controls luminescence in marine bacteria and virulence in several pathogens. American Society of Microbiology News 63:371–377.
Handelsman J. 2005 (March 16). Session I: Host-Pathogen Interactions: Defining the Concepts of Pathogenicity, Virulence, Colonization, Commensalism, and Symbiosis. Presentation at the Forum on Microbial Threats Workshop Ending the War Metaphor: The Changing Agenda for Unraveling the Host-Microbe Relationship, Washington, D.C., Institute of Medicine, Forum on Microbial Threats.
Hastings JW, Greenberg EP. 1999. Quorum sensing: The explanation of a curious phenomenon reveals a common characteristic of bacteria. Journal of Bacteriology 181(9):2667–2668.
Hemmer B, Gran B, Zhao Y, Marques A, Pascal J, Tzou A, Kondo T, Cortese I, Bielekova B, Straus SE, McFarland HF, Houghten R, Simon R, Pinilla C, Martin R. 1999. Identification of candidate T-cell epitopes and molecular mimics in chronic Lyme disease. Nature Medicine 5(12):1375–1382.
Hilton E, Kolawowski P, Singer C, Smith M. 1997. Efficacy of Lactobacillus GG as a diarrheal preventive in travelers. Journal of Travel Medicine 4:41–43.
Hooper LV, Wong MH, Thelin A, Hansson L, Falk PG, Gordon JI. 2001. Molecular analysis of commensal host-microbial relationships in the intestine. Science 291(5505):881–884.
Hooper LV, Midtvedt T, Gordon JI. 2002. How host-microbial interactions shape the nutrient environment of the mammalian intestine. Annual Review of Nutrition 22:283-307.
Hooper TL, Hopkinson DN, Bhabra MS. 1998. Pulmonary graft preservation: A worldwide survey of current clinical practice. Journal of Heart and Lung Transplantation 17(5):525–531.
Hugot JP, Chamaillard M, Zouali H, Lesage S, Cezard JP, Belaiche J, Almer S, Tysk C, O’Morain CA, Gassull M, Binder V, Finkel Y, Cortot A, Modigliani R, Laurent-Puig P, Gower-Rousseau C, Macry J, Colombel JF, Sahbatou M, Thomas G. 2001. Association of NOD2 leucine-rich repeat variants with susceptibility to Crohn’s disease. Nature 411(6837):599–603.
Hylemon PB, Harder J. 1998. Biotransformation of monoterpenes, bile acids, and other isoprenoids in anaerobic ecosystems. FEMS Microbiology Reviews 22(5):475–488.
IOM (Institute of Medicine). 2003a. Microbial Threats to Health: Emergence, Detection, and Response. Washington, DC: The National Academies Press.
IOM. 2003b. The Resistance Phenomenon in Microbes and Infectious Disease Vectors. Washington, DC: The National Academies Press.
IOM. 2004. The Infectious Etiology of Chronic Diseases. Washington, DC: The National Academies Press.
ISAPP (International Scientific Association for Probiotics and Prebiotics). 2005. About ISAPP. [Online]. Available: http://www.isapp.net/IS_about.htm [accessed on December 19, 2005].
Isenberg HD. 1988. Pathogenicity and virulence: Another view. Clinical Microbiology Reviews 1(1):40–53.
Joint FAO/WHO Working Group. 2002. Guidelines for the evaluation of probiotics in food. [Online]. Available: http://www.fao.org/es/ESN/food/foodandfood_probio_en.stm [accessed December 19, 2005].
Kagnoff MF, Eckmann L. 1997. Epithelial cells as sensors for microbial infection. Journal of Clinical Investigation 100(1):6–10.
Kalliomaki M, Salminen S, Arvilommi H, Kero P, Koskinen P, Isolauri E. 2001. Probiotics in primary prevention of atopic disease: A randomized placebo-controlled trial. Lancet 357(9262):1076–1079.
Kindhauser MK. 2003. Communicable Diseases 2002: Global Defense Against the Infectious Disease Threat. Geneva, Switzerland: WHO.
Kobayashi KS, Chamaillard M, Ogura Y, Henegariu O, Inohara N, Nunez G, Flavell RA. 2005. Nod2-dependent regulation of innate and adaptive immunity in the intestinal tract. Science 307(5710):731–734.
Kopp E, Medzhitov R. 2003. Recognition of microbial infection by Toll-like receptors. Current Opinion in Immunology 15(4):396–401.
Kozlowski PA, Neutra MR. 2003. The role of mucosal immunity in prevention of HIV transmission. Current Molecular Medicine 3(3):217–228.
Kroes I, Lepp PW, Relman DA. 1999. Bacterial diversity within the human subgingival crevice. Proceedings of the National Academy of Sciences USA 96(25):14547–14552.
Lederberg J. 2000. Infectious history. Science 288(5464):287–293.
MacDonald TT, Monteleone G. 2005. Immunity, inflammation and allergy in the gut. Science 307(5717):1920–1925.
Madara JL, Nash S, Moore R, Atisook K. 1990. Structure and function of the intestinal epithelial barrier in health and disease. Monographs in Pathology 31:306–324.
Maeda S, Hsu LC, Liu H, Bankston LA, Iimura M, Kagnoff MF, Eckmann L, Karin M. 2005. Nod2 mutation in Crohn’s disease potentiates NF-kappaB activity and IL-1beta processing. Science 307(5710):734–738.
Majamaa H, Isolauri E. 1997. Probiotics: A novel approach in the management of food allergy. Journal of Allergy and Clinical Immunology 99(2):179–185.
Manger ID, Relman DA. 2000. How the host “sees” pathogens: Global gene expression responses to infection. Current Opinion in Immunology 12(2):215–218.
McNeill WH. 1976. Plagues and Peoples. New York: Anchor Books. P. 369.
Med-Vet-Net. 2005. MedVetNet Homepage. [Online]. Available: http://www.medvetnet.org/cms/ [accessed December 19, 2005].
Merell DS, Falkow S. 2004. Frontal and stealth attack strategies in microbial pathogenesis. Nature 430(6996):250–256.
Metchnikoff E. 1901. Sur la flore du corps humain. Manchester Literary and Philosophical Society 45:1–38.
Morelli L. 2005 (March 17). Session IV: Novel Approaches for Mitigating the Development of Resistance. Presentation at the Forum on Microbial Threats Workshop Ending the War Metaphor: The Changing Agenda for Unraveling the Host-Microbe Relationship, Washington, D.C., Institute of Medicine, Forum on Microbial Threats.
Nase L, Hatakka K, Savilahti E, Saxelin M, Ponka A, Poussa T, Korpela R, Meurman JH. 2001. Effect of long-term consumption of a probiotic bacterium, Lactobacillus rhamnosus GG, in milk on dental caries and caries risk in children. Caries Research 35(6):412–420.
Neutra MR. 2005 (March 17). Session II: Ecology of Host-Microbe Interactions. Presentation at the Forum on Microbial Threats Workshop Ending the War Metaphor: The Changing Agenda for Unraveling the Host-Microbe Relationship, Washington, D.C., Institute of Medicine, Forum on Microbial Threats.
Neutra MR, Kraehenbuhl JP. 1994. Cellular and molecular basis for antigen transport in the intestinal epithelium. In: Ogra PL, Mestecky J, Lamm ME, Strober W, McGhee JR, Bienstock J, eds. Handbook of Mucosal Immunology. Boston, MA: Academic Press, Inc. Pp. 27–39.
Neutra MR, Mantis NJ, Kraehenbuhl JP. 2001. Collaboration of epithelial cells with organized mucosal lymphoid tissues. Nature Immunology 2(11):1004–1009.
Nowak-Wegryzn A. 2005. Future approaches to food allergy. Pediatrics 111(6):1672–1680.
Ogura Y, Bonen DK, Inohara N, Nicolae DL, Chen FF, Ramos R, Britton H, Moran T, Karaliuskas R, Duerr RH, Achkar JP, Brant SR, Bayless TM, Kirschner BS, Hanauer SB, Nunez G, Cho JH. 2001. A frameshift mutation in NOD2 associated with susceptibility to Crohn’s disease. Nature 411(6837):603–606.
Peyromaure M, Zerbib M. 2004. T1G3 transitional cell carcinoma of the bladder: Recurrence, progression and survival. BJU International 93(1):60–63.
Querishi ST, Medzhitov R. 2003. Toll-like receptors and microbial infection. Genes and Immunity 4(2):87–94.
Rawls JF, Samuel BS, Gordon JI. 2004. Gnotobiotic zebrafish reveal evolutionarily conserved responses to the gut microbiota. Proceedings of the National Academy of Sciences USA 101(13): 4596–4601.
Relman DA. 2002. New technologies, human-microbe interactions, and the search for previously unrecognized pathogens. Journal of Infectious Diseases 186(Suppl 2):S254–S258.
Relman DA. 2005 (March 17). Session II: Ecology of Host-Microbe Interactions. Presentation at the Forum on Microbial Threats Workshop Ending the War Metaphor: The Changing Agenda for Unraveling the Host-Microbe Relationship, Washington, D.C., Institute of Medicine, Forum on Microbial Threats.
Relman DA, Loutit JS, Schmidt TM, Falkow S, Tompkins LS. 1990. The agent of bacillary angiomatosis. An approach to the identification of uncultured pathogens. New England Journal of Medicine 323(23):1573–1580.
Relman DA, Schmidt TM, MacDermott RP, Falkow S. 1992. Identification of the uncultured bacillus of Whipple’s disease. New England Journal of Medicine 327(5):293–301.
Rojo E, Martin R, Carter C, Zouhar J, Pan S, Plotnikova J, Jin H, Paneque M, Sanchez-Serrano JJ, Baker B, Ausubel FM, Raikhel NV. 2004. VPE-gamma exhibits a caspase-like activity that contributes to defense against pathogens. Current Biology 14(21):1897–1906.
Saggioro A. 2004. Probiotics in the treatment of irritable bowel syndrome. Journal of Clinical Gastroenterolgy 38(Suppl 6):S104–S106.
Salyers A. 2005 (March 16). Session I: Host-Pathogen Interactions: Defining the Concepts of Pathogenicity, Virulence, Colonization, Commensalism, and Symbiosis. Presentation at the Forum on Microbial Threats Workshop Ending the War Metaphor: The Changing Agenda for Unraveling the Host-Microbe Relationship, Washington, D.C., Institute of Medicine, Forum on Microbial Threats.
Sartor RB. 2004. Therapeutic manipulation of the enteric microflora in inflammatory bowel diseases: Antibiotics, probiotics, and prebiotics. Gastroenterology 126(6):1620–1633.
Sartor RB. 2005 (March 16). Session I: Host-Pathogen Interactions: Defining the Concepts of Pathogenicity, Virulence, Colonization, Commensalism, and Symbiosis. Presentation at the Forum on Microbial Threats Workshop Ending the War Metaphor: The Changing Agenda for Unraveling the Host-Microbe Relationship, Washington, D.C., Institute of Medicine, Forum on Microbial Threats.
Savage DC. 1977. Microbial ecology of the gastrointestinal tract. Annual Reviews in Microbiology 31:107–133.
Schultz M, Timmer A, Herfarth HH, Sartor RB, Vanderhoof JA, Rath HC. 2004. Lactobacillus GG in inducing and maintaining remission of Crohn’s disease. BMC Gastroenterology 4:5.
Smoot LM, Smoot JC, Smidt H, Noble PA, Konneke M, McMurry ZA, Stahl DA. 2005. DNA microarrays as salivary diagnostic tools for characterizing the oral cavity’s microbial community. Advances in Dental Research 18(1):6–11.
Stahl D. 2005 (March 17). Session II: Ecology of Host-Microbe Interactions. Presentation at the Forum on Microbial Threats Workshop Ending the War Metaphor: The Changing Agenda for Unraveling the Host-Microbe Relationship, Washington, D.C., Institute of Medicine, Forum on Microbial Threats.
Staskawicz BJ. 2005 (March 16). Session I: Host-Pathogen Interactions: Defining the Concepts of Pathogenicity, Virulence, Colonization, Commensalism, and Symbiosis. Presentation at the Forum on Microbial Threats Workshop Ending the War Metaphor: The Changing Agenda for Unraveling the Host-Microbe Relationship, Washington, D.C., Institute of Medicine, Forum on Microbial Threats.
Staskawicz BJ, Mudgett MB, Dangl JL, Galan JE. 2001. Common and contrasting themes of plant and animal diseases. Science 292(22):2285–2289.
Stewart WH. 1967. “A Mandate for State Action,” presented at the Association of State and Territorial Health Officers, Washington, D.C., Dec. 4, 1967. Taken from: Garrett L. 1994. The Coming Plague: Newly Emerging Diseases in a World Out of Balance. New York: Penguin Books. P. 33, footnote 9.
Sutton CL, Kim J, Yamane A, Dalwadi H, Wei B, Landers C, Targan SR, Braun J. 2000. Identification of a novel bacterial sequence associated with Crohn’s disease. Gastroenterology 119(1): 23–31.
Szajewska H, Mrukowicz JZ. 2001. Probiotics in the treatment and prevention of acute infectious diarrhea in infants and children: A systematic review of published randomized, double-blinded, placebo-controlled trials. Journal of Pediatric Gastroenterology and Nutrition 33(Suppl 2): S17–S25.
Vaillancourt R. 2005 (March 17). Session IV: Novel Approaches for Mitigating the Development of Resistance. Presentation at the Forum on Microbial Threats Workshop Ending the War Metaphor: The Changing Agenda for Unraveling the Host-Microbe Relationship, Washington, D.C., Institute of Medicine, Forum on Microbial Threats.
Van Niel CW, Feudtner C, Garrison MM, Christakis DA. 2002. Lactobacillus therapy for acute infectious diarrhea in children: A meta-analysis. Pediatrics 109(4):678–684.
Vanderhoof JA. 2001. Probiotics: Future directions. American Journal of Clinical Nutrition 73(6): 1152S–1155S.
Whittle G, Shoemaker NB, Salyers AA. 2002. The role of Bacteroides conjugative transposons in the dissemination of antibiotic resistance genes. Cellular and Molecular Life Sciences 59(12):2044–2054.
WHO (World Health Organization). 2000. Factors contributing to resistance. In: World Health Organization Report on Infectious Diseases 2000: Overcoming Antimicrobial Resistance. [Online]. Available: http://www.who.int/infectious-disease-report/2000/ [accessed January 6, 2005].
Wiemann B, Starnes CO. 1994. Coley’s toxins, tumor necrosis factor and cancer research: A historical perspective. Pharmacology and Therapeutics 64(3):529–564.
Wilson B, Salyers AA. 2003. Is the evolution of bacterial pathogens an out-of-body experience? Trends in Microbiology 11(8):347–350.
Winter C, Smit A, Herndl GJ, Weinbauer MG. 2004. Impact of virioplankton on archaeal and bacterial community richness as assessed in seawater batch cultures. Applied Environmental Microbiology 70(2):804–813.
Woolhouse ME. 2002. Population biology of emerging and re-emerging pathogens. Trends in Microbiology 10(Suppl 10):S3–S7.
Woolhouse M. 2005 (March 17). Session II: Ecology of Host-Microbe Interactions. Presentation at the Forum on Microbial Threats Workshop Ending the War Metaphor: The Changing Agenda for Unraveling the Host-Microbe Relationship, Washington, D.C., Institute of Medicine, Forum on Microbial Threats.
Woolhouse M, Dye C. 2001. Population biology of emerging and re-emerging pathogens. Philosophical Transactions of the Royal Societies of London Series B Biological Sciences 356:979–1106.
Woolhouse M, Webster JP, Domingo E, Charlesworth B, Levin B. 2002. Biological and biomedical implications of the co-evolution of pathogens and their hosts. Nature Genetics 32(4):569–577.
Xiao SD, Zhang de Z, Liu HY, Wang GS, Xu GM, Zhang ZB, Lin GJ, Wang GL. 2003. Multicenter, randomized, controlled trial of heat-killed Lactobacillus acidophilus LB in patients with chronic diarrhea. Advances in Therapy 20(5):253–260.
Xu J, Gordon JI. 2003. Inaugural article: Honor thy symbionts. Proceedings of the National Academy of Sciences USA 100(18):10452–10459.


































