
1
Role and Importance of Pollinators
The angiosperms—flowering plants that produce seeds often enclosed within an edible fruit—are among the planet’s most successful life forms. More than 250,000 species of flowering plants have been described, and an equivalent number could await discovery (Davies et al., 2004). At the base of most terrestrial and many aquatic food webs, the angiosperms are the principal providers of nutrients and resources to most other organisms, and they provide multidimensional structure for the majority of terrestrial and freshwater ecosystems. Humans derive food, fiber, drugs, and fuel, either directly or indirectly from angiosperms. Moreover, angiosperms have enhanced aesthetic, recreational, and cultural pursuits since before recorded history.
Reproductive systems of angiosperms vary greatly among species (Box 1-1), but two processes are necessary for sexual reproduction in all angiosperms: pollination—the transfer of pollen from the anthers of a stamen to the stigma of a pistil—and fertilization—the fusion of the sperm nuclei from pollen with the egg nucleus in the ovary to produce an embryo. Some plants self-pollinate, that is, pollen transfer occurs within the same flower or among the flowers on a single plant, usually because the anthers touch the adjacent stigma. The majority of flowering plants, however, depend on the transfer of pollen from other individuals (cross-pollination).
Although some species rely on abiotic forces, including wind and water, for pollen transfer, more than three-fourths of the planet’s angiosperms rely on over 200,000 species of animal pollinators to various extents to meet their reproductive needs. Fossil records show that angiosperms underwent a remarkable diversification between 130 million and 90 million years ago
|
BOX 1-1 Barriers to Self-Fertilization Flowers can be staminate (bearing only male reproductive organs), pistillate (bearing only female reproductive organs), or perfect (bearing male and female reproductive organs). Individual plants can be monoecious (bearing staminate and pistillate flowers), dioecious (staminate and pistillate flowers borne on separate plants), or even trioecious (staminate, pistillate, and perfect flowers borne on separate plants). Within dioecy, various conditions can be found in different species; gynodioecy, for example, is the term applied to the breeding system of species in which individuals bear either female or hermaphrodite flowers (Richards, 1997). Almost three-quarters of all plant species produce perfect flowers. Approximately 5 percent are dioecious, and slightly more than 5 percent are monoecious (Molnar, 2004). Pollination can occur within the flowers of a single plant, among different flowers of a single plant, and among flowers of different plants. A plant that is self-fertile and self-pollinating is called autogamous if pollination and fertilization take place within the same flower. A plant is geitonogamous if pollination and fertilization take place between flowers of the same plant, whereas a plant that is cross-pollinated and cross-fertilized is xenogamous. It is common for plants to receive mixtures of self and outcross (nonself) pollen grains, especially if the male and female parts are in the same flower (Plate 1—a perfect or hermaphrodite flower). Perpetual self-fertilization could be problematic for plants because of the many potential genetic complications associated with inbreeding (Charlesworth and Charlesworth, 1987). Accordingly, adaptations that reduce the likelihood of selfing exist in many taxa. Dioecy and monoecy promote outcrossing, |
and that they achieved ecological dominance 100 million to 70 million years ago (Davies et al., 2004). Chief among the many explanations offered for their spectacular ascendancy is the development of mutualistic associations with animals for the dispersal of pollen (Baker and Hurd, 1968; Faegri and van der Pijl, 1979; Labandeira et al., 1994; Stebbins, 1950, 1974) and seeds (Herrera, 1989; Kevan, 1984; van der Pijl, 1982). Mutualistic associations with animals provide mobility of gametes to otherwise predominantly sessile terrestrial plants, which allows for greater genetic variation in plants as well as access to a wider range of ecological opportunities through seed dispersal. For flowering plants, use of an animal partner to transport pollen increases the area in which potential mates can be found and promotes outcrossing, the merger of gametes from genetically distinct individuals. Increasing genetic variability through recombination associated with outcrossing is key
|
although monoecious plants can receive self-pollen from male flowers on the same plant. Many monoecious species produce male and female flowers at different times, and the probability of selfing is reduced. Similarly, in plants with hermaphrodite flowers, self-pollination within flowers is avoided when the male and female floral parts mature at different times. In some species, the chance of self-pollination is reduced because the male and female parts of the same flowers are separated. In a subset of those species, the male and female parts of the flower move closer together as the flower ages, allowing self-pollination as a “last resort” before the flower is too old to set fruit. As a further deterrent to selfing, many flowering plant species are self-incompatible—that is, pollen that is deposited on a stigma within the same flower (or another flower on the same plant) is unable to achieve fertilization. Self-incompatibility is controlled in complex and variable ways, and it involves the interplay of incompatibility alleles (of which there may be many) and their effects in the two parent plants (Matton et al., 1994). The effectiveness of self-incompatibility mechanisms ranges from absolute to weak, and the mechanisms for blocking self-fertilization can break down as a result of aging or external factors, especially heat. Breaking those barriers down ensures sexual reproduction (seed set and fruit set) even when cross-pollination is not possible. It is important to note, however, that despite the ubiquity of outbreeding, many species persist exclusively and successfully with self-pollinating and self-fertile flowers. Moreover, some self-fertile plants that can self-pollinate (including some legumes) are of agricultural importance. They can establish themselves in nonindigenous areas where their natural pollinators are absent. The nature and evolutionary biology of plant-breeding systems are presented in detail by Richards (1997). |
to allowing organisms to adapt to spatially and temporally variable environments (Box 1-1). Genetic variability in plant populations could help to facilitate the evolution of resistance to pathogens and herbivores.
After fertilization is complete, the production of fruit ensues. A flower’s ovary may contain a single ovule and produce a fruit that bears only a single seed (as in the almond, avocado, coconut, plum, or cherry), or it may contain hundreds of ovules and produce a fruit bearing hundreds of seeds (as in the tomato, kiwi fruit, cucumber, watermelon, or squash). Because each seed results from the union of a sperm cell from a pollen grain and an egg cell, some plants require many hundreds of pollen grains to fertilize all of the available egg cells. If a flower receives an inadequate number of pollen grains, some of the egg cells will not be fertilized and accordingly seeds will not develop. Economic consequences of such incomplete fertilization include
production of undersized or misshapen fruit that, for a market crop, has less value. Adequate pollination often requires that individual flowers be visited by many pollinators or that one to several pollinators make multiple trips to the same flower.
Some fruits of economic importance are seedless by design. They are generally the product of selective breeding or genetic manipulation that would not be sustainable in nature (Schery, 1972). Seedless bananas, for example, are the products of sterile triploid plants arising either spontaneously or as a result of hybridization of diploid and tetraploid individuals and are propagated vegetatively. Parthenocarpic fruits, such as clementines (seedless tangerines), are those in which fruits develop in the absence of successful fertilization; fertilization could fail because these self-incompatible cultivars are grown in monoculture orchards. Seedless grapes, in contrast, are stenospermocarpic; fertilization takes place, but the resulting fruit is seedless because the immature embryo fails to develop (Schery, 1972).
POLLINATORS IN NATURAL AND AGRICULTURAL ECOSYSTEMS
In view of the fact that pollinator-plant interactions encompass almost 400,000 species, the precise nature of the relationship between plant and pollinator varies enormously. Although some animals visit flowers for nectar or pollen, not all flower visitors bring about pollination. Effective pollinators often have behavioral and anatomical traits that greatly increase the efficiency and accuracy of pollen delivery (Barth, 1991; Faegri and van der Pijl, 1979; Proctor et al., 1996). In general, pollination is a mutually beneficial interaction; pollinating animals receive some form of nutritional “reward” for visitation and pollen delivery. Pollen itself can be a reward, serving as the primary food resource for most larval bees and as an important source of protein for some flies, butterflies, birds, and bats (Roulston and Cane, 2000). Other plants provide nectar, oils, resins, fragrances, pheromone precursors, and other resources to induce visitation and pollen delivery (Barth, 1991; Buchmann, 1987; Dafni et al., 2005; Roulston and Cane, 2000; Roulston et al., 2000).
Plants and pollinators vary in their degree of interdependence. Some plant species depend primarily on a single species or genus of pollinator, which in turn has restricted sources of pollen or nectar. An example of a closely dependent association is the relationship between plants in the genus Yucca (Agavaceae) and their pollinators, the aptly named yucca moths of the genus Tegeticula (reviewed in Pellmyr, 2003). In this mutualism, estimated to be more than 40 million years old, the adult yucca moth is the primary pollinator of yucca and the developing yucca seeds the main nutritive source for the caterpillar. The female moths have unique structures, called tentacles,
which are used to collect and compact comparatively large quantities of pollen (up to 10 percent of the moth’s weight) from yucca flowers. After gathering a pollen mass, the moth flies off and visits another flower, in which she lays eggs. Then, in a distinctive series of stereotyped behaviors, she places part of the pollen load directly on the stigma surface to achieve pollination and subsequent fertilization, thereby guaranteeing a food source for her offspring.
Such specialized relationships, however, are the exception in plant-pollinator interactions. In many cases, if not most, associations are highly opportunistic. Over a 2-year period, for example, at least 45 species of insects in 5 orders were observed to visit Geranium thunbergii flowers in a natural population; of these, 11 species in 3 orders served as principal pollinators (Kandori, 2002). Principal pollinators of a particular species can vary spatially as well as temporally; the alpine sky pilot, Polemonium viscosum, is pollinated primarily by bumble bees at high elevations and by flies at low elevations in its native Rocky Mountain range (Galen et al., 1987). Humans have understood the agricultural importance of pollination—that plants require pollen transfer to produce fruits and set seed—for at least 3,500 years. However, the idea that seeds result ultimately from the deposition of pollen grains on stigmatic surfaces was not clearly articulated until the 17th century (Camerarius, 1694) and even then was slow to gain acceptance. The systematist Carolus Linnaeus, for example, identified the sexual organs of plants as important components of his classification system, to the disapprobation of the 18th century’s religious establishment.
The idea that animals play a role in cross-pollination, the transfer of pollen from one individual to another, was not clearly articulated until close to a century later (Kölreuter, 1761; Sprengel, 1793). Joseph Gottlieb Kölreuter, a professor of natural history at the University of Karlsruhe, Germany, demonstrated that insect visitation was a prerequisite for seed production in several economically important fruits, vegetables, and ornamental flowers and put his knowledge to practical use by developing technique for artificial fertilization and conducting the first cross-hybridization of two plant species (Mayr, 1986; Sinnott, 1946).
The great insights and practical achievements of Kölreuter and Sprengel failed for the most part to inspire their contemporaries, but a half-century later their work had a profound influence on the thinking of another biologist—Charles Darwin. The publication of The Origin of Species by Means of Natural Selection or The Preservation of Favoured Races in the Struggle for Life (The Origin of Species, in short) in 1859 ushered in a new era of experimental pollination biology. In his preface, Darwin described the process of coadaptation, which is what allows living organisms to serve as selective agents in the same manner as abiotic forces and specifically mentioned pollination as an example.
Darwin’s extensive writings on plant-pollinator relationships, not only in The Origin of Species but also in The Various Contrivances by which British and Foreign Orchids are Fertilised by Insects and the Good Effects of Intercrossing in 1862 and other subsequent publications, summarized an extensive literature and described his own meticulous experiments, thereby providing compelling evidence of the significance of pollination in the lives of plants. Demonstration of the evolutionary mechanism to account for the reciprocally adaptive relationship between plants and their pollinators provided the impetus for an explosion of interest in pollination biology (Delpino, 1868–1875; Knuth, 1906, 1908, 1912; Müller, 1869; Müller and Delpino, 1869) that laid the foundation for agricultural applications and for contemporary experimental studies.
POPULATION MANAGEMENT
Active Management
Recognition of the mechanisms of biotic pollination led to important agricultural innovation, with extensive economic consequences (Box 1-2); management of pollinator species allowed for enhanced crop productivity and for commercialization (and export) of numerous crop plants. In North America, only a handful of pollinator species are actively managed—that is, they are semidomesticated, produced in large quantities, and bought and sold commercially. Of these, Apis mellifera L., the western honey bee, is the premier actively managed pollinator worldwide, highly valued for its activity as a pollinator and for its production of wax and honey (Delaplane and Mayer, 2000; Free, 1993; Kearns et al., 1998; McGregor, 1976).
Native to Eurasia, the honey bee has been hunted for its honey and wax for at least 6,000 years (Crane, 1983, 1990) and records of semidomestication and hive management date back to ancient Egypt (Crane, 1999). A. mellifera rapidly became the primary pollinator for modern agriculture, and managed colonies were transported around the world, first arriving in North America with European colonists in the 1600s (Sheppard, 1989a). Modern apiculture in North America dates to 1862, when L. L. Langstroth, a Philadelphia minister who kept bees as a hobby, exploited the concept of “bee space” to construct movable-frame, top-bar hives that allowed beekeepers to harvest honey, manipulate their colonies, and increase efficiency without harming the bees (Langstroth, 1862). Langstroth’s invention resulted in the large-scale commercial beekeeping and honey industry that exists today.
Honey bees pollinate more than 100 commercially grown crops in North America (Delaplane and Mayer, 2000; Free, 1993; Kearns et al., 1998; McGregor, 1976). In the United States, about 135,000 beekeepers
manage 2.4 million colonies of honey bees. Most beekeepers (about 94 percent) are hobbyists with 25 colonies or fewer. Another 5 percent are called sideliners, each managing 25–300 colonies. Only about 1 percent are commercial beekeepers and they generally manage between 300 and 60,000 colonies each to provide most of the nation’s pollination services (D. Weaver, The American Beekeeping Federation, presentation to the committee on October 19, 2005).
Beekeepers in the United States have formed hundreds of local associations and two national trade organizations. The American Beekeeping Federation (ABF) has about 1200 members (ABF, 2005), and the American Honey Producers Association (AHPA) has about 500 members (S. Park, AHPA, personal communication, June 12, 2006). The Eastern Apicultural Society, Heartland Apicultural Association, and the Western Apicultural Society meet annually and provide extensive educational opportunities for beekeepers. Many beekeepers, however, do not belong to any formal organization.
Other species of pollinators for which active management systems have been developed include several species of bumble bees (Bombus), mainly for pollination of greenhouse tomatoes (de Ruijter, 1997; Hughes, 1996; Kevan, et al., 1991; Macfarlane et al., 1994; Plowright, 1996; van Heemert et al., 1990), and leafcutting bees (Megachile rotundata) (Bohart, 1972a; Frank, 2003), which pollinate most of the alfalfa in parts of the arid Pacific Northwest (R. Bitner, presentation to the committee, January 14, 2006). To a lesser extent, alkali bees (Nomia melanderi) (Bohart, 1972a) also are managed for alfalfa pollination (Stephen, 2003).
Mason bees, including the Japanese horn-faced bee, Osmia cornifrons, are managed to some extent, mainly for pollination of apple orchards in the eastern United States (Batra, 1982; Bohart, 1972b), although they are used extensively in Japan for pollinating the entire apple crop. Several native Osmia species, notably O. lignaria, are used to pollinate apples in the northwestern United States (Bosch and Kemp, 2002) and in eastern Canada (Sheffield, 2006) and to pollinate cherries (Bosch and Kemp, 1999, 2000, 2001). Methods for cultivating this species are well developed (Griffin, 1993; Torchio, 2003).
Literature on the culture and management of many alternative pollinators is available (Batra, 1994a,b; Bosch and Kemp, 2001; Free, 1970; Kevan et al., 1990; Shepherd et al., 2003; Torchio, 1990, 2003). For some crops, bumble bees, megachilids, and other native bees are more efficient pollinators than are honey bees (Cane, 2002; Javorek et al., 2002; Maeta and Kitamura, 1981; Tepedino, 1997) and Osmia species serve as alternative pollinators for almonds (Bosch and Kemp, 2000; Torchio, 2003), red raspberries and blackberries (Cane, 2005), pears (Maeta et al., 1993), blueberries (MacKenzie et al., 1997; Stubbs and Drummond, 1997a,b; Stubbs et al., 1997), and sweet clover (Richards, 2003).
|
BOX 1-2 What Is Pollination Worth? Pollination has value in two very different senses. Its intrinsic value or essential worth is conceptual, so it cannot be measured easily. The economic value of pollination is its worth for human ends, as determined through exchanges of goods or services (NRC, 2005). The aggregate economic value of pollination services is the difference between what consumers are willing to pay (demand) and what it costs producers to provide those services (supply). Consumers’ willingness to pay comes not only from direct “use” of pollination (for example, eating fruits of pollinated blossoms or enjoying the aesthetics of butterflies visiting garden flowers), but also from appreciation for the existence of pollinators and their services to future generations. Where markets do not exist (as for pollination services provided by wild pollinators), it is difficult to estimate economic value, although environmental economists have developed methods of approximation (NRC, 2005). Where markets exist (as for agricultural crop pollination), economic values can be estimated for discrete changes in supply and demand. For the case of commercial honey bee pollination services, the consumers are the crop growers and the producers are the beekeepers. The demand curve that describes the number of honey bee colonies the growers are willing to rent at different prices for pollination is derived from what individual growers expect to earn from yield gains attributable to pollination; their demand depends on expected crop prices, expected yield gains, and the costs of available alternative means of pollination. Because growers raise different crops under different conditions, some are willing to pay more than others. An almond grower whose production is worth $2,000 per acre will be willing to pay more than will a grower of apples for cider that is worth less than $500 per acre. The supply curve that describes how many honey bee colonies individual beekeepers are willing to rent out depends on the beekeepers’ costs of production and what they can earn from alternative uses of their bees. Beekeeping also differs in costs and earning opportunities. A beekeeper who must transport bees a long distance will have higher costs than will one who |
An improved understanding of the mechanics of pollination and of its active management led to the commercialization and worldwide expansion of many crops, hitherto an impossibility. Common figs (Ficus carica) and Smyrna figs in California are a case in point. California, second only to Turkey in fig production, has 18,357 acres in fig production that was worth nearly $10 million in 1998 (Farrar, 1999). Ficus species are primarily pollinated by agaonid fig wasps in highly species-specific associations (Bronstein, 1988; Machado et al., 2005). Fig production did not become established in
|
is close to an orchard that needs pollination. More details on supply and demand effects and an example relating to almond pollination are presented in Chapter 4. Three basic methods have been used to estimate the value of commercial honey bee pollination services (Table 1-1). The first is simply to equate the value of services with the amount paid for them (Rucker et al., 2005). The approach does not capture potential consumer willingness to pay, nor does it account for beekeepers’ production costs. The second approach also is the most common. This method is to estimate the value of pollination services by taking the total value (market price multiplied by quantity) of a crop and multiplying that value by a coefficient for the crop’s estimated dependence on commercial pollination (Levin, 1983; Morse and Calderone, 2000; Robinson et al., 1989b). That approach captures consumer willingness to pay, but fails to subtract beekeepers’ production costs. It attributes all crop value to pollination and ignores other inputs required to produce the crop. Neither of the first two methods considers that a shift in honey bee supply (for example, because of a new disease or pest) could raise crop prices and thus alter grower demand for pollination services (Kevan and Phillips, 2001). Southwick and Southwick (1992) attempted to capture that effect by estimating the price elasticities of demand for U.S. agricultural crops. Even where markets exist and price effects are considered, it is impossible to make reliable estimates of the total value of an ecosystem service such as pollination. The market value of pollination supply shifts can be reliably estimated only for relatively small perturbations from values that have been observed in the past. Even the threat of a complete loss of pollination services would induce some people to pay extraordinarily high prices to prevent a total loss of the service. Others, however, would do without. Such price-quantity relationships fall well outside prior experience. For pollination services provided by wild pollinators where markets do not exist, current estimates of nonmarket value are fraught with limiting assumptions. The economic value of extreme deviations, such as losing all pollination services, cannot be soundly estimated (Heal, 2000). If calculable, the economic value of keeping pollination services would be very high, similar to their intrinsic value. |
California until fig wasps were imported in the 1890s for pollination and growers learned to identify the proper species for pollination and determine overwintering requirements to synchronize wasp life cycles with the plants (McGregor, 1976; Swingle, 1908).
Although pollinators are in most cases managed for crop pollination, there are examples of pollinator management to achieve other goals. Hobby beekeepers often keep bees primarily for honey production or for personal satisfaction rather than for pollination. Honey bees have been recruited
for delivery of various biological control agents to protect field and greenhouse-grown crops against fungal pathogens and pests (Kevan et al., 2005). Bees are used to deliver Bacillus subtilis to blueberry flowers to suppress Monilinia vaccinicorymbosi, or mummy berry disease, a devastating fungus (Dedej et al., 2004). They also have been used to deliver Trichoderma harzianum 1295-22, a commercially produced agent for control of the pathogenic fungus Botrytis cinerea on strawberries (Kovach et al., 2000). There is a continuing effort to investigate the potential of honey bees as biological monitors for environmental contaminants (Bromenshenk et al., 1995) and land mines (Bromenshenk et al., 2003).
Passive Management
In addition to active management, pollinators can also be managed passively—that is, their activities can be manipulated by altering environmental conditions to promote their diversity and population growth or to influence particular behavior (Shepherd et al., 2003; Vaughn et al., 2004). Passive management includes farming to promote the growth of floral resources, providing artificial nest materials and nest sites, and protecting nesting habitat. All of these practices are designed to increase the diversity of the pollinator community and the abundances of particular species (Kevan et al., 1990; Kremen and Chaplin, in press).
VALUE OF POLLINATION
Pollination as a biotic process has both commercial and ecological value. In the context of agriculture, pollination provides a wide range of benefits to a broad diversity of commodities across North America. In some cases, production of the commodity itself results directly from the act of pollination (for example, fruit production). In other cases, although pollination does not result in production of the commodity itself, the process contributes to crop propagation (for example, production of seeds used to grow a root crop such as carrots) or quality (for example, size of tomatoes has been linked to repeated pollination). There are indirect benefits as well, through food-chain relationships. Alfalfa seed, a bee-pollinated crop with an annual value of $109 million (direct effect), is used to produce hay for livestock forage that is valued at $4.6 billion per year (indirect effect) (Morse and Calderone, 2000). Although these indirect effects tend to exaggerate the economic value of pollination, they have been used in several widely cited studies (see Table 1-1).
The annual value of honey bee pollination to U.S. agriculture has been variously estimated at $150 million (Rucker et al., 2005), $1.6–5.7 billion (Southwick and Southwick, 1992), $9 billion (Robinson et al., 1989a),
TABLE 1-1 Value of U.S. Agricultural Production Attributable to Honey Bee Pollination: Comparison of Studies
|
Study |
Reference Year |
Total Value ($ billion) |
Direct Crop Value ($ billion) |
Indirect Crop Valuea($ billion) |
Animal Value ($ billion) |
Notes |
|
Levin, 1983 |
1980 |
19.0 |
5.9 |
6.0 |
7.2 |
Author attributes 10% of cattle value to bees via alfalfa hay |
|
Robinson et al., 1989a,b |
1985 |
9.7 |
6.1 |
3.6 |
0 |
|
|
Southwick and Southwick, 1992b |
1986 |
5.7 |
5.7 |
0 |
0 |
Value based on price elasticity of supply change |
|
Morse and Calderone, 2000 |
1996–1998 |
14.6 |
7.8 |
6.7 |
0 |
|
|
Rucker et al., 2005 |
2004 |
0.15 |
0.15 |
0 |
0 |
Value is pollination fees paid |
|
aCrops that receive indirect benefits include alfalfa hay, asparagus, broccoli, carrot, cauliflower, celery, onion, and sugar beet. bEstimate for no replacement of bees; no price effects. |
||||||
$14.6 billion (Morse and Calderone, 2000), and $18.9 billion (Levin, 1983). The annual benefit of honey bee pollination in Canada has been estimated at $443 million by Scott-Dupree and colleagues (1995). More recent data are shown on the website of the Canadian Honey Council (http://www.honeycouncil.ca/users/folder.asp). The lowest U.S. figure is an estimate of the annual value of pollination fees actually paid by farmers (Rucker et al., 2005)—a value that does not capture the higher fees that farmers would be willing to pay to ensure good pollination. Table 1-1 compares studies that include estimates of such willingness to pay for pollination services and it provides a breakdown of total reported values as direct benefits to crops, indirect benefits to crops, and indirect benefits to livestock. The value of direct benefits to crops clusters in the range of $5 billion to $10 billion (the higher end adjusted to 2005 dollars). Values reported by Morse and Calderone (2000) and by Levin (1983) include indirect benefits of the honey bee pollination required for seed production in alfalfa hay, asparagus, broccoli, carrot, cauliflower, celery, onion, and sugar beet. Levin (1983) included 10
percent of the value of cattle and dairy production that he attributed to alfalfa hay whose seed requires bee pollination. Attributing the full market value of such indirect effects to pollination exaggerates the economic value of pollination services, because indirect products like alfalfa hay or cattle require many production inputs besides alfalfa seed. Even the alfalfa seed made possible by pollination requires that farmers provide other costly production inputs. These and other limitations of estimating economic values are discussed in Box 1-2. Given the estimates currently available, consistent comparisons can be made across those economic values based on the direct effects of pollinators.
The contributions of A. mellifera are not unique: Alfalfa leafcutting bees and bumble bees also pollinate crops. An estimated $2 billion to $3 billion value in annual crop pollination can be attributed to the activities of native bees and other insects (Losey and Vaughn, 2006; Prescott-Allen and Prescott-Allen, 1986; Southwick and Southwick, 1992).
Some vertebrates also operate as pollinators of ecologically and economically important plants. Tropical trees of the family Bombacaceae, which includes species used for timber, silk cotton, balsa wood, and other products, rely primarily on bats for pollination (Bawa, 1990; Watson and Dallwitz, 1992). Many columnar cacti and agaves, which are important sources of alcoholic beverages (tequila, mescal) and other products (sisal fibers), also depend on bats and birds for pollination (Arizaga and Ezcurra, 2002; Arizaga et al., 2002; Fleming et al., 2001a,b; Grant and Grant, 1979; Rocha et al., 2005; Valiente-Banuet et al., 1996; but see also Slauson, 2000, 2001). Globally, pollinators are fundamentally important for the production of roughly 30 percent of the human diet and most fibers (cotton and flax), edible oils, alcoholic beverages, nutraceuticals, and medicines created from plants (Buchmann and Nabhan, 1996; McGregor, 1976; Roubik, 1995).
Estimating the value of pollinators and pollination in natural ecosystems and predicting the consequences of their losses are considerably more challenging than estimating their economic value in agriculture. Such estimates are complicated by both the number of species involved (globally, more than 400,000) and the relative paucity of information available for most of those species. For example, in their effort to calculate the economic value of ecological services provided by insects, Losey and Vaughan (2006) did not attempt to place a dollar value on the contributions of pollinators to maintenance of natural plant communities, although it is reasonable to assume that a significant proportion of plants in uncultivated terrestrial communities rely upon pollinators. These plants, in turn, contribute to many ecosystem services of value to humans, such as water filtration, carbon sequestration, and flood and erosion control (Daily et al., 1997). An added complication is that insect pollinators may contribute ecosystem services other than pollination in their larval stages. The value of these services is
equally difficult to calculate, particularly without a complete understanding of all aspects of pollinators’ life histories.
Dobson et al. (2006), however, developed a system for assessing the susceptibility of different ecosystem services to species loss. According to this system, which takes into consideration trophic level interactions, redundancies, and competition, ecosystem services are classified into Types A through E, with Type A services at one extreme identified as those in which species losses are mostly compensated for by co-occurring species and Type E services identified as “the most brittle services; for these services, small changes in species biodiversity result in large changes in the provisioning of ecosystem services.” In Type C, an intermediate response, a linear decline in ecosystem service is expected with each species loss. In this system, pollination is considered a Type C or E service for most ecosystems, with species losses having significant impacts on trophic stability. Indeed, pollination is the only mutualistic association singled out by Dobson et al. (2006) for assessment.
STATE OF KNOWLEDGE
The study of pollinator-plant interactions is a thriving, albeit small, area of inquiry. There are no professional societies dedicated to this pursuit, and publications in the field appear in a wide range of journals. A search of the Ovid serials database with the keyword “pollination” yields 6906 publications for the period 1990–2005, with the numbers steadily increasing from 2000 onward (see also Figure 1-1 in Kearns and Inouye, 1993). Although several universities alone or in partnership with state agencies offer classes on bees and beekeeping (for example, http://www.masterbeekeeper.org/masterbeekeeper.htm, http://www.ento.vt.edu/~fell/apiculture/summer/summerb.htm, http://www.life.uiuc.edu/entomology/bee-course.html, http://www.news.cornell.edu/Chronicle/00/5.11.00/bee-course.html, http://neipmc.org/ipm_news_popover.cfm?id=821), courses in pollination biology are rare, although it often is included as a topic in seminars on plant-insect interactions (University of Southern Mississippi, Auburn University, Mesa State College, University of Toronto, Canada). Classes in pollination biology have been taught in recent years at the University of Arizona, University of California (Davis and Berkeley), University of Maryland, University of Texas at Austin, St. Louis University, Humboldt State University, and California State University at Fullerton. In Canada, pollination biology courses have been recently offered at the University of Guelph, Ontario, and at the University of Manitoba. In Puerto Rico, a course is offered at the Departamento de Química y Biología (http://mail.udlap.mx/~cvergara/EcolPol/EcolPol.html). Many pollination biologists, conservationists, and land managers benefit from an annual 10-day class (the Bee Course, now in its ninth year,
http://research.amnh.org/invertzoo/beecourse) on the systematics and biology of bees native to the southwestern United States and Mexico that is presented by the American Museum of Natural History-Southwestern Research Station at Portal, Arizona. In 2006, the McGuire Center for Lepidoptera & Biodiversity in Gainesville, Florida, announced plans to offer a course on “natural history and identification, captive propagation, host plant care and needs, field/lab research techniques, permitting/recovery planning basics, population monitoring, habitat restoration, butterfly exhibitry, education and outreach, partnership building,” and the like (http://www.aza.org/prodev/ButterflyCon/).
Despite the widely held assumption that “the birds and the bees” in its literal sense is a concept that is familiar even to schoolchildren, incorporation of pollination biology into primary and even secondary science curricula is far from routine. There are no specific references, for example, to pollination biology or pollinators in the National Academy of Sciences’ own set of science education standards (NRC, 1996) or in the benchmarks for scientific literacy published by the American Association for the Advancement of Science (AAAS, 1994).
HISTORY OF CONCERN
Concerns about the status of pollinators in North America over the last quarter-century have arisen in two different contexts. The agricultural community has voiced concerns over fluctuations in the health and availability of A. mellifera, the principal managed pollinator in the United States, and associated impacts on crop production, whereas the ecological community has noted declines worldwide as part of a larger effort addressing biodiversity losses and associated impacts on ecosystem services.
Managed Pollinators
Although the U.S. Department of Agriculture’s (USDA’s) National Agricultural Statistics Service (NASS) reported that 4.2 million colonies of honey bees were managed by beekeepers in the United States in 1981, the detection of the parasitic honey bee mites Acarapis woodi Rennie and Varroa destructor (formerly V. jacobsoni Oud; see Anderson and Trueman, 2000) in the United States in 1984 (Anonymous, 1987; Delfinado-Baker, 1984) and in 1987 (Delfinado-Baker 1988), respectively, ushered in an era that has been marked by fluctuations in colony numbers that is overlaid by a general downward trend (Chapter 2). Although A. woodi was very damaging immediately after its introduction, North American honey bee populations exhibited some resistance (Chapter 3) and, after several years, deaths from that parasite appeared to decline. However, the actual contribution of this
parasite to changes in honey bee colony numbers was never clearly established. Today, the primary concern among beekeepers is the varroa mite, which continues to cause major losses of managed hives (Caron and Hubner, 2001; Finley et al., 1996; Lumkin, 2005). Infection with V. destructor is fatal to most honey bee colonies of European ancestry (Beetsma, 1994; DeJong, 1990; DeJong et al., 1984; Matheson, 1994; Morse and Gonçalves, 1979). The high rate of mortality is the combined result of several factors, including low levels of natural resistance to mites in the honey bee population; inadequate stock development and production facilities; widespread use of pesticides, which helps to maintain mite-susceptible genotypes in the honey bee population; and widespread pesticide resistance in the mite population.
Few honey bee breeding programs (artificial selection) have successfully consolidated low levels of existing mite resistance into strains with significant levels of mite resistance (Harbo and Harris, 1999a). Although these strains hold promise for mite management, they are slowly being integrated into beekeepers’ management programs (Chapter 6; Sheppard, 2006).
Honey bees have been widely regarded as having suffered under the weight of those stresses. NASS’s 2005 estimate of 2.41 million honey bee colonies in the United States is a 28 percent decline from the pre-mite 1981 numbers, after correction for a 1985 change in NASS methodology1 (Chapter 2). Also, parasitic mites had, by all accounts, an even more serious and negative effect on the population of feral honey bee colonies (Hoopingarner, 1991; Kraus and Page, 1995; Loper, 1995, 1996, 1997).
These losses occurred as demand for agricultural pollination services was increasing dramatically, particularly for crops that depend completely on pollinators. The almond-growing business presents a compelling example (Figure 1-1). Over the 25 years between 1980 and 2005, U.S. almond acreage increased by nearly 70 percent. Approximately 1.4 million bee colonies are needed to pollinate 550,000 acres of almonds in California, and in 2005 a shortage of colonies led to imports of colonies from Australia (Flottum, 2005). According to forecasts of the California Almond Board (cited in Sumner and Boriss, 2006), almond growing will expand to 850,000 acres by 2012, eventually requiring the services of 2.12 million colonies of pollinators. Other pollinator-dependent crops include squash (Figure 1-2) and muskmelon (cantaloupe) (Figure 1-3); domestic production of these crops has nearly doubled over the past two decades.
The growth in demand for crop pollination is not restricted to field crops. Although USDA does not collect data on acreage of greenhouse tomatoes, other sources suggest a recent large increase. Total greenhouse crop
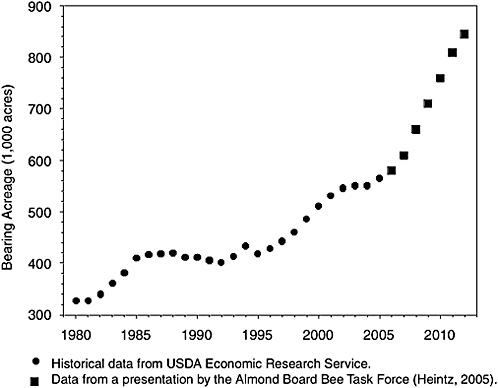
FIGURE 1-1 Bearing acreage of U.S. almond trees in 1980–2010. Almost all almonds are grown in California.
acreage increased by 40 percent between 1996 and 1999 (Dodson et al., 2002). In Mississippi, the number of commercial growers went from 15 in 1988 to about 130 in 2006 (Mississippi State University Extension Service, 2006; Snyder, 2006). The top 15 states in greenhouse tomato production had almost 600 acres in cultivation in 1999 (Snyder, 1999); in 2002, the total was about 750 acres (Dodson et al., 2002). Greenhouse tomatoes require pollination that is now accomplished mainly by managed colonies of bumble bees (Chapter 3). The growth in production suggests the demand for those bees will increase as well.
Wild Pollinators
Concerns about pollinator status are in at least one way unique in discussions of threats to biodiversity in general, in that such concerns are often directly and explicitly linked to concerns about the status of another group of organisms—the mutualistic flowering plant partners of pollinators. Over the past quarter-century, declines in wild pollinator populations
of various descriptions have been reported in Europe, Asia, Central and South America, Africa, and Australia (Allen-Wardell et al., 1998; Biesmeijer et al., 2006; Donaldson, 2002; Kearns et al., 1998; Oldroyd and Wongsiri, 2006; de Ruijter, 2002). By contrast, few pollinator species are monitored in North America. Declines in bat populations have been so dramatic that two of the three U.S. species—the lesser long-nosed bat (Leptonycteris curasoae) and the Mexican long-nosed bat (L. nivalis)—are now listed as endangered under the terms of the U.S. Endangered Species Act. All three species are listed on the federal list of species at risk of extinction in Mexico (SEMARNAT, 2002). Around the world, almost half of the insect extinctions documented have involved flower-visiting species (Appendix C). The first insect to be listed as officially endangered in the United States (Appendix D) was the Schaus swallowtail, Papilio aristodemus, a flower-visiting species and a presumed pollinator. Flower-visiting Lepidoptera, many of which are actual or potential pollinators, currently dominate the list of endangered species: 17 species of butterfly and 3 species of moth constitute more than half of all insect species listed as endangered (http://ecos.fws.gov/servlet/TESSWebpage) (Appendix D). Evidence for population decline
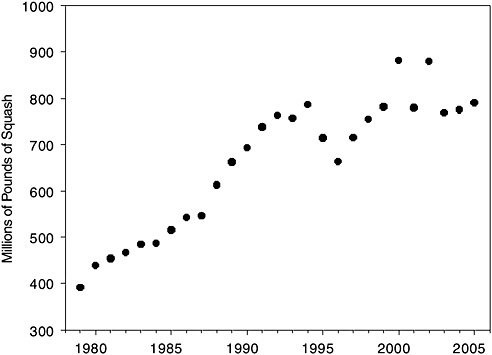
FIGURE 1-2 U.S. squash production, 1980–2005. Available evidence shows that squash plants must be pollinated by insects, and that honey bees are the chief pollinators (McGregor, 1976). Source: USDA-NASS.
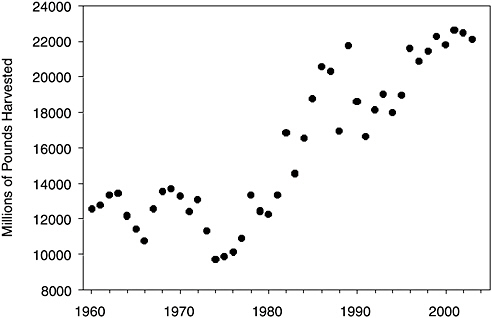
FIGURE 1-3 U.S. muskmelon production, 1960–2006. Flowers must be pollinated to produce cantaloupes. Source: USDA Economic Research Service.
(Appendix E) and local population extinction of other pollinators has begun to accumulate (Kremen et al., 2002a; Larsen et al., 2005).
Comprehensive recognition of the value of ecosystem services provided by pollinators (Daily et al., 1997; Kremen et al., 2002a, Kremen and Chaplin, in press) is relatively recent and quantitative studies on pollinator populations in North America are few in number. Among the possible causes suggested for declines in wild pollinator numbers are fragmentation, degradation and loss of habitat, nontarget effects of agricultural pesticides, competition from invasive species, and introduced diseases (Johansen, 1977; Kearns et al., 1998; Kevan, 1974, 1975a, 1999, 2001; Kremen and Ricketts, 2000; Morandin et al., 2005; Rathcke and Jules, 1993).
Efforts to Address Concern
The concept of a pollinator “crisis”—localized extinctions and possibly a global decline in the number and viability of pollinating species contributing to trophic collapse (sensu Dobson et al. 2006)—began to gain recognition as a critical issue in the early 1990s. Major losses in managed honey bee colonies led to concern in the United States about this critical pollinator (Watanabe, 1994). The publication of The Forgotten Pollinators (Buchmann and Nabhan, 1996), which extended the concern to nonmanaged species
and placed the issue within the greater context of biodiversity decline, galvanized the ecological community. Buchmann and Nabhan’s book concluded with a call for a U.S. national policy on pollination and pollinators. Subsequent emphasis of pollination as a fundamental “ecosystem service” (Daily et al., 1997) led to an explosion of interest in the international policy arena (Allen-Wardell et al., 1998; Costanza et al., 1997; Eardley et al., 2006).
Among the first international efforts to address concern specifically about the status of pollinators was a proposal by the government of Brazil at the second meeting of the Subsidiary Body on Scientific, Technical and Technological Advice (SBSTTA) of the Convention on Biodiversity, which convened in Montreal, Quebec, September 2–6, 1996 (Appendix F). The purpose was to establish a program in agricultural biodiversity, which includes an “international pollinator conservation initiative” (Campanhola et al., 1998). Subsequently, the Third Conference of the Parties (COP3) to the Convention on Biodiversity (CBD), which met in Buenos Aires, Argentina, November 4–15, 1996, drafted Decision III.11, “Conservation and Sustainable Use of Agricultural Biodiversity,” identifying pollinators as a “priority group” for case studies. That, in turn, led to an International Workshop on Conservation and Sustainable Use of Pollinators in Agriculture, which placed an emphasis on bees, when it met in São Paulo, Brazil, in October 1998. The workshop report, Pollinating Bees: The Conservation Link Between Agriculture and Nature. Proceedings of the Workshop on the Conservation and Sustainable Use of Pollinators in Agriculture, with Emphasis on Bees (Kevan and Imperatriz Fonseca, 2002), included the “São Paolo Declaration on Pollinators,” which proposed the establishment of an International Pollinator Conservation Initiative (Dias et al., 1999). An early response to that call was made at the first meeting of the Systematics Society of Southern Africa, which founded the African Pollinator Initiative in January 1999.
The São Paolo proposal was ultimately presented by the Brazilian government to the SBSTTA at its fifth meeting and, at COP5—the fifth meeting of the Conference of the Parties—the International Initiative for the Conservation and Sustainable Use of Pollinators was created. That initiative was to promote international cooperation with several goals:
-
Monitor pollinator decline, its causes, and its impact on pollination services.
-
Address the lack of taxonomic information on pollinators.
-
Assess the economic value of pollination and the economic impact of the decline of pollination services.
-
Promote the conservation and the restoration and sustainable use of pollinator diversity in agriculture and related ecosystems (CBD, 2005, p. 2).
The United Nations Food and Agriculture Organization was invited to coordinate the initiative among the participating nations and agencies and to devise a plan that was based on the recommendations in the São Paulo Declaration on Pollinators. In April 2002, the International Pollinator Initiative was approved at the sixth meeting of the Conference of the Parties, which was convened in the Netherlands (www.biodiv.org/doc/meetings/sbstta/sbstta-07/official/sbstta-07-09-add1-en.doc). In March 2006, at the United Nations Convention on Biodiversity, Conference of the Parties, general meeting 8 in Curitiba, Brazil, the publication, Pollinators and Pollination: A Resource Book for Policy and Practice (Eardley et al., 2006), was heralded.
Between 1995 and 1999, the tri-national (United States, Canada, Mexico) Forgotten Pollinators (FP) Campaign was co-founded and directed by Gary Nabhan and Stephen Buchmann at the Arizona-Sonora Desert Museum in Tucson, Arizona. During its later years, the FP Campaign shifted its emphasis to focus on migratory pollinators (bats, hummingbirds, white-winged doves, monarch butterflies) in the United States and Mexico. In 1999, the North American Pollinator Protection Campaign (NAPPC) was launched by the Coevolution Institute in San Francisco, California, in collaboration with the National Fish and Wildlife Foundation. NAPPC is a tri-national public-private partnership of more than 100 organizations and agencies that promote pollinator awareness, policies, educational outreach, research, and conservation. Also in 1999, USDA and the U.S. Geological Survey (USGS) sponsored a joint meeting in Logan, Utah (Tepedino and Ginsberg, 2000), to explore the issue of pollinator decline. In November 1999, the USDA Agricultural Research Service (ARS) convened a workshop to obtain stakeholder input from state departments of agriculture, universities, pesticide companies, beekeepers, and extension agents on research priorities for the Bees and Pollination Component of the Crop Production National Program Writing Teams. These teams were formed at each of the workshops and consisted of ARS scientists and members of the ARS National Program Staff. The first function of each was to identify problem area topics for inclusion in the National Program Action Plan. Subsequently, individual team members were assigned as principal authors for each area identified. Writing teams and individual writers used input from the workshops, their own knowledge, and input from other ARS scientists and cooperators to identify research goals and activities to develop this action plan (http://www.ars.usda.gov/research/programs/programs.htm?np_code=305&docid=883).
Also in 1999, the National Science Foundation (NSF) funded a meeting at the National Center for Ecological Analysis and Synthesis (NCEAS) on pollinator decline, the proceedings of which were published in 2001 (http://www.ecologyandsociety.org/vol5/iss1/art1/) in the journal Conserva-
tion Ecology (now Ecology and Society). NSF also provided funds in fiscal years 2005–2007 for a series of NCEAS workshops to determine how to restore pollinators and pollination function in degraded landscapes. Other federal agencies with an interest in pollinator status include the U.S. Fish and Wildlife Service’s Division of Environmental Quality, which maintains a pollinator website (http://www.fws.gov/contaminants/Issues/Pollinators.cfm) that has statements on pollinator decline. The USGS National Biological Information Infrastructure, a collaborative program designed to provide access to data on natural resources within U.S. borders, also has a pollinator decline program (http://www.nbii.gov).
CHARGE TO THE COMMITTEE
In 2002, NAPPC approached the National Research Council with a request for a study to review the literature on pollinating animals in North America. With funding provided by USDA, USGS, and The National Academies, the National Research Council’s Board on Life Sciences and Board on Agriculture and Natural Resources jointly convened an ad hoc committee to document the status of pollinating animals in North America. The questions to be addressed included whether, and to what degree, pollinators are experiencing serious decline; in cases where decline can be established by available data, what its causes are; and what the potential consequences of decline might be in both agricultural and natural ecosystems. The study committee was tasked to make recommendations on research and monitoring needs to provide improved information and on any conservation or restoration steps that could prevent, slow, or reverse potential decline. The committee also was asked to compile and analyze the published literature, determine the current state of knowledge on pollinator status, identify knowledge gaps, and establish priorities for addressing these gaps.
To address its charge, the committee assessed the status of pollinators in the United States, Canada, and Mexico between the Atlantic and Pacific Oceans (Chapter 2) and some of the causes of decline in pollinator populations (Chapter 3). However, the extent to which the committee could discuss each species in different regions depended largely on the availability of data. The proportion of the report devoted to honey bees reflects the amount of knowledge and data available for this species. In contrast, the proportion of the report devoted to wild pollinators reflects the sparse data and our incomplete knowledge on those groups despite their critical role in ecosystem functioning. The later chapters of the report discuss the potential impact of pollinator decline in agriculture and natural areas (Chapter 4) and suggest ways to monitor, conserve, and restore managed and unmanaged pollinators (Chapters 5 and 6).





















