
15
Biological Design in Science Classrooms
EUGENIE C. SCOTT and NICHOLAS J. MATZKE
Although evolutionary biology is replete with explanations for complex biological structures, scientists concerned about evolution education have been forced to confront “intelligent design” (ID), which rejects a natural origin for biological complexity. The content of ID is a subset of the claims made by the older “creation science” movement. Both creationist views contend that highly complex biological adaptations and even organisms categorically cannot result from natural causes but require a supernatural creative agent. Historically, ID arose from efforts to produce a form of creationism that would be less vulnerable to legal challenges and that would not overtly rely upon biblical literalism. Scientists do not use ID to explain nature, but because it has support from outside the scientific community, ID is nonetheless contributing substantially to a long-standing assault on the integrity of science education.
Nature is full of complex biological adaptations such as the camera eye, the bird wing, the bacterial flagellum, the mammalian immune system, or the complex traps of orchid flowers. Evolutionary biology continues to make progress in explaining such fascinating structures through the scientific process of positing natural explanations
National Center for Science Education, Inc., 420 40th Street #2, Oakland, CA 94609-2509.
and testing them against the natural world. Nevertheless, in recent years scientists have been forced to confront a resurgence of opposition to evolution in the political realm of public education. This new antievolutionism is called “intelligent design” (ID). Its proponents allege that it is a revolutionary new scientific explanation for complex adaptations, that it is purely secular and definitely not creationism, and that it is therefore pedagogically and legally appropriate for public school biology classrooms. However, an analysis of ID shows that in both content and history, it is a subset of an earlier antievolution movement known as creation science.
HISTORICAL BACKGROUND: FUNDAMENTALIST OPPOSITION TO EVOLUTION EDUCATION IN THE UNITED STATES
The creationism/evolution battle began in the 1920s as a by-product of the acrimonious split of American Protestantism into “fundamentalist” and “modernist” camps. Fundamentalism arose in the early 20th century in reaction to issues such as modern historical criticism of the Bible, technological and social progress, and evolution (Marsden, 1991; Armstrong, 2000). Modernists moved toward viewing the Bible as allegorical and as a product of human history, whereas fundamentalists tried to defend what they viewed as “the fundamentals” of the Christian faith by adopting a strict doctrine of biblical inerrancy, wherein the entire text of the Bible was considered to be divinely inspired truth and without error (and usually, but not always, to be interpreted literally).
Open conflict between modernists and fundamentalists was suppressed by the drive for Prohibition and by World War I. But after the war, the populist politician William Jennings Bryan decided that “Darwinism” had been the cause of German militarism as well as a threat to traditional religion and morality (Marsden, 1991). In the early 1920s, he spearheaded a national crusade against the teaching of evolution in the public schools, which in the previous decades had become common in textbooks and thus in the curriculum (Grabiner and Miller, 1974). Bans on teaching evolution were passed in several states (Larson, 2003).
Bryan’s campaign peaked in the 1925 Scopes Monkey Trial in Dayton, TN, where he was humiliated on the stand by Clarence Darrow; he died a few days later. But although fundamentalism was discredited in the eyes of the media, Tennessee’s ban on teaching evolution was not overturned. Other states and many local jurisdictions enacted laws or policies that discouraged or forbade the teaching of evolution, and evolution rapidly disappeared from high school textbooks.
Evolution was not part of the precollege curriculum for 40 years, until fears of technologically falling behind the Soviet Union led in the late 1950s to federal money for new science textbooks—unusually for the time,
written by scientists (Larson, 2003). The Biological Sciences Curriculum Study’s series of biology textbooks reintroduced evolution. The Arkansas Education Association, concerned about teachers being caught between a state ban on evolution and district requirements to use textbooks that included evolution, challenged the state’s antievolution law. This suit resulted in the 1968 Supreme Court decision Epperson v. Arkansas (1968), which ruled that bans on teaching evolution were an unconstitutional favoring of the fundamentalist religious view.
“Creation science” arose on the national scene in the late 1960s as a counter to the reintroduction of evolution into the curriculum. The person largely responsible for its invention was Henry M. Morris, who declared, “Creationism is on the way back, this time not primarily as a religious belief, but as an alternative scientific explanation of the world in which we live” (Numbers, 2006). Morris’s creation science was his literal interpretation of Genesis (including a young Earth, global flood, and special creation of plants and animals) expressed in scientific terminology. Explicit references to the Bible were optional: Morris’s 1974 book Scientific Creationism (Morris, 1974) came in two versions, one with Bible quotes, and one without.
In 1972, Morris founded the best-known creation science organization, the Institute for Creation Research (ICR), now in Santee, CA, and served as its president until his retirement in 1996. Even after retirement, Morris continued to promote creation science until his death in 2006 at the age of 87. Morris and the ICR have spun-off or inspired other organizations promoting creation science, the most important of which is the Kentucky-based ministry Answers in Genesis. Answers in Genesis rivals ICR in size and influence, and plans to open in 2007 a 50,000-square-foot museum promoting a literal Genesis creation about 10,000 years ago. Dozens of smaller institutions and active independent creation science ministries, fundamentalist churches, and several television evangelists also contribute to the movement (Scott, 2004).
Despite its scientific veneer, creation science was ruled to be clearly religious and therefore unconstitutional to advocate in the public schools in the district court decision McLean v. Arkansas (1982) and the Supreme Court decision Edwards v. Aguillard (1987). As will be shown below, ID arose as a direct response to these defeats. However, even though ID recently has attracted more national media attention, partially as a result of the Kitzmiller v. Dover case (2005) where it too was ruled unconstitutional, creation science remains the larger of the two movements and generates much grass-roots activity.
INTELLIGENT DESIGN ARGUMENTS
The ID movement has its de facto headquarters at the Discovery Institute, a Seattle-based, policy-oriented think tank founded in 1990. In 1996, the Discovery Institute added ID to its agenda by opening the Center for Renewal of Science and Culture. In 2002, the words “Renewal of” were deleted from the name, producing the Center for Science and Culture, probably to appear more secular. As documented by Forrest and Gross’s examination of the “Wedge Document,” a fund-raising proposal prepared by staff at the Center for Science and Culture, Christian cultural renewal is precisely the goal of the ID movement (Forrest and Gross, 2004). Although the Discovery Institute has vociferously claimed that ID is a scientific research program and “not creationism,” in reality, many of the movement’s claims are derived directly from creation science with no modification. However, the ID movement promotes a few phrases and concepts that at first glance seem to be novel.
As defined by William Dembski (1999), the most prolific ID proponent:
Intelligent design is three things: a scientific research program that investigates the effects of intelligent causes, an intellectual movement that challenges Darwinism and its naturalistic legacy, and a way of understanding divine action.
Challenging the alleged “naturalistic legacy” of “Darwinism” and “understanding divine action” are not scientific endeavors, so the scientific component of ID defaults to the investigation of “the effects of intelligent causes.” It might seem, then, as though ID is intended to be a contribution to psychology, ethnology, or archaeology, all scientific fields that involve the effects of uncontentiously intelligent causes. Not so: ID is conspicuously absent from the scientific literature of those fields, as indeed from the scientific literature in general. In any case, ID’s proclaimed goal is significantly more ambitious: to detect intelligent design in nature.
ID proponents claim to be able to distinguish complex things that are the result of unintelligent causes and those that are the result of intelligent causes. The differentiation is supposedly accomplished through a variety of approaches; the two most popular being “irreducible complexity,” promoted by biochemist Michael Behe in Darwin’s Black Box (1996), and Dembski’s “specified complexity,” which leads to a “design inference” (Dembski, 1998).
Behe defines an irreducibly complex structure as one with many components, all of which must be in place for the structure to function. He typically illustrates the concept with a mousetrap, which requires the simultaneous presence of a spring, bar, platform, and some other parts to
catch a mouse, but his favorite biological example is the bacterial flagellum. Behe contends that all of the more than 40 different proteins that make up the flagellum must be present for the flagellum to function. He then infers that the incremental process of mutation and selection, requiring a selective benefit at each step of construction, cannot (or is extraordinarily unlikely to) produce such a system, “because any precursor to an irreducibly complex system that is missing a part is by definition nonfunctional” (Behe, 1996). Instead, the functional system must have been produced all at once, as a “purposeful arrangement of parts,” much like a watch or any other human-designed machine. Hence, irreducibly complex structures, like human machines, are the product of an intelligent agent, not natural selection.
Dembski’s “design inference” resembles Behe’s ID criterion, but Dembski’s arguments tend to be conducted at a high level of abstraction, ornamented with mathematical notation of dubious utility. In brief, Dembski contends that if a given event or object has a low probability of occurrence on all of the nondesign hypotheses available, then it exhibits what he calls specified complexity; specified complexity is, he argues, a reliable indicator of design. The only biological structure to which Dembski attempts to apply his method is the bacterial flagellum (Dembski, 2002). Noting that the chance of its parts assembling at random is astronomically low, and relying on Behe’s argument to exclude gradual evolutionary assembly, Dembski concludes that it was intelligently designed. Like Behe, however, he asserts that science is incapable of proceeding further to determine the nature of the designer or the means by which the design was instantiated.
The design inference and irreducible complexity consist of two components: an extensive negative argument against the plausibility of evolutionary explanations, and then a brief attempt at a positive argument relying on an analogy between biological adaptations and human artifacts. Behe’s negative argument against stepwise assembly of “irreducible” systems fails because it mistakenly assumes that evolution proceeds only by improvement of an extant function, whereas evolutionary theory extending back to Darwin has always emphasized the importance of changes of function in the origin of complex adaptations (Darwin, 1859b, 1862; Mayr, 1960; Jacob, 1977; Ganfornina and Sanchez, 1999; True and Carroll, 2002). The flagellum, although elucidated long after Darwin, is a useful case to examine. Contrary to the assertions of Behe and Dembski, a survey has shown that only 23 of the 42 proteins of the Salmonella typhimurium flagellum are universally required in bacterial flagella; and of those, 21 have already been found to have homologous related proteins that function in other, simpler biochemical systems (Pallen and Matzke, 2006). It is therefore not true that simpler precursors would be nonfunctional; they clearly
could have had different functions, just like the related systems in existence today. Deleting parts from a modern system does not simulate evolution in reverse, any more than decapitating modern vertebrates provides information about the origin of cephalization in early invertebrates.
The scientific criticisms of ID’s objections to evolution will not be treated in depth here, but it is important for scientists to be aware of and have ready reference to the most detailed scholarly critiques of the ID movement’s claims. These include rebuttals to the movement’s claims about the philosophy of science and the nature of science (Fitelson et al., 1999; Pennock, 1999; Smith, 2001; Peterson, 2002). Dembski’s argument inferring design from specified complexity, besides relying entirely on Behe’s argument for its application to biology, has been shown to rely on misconstruals of probability and information theory (Wilkins and Elsberry, 2001; Sober, 2002; Perakh, 2004; Shallit and Elsberry, 2004). The ID movement’s common claim that evolution cannot produce “new genetic information” is contradicted by numerous papers documenting the origin of new genes (e.g., Long et al., 2003) or even entire multiprotein catabolic pathways for artificial compounds that humans have released into the environment in recent decades (Copley, 2000; Johnson and Spain, 2003). Behe’s claim has been rebutted in general (Miller, 1999; Thornhill and Ussery, 2000; Matzke and Gross, 2006) and for specific complex systems such as bird wings (Gishlick, 2004), the vertebrate blood clotting cascade (Davidson et al., 2003), the vertebrate immune system (Bottaro et al., 2006), and the ID movement’s favorite system, the bacterial flagellum (Miller, 2003; Musgrave, 2004; Pallen and Matzke, 2006). Faced with such rebuttals, Behe and Dembski typically make the unsupported assertion that indirect pathways are highly improbable or, ironically, given the absence of any detail in their own explanation, complain that the proffered explanations lack sufficient detail to be empirically tested.
THE ARTIFACT ANALOGY AND DESIGN
The ID movement’s negative arguments against evolution are numerous, but its positive argument for design consists of variations on an analogy between biological systems and human artifacts. Behe and other ID proponents will often analogize the recognition of design in biology to the recognition of design in human-made artifacts, for example, Mount Rushmore. Behe writes, “unintelligent physical forces like plate tectonics and erosion seem quite sufficient to account for the origin of the Rocky Mountains. Yet they are not enough to explain Mount Rushmore” (Behe, 2005a). Intelligence is required to explain the purposeful arrangement of stone surfaces into the faces of four presidents. Similarly, ID proponents argue, when a biological structure exhibits a complex and purposeful
arrangement of parts, “intelligence” is the obvious explanation once evolutionary processes have been eliminated.
As a scientific explanation, this argument is exceedingly vague. The artifact analogy proposes that a structure like the flagellum “looks” designed, evolution cannot explain it, and therefore it is designed. The indicators of design are complexity and/or a “purposeful arrangement of parts.” But complexity is not a reliable marker of intelligent agency: A paperclip is also the product of an intelligent agent, but it is certainly not complex. The only observable designers, humans, seem to favor simplicity as often as complexity in their designs; simple designs are often more efficient to manufacture and use, and less prone to breakage and user error. On the other hand, even outside of biology any number of physical forces can produce complexity so extreme that it is far beyond the capacity of any known or even theoretical computer to precisely model; chaotic systems such as weather are examples.
ID proponents often present archaeology as an example of how scientists search for complexity, and thus by analogy, ID is a scientific field. Yet archaeologists are not seeking complexity in the discovery of human artifacts. When a stone implement is discovered, it may be highly complex, with many facets removed to produce a specific shape, such as an intricate, fluted Native American Folsom point. But a human artifact may also be quite simple—such as a unifacially flaked chopper from Olduvai Gorge made by early humans. What distinguishes both a Folsom point and an Oldowan chopper from unworked stone is not complexity but the different chipping patterns produced by human manufacture versus natural weathering, and perhaps most importantly, the context of the discovery. As Hurd explains, “We [archaeologists] have three sources of information: practical experience with the materials used, evaluation of objects in their context, and the commonality between contemporary behaviors and ancient behaviors” (Hurd, 2004).
The “purposeful arrangement of parts” criterion invites the question of just what the purposes are supposed to be for a given system. Human purposes are well known, and are reflected in the kinds of artifacts they design: cutting implements are devised for cutting, etc. But what is the purpose of the bacterial flagellum? When queried about this on the stand in Kitzmiller, Behe gave the unhelpful reply that the only purpose that could be inferred was that the designer wanted to make a bacterial flagellum. The artifact analogy, then, fails even at the first, definitional stage.
In actual scientific research, inferences of design are not made by using the vague criteria put forward by ID advocates. Archaeological artifacts or constructions like Mount Rushmore are recognized as having been designed (by humans) because a great deal of background knowledge is available about human design, including methods and motives. The man-
ufacture of stone tools has been observed and replicated, as has the design and manufacture of sculptures such as Mount Rushmore. In archaeology, real design events are reconstructed in detail, including the time, location, materials, tools, techniques, motivation, and culture that produced an artifact, and these, in addition to basic physical laws that humans must follow such as conservation of mass, result in a highly constrained explanatory hypothesis that is readily testable with additional data. ID offers none of this. It invokes an unidentified, unconstrained agent (the intelligent designer) who makes complex biological structures such as the bacterial flagellum for an unknown purpose, using unknown techniques and unknown materials. Even questions such as the time of origin and whether or not mass and energy were conserved remain unanswered. ID provides none of the information that we have about human artifacts and their creators that allow us to make the decision that a given object is artificial rather than occurring naturally (Wilkins and Elsberry, 2001).
ID proponents regularly analogize machines (truly “purposeful arrangements of parts”) with multipart molecular structures and processes. Yet, on inspection, such analogies break down. The differences between biological phenomena and human-built machines easily outweigh the superficial similarities. Machines and other artifacts serve human purposes, whereas biological designs serve only the ultimate “purpose” of self-replication. Machines made by humans consist of parts designed for the task; complex biological “machines” are always, upon investigation, found to be cobbled together from preexisting modules with other functions. Biological designs are not really “purposeful arrangements of parts,” they are really adaptations of parts originally used for some other purpose. Some differences are even more fundamental. As Woese (2004) notes, “The machine metaphor certainly provides insights, but these come at the price of overlooking much of what biology is. Machines are not made of parts that continually turn over, renew. The organism is.” Woese suggests that organisms are like eddies in a current, “resilient patterns in a turbulent flow—patterns in an energy flow” (Woese, 2004).
ID proponents contend that scientists reject ID for religious/ philosophical reasons, allegedly to promote a materialistic worldview (Johnson, 1991). But as this discussion shows, ID has been rejected for its scientific failings. Its negative arguments against evolution are based on a strawman version of evolutionary theory and ignorance about the data and the literature. Its positive argument approaches the problem of biological design from an erroneous premise of an inaccurate analogy. Living things may be composed of individual parts, and may be highly complex, but they are not artifact-like in any way that would help explain their origins. Scientists who have examined claims of ID reject it because ID does not adequately explain the natural world. Significantly, these
scientific criticisms of ID come both from scientists who believe in God, such as Kenneth R. Miller (1999), and those who do not, such as Richard Dawkins (2006).
But if ID is flawed on so many levels, why does it exist at all? The answer is found in its historical origins.
DESIGN IN CREATION SCIENCE
Long before the ID movement arose, creation scientists constantly invoked design arguments. Some deny this connection (Ratzsch, 2005), but an extensive 1989 survey (McIver, 1989) of creationist literature notes the ubiquitous role of design:
The venerable Argument from Design remains the chief weapon in creationist apologetics. Creationists consider it self-evident and incontrovertible. Although the theory of evolutionary adaptation stood the design argument completely on its head, creationists continue to appeal to Design without even a trace of defensiveness. It is featured in virtually every book or article promoting creation-science. “Actually,” says John Morris [(1989)], Henry Morris’s son, “any living thing gives such strong evidence for design by an intelligent designer that only a willful ignorance of the data (II Peter 3:5) could lead one to assign such intricacy to chance.”
Design as an argument against evolution has historically been a constant theme in creationist periodicals such as the Creation Science Research Quarterly. A cursory search shows that design arguments are invoked for tetrapod limbs (Davis, 1965), the yucca and its moth (Clark, 1965), the hummingbird (Keithley, 1977), and long lists of adaptations from across biology (Shute, 1965a,b). All of these examples of design use some version of Behe’s irreducible complexity argument, and even Behe’s mousetrap is presaged by numerous articles claiming design for the traps of carnivorous plants (Keithley, 1972, 1982; Howe, 1978). Even the bacterial flagellum, the iconic example of the ID movement, is found in the creation science literature before Behe promoted it (Anonymous, 1992; Lumsden, 1994). In fact, creation science leaders have criticized the ID movement for stealing their arguments.
Dembski often refers, for example, to the bacterial flagellum as a strong evidence for design (and indeed it is); but one of our ICR scientists (the late Dr. Dick Bliss) was using this example in his talks on creation a generation ago. And what about our monographs on the monarch butterfly, the bombardier beetle, and many other testimonies to divine design? Creationists have been documenting design for many years, going back to Paley’s watchmaker and beyond (Morris, 2005).
The concept of design thus is central to both creation science and ID. Although ID claims to be agnostic on much of creation science, such as the age of the Earth, Noah’s Flood, and the like, when it comes to design, creation science and ID speak in one language. This language is that of William Paley, whose argument from design in his 1802 Natural Theology proclaimed that structural complexity of biological organisms was evidence for the existence of God (Paley, 1802).
Like the irreducible complexity argument, the other prominent claims made by the ID movement, and often the specific terminology, trace back to creation science. “Specified complexity” entered the antievolution literature in Thaxton et al. (1984), in the midst of a chapter that attempted to repair the infamous creation science shibboleth, much ridiculed by scientists, that a decrease in entropy in biological systems contradicts the Second Law of Thermodynamics. The authors grudgingly conceded that local decreases in entropy were not prohibited in open systems like the earth, which experience a continuous energy flow, but claimed that genetic information exhibits specified complexity, and that thermodynamic limitations block any nonintelligent increase in information. More generic “no new information” arguments had been made by the European creation scientist A. E. Wilder-Smith, who has been repeatedly cited as an inspiration by many ID proponents (Touchstone, 2004). Other ID arguments, such as the claim that there are no transitional fossils in the fossil record or that “microevolution” is proven but “macroevolution” is dubious, are indistinguishable from those in the creation science literature (Matzke and Gross, 2006).
The microevolution/macroevolution distinction is particularly revealing. In evolutionary biology, microevolution refers to evolutionary processes operating within a species. Although scientists sometimes colloquially refer to macroevolution as “evolution above the species level,” this definition does not do justice to the complexity of topics included within the concept. Macroevolution refers to patterns that emerge as species and lineages branch through time, including the rate and pace of evolutionary change, adaptive radiation, morphological trends in lineages, extinction or branching of a lineage, concepts such as species sorting, and the emergence of major new morphological features (such as segmentation, or shells, or the fusion or loss of bones). Decades ago, creationists began to use microevolution and macroevolution idiosyncratically. Creationists’ use of “microevolution” is not dissimilar to that of evolutionary biologists, although they apply it not just to species but to evolution within the limits of a specially created “kind” of organism. When ID supporters and other creationists claim to accept some evolution, they generally mean it in this limited sense of evolution “within the kind.” A larger distinction occurs in the creationist definition of macroevolution, which to them
refers to (unacceptable) common ancestry of different created kinds. It also refers to the acquisition of major morphological features or body plan changes, also considered impossible without the direct involvement of God. Both creation science and ID approach the micro/macro divide similarly: microevolution is accepted, and macroevolution (their definition) is rejected.
SPECIAL CREATION
The conservative Christian theological doctrine of special creation is central to creation science. Special creation insists on the creation of natural phenomena in their present form, although variations occur. The most extreme special creationists believe that the entire universe (galaxies, stars, the earth, and living things on the earth) was created essentially as we see it today, with only limited change since the Creation. Young-earth creationists such as Henry Morris accept such a view. Various schools of old-earth creationism accept cosmological evolution, but all reject biological evolution. For them, God specially creates organisms intermittently over the millions of years of the earth’s history.
The idea of specially created “kinds” of organisms derives from the Book of Genesis:
And God made the beast of the earth after his kind, and cattle after their kind, and everything that creepeth upon the earth after his kind: and God saw that it was good (Genesis 1:25, King James version).
For conservative Christians who believe that every word of the Bible is inerrant truth, biblical “kinds” are highly significant, because the language is plain and the phrase is repeated again and again in Genesis. “Kinds” have enough genetic variability to adapt to local conditions, but adaptation is strictly limited to the boundaries set by God; because kinds are specially created, common ancestry between created kinds is impossible by definition. Creationists have made efforts to discern the limits of the created kinds, but applying the doctrine to profligate biological diversity has proven difficult. According to Duane Gish, a biochemist who recently retired from the ICR, a “kind” might correspond to a whole phylum in the case of invertebrates, a family for some vertebrates, or a species in the case of humans (Gish, 1985).
The denial of common ancestry is unsurprising in creation science, but it is a common misconception that ID advocates accept common ancestry and “macroevolution.” In fact, the vast majority of ID proponents deny the common ancestry of humans and apes. Behe is the only significant exception, although he is much-touted by those who wish to
portray ID as a moderate position. Even Behe’s support is lukewarm; in 2005, he wrote that “my Intelligent Design colleagues who disagree with me on common descent have greater familiarity with the relevant science than I do” (Behe, 2005b). Dembski’s position is typical, accepting “some change in the course of natural history,” but believing “that this change has occurred within strict limits and that human beings were specially created” (Dembski, 1995). This is the standard position of an ID advocate. In May 2005, ID supporters on the Kansas Board of Education held hearings to support ID-friendly science standards. Mainstream scientists boycotted the hearings, but a series of pro-ID witnesses, mostly teachers and academics (but few professional biologists) testified in support of the standards. During cross-examination, only 2 of 19 witnesses accepted the common ancestry of humans and apes. One was an independent scholar who clarified that although he supported the Kansas standards, he was not an ID advocate; and the other was Behe. The rejection of evolution by the vast majority of ID witnesses at the Kansas hearings parallels the rejection of evolution by ID proponents in general.
THE EMERGENCE OF INTELLIGENT DESIGN
Although the content of ID suggests that it is derived from creation science, the recently uncovered historical origin of ID illustrates this even more clearly.
The creation science movement reached its peak in the early 1980s. Equal time for evolution and creation science bills were proposed in at least 27 states in 1980 and 1981 (Scott, 2004). Arkansas and Louisiana passed laws mandating “equal time” for the “two models” of evolution and creation science. Arkansas Methodist minister Bill McLean and other plaintiffs, most of them professional clergy from various Christian denominations, brought suit against their state’s equal time law in federal district court, and the trial was held in December 1981. McLean v. Arkansas pitted a team of plaintiffs’ witnesses that included eminent scientists such as Francisco Ayala, Stephen Jay Gould, Harold Morowitz, and G. Brent Dalrymple against a team of creationist defense witnesses who were largely unknown in the world of science, and who had the impossible task of defending the scientific merits of a young earth and global flood.
McLean put creation science on trial, and creation science lost badly. In the January 1982 decision, the judge wrote that creation science was biblical literalist Christianity in disguise, and that to teach it would be to promote a sectarian religious view, which he held to be unconstitutional under the First Amendment of the Constitution (1982). The judge in McLean also noted a characteristic of creation science he termed a “contrived dualism” wherein evidence against evolution was considered to be
evidence supporting special creation. This was inadequate for a proper scientific explanation and “has no scientific factual basis or legitimate educational purpose.”
Even conservative Christians recognized that creation science had been a legal disaster. Christianity Today editorialized, “Evangelicals are appalled at the adverse publicity given biblical faith by the public media as a result of the recent creation/evolution trial in Arkansas” (Kantzer, 1982). The fundamentalist Moody Monthly published a story asserting that Arkansas was “Where Creationism Lost its Shirt” and, despite being squarely behind creation science, concluded that the problem with the creation science witnesses had been the lack of published research supporting creation (Mawyer, 1982).
Despite their loss in Arkansas, the creationists had high hopes for the parallel bill enacted in Louisiana. The Louisiana bill was drafted with more deliberation and was more vague about the tenets of “creation science,” leaving out explicit mention of the young earth and global flood. Furthermore, the state of Louisiana deputized the creationist lawyer Wendell Bird, ensuring that a highly motivated expert would defend the law from the inevitable American Civil Liberties Union challenge (Larson, 2003).
In the midst of the 1981 wave of creation science legislation and litigation came the first hints of the book that would later introduce the world to ID. The Fall 1981 issue of a creationist student newspaper carried the front-page headline, “Lawsuit prospects dim in Arkansas, bright in Louisiana.” Below the main story was a short announcement for a “high school biology textbook” that would “present both evolution and creation while limiting discussion to scientific data” (Anonymous, 1981). Those interested in the project were urged to contact the Foundation for Thought and Ethics (FTE). FTE is a self-described “Christian think tank” located in Richardson, Texas. It was founded in December 1980 by Jon Buell, who had previously worked at the old-earth creationist Probe Ministries, also in Richardson. A document filed with the IRS in 1981 entitled “What is the Foundation for Thought and Ethics?” declared that:
The Foundation for Thought and Ethics has been established to introduce biblical perspective into the mainstream of America’s humanistic society, confronting the secular thought of modern man with the truth of God’s Word.
FTE described two projects in the works to carry out its goals.
[O]ur first project is a rigorous scientific critique of the theory of prebiotic evolution. Next, we will develop a two-model high school biology textbook that will fairly and impartially view the scientific evidences for
creation side by side with evolution. (In this case Scripture or even religious doctrine would violate the separation of church and state.)
The first project materialized as The Mystery of Life’s Origin (Thaxton et al., 1984), written by Charles B. Thaxton, Walter Bradley, and Roger B. Olsen, three conservative evangelicals who accepted an old earth but who were firmly against a natural origin of life or any substantial biological evolution. The book presented the problem of the origin of life as a scientifically unsolvable mystery and, in a postscript, endorsed divine creation as a better answer. Although ID proponents point to The Mystery of Life’s Origin as being the foundational publication for the movement that came to be called intelligent design, it was just one of many books written in the early 1980s that represented attempts by believers in biblical inerrancy to develop a creationist science that avoided the pitfalls of more traditional creation science, such as hostility to an old earth (Pun, 1982; Wiester, 1983; Lester et al., 1984; Pitman, 1984).
Much as had the creation scientists, the authors of The Mystery of Life’s Origin proposed that the origin of life was not simply an extraordinarily difficult problem upon which the research community had not yet reached consensus. Instead, it was a problem that was categorically unsolvable by appeal to natural causes: The first cell was simply too complex to have been produced through natural—equated with “chance” or unguided—processes. Dean Kenyon, then a biologist at San Francisco State University, wrote in the introduction, “… it is fundamentally implausible that unassisted matter and energy organized themselves into living systems” (Thaxton et al., 1984). The authors proposed that in the absence of any possible natural causes, the origin of life must therefore be the result of intelligent agency. The agent, they hastened to add, did not have to be God: it could be, perhaps, an intelligent alien.
Even before The Mystery of Life’s Origin was published, FTE’s Buell had begun work on the second project mentioned above: the “two-model high school biology textbook.” This was published in 1989 as Of Pandas and People, later described by Buell as “the first place where the phrase “intelligent design’ appeared in its present use” (Buell, 2004). (Buell’s remark occurs in his preface to the third edition of Of Pandas and People, temporarily available on Dembski’s web site in 2004.) Credit for authorship was given to Percival William Davis and Dean Kenyon. Davis was described as a biology instructor, and Kenyon as a biology professor. These descriptions, while true, left unsaid the fact that both were traditional creation scientists. Davis was the coauthor of a creationist book (Frair and Davis, 1967) and articles in the Creation Research Society Quarterly (Davis, 1965; Howe and Davis, 1971). Kenyon in 1981 had been scheduled as a defense (i.e., creation science) witness in the McLean trial (although he
did not testify). He was to be Wendell Bird’s lead expert in the Louisiana litigation, Edwards v. Aguillard. He also had authored several forewords for creationist books and stated in interviews that he believed there were “no errors in the Bible,” that “10,000 to 20,000 years ago—the entire cosmos was brought into existence out of nothing at all by supernatural creation,” (Salner, 1980) and that he had converted to scientific creationism after reading books by Wilder-Smith and Henry Morris (Pearcey, 1989). Two unacknowledged authors of Of Pandas and People are of interest: Nancy Pearcey, author of the Overview chapter, was another young-earth creationist and an editor at the Bible-Science Newsletter, and Michael Behe wrote much of the chapter on biochemistry for the second edition, published in 1993.
The history of the writing of Of Pandas and People illustrates the creationist roots of ID. During the course of the Kitzmiller v. Dover trial, early manuscript versions were subpoenaed by the plaintiffs’ legal team and introduced into evidence as exhibits. By comparing these manuscripts, it was possible to document a transition from creationist to ID terminology during the preparation of the book. The titles of the manuscripts changed over time: in order, the early manuscripts (numbers in parentheses refer to court exhibits from the Kitzmiller v. Dover trial, on file at the National Center for Science Education) are titled Creation Biology (1983) (P-563), Biology and Creation (1986) (P-560), and Biology and Origins (1987) (P-561). In 1987, the title was changed to Of Pandas and People; there were two 1987 (1987-1: P-562; 1987-2: P-652) manuscripts with this title. In 1989, the first edition was published by a small Dallas publisher (Davis et al., 1989), and in 1993, the second edition appeared (Davis et al., 1993).
On June 19, 1987, the Supreme Court decided in Edwards v. Aguillard that teaching creation science was unconstitutional. Although Wendell Bird argued strenuously before the Court that Kenyon’s expert witness affidavit showed that creation science was scientific and nonreligious, the justices voted 7–2 that supernatural creation was a religious view and that the Louisiana legislature had violated the Establishment Clause by promoting it in public schools. Creation science as a legal strategy was no longer viable.
The Pandas manuscripts reflect this important legal decision. During the Kitzmiller case, word counts for the terms “creationist” or cognates, and for the phrase “intelligent design” were compared across the manuscripts. When graphed, it becomes clear that one set of terminology was substituted for the other, with the change taking place between the two 1987 manuscripts (Fig. 15.1).
Another comparison, this time of a key sentence defining creationism, similarly illustrates the substitution of “intelligent design” for cognates
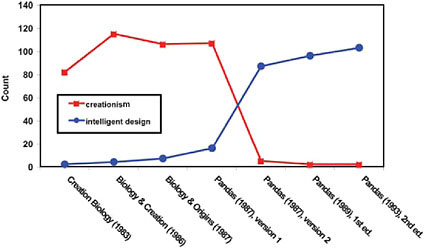
FIGURE 15.1 A comparison of phrasing in the prepublication manuscripts of the ID textbook Of Pandas and People. Early manuscripts freely used cognates of “creation” (creationism, creationist), but these terms were replaced by the phrase “intelligent design” after the mid-1987 Edwards v. Aguillard Supreme Court decision outlawing the teaching of creationism.
of creationism. In the 1986 manuscript Biology and Creation, a paragraph appears that reads:
Creation means that the various forms of life began abruptly through the agency of an intelligent creator with their distinctive features already intact—fish with fins and scales, birds with feathers, beaks and wings, etc. (emphasis added) (p. 2-10)
This paragraph is repeated with only small changes (capitalization or punctuation) in early manuscripts, and appears also in the first of the two 1987 drafts titled Of Pandas and People:
Creation means that various forms of life began abruptly through the agency of an intelligent Creator with their distinctive features already intact—fish with fins and scales, birds with feathers, beaks and wings, etc. (emphasis added) (pp. 2-14, 2-15)
In the second 1987 manuscript, the paragraph’s wording has been changed:
Intelligent design means that various forms of life began abruptly through an intelligent agency with their distinctive features already intact—fish with fins and scales, birds with feathers, beaks and wings, etc. (emphasis added) (p. 2-15)
Also introduced into evidence during the Kitzmiller case was chapter six of the manuscript for the third edition of Of Pandas and People, in preparation, also to be published by FTE, which will be given the new title, The Design of Life (P-775). This edition has new authors: William Dembski and fellow ID proponent Jonathan Wells. Perhaps considering that “intelligent agency” (or any creative source embodied by an agent) might imply creation by God, the newest version proposes an agent-free form of creationism:
Sudden emergence holds that various forms of life began with their distinctive features already intact, fish with fins and scales, birds with feathers and wings, animals with fur and mammary glands. (emphasis added)
When these comparisons were presented during the Kitzmiller trial, they had a powerful effect. In his decision, the judge called this blatant switch of terminology “astonishing.”
The manuscript drafts also preserved a further piece of evidence of the evolution of creationism into ID, although this was not presented in trial. A textual transitional fossil was discovered by Kitzmiller expert witness Barbara Forrest during her study of the trial exhibits. Shown in Fig. 15.2 for comparison are two excerpted passages from the two 1987 drafts. Evidently, the editor of the drafts was deleting the word “creationists” and inserting the phrase “design proponents” throughout the document. During the tedious procedure, the editor evidently forgot to delete the “c” and “ists” from the word “creationists.”

FIGURE 15.2 The missing link between creation science and intelligent design. In the early 1987 manuscript (a) of Of Pandas and People, the original wording of a sentence in chapter 3 reads, “Evolutionists think the former is correct, creationists accept the latter view.” In the second 1987 manuscript (b), an incomplete (and uncorrected) block-and-paste of “design proponents” for the term “creationists” leaves “cdesign proponentsists” (sic), forming a missing link between creationism and ID.
In conclusion, examination of both the history and content of ID shows that it is a form of creationism, despite the persistent efforts of proponents to obscure this connection. Creation science was struck down because teaching it would be a form of religious advocacy. ID was invented as a way to circumvent the constitutional barrier to creation science, but when the constitutionality of ID was tested in Kitzmiller, it met the same fate (2005). However, unlike a Supreme Court decision, a district court decision such as Kitzmiller only sets a local precedent, and future attempts to incorporate ID in public school curricula are likely. However, because the Kitzmiller opinion was so thorough and powerful, it will undoubtedly discourage communities that may be contemplating ID policies. But just as creation science continued after McLean v. Arkansas, so ID will continue after Kitzmiller, even if in a reduced form.
INTELLIGENT DESIGN AND EDUCATIONAL POLICY
Despite its scientific shortcomings, the ID movement should be taken seriously because it has been disquietingly effective in reinforcing the sentiment, originally exploited by proponents of creation science, that evolution is inadequate science and that creationism is a valuable approach that students deserve to learn about in public school science classes. Recent survey data from the United States and foreign countries indicate that the United States is distinctive among developed nations for its unusually low level of acceptance of evolution (Miller et al., 2006). Because evolution is rejected by so many, and because American education is highly decentralized and unusually politicized, it is not surprising to learn that evolution is under attack in many communities around the country. Such attacks take one or both of two forms: efforts to promote creation science or ID, and efforts to compromise or reduce the teaching of evolution.
The National Center for Science Education collects data on controversies over evolution education in the United States. Although these data are possibly incomplete, it is apparent that the country has experienced another wave of antievolutionism at the state level since the late 1990s. States have been revising their science education standards in response to the No Child Left Behind Act’s mandate requiring students to be tested in science beginning in 2007. Between 2000 and 2006, the National Center for Science Education has monitored conflicts over the treatment of evolution in state science standards being developed or revised in Alaska, Arizona, Alabama, Georgia, Hawaii, Kansas, Minnesota, New Mexico, Nebraska, Ohio, Pennsylvania, North Carolina, South Carolina, and West Virginia.
The National Center for Science Education has also monitored attempts to undermine the teaching of evolution by state legislatures. In 2006 alone, legislation was introduced in nine states that would have either promoted
creationism/ID or inhibited the teaching of evolution. Most of these bills did not pass, due to action by citizens, including scientists, who persuaded legislators to vote against the bills. The states included (HB for House Bill, and SB for Senate Bill) Alabama (HB 106, SB 45), Indiana (HB 1388), Michigan (HB 5606, HB 5251), Missouri (HB 1266), Mississippi (HB 953, SB 2427, HB 214), Oklahoma (SB 1959, HB 2526, HB 2107, HB 1003), Utah (SB 96), New York (AB 8036), and South Carolina (SB 114). In Mississippi, a watered-down version of a bill appended to another bill did pass (HB 214, appended to SB 2427). This bill originally called for the teaching of “flaws or problems” in evolution and encouraged the teaching of ID. The final, reduced bill provided that “No local school board, school superintendent or school principal shall prohibit a public school classroom teacher from discussing and answering questions from individual students on the origin of life.”
Although there are still rare attempts to promote creation science at the state level, most of the school board or legislative antievolutionism today is directed toward promoting ID and/or promoting the teaching of alleged “evidence against evolution.” The latter strategy consists of taking the creationist objections to evolution and stripping them of any mention of a positive explanation of biology, such as creation or design. By avoiding explicit or implied reference to God or a Designer, creationists hope to survive constitutional challenges.
A number of phrases are being used to promote this “evidence against evolution” approach, including requiring students to “critically analyze evidence for evolution,” to learn “both evidence for and evidence against evolution,” to study “both the strengths and weaknesses of evolution,” or to have evolution presented as “theory not fact.” Teachers are also exhorted to “teach the full range of views about origins” and, in the slogan of the Discovery Institute, “teach the controversy.” The vagueness of “teach the controversy” is its strength: The public is told by media sources that evolution is socially controversial and infers that evolution is also controversial among scientists. “Teach the controversy” does not mean that teachers should have students debate actual controversial scientific issues; it is rather an exhortation to teachers to instruct students that common ancestry (evolution) is a serious issue of contention among scientists.
Antievolutionists have also proposed policies and legislation that contend that it is the students’ right to know and the teacher’s right to teach creation science, intelligent design, or “evidence against evolution.” Such “fairness” arguments resonate with the American public, which responds to the cultural attractiveness of hearing all sides to an issue but which by and large fails to understand that there is no serious scientific challenge to evolution.
ID therefore is making a serious challenge not in the world of science, but in the world of public educational policy. It aims to be a “big tent” presenting a minimalist form of creationism on which all creationists can agree (Scott, 2001), focusing on the supposed impossibility of the natural origin of biological complexity. In addition to its unsuitability for the public school classroom because of its promotion of a sectarian religious position, ID is also a failure as science and has not earned the right to be taught in precollege classrooms. For all its opportunistic use (and misuse) of recent biological discoveries, ID offers only a premodern and impoverished perspective to explain complex functional biological phenomena, a perspective different indeed from the fertile and unifying evolutionary principles underlying the field of evolutionary biology.
ACKNOWLEDGMENT
We thank Glenn Branch for very useful comments on both content and style.




















