
13
Climatic Acme Events in the Sea and on Land
BILAL U.HAQ
Woods Hole Oceanographic Institution
INTRODUCTION
Major climatic events in the geologic past have left a variety of retrievable records in the sediments, both on land and sea. Some marked fluctuations in the climates are evident from widely distributed sedimentological, geochemical, and paleontological features. Here we are mainly concerned with the paleontological record of widespread latitudinal domination of certain floral and faunal elements during times of extreme climatic change. Numerous intervals during which biogeographic patterns show poleward expansions of tropical assemblages, or equatorward extensions of cold, higher-latitude assemblages, have been documented for the Cenozoic. These events have been informally termed climatic acme events (indicating peak climatic change) and have been used successfully in the reconstruction of paleoclimate. This paper describes and illustrates the concept of climatic acme events with examples from the Cenozoic marine and terrestrial record and, where available, gives evidence from the oxygen isotope record that corroborates the paleontological conclusions. The usefulness of acme events in biostratigraphic resolution of higher latitudes is also discussed.
The concept of acme events was informally introduced by Haq and Lohmann (1976). The maxima of assemblage migrationary cycles were termed “acme horizons” and defined as the levels in relatively continuous sequences characterized by the maximum abundance of an environmentally sensitive assemblage. If one assumes that times of extreme environmental (climatic, oceanographic) change are, geologically speaking, contemporaneous over large geographic areas, then the acme events will define synchronous horizons. Haq and Lohmann (1976) suggested that the precision with which acme horizons approximate synchronous surfaces depends on our ability unambiguously to identify the acme events (maximum geographic shifts of assemblages). This is affected both by the complexity of the migrationary cycles and by resolution of the method used to delineate them.
The recognition of acme events involves the use of paleobiogeographic data to delineate the cycles of latitudinal migrations of assemblages in stratigraphic sequences over wide geographic areas. The method has been described by Haq and Lohmann (1976) and further discussed by Haq et al. (1977) and Haq (1980). It essentially involves the following steps: (1) census of a particular fossil group from relatively continuous
sequences in sediment samples in which the fossils are relatively well preserved; (2) standardization and reduction of data by means of an appropriate quantitative analytical technique, which reduces the raw census data into few “factors” (assemblages) that explain a large share of the variance in the data matrix; and (3) delineation of the distribution patterns of the quantitatively defined assemblages on a time-space grid and recognition of individual migrationary events.
CENOZOIC EXAMPLES
In a series of recent studies, the Cenozoic biogeography of calcareous plankton (nannoplankton and planktonic foraminifera) has been delineated for the Atlantic Ocean (Haq and Lohmann, 1976; Haq et al., 1977; Haq, 1980). Numerous latitudinal migrations of assemblages have been recognized in both the Paleogene and the Miocene. Examples of some of the more prominent acme events are included below, and the results are compared to oxygen-isotopic data, where available.
Figure 13.1 shows the nannofloral and planktonic foraminiferal migrationary patterns recognized by Haq et al. (1977) in the North Atlantic Paleogene [65–25 million years ago (Ma)]. The most prominent acme events that are evident from both the nannofossil and the foraminiferal migrationary patterns are described below.
-
The shift during the middle Paleocene (60–57 Ma) of both high-latitude nannofloral assemblages to low latitudes and of mid-latitude “low-spired subbotinid” foraminiferal assemblage to low latitudes is in response to a marked cooling at this time.
-
A shift during the latest Paleocene-early Eocene (53–49 Ma) of low-latitude nannofloral assemblages to high latitudes combined with a withdrawal of high-latitude assemblages, combined with a maximum incursion of low-latitude morozovellid foraminiferal assemblage to mid-latitudes, as well as the mid-latitude “low-spired subbotinid” assemblage to higher latitude also takes place as a result of a peak warming.
-
A middle Eocene (46–44 Ma) incursion of higher-latitude assemblages into lower latitudes in response to a cooling, when high-latitude nannofossil assemblages once again return to temperate and tropical areas and the high-latitude globigerinid assemblage makes its first prominent appearance in mid latitudes.
-
A second incursion of the globigerinid assemblage into middle and lower latitudes in the late Eocene-early Oligocene (38–35 Ma) indicates a marked cooling at this time. Nannofossil data are lacking in latest Eocene, but early Oligocene data show a sharp incursion of high-latitude assemblages into low latitudes at 35 Ma.
-
A third incursion of the high-latitude globigerinid assemblage into lower latitude occurs in the middle Oligocene (32–31 Ma). This cooling event is also indicated by the migration of high-latitude nannofossil assemblages into low latitudes at this time. This acme event seems to have been at least equal in intensity to the latest Eocene-early Oligocene event of marked climatic deterioration between 38–35 Ma.
Supporting evidence for most of these paleoclimatic conclusions based on acme events of calcareous plankton comes from other independent sources. On land, the early Eocene acme event, indicating peak warming, manifests itself in a major expansion of tropical-subtropical land plants as far north as 40–45° N latitude (Wolfe, 1978) on the west coast of North America and in the presence of subtropical floral elements as far north as 60° N in the early Eocene in the Gulf of Alaska borderlands area, based on a revised age estimate (see Chapter 16). In general, land plants indicate significantly higher mean annual temperatures in the northern hemisphere and low-latitudinal temperature gradients in the early Eocene (Wolfe, 1978).
The evidence from land plants also corroborates the late Eocene cooling episode: the North American flora indicates that the mean annual temperatures on land were significantly lower than in early Eocene, and, at the same time, mean annual range of temperatures increased dramatically (Wolfe, 1978),.supporting a scenario marked by climatic deterioration. Sporomorph evidence from the southeastern United States (Gulf of Mexico and upper Coastal Plains) also suggests a rapid and marked climatic decline near the end of the Eocene, when climates became cooler and drier (Frederiksen, in press). This regime persisted into the early Oligocene,
Oxygen isotopic data from various parts of the ocean also lend support to the marked climatic changes indicated by the Paleogene calcareous plankton temporal and spatial migrations (see Figure 13.2). A decrease in benthic isotopic temperature in the middle Paleocene has been documented by Boersma and Shackleton (1977) from Deep Sea Drilling Project (DSDP) Site 357 on the Rio Grande Rise (South Atlantic) and by Boermsa et al. (1979) from DSDP Site 384 (western North Atlantic). At the latter site the planktonic foraminiferal (morozovellid) data also show low isotopic temperature in the middle Paleocene between 61–60 Ma. Buchardt (1978) recorded similarly low temperatures in the middle Paleocene based on oxygen isotope analyses of mollusc shells from north-west Europe.
The latest Paleocene-early Eocene climatic amelioration has been documented in numerous oxygen isotopic studies (Figure 13.2). Shackleton and Kennett (1975) recorded high isotopic temperatures based on both planktonic and benthic foraminifera in southern high-latitude DSDP Site 277 on Campbell Plateau—the temperatures recorded by them were highest in the entire late Paleocene to Recent stratigraphic interval. Buchardt’s (1978) molluscan oxygen isotopic curve shows a sharp rise in paleotemperature of shallow marginal seas in northwest Europe in the late Paleocene-early Eocene. Similar trends of isotopic temperature elevation have also been recorded in the Late Paleocene-early Eocene sequences at DSDP sites 398 and 401 in the North Atlantic by Vergnaud-Grazzini et al. (1978) in both the benthic and planktonic foraminifera.
All the data cited above bear on the conclusion that the latest Paleocene-early Eocene represents the warmest period of the entire Cenozoic in the marine realm. On land, mean annual temperatures were also considerably higher than at present (Wolfe, 1978). This peak climatic amelioration also seems
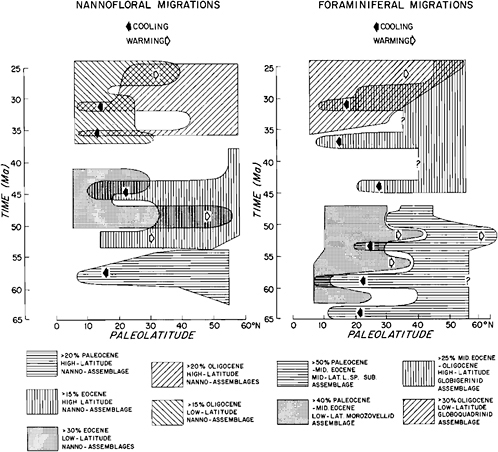
FIGURE 13.1 A summary of the major nannofloral and foraminiferal migrationary patterns in the North Atlantic Ocean through the Early Cenozoic. Migrations toward higher latitudes are interpreted as being caused by climatic warming and toward lower latitudes by climatic cooling. The patterns delineated enclose all samples that contain abundances greater than those indicated in the legend. Arrows indicate the direction of the major shifts of assemblages. Major and minor nannofloral assemblages with similar latitudinal preferences have been combined to obtain composite patterns in some cases. (From Haq et al., 1977, with permission of the American Geophysical Union.)
to have triggered higher evolutionary turnover in phytoplankton and foraminifera, when both nannoplankton and planktonic foraminifera show high evolutionary rates (Berggren, 1969; Haq, 1973), culminating in a peak in pelagic diversity in the middle Eocene (Fischer and Arthur, 1977).
The other nannofloral acme events observed in the Atlantic Ocean are also supported by oxygen isotopic data to varying degrees, The later middle Eocene cooling has been recorded at DSDP site 277 (Shackleton and Kennett, 1975), DSDP site 357 (Boersma and Shackleton, 1977) and in northwest Europe (Buchardt, 1978). The late Eocene-early Oligocene cooling event has been particularly well documented in the oxygen isotopic record. At DSDP site 277 a sharp drop in both planktonic and benthic isotopic temperature is evident (Shackleton and Kennett, 1975); at DSDP sites 398 and 401 (North Atlantic) a marked cooling trend has been recorded (Vergnaud-Grazzini et al., 1978) and the northwest European data show a similar precipitous drop in paleotemperature in the late Eocene-early Oligocene interval (Buchardt, 1978). The slight offset in the timing of this event between DSDP site 277
(Shackleton and Kennett, 1975) and elsewhere is most probably due to the relatively less accurate biochronologic resolution capability because of the high-latitude location of DSDP site 277. The mid-Oligocene cooling event is not so obvious in the isotopic record, which shows relatively low-amplitude variations during most of the Oligocene.
Figure 13.3 summarizes the nannofloral migrations in the North Atlantic in response to climatic acmes during the Miocene (Haq, 1980). Most of the biogeographic “activity” is confined to the mid to high latitudes, which show the most distinct migrationary patterns. This record shows four cooling events, alternating with four warming events of varying in-
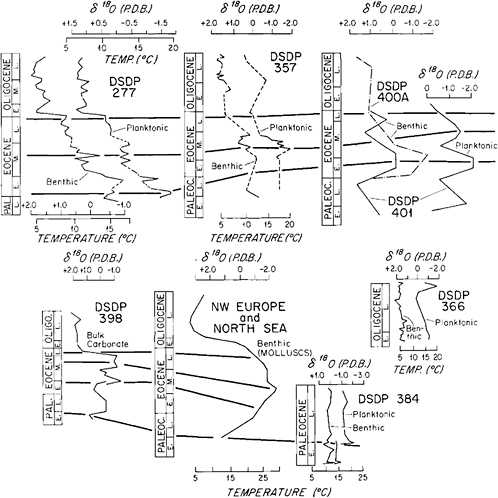
FIGURE 13.2 Paleogene oxygen isotope stratigraphy of DSDP sites from published sources. Planktonic and benthic foraminiferal δ18O curves from DSDP sites 277 (Shackleton and Kennett, 1975); 357 (Boersma and Shackleton, 1977); and 398, 400, and 401 (Vernaud-Grazzini et al., 1978); generalized mollusc δ18O curve from northwest Europe (Buchardt, 1978); planktonic and benthic foraminiferal curves from Paleocene of DSDP site 384 (Boersma et al., 1979); and Oligocene planktonic and benthic curves from DSDP site 366 (Boersma and Shackleton, 1978). Lines are drawn through the climatic acme events that have been identified through calcareous plankton migrationary patterns (Haq et al., 1977).
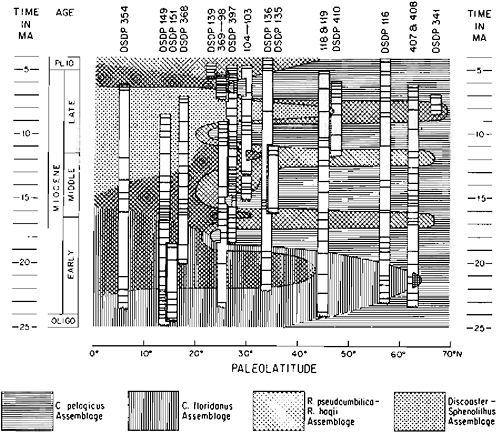
FIGURE 13.3 A summary of nannofloral migrationary patterns in the North Atlantic Ocean through the Miocene. DSDP cores are represented by the appropriate site number, and sample levels in these cores are represented by lines. The Miocene record shows four climatic warming episodes as indicated by low- and mid-latitude assemblage incursions into higher latitudes, and four cooling episodes are indicated by expansion of Coccolithus pelagicus (high latitude) assemblage into lower latitudes. (After Haq, 1980.)
tensity and duration. Earliest Miocene is generally cooler, when little or no biogeographic activity is observed and a stress-adapted, cosmopolitan assemblage dominates most latitudes. The first warm acme occurs between 22–20 Ma, with a peak at about 21 Ma. The second warming event occurs between 17–15 Ma, when mid-latitude nannoflora expands into higher latitudes. This is followed by a generally cooler interval up to 12.5 Ma, to be followed by a second incursion of mid-latitude nannofloral element into higher latitudes, indicating a warming between 12.5–11.5 Ma with a peak around 12 Ma. The third expansion of mid-latitude assemblage into higher latitudes occurs between 9–7.5 Ma with a peak around 8 Ma. The late Miocene interval, after this last warming, is characterized by cool climates, and high-latitude nannofloral elements are found as far south as 25° N.
Some of these Miocene acme events also manifest themselves on land, as indicated by the terrestrial plant record from the Pacific Northwest and Alaska (see Chapter 16). A temperature rise in the middle Miocene is indicated between 17–15 Ma, which is followed by a cooling, and then a renewed warming at 8 Ma. The terrestrial record also supports the suggestion that the latest Miocene was a generally cool interval.
The oxygen isotopic record shows trends that are in general agreement with the nannofloral record (Figure 13.4). The warming trend between 22–20 Ma can be observed in the planktonic and benthic isotopic record from the South Pacific, high-latitude DSDP site 179 (Shackleton and Kennett, 1975), from the benthic and planktonic record at North Atlantic DSDP sites 398 (Vergnaud-Grazzini et al., 1978) and 116 (C. Vergnaud-Grazzini, University of Pierre and Marie Curie, Paris, personal communication), and in the record at South Atlantic DSDP site 357 (Boersma and Shackleton, 1977). The most obvious and the warmest Miocene event is a marked decrease in δ18O values at the early-middle Miocene boundary (shown by a line through the peaks in various oxygen isotope curves reproduced in Figure 13.4). This event corresponds also to the first incursion of mid-latitude assemblages into high latitudes between 17–16 Ma (see Figure 13.3), which
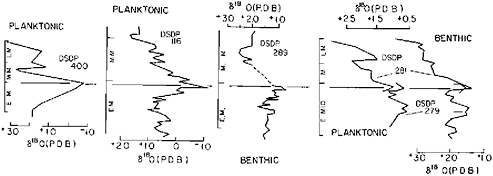
FIGURE 13.4 Miocene oxygen isotope stratigraphy of DSDP sites from various sources; planktonic foraminiferal curve for DSDP sites 400 (Vergnaud-Grazzini et al., 1978) and 116 [Rabussier-Lointier and others (C. Vergnaud-Grazzini, University of Pierre and Marie Curie, Paris, personal communication. 1980)]; and planktonic and benthic foraminiferal curves from DSDP sites 279 and 281 (Shackleton and Kennett, 1975). Line connects the major warm acme between 17–16 Ma (see text).
is also observed in the southern Atlantic Ocean (Haq, 1980). It seems to have been the salient climatic event of the Miocene epoch. The relatively sharp drop in temperatures that followed this warm episode is also clearly indicated by isotopic curves (Figure 13.4) and seems to have been related to the onset of extensive glaciation on Antarctica. A late Miocene warming between 9–7.5 Ma and the cooling event that followed are also evident as well from the δ18O curves from DSDP sites 400 and 281 (Figure 13.4) and from DSDP sites 357 (Boersma and Shackleton, 1977) and 398 (Vergnaud-Grazzini et al., 1978).
These examples from the Cenozoic acmes and corroborative evidence from oxygen isotopic data indicate that most of these events were widespread and at least some of them were of global extent.
STRATIGRAPHIC IMPLICATIONS
Stratigraphy is primarily an exercise of arranging the succession of “events” aimed at achieving better correlation capability for the documentation of historical geology. Biostratigraphy, one aspect of stratigraphy, utilizes the sequences of biostratigraphic “events” (first and last occurrences of taxa and some-times their dominances) and their inferred (or interpolated) ages to arrive at a utilitarian biochronology. The extent to which biostratigraphic zones or datum events can be applied for correlations is limited by the geographic distribution of the defining taxa. This is the basic cause of the difficulties in biostratigraphically correlating high- and low-latitude sequences.
The main reason for the latitudinal differences in faunal and floral assemblages is the ecologic exclusion of most low-latitude taxa from the higher latitudes; the high-stress environments of the high latitudes inhibit the development of diversified assemblages, and thus much of the time the high latitudes are populated with few, cosmopolitan, robust, and well-adapted eurythermal taxa, with long-stratigraphic ranges that are only marginally, if at all, useful in biostratigraphy. It is only during periods of marked climatic ameliorations (warm acme events) that tropical-temperate assemblages expand into higher latitudes, making lower latitude zonal schemes applicable in higher latitudes for the duration of the event. A good example of this is the late Paleocene-early Eocene and the early middle Miocene intervals when global warming episodes make tropical-subtropical biostratigraphies more readily applicable in higher latitudes. During other times in the Cenozoic the low- and high-latitude correlations are difficult at best.
The delineation of migrationary patterns of assemblages in response to climatic acmes can provide a solution to the highto low-latitude correlation dilemma. As mentioned earlier, if episodes of extreme environmental changes are geologically contemporaneous, then acme events will approximate synchronous horizons. Acme horizons (Figure 13.5), or maxima in migrationary cycles, will define approximate time lines, and both the warm and cold acme events can be conveniently used for correlation purposes. The acme horizons are similar to, but more narrowly defined than, acme or peak zones, which are characterized by the “exceptional” abundance of a
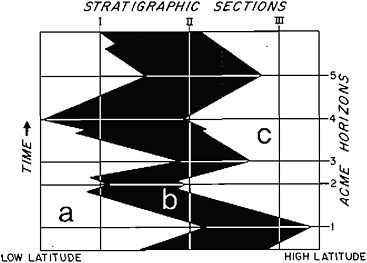
FIGURE 13.5 Acme horizons defined by maximum latitudinal migrations of assemblages in response to climatic acme events. Areas labeled a, b, and c delineate the latitudinal distribution of hypothetical assemblages and record latitudinal migrations of these assemblages through time. Acme horizons 1–5 approximate isochrons and help in the stratigraphic correlation of sections I, II, and III. A given acme horizon may be defined by abundances of different assemblages in different areas, allowing the horizon to be extended across all latitudes. At a given latitude it may not be possible to identify uniquely a given acme horizon from the many existing within a stratigraphic sequence. Thus stratigraphic sections must first be approximately correlated by the usual biostratigraphic methods, and then the acme horizons are used to refine the time stratigraphy. (After Haq and Lohmann, 1976.)
taxon rather than by the maximum abundance of an environmentally sensitive assemblage (Haq and Lohmann, 1976).
The use of acme horizons as a correlation tool has been illustrated in Figure 13.5. Essentially, the procedure involves three steps: (1) delineation of cycles of latitudinal migration of assemblages recorded in stratigraphic sequences, (2) approximate correlations of these cycles at different latitudes by the usual biostratigraphic methods, and (3) refinement of this approximate time framework using migrationary maxima as time lines. This scheme is analogous to the method of local time correlation that uses position within bathymetric cycles recorded in transgressive-regressive stratigraphic sequences.
ACKNOWLEDGMENTS
The author is indebted to C.Vergnaud-Grazzini and F. Woodruff for providing copies of the unpublished isotopic curves from DSDP sites 116 and 289, respectively. This paper was reviewed by W.A.Berggren, B.H.Corliss, and F. Thayer. The author’s research is supported by a grant from the National Science Foundation, Division of Submarine Geology and Geophysics, No. OCE78–19769. This is Woods Hole Oceanographic Institution Contribution Number 4617.
REFERENCES
Berggren, W.A. (1969). Rates of evolution in some Cenozoic plank tonic foraminifera, Micropaleontology 15, 351–365.
Boersma, A., and N.J.Shackleton (1977). Tertiary oxygen and carbon isotopic stratigraphy, Site 357 (mid-latitude South Atlantic), in Initial Reports Deep Sea Drilling Project 39, U.S. Government Printing Office, Washington, D.C., pp. 911–924.
Boersma, A., and N.J.Shackleton (1978). Oxygen and carbon isotope record through the Oligocene, DSDP Site 366, Equatorial Atlantic, in Initial Reports Deep Sea Drilling Project 41, U.S. Government Printing Office, Washington, D.C., pp. 957–962.
Boersma, A., N.J.Shackleton, M.Hall, and Q.Given (1979). Carbon and oxygen isotope records at DSDP site 384 (N. Atlantic) and some Paleocene paleotemperatures and carbon isotope variations in the Atlantic Ocean, in Initial Reports Deep Sea Drilling Project 43, U.S. Government Printing Office, Washington, D.C., pp. 698–717.
Buchardt, B. (1978). Oxygen isotope palaeotemperatures from the Tertiary period in the North Sea area, Nature 275, 121–123.
Fischer, A.G., and M.L.Arthur (1977). Secular variations in the pelagic realm, in Deep Water Carbonate Environments, H.E. Cook and P.Enos, eds., Soc. Econ. Paleontol. Mineral. Publ. No. 25, pp. 119–150.
Fredericksen, N.O. (in press). Mid-Tertiary climate of southeastern United States: The sporomorph evidence, J. Paleontol.
Haq, B.U. (1973). Transgressions, climatic change and diversity of calcareous nannoplankton, Mar. Geol. 15, 25–30.
Haq, B.U. (1980). Biogeographic history of Miocene calcareous nannoplankton and paleoceanography of the Atlantic Ocean, Micropaleontology 26, 414–443.
Haq, B.U., and G.P.Lohmann (1976). Early Cenozoic calcareous nannoplankton biogeography of the Atlantic Ocean, Mar. Micropaleontol. 1, 119–194.
Haq, B.U., I.Premoli-Silva, and G.P.Lohmann (1977). Calcareous plankton paleobiogeographic evidence for major climatic fluctuations in the Early Cenozoic Atlantic Ocean, J. Geophys. Res. 82, 3861–3876.
Shackleton, N.J., and J.P.Kennett (1975). Paleotemperature history of the Cenozoic and initiation of Antarctic glaciation: Oxygen and carbon isotope analyses in DSDP Sites 277, 279, and 281, in Initial Reports Deep Sea Drilling Project 29, U.S. Government Printing Office, Washington, D.C., pp. 743–755.
Vergnaud-Grazzini, C., C.Pierre, and R.Le’Tolle (1978). Paleoenvironment of the N.E. Atlantic during Cenozoic: Oxygen and carbon isotope analyses at DSDP sites 398, 400A and 401, Oceanol. Acta 1, 381–390.
Wolfe, J.A. (1978). A paleobotanical interpretation of Tertiary climates in the Northern Hemisphere, Am. Sci. 66, 694–703.







