

















Below is the uncorrected machine-read text of this chapter, intended to provide our own search engines and external engines with highly rich, chapter-representative searchable text of each book. Because it is UNCORRECTED material, please consider the following text as a useful but insufficient proxy for the authoritative book pages.
3 Patterns of Biodiversity and Endemism on Indo-West Pacific Coral Reefs MARJORIE L. REAKA,* PAULA J. RODGERS,* and ALEXEI U. KUDLAâ Diversity of the primary groups of contemporary Indo-West Pacific coral reef organisms, including mantis shrimps (stomatopod crus- taceans), peaks in the Indo-Australian Archipelago (IAA), reaches a lower peak in East Africa and Madagascar [Indian Ocean con- tinental (IOC)], and declines in the central Indian Ocean (IO) and Central Pacific (CP). Percent endemism in stomatopods (highest in the IAA, high in the IOC, lower in regions adjacent to centers, and moderate in the CP) correlates positively with species diver- sity (this varies with scale) and inversely with species body size. Because it constrains reproductive traits and dispersal, body size is a reliable indicator of speciation and extinction potential in reef stomatopods and probably most marine organisms. Assemblages are dominated by small-sized species in the IAA and IOC. Both speciation and extinction likely are high, resulting in especially high endemism (small ranges reflect both originating and disap- pearing species) in these regions. Rates of speciation exceed extinction, yielding centers of diversity (especially in the IAA). Dis- persal slows speciation and extinction in areas adjacent to these centers. Body size declines toward the CP, especially in atoll environments. Here the wheels of speciation and extinction again spin rapidly but in the opposite direction (extinction > speciation), *Department of Biology, University of Maryland, College Park, MD 20742; and â Georgetown University School of Medicine, Washington, DC 20007. 45
46â /â Marjorie L. Reaka, Paula J. Rodgers, and Alexei U. Kudla yielding low diversity and moderate endemism. We conclude that life histories, dispersal, and speciation/extinction dynamics are primary agents that mold patterns of diversity and endemism. His- torical factors, currents, productivity, and species diversity itself (through ecological interactions) also influence these patterns, in some cases by altering body size. B eing repositories of ancient phyla as well as more recent special- ized taxa, coral reefs are among the most spectacular, produc- tive, diverse (per unit area), and threatened ecosystems on Earth. Organisms they house provide a critical source of protein for people in many tropical countries, and reefs themselves protect human populations from storm and wave damage. Coral reefs provide aesthetic beauty (and the bioeroded sand on beaches) for tourismâan increasingly important economic resource for developing tropical countries. However, tourism and other uses of reefs must be carefully managed to be of sustainable economic benefit. A fundamental value of coral reefs is that they provide an aesthetic, intellectual, educational, and cultural heritage for present and future generations. Threats to global coral reefs, however, are severe and well documented (Bryant et al., 1998; Burke et al., 2002; Gardner et al., 2003; Hughes et al., 2003; Pandolfi et al., 2003, 2005; Bellwood et al., 2004; Burke and Maidens, 2004; Hoegh-Guldberg et al., 2007). Overexploitation has been identified as an especially serious problem, but other threats include coastal develop- ment and sedimentation, pollution, global warming, disease, and ocean acidification. The Global Coral Reef Monitoring Network reports that 20% of global coral reefs already have been degraded beyond recovery, an addi- tional 24% are under imminent threat of collapse, and a further 26% are at longer-term risk (Wilkinson, 2004). In the Indo-West Pacific (IWP), 88% of Southeast Asian reefs and 61% and 54% of Middle Eastern and Indian Ocean (IO) reefs are at medium to high risk (Bryant et al., 1998; Burke et al., 2002). Human population density near reefs is particularly high in Southeast Asia and the IO. Pacific reefs are in better condition (59% at low risk), with more protected area, than those in other regions. Despite the biological, cultural, and economic value of coral reefs and the widely publicized alarm at their global decline, it remains astonish- ing how little we know about the patterns of diversity (ââdiversityââ will be used equivalently to species richness) on coral reefs that would help us manage and conserve them. This chapter will (i) review and provide new information on the geography of coral reef diversity in the IWP using information from the most ecologically important and well-known groups of reef organisms, (ii) provide a brief overview of the major factors that
Patterns of Biodiversity and Endemism on Indo-West Pacific CoralReefsâ /â 47 generate these patterns, and (iii) briefly review ââhotspotsââ and provide new information on endemism in reef stomatopods. (iv) We will then examine information on body size, life history characteristics, geographic ranges, and speciation/extinction dynamics of reef stomatopods and other organisms to suggest mechanisms that, in combination with environmen- tal factors, can explain the observed patterns of IWP coral reef diversity and endemism. We will focus on reef-dwelling mantis shrimps as a model taxonomic group for such analyses because these crustaceans are important members of the benthic community. All stomatopods are predators with a pair of enlarged, equally sized raptorial claws that are used to smash and spear prey, competitors, and predators (Fig. 3.1). Protective holes in the substrate are a limiting resource because of strong fish predation; stomatopods exhibit colorful communicatory displays and intense territorial fighting to maintain exclusive ownership of these holes (Caldwell and Dingle, 1975; Reaka and Manning, 1981). PATTERNS OF CORAL REEF DIVERSITY Species diversity for several marine taxa (fishes, corals, lobsters, and snails) reaches a global maximum in the ââEast Indies triangleââ (Malay- sia, Indonesia, New Guinea, and the Philippines) of the Indo-Australian Archipelago (IAA), declines in the IO (with heightened diversity in some parts of the western IO for many taxa), declines eastward across the Cen- tral Pacific (CP), and peaks again in the Caribbean (Steli and Wells, 1971; Briggs, 1995, 1999a, 2000, 2003, 2007; Veron, 1995; Paulay, 1997; Bellwood and Hughes, 2001; Hughes et al., 2002; Roberts et al., 2002; Meyer, 2003; Mora et al., 2003; Bellwood et al., 2005). The first diversity contour map for reef stomatopods (Fig. 3.2) shows a similar pattern, with a high, sharp peak in the central IAA and an area of elevated diversity in the northwestern IO that increases southward to a secondary peak around Madagascar. Similar to most other taxa where contours of diversity are known, tails of diversity extend from the IAA toward the southeast and northeast. EXPLANATIONS OF IWP DIVERSITY GRADIENTS After considering all explanations for patterns of IWP reef biodiver- sity, we identify here only those that are most applicable to the present study.
48â /â Marjorie L. Reaka, Paula J. Rodgers, and Alexei U. Kudla FIGURE 3.1â Dorsal view of typically sized reef stomatopods (large Gonodactyla- ceus falcatus, small Gonodactylellus incipiens; scale bar for both is 10.0 mm), lateral view of raptorial claw, and photograph of individual delivering a threatening display at the entrance of its burrow (Gonodactylaceus ternatensis, courtesy of Roy Caldwell). Drawing of the raptorial claw shows the merus (m), meral spot (ms, which varies in color from white through yellow, orange, red, pink, purple, and blue among species), propodus (p), and dactyl (d); p and d normally are folded against m but are opened (arrow) either together as a hammer or with d projected zpq9990837580001.c.eps as a spear. Coloration of the flared antennal scales and setae also varies from yel- low to red, blue, and green among species. (20p6) Faunal Carryover Hypothesis Species from a Mesozoic/early Cenozoic center of diversity in Europe migrated east to the IAA and probably south along the continental margin of the Indian Ocean (IOC) as the Tethys Seaway was closed by the collision of Africa with Eurasia between the Paleocene and Miocene (Briggs, 2000, 2003, 2007). High stomatopod diversity in the IAA and in the western/ southwestern IO (Fig. 3.2) is consistent with this hypothesis. Center of Accumulation Hypothesis Species originate in small peripheral populations, larvae from periph- eral regions are carried by currents into central areas favorable for reef growth (arrows in Fig. 3.2), and species accumulate in these current-fed
Patterns of Biodiversity and Endemism on Indo-West Pacific CoralReefsâ /â 49 6-10 <5 6-10 16 21 - 2 -2 11-15 <5 0 5 16-20 0 6-10 -2 6-10 11 16 11-15 - 15 46-50 11-15 <5 21 26 21-25 -2 -30 5 16 5 -2 -1 6-10 0 11 FIGURE 3.2â Contours of species richness for reef stomatopods in the IWP. Num- bers represent species present in each contour. Arrows indicate major currents. All species of Alainosquillidae, Gonodactylidae, Odontodactylidae, Protosquillidae, and Takuidae are included; Pseudosquillidae occur on reefs but are excluded from analysis because their reproductive, larval, and life history patterns differ from those of other reef-dwelling families (Reaka, 1979, 1980; Reaka and Manning, 1987a). Data are from our own collections, National Museum of Natural History collections, and published literature [updated to currently accepted taxonomy (Ahyong, 2001; Schram and Muller, 2004)]. regions over time (Ladd, 1960; Jokiel and Martinelli, 1992; Briggs, 1995; Connolly et al., 2003). Peaks of stomatopod diversity in the IAA and west- ern IO are consistent with this hypothesis, but Barber and Bellwood (2005) and the present study find speciation and endemism in both peripheral regions and diversity centers. Energy/Productivity Hypothesis Higher productivityâthe rate at which energy flows through an eco- systemâallows an ecosystem to support more species (although diversity often declines at very high levels of productivity) (Rosenzweig, 1995). Similarly, increased temperature accelerates speciation (Allen and Gillooly, 2006; Allen et al., 2006), but Bellwood et al. (2005) find no relationship between sea surface temperature and diversity of reef corals and fishes. Phytoplankton abundance has not been compared previously with con- tours of reef diversity. The general pattern of stomatopod diversity cor- relates fairly well with phytoplankton productivity (Figs. 3.2 and 3.3). We later infer that phytoplankton productivity affects body size and extinc- tion/speciation dynamics of stomatopods on high (volcanic peaks with
50â /â Marjorie L. Reaka, Paula J. Rodgers, and Alexei U. Kudla FIGURE 3.3â Distribution of global phytoplankton pigment concentration (adapt- ed with permission from the National Aeronautics and Space Administration). Among reef areas, phytoplankton pigments are particularly abundant (0.3â0.4 mg/m3) off the Arabian Peninsula and west coast of India/Sri Lanka, around the Malay Peninsula/Indonesia, and around New Guinea/northern Australia. Pigment concentrations decline to 0.15 mg/m3 in a relatively narrow band ocean- ward from the above areas; immediately offshore from the continental margin of western Australia, eastern Africa, Madagascar; and in an equatorial band extend- ing westward from the central East Pacific. Still further offshore, phytoplankton pigments decline (0.05 mg/m3) until they reach the very low levels characteristic zpq9990837580003.c.eps of the centers of the northern and southern gyres of the Indian and Pacific Oceans (<0.05 mg/m3, smallest oceanic circles). We drew black lines for each of these 20p6 contours by eye from map coloration. extensive terrestrial area and soil) vs. low (carbonate atolls with little elevation or nutrient runoff) mid-Pacific islands. The fact that terrestrial runoff elevates productivity around high vs. low Pacific islands, enhanc- ing the survival of phytoplankton-feeding starfish larvae and fostering crown of thorns starfish population explosions (Birkeland, 1982; Brodie et al., 2005; Houk et al., 2007), suggests that productivity can have impor- tant effects on larval recruitment, local reef populations, and hence reef biodiversity. Center of Origin Hypothesis ââSuccessfulââ lineages originate in the East Indies; species subsequently migrate into peripheral regions, where they remain as relicts; gaps in spe- cies distributions suggest high extinction as well as origination in the East
Patterns of Biodiversity and Endemism on Indo-West Pacific CoralReefsâ /â 51 Indies (Briggs, 1995, 1999a,b, 2000, 2003, 2007). Although much evidence supports this hypothesis, species of reef fish are not consistently young in the IAA and old in peripheral regions (Barber and Bellwood, 2005), and the present study suggests that, in reef stomatopods, both speciation and extinction are high in peripheral areas as well as in the IAA. Species Diversity Hypothesis High species diversity itself may promote diversification [Emerson and Kolm (2005), but see Cardena et al. (2005)], probably through species interactions. Speciation rates in three groups of fossil plankton over 2â20 Myr correlate with species diversity independent of sampling intensity and area (Allen and Gillooly, 2006). In the present study, species interac- tions in diverse assemblages may cause shifts in body size and consequent changes in life history patterns and speciation/extinction patterns. Life History Speciation/Extinction Hypothesis We will suggest that the biotic and environmental processes that gov- ern body size and life history traits drive rates of speciation/extinction and thus patterns of diversity in IWP stomatopods and other reef organisms. ENDEMISM AND HOTSPOTS Endemism has been of particular interest as an indicator of extinc- tion. The concept of biodiversity hotspotsâconcentrations of endemic species that are at exceptional riskâwas motivated by the need to estab- lish conservation priorities (Myers, 1988; Reid, 1998; Myers et al., 2000; Mittermeier et al., 2004). However, some authors have argued that high overall diversity or phylogenetically unique taxa or habitats deserve pri- ority attention, and others have shown that centers of endemism do not always coincide among taxa or with degree of threat (Prendergast et al., 1993; Williams et al., 1996; Bonn et al., 2002; Orme et al., 2005). For some of the same reasons, hotspots on global coral reefs have been controversial (Baird et al., 2002; Briggs, 2002; Hughes et al., 2002; Roberts et al., 2002). Here we examine patterns of endemism in the context of both speciation and extinction, because limited ranges occur during both processes. Con- servation of areas rich in endemics is important not only because they are at particular risk of species loss but also because they represent potential sources of diversification. Endemism in reef stomatopods varies with scale. When stomatopods are known from only a single locality, these ââlocal endemicsââ are widely scattered (Fig. 3.4), with no significant relationship between number of
52â /â Marjorie L. Reaka, Paula J. Rodgers, and Alexei U. Kudla FIGURE 3.4â ââLocally endemicââ species of reef stomatopods. endemics and number of species found at each local site (x2 = 12.72, df = 20, P > 0.05). Average endemism for the 21 local sites is 10.5 ± 2.6% (SE). ââRestricted regional endemicsâââthose known from local sites, archi- pelagos, and parts of regionsâare widespread but reach their highest incidences (>15%) in the Malay Peninsula/Indonesia/Philippines, Red Sea, Mascarene Islands, Madagascar/Comores Islands, Society Islands, and zpq9990837580004.g.eps However, the distribution of restricted Hawaiian Islands (Fig. 3.5). regional endemics does not differ from those expected when the diversi- ties of their subregional stomatopod faunas are considered (x2 = 25.13, df 42 to 27 = 23, P > 0.05). Endemism across the 24 subregions averages 14.5 ± 2.1%. To examine patterns of endemism at a larger scale, the IWP was divided into major ââregionsââ of continental and oceanic habitat types. At this scale, percent endemism (squares) is significantly concentrated in the IAA and drops in the adjacent oceanic regions but then rises toward the FIGURE 3.5â âRestricted regional endemicâ species of reef stomatopods. Of the 24 subregions considered, those that exceed 10% are shown. Reaka.Fig5.B&W.tif 40 to 27
Patterns of Biodiversity and Endemism on Indo-West Pacific CoralReefsâ /â 53 FIGURE 3.6â Endemism and body size of reef stomatopods in six different regions (dashed circles) consisting of the IOC, IO, IAA (the western margin of the IAA abuts the IOC off Burma), Western Pacific margin (WPM), West Central Pacific Reaka.Fig6.B&W.tif (WCP), and CP. Numbers in each region represent (i) percentage of species in each regionto 27 endemic (square, top number in each region; percentage is used 54 that are to avoid confounding diversity with endemism and because the regions are not equal in area), (ii) percentage of species in each region <40 mm in body length (oval, middle number in each region; 40 mm is the median body size across all regions), and (iii) median body size (millimeters total length) among species in each region (underline bar, bottom number in each region). west in the IOC and, to a lesser extent, toward the east in the West Central Pacific and especially the CP (x2 = 26.23 using raw numbers of endem- ics/nonendemics, df = 5, P < 0.001; Fig. 3.6). Average endemism among the six regions is 20.2 ± 6.1%. The percent regional endemism increases significantly with regional species richness (Fig. 3.7; r2 = 0.74, P = 0.03, F = 11.41, y = 0.08x + 1.61, square-root-transformed data). The percent regional endemism is inversely related to body size among regions, decreasing with increased median body size (Fig. 3.7; r2 = 0.65, P = 0.05, F = 7.53, y = 0.10x + 8.16, square-root-transformed data) and increasing with percentage of species in each region that are <40 mm in body size (r2 = 0.68, P = 0.04, F = 8.39, y = 0.08x â 0.15, square-root- transformed data). Species are concentrated in small body size classes in the IAA and IOC, although the range of body sizes is large in these regions. Typical body sizes are larger in the oceanic regions adjacent to the IAA and IOC but decline (with an absence of large-sized species) toward the CP (Fig. 3.6; number of species >40 mm and <40 mm for all regions, x2 = 11.07, df = 5, P = 0.02). Hypothesizing that productivity influences body sizes and life his- tories of reef stomatopods, we further categorized the species in the six regions according to whether they inhabited productive or unproductive
54â /â Marjorie L. Reaka, Paula J. Rodgers, and Alexei U. Kudla Species Richness 0 20 40 60 % Regional Endemics 50 40 30 20 10 0 0 20 40 60 Median Body Size of Species in Region FIGURE 3.7â The percentage of regional endemics vs. median body size of species in each region (open diamonds) and species richness in each region (filled squares) for reef stomatopods. Residual analyses show no difference in and statistical zpq9990837580006.g.eps results are similar for14 p to 27 transformed and untransformed data; untransformed data are shown here. environments. Because of terrestrial runoff, continental regions and high islands are expected to have higher nearshore productivity than low oceanic islands. Analysis of maximal speciesâ body sizes on continental and high vs. low island environments shows, as above, that endemism is consistently associated with small body size (F = 39.27, df = 157, P < 0.001, type 3 ANOVA tests of fixed effects for regional endemism, terrain height, and region, log-transformed data; analysis using species median body size yields the same result). Terrain height is not consistently related to body size across all six regions, probably because of the large species scattered throughout the IOC, IO, and IAA (type 3 ANOVA of fixed effects as above, P > 0.05; for number of species >40 mm and <40 mm on continental/high vs. low islands across the region, x2 = 15.3, df = 9, P = 0.08). However, a previous study of reef stomatopods showed that body size of individual populations within each of four species complexes of reef stomatopods declines significantly from the IAA toward the CP and that populations on high islands reach significantly larger body sizes than those inhabiting atolls in these regions (Reaka-Kudla, 2000). Guided by the previous study, we analyzed maximal and median body sizes of species assemblages from high vs. low islands from the West Central Pacific and CP. High islands support species of significantly larger body size than low islands in these regions (F = 4.79, df = 40, P = 0.03; type 3 ANOVA of fixed effects for region, island height, and regional endemism, log-transformed data; analysis of median body size of species yields a similar result).
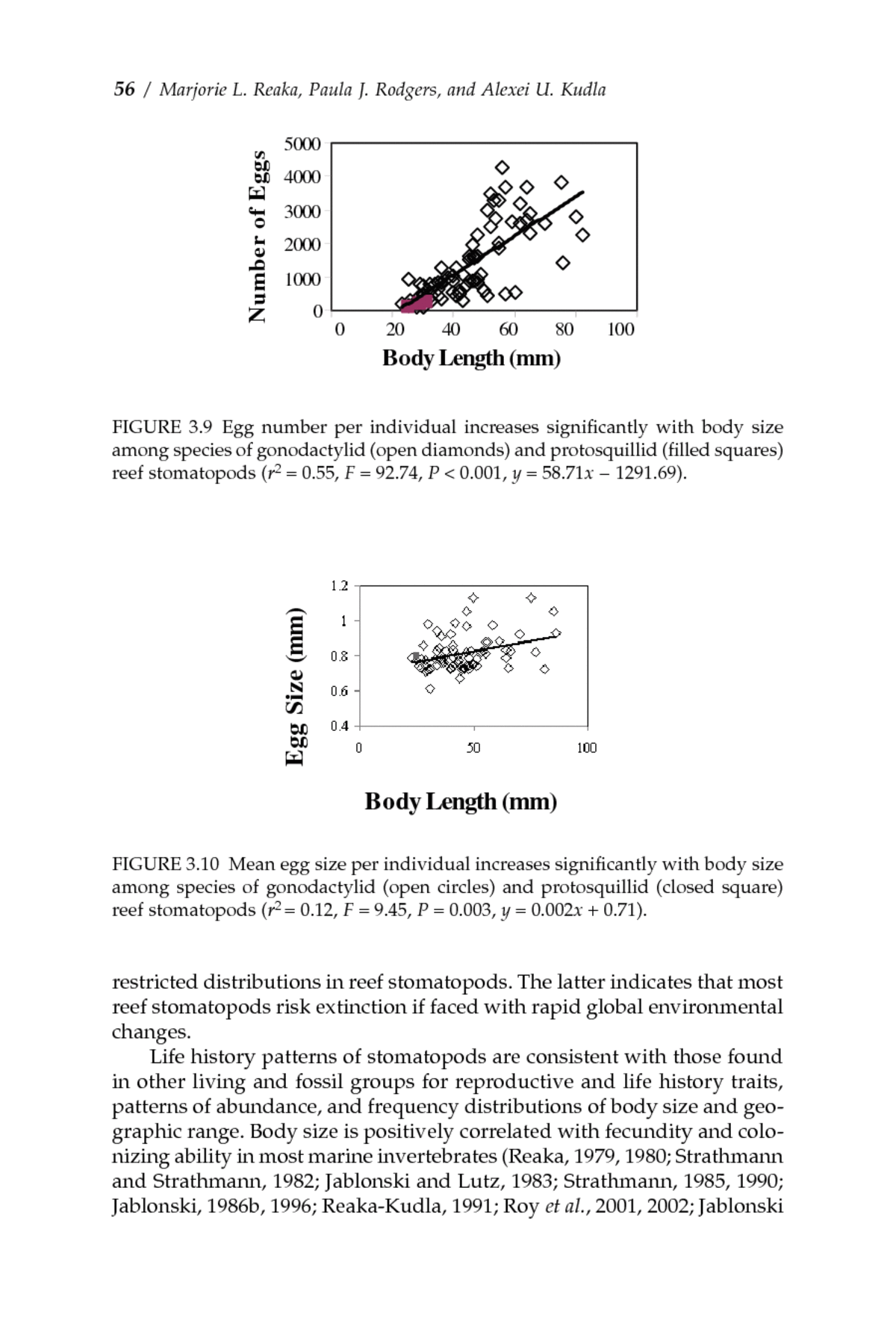
Patterns of Biodiversity and Endemism on Indo-West Pacific CoralReefsâ /â 55 100 All species 80 High Islands Number 60 Low Islands 40 20 0 Endemic Nonendemic FIGURE 3.8â Number of regional endemic vs. nonendemic species of reef stomato- pods and those occurring on high vs. low islands. Number of endemics and nonendemics on high vs. low islands did not zpq9990837580007.g.eps vary significantly across regions (P > 0.05), so all regions were combined. Significantly fewer endemics occur on low than high islands (x2 = 19.16, df = 1, P < 0.001; Fig. 3.8). LIFE HISTORY PATTERNS OF REEF STOMATOPODS We propose that the ecological and environmental factors that govern body size and life history traits drive patterns of diversity and endemism in reef organisms. Large body size in reef stomatopods is significantly correlated with massive reproductive output (more and larger eggs; Figs. 3.9 and 3.10), greater planktonic larval dispersal, larger geographic ranges, and greater saturation of available reef habitat within ranges (Fig. 3.11), whereas small body size correlates with restricted reproductive capacity, restricted larval dispersal, and relative rarity (low abundance, few sites, small geographic ranges) (Reaka, 1979, 1980; Reaka and Manning, 1987a; Reaka-Kudla, 1991; Wilkinson, 2004). This correlation occurs because small body size constrains reproductive traits in marine organisms. Small-bod- ied species cannot produce enough small plankton-feeding larvae to leave one surviving offspring given the high mortality rate suffered by these long-lived larvae. Small species must endow fewer, larger larvae with yolk supply, often brooding them before a relatively brief planktonic period, to increase survivorship. The body volume of larger species allows them to produce sufficient numbers of small larvae that feed in the plankton for long periods that some offspring survive despite heavy mortality (Menge, 1975; Strathmann and Strathmann, 1982). Size frequency distributions for both body size (Fig. 3.12) and geographic range (Fig. 3.13) are strongly shifted toward diminutive sizes (particularly in endemic species) and
56â /â Marjorie L. Reaka, Paula J. Rodgers, and Alexei U. Kudla 5000 Number of Eggs 4000 3000 2000 1000 0 0 20 40 60 80 100 Body Length (mm) FIGURE 3.9â Egg number per individual increases significantly with body size Xzpq9990837580008.g.eps among species of gonodactylid (open diamonds) and protosquillid (filled squares) 12 to 27 reef stomatopods (r 2 = 0.55, F = 92.74, P < 0.001, y = 58.71x â 1291.69). Egg Size (mm) Body Length (mm) FIGURE 3.10â Mean egg size per individual increases significantly with body size Reaka.Fig10.B&W.tif among species of gonodactylid (open circles) and protosquillid (closed square) reef stomatopods (r2 = 0.12, F = 9.45, P = 0.003, y = 0.002x + 0.71). restricted distributions in reef stomatopods. The latter indicates that most reef stomatopods risk extinction if faced with rapid global environmental changes. Life history patterns of stomatopods are consistent with those found in other living and fossil groups for reproductive and life history traits, patterns of abundance, and frequency distributions of body size and geo- graphic range. Body size is positively correlated with fecundity and colo- nizing ability in most marine invertebrates (Reaka, 1979, 1980; Strathmann and Strathmann, 1982; Jablonski and Lutz, 1983; Strathmann, 1985, 1990; Jablonski, 1986b, 1996; Reaka-Kudla, 1991; Roy et al., 2001, 2002; Jablonski
Patterns of Biodiversity and Endemism on Indo-West Pacific CoralReefsâ /â 57 40 % Sat. Habitat 30 20 10 0 0 20 40 60 80 100 120 Body Length (mm) FIGURE 3.11â The percentage saturation of available reef habitat within each spe- ciesâ geographic range increases significantly with median body size among spe- zpq9990837580009.g.eps cies of gonodactylid and protosquillid stomatopods (r2 = 0.54, F = 47.68, P < 0.001, y = 0.19x â 3.83). The percentage saturation of each speciesâ range is the proportion of all 5 à 5° latitudinal and longitudinal quadrants containing habitable reef that is occupied by that species. 40 All species 35 Endemics 30 Frequency Nonendemics 25 20 15 10 5 0 . 99 . 99 . 99 . 99 . 99 . 99 . 99 . 99 . 99 0 -9 2 0- 29 4 0- 49 6 0- 69 8 0- 89 10 9 12 9 14 9 16 9 1 00- 1 20 - 1 40 - 1 60 - Maximum Body Size Classes (mm) FIGURE 3.12â Size frequency distributions of maximum body sizes for total spe- cies, regional endemics, and nonendemic species of reef stomatopods. Endemics zpq9990837580010.g.eps are significantly smaller than nonendemics (t = 6.40, df = 146, P < 0.001; two- sample t test assuming unequal variance). Analysis of median body size for each species gives a similar graphical and statistical result. et al., 2003a). Stomatopods share the following characteristics with taxa in both marine and terrestrial environments. (i) Body size frequency dis- tributions are usually shifted toward small size classes (May, 1978, 1986, 1988; Rosenzweig, 1995; Gaston and Blackburn, 2000; Roy et al., 2000, 2002). (ii) Species abundances are biased toward few common and many rare species (Hubbell, 2001). (iii) Local abundance increases with range size (Gaston, 1994). (iv) Geographic range size increases with body size
58â /â Marjorie L. Reaka, Paula J. Rodgers, and Alexei U. Kudla Number of Species 40 30 20 10 0 5 10 15 20 25 30 35 40 45 50 55 60 65 70 Size of Geographic Range (No. of 5x5º Lat. x Long. Quadrats) FIGURE 3.13â Size frequency distribution of geographic ranges in reef stomato- zpq9990837580011.g.eps pods (Lat, latitude; Long, longitude). Other measures of geographic range size [latitudinal distance, longitudinal distance, square root of (latitude à longitude)2] yield a similar plot. (Gaston and Blackburn, 1996). (v) Frequency distributions for geographic ranges are shifted toward small ranges [Gaston (1994, 1998), Gaston and Chown (1999), and Roberts and Hawkins (1999), but see Hughes et al. (2002)]. Commonalities in patterns of body size, life history, and distribu- tion between reef stomatopods and other taxa suggest that the relationship between life history mechanisms and patterns of diversity and endemism we find in benthic reef organisms also may operate in other systems. SPECIATION AND EXTINCTION Factors that influence the relative rates of speciation vs. extinction control the geography of species diversity and endemism. This section will review briefly some of the factors thought to determine rates of extinction and speciation in marine and other organisms, and Discussion will apply these generalities to the patterns of diversity and endemism observed in IWP reef stomatopods. Species or genera that are widespread, abundant, and dispersive resist extinction in both marine and terrestrial environments and both fossil and contemporary lineages (Jablonski, 1986a,b, 1987, 1991; Gaston, 1994; Hubbell, 2001; Jablonski et al., 2003a). In addition to geographic range, which is sufficient on its own to explain species survival (Jablonski and Hunt, 2006), the presence of long-lived larvae and species richness of the clade confer resistance to background extinction in fossil marine bivalves (Jablonski, 1986a, 1991). Broad distribution of the clade confers protection against mass extinction (Jablonski, 1986a, 1991, 2007; Powell, 2007a). In addition, latitudinal distribution affects extinction, with the tropicsâ especially reef faunasâbeing subject to repeated upheaval, particularly during mass extinctions (Jablonski, 1991, 1993; Powell, 2007b).
Patterns of Biodiversity and Endemism on Indo-West Pacific CoralReefsâ /â 59 In stomatopods, evidence of extinction is derived from conspicuous gaps in regional distributions. For example, when a lineage occurs in the IWP, East Atlantic, and East Pacific but is absent from the West Atlantic, the most parsimonious explanation, given what is known about Cenozoic extinctions in the Americas (Jackson et al., 1996; Budd, 2000; Jackson and Johnson, 2000; Todd et al., 2002), is that it became extinct in the West Atlan- tic. Such ââapparent extinctionsââ are significantly elevated in lineages with restricted dispersal and small ranges as well as those from coral reefs and tropical latitudes (Reaka and Manning, 1987a). Although adult body size is not significantly associated with such gaps, small body size correlates significantly with reduced larval dispersal and small geographic ranges and can be used as an indicator of extinction risk (Reaka, 1980; Reaka and Manning, 1987a). Speciation also is related to life history characteristics and geographic range size. Although the relationship between geographic range size and speciation has been debated (Rosenzweig, 1995; Chown, 1997; Gaston, 1998; Gaston and Chown, 1999; Jablonski, 2007), range size is positively related to dispersal ability and gene flow in many marine and terres- trial taxa (Reaka, 1980; Reaka and Manning, 1981, 1987a; Jablonski and Lutz, 1983; Jablonski et al., 2003a; Paulay and Meyer, 2006), and these factors tend to dampen rates of speciation (Mayr, 1963; Hubbell, 2001; Jablonski et al., 2003a). Paleontological approaches allow measurement of both speciation rates and geographic range sizes over time. Range size is significantly inversely related to speciation rates in fossil gastropods and brachiopods (Jablonski and Roy, 2003; Jablonski et al., 2003a; Jablonski, 2007; Powell, 2007b). Because it constrains reproductive traits and geographic range, body size can be used as an indicator of speciation rate. Evidence that small- bodied species are engines of diversification comes from studies of mor- phological and taxonomic divergence of stomatopod lineages across bio- geographic barriers (Reaka and Manning, 1981, 1987a). Large-bodied lineages and those with long-lived larvae remain conspecific (morpho- logically indistinguishable) or closely related (cognate species) when separated by a barrier (e.g., the Central American isthmus) significantly more frequently than small-bodied species with abbreviated larval devel- opment. Conversely, the percentage of endemic species without cognates elsewhere and the percentage of species within locally radiating clusters of species (more closely related to each other than to any species outside the region) are significantly elevated in lineages of small body size, of limited larval dispersal, and from reefs. Recent molecular studies, including those on gastropods and small- bodied stomatopods, show that incipient or full speciation is more com- mon in reef fauna than previously thought (Barber et al., 2000, 2002, 2006;
60â /â Marjorie L. Reaka, Paula J. Rodgers, and Alexei U. Kudla Meyer, 2003; Meyer et al., 2005). In addition to the fact that most benthic reef species are small in body size (Reaka-Kudla, 1997; Bouchet et al., 2002; Meyer et al., 2005) with restricted or moderate colonizing ability, the behavior of reef larvae further reduces dispersal. Stomatopod larvae (and those of most other reef taxa) exhibit diurnal vertical migrations, hiding in reef rubble by day and migrating into the water column at dusk and dawn (Robichaux et al., 1981), which reduces exposure to currents. Although panda clownfish have a 9- to 12-day pelagic phase, one-third of marked juveniles settle within their natal area, many within 100 m of their birth site (GP Jones et al., 2005). DISCUSSION Using body size as an indicator of speciation and extinction rates, we infer that the IAA, and to a lesser extent the IOC, are areas of both high origination and high extinction in reef stomatopods. However, rates of origination must exceed those of extinction in these areas, yielding the high biodiversity observed. Endemism results from either newly origi- nated or almost extinct species and thus is expected to be especially high if both speciation and extinction are elevated, as is observed. Although species are concentrated in small size classes in the IAA and IOC, the range of body sizes is large in these areas (see Fig. 3.14). Historical factors (faunal carryover from the Tethys Seaway), productivity in the continental areas, currents, and species diversity itself (via ecological interactions between species) likely have contributed to the species richness and range of body sizes in the IOC and IAA. The dispersal and colonizing capability of large-sized species in these areas allows them to disproportionally colo- nize adjacent oceanic regions, where extensive larval immigration lowers extinction and retards speciation, yielding moderately diverse, somewhat larger-sized assemblages with low endemism. In the center of the IO and in the broader expanse of the Pacific, how- ever, larval immigrants have been filtered by starvation, predation, and distance. Given enough time, it is likely that larvae from diversity centers reach mid-ocean islands. However, both diversity and adult body size of reef stomatopods decline in the mid-Pacific, and body size is smaller on mid-Pacific atolls than on high islands, suggesting that productivity of the island environment, as well as propagule pressure, influences suc- cessful colonization. Dwarfed by limited productivity, populations cannot produce sufficient propagules to reach another island archipelago and are unlikely to receive many immigrants from ancestral populations to the west. They diverge into new species; endemism increases toward the CP. However, extinction also must be exceedingly high in these small-sized peripheral species. Endemics are missing from atolls, probably reflect-
Patterns of Biodiversity and Endemism on Indo-West Pacific CoralReefsâ /â 61 Body Size (mm) Geographic Region FIGURE 3.14â Body sizes of species on continental and high vs. low islands from the IOC to the CP. (H) and (L) refer to high and low island environments. Regional abbreviations are as in Fig. 3.6. Data for each species represent maximal recorded body length; plots for median body length provide a very similar pattern. Squares Reaka.Fig14.B&W.tif with crossbars are means. 37 to 27 ing the difficulty of establishing successful populations in these low- productivity environments that are heavily dominated by top predators (M.L.R., personal observation). We have observed one instance of popula- tion extinction in a small-sized reef stomatopod from a mid-Pacific atoll (Reaka and Manning, 1987b). Consequently, the wheels of speciation and extinction turn rapidly, but in reverse direction. If species arrive, specia- tion is high but extinction even higher; thus, diversity is low in remote oceanic regions of the IO and CP. Although the available evidence from life histories, geographic ranges, and extinction/speciation in stomatopods and other organisms supports this interpretation, molecular evidence on ages of species also is needed. We conclude that life history patterns and dispersal are the primary mediators of the rate and direction of the speciation/extinction cycle, which in turn determines the geography of diversity and endemism. However, productivity, historical factors (antecedent faunas), and currents likely influence diversity in particular localities. In addition, productivity, historical factors (lineage history), and species diversity itself (through ecological interactions) alter body size and thus influence life history and dispersal. ACKNOWLEDGMENTS We thank F. J. Ayala and J. C. Avise and two reviewers for helpful comments that improved the chapter.
