
Estimating Population Size and Growth Rates
Understanding the number and distribution of free-ranging horses and burros on their range is explicitly part of the mandate to the Bureau of Land Management (BLM) in the Wild Free-Roaming Horses and Burros Act of 1971 (P.L. 92-195). That act, as amended by the Public Rangelands Improvement Act of 1978 (P.L. 95-514), states that BLM “shall maintain a current inventory of wild free-roaming horses and burros on given areas of the public lands” to, in part, “make determinations as to whether and where an overpopulation exists and whether action should be taken to remove excess animals.”1 Thus, nearly all the management actions that BLM takes on Herd Management Areas (HMAs) are predicated on the population-size estimates of equids on the range. Population estimates aid in allocation and management of forage and habitat and underlie the establishment of appropriate management levels (AMLs). In addition, data on changing horse and burro abundance provide information that can be used to estimate population growth rates; aid in accruing knowledge to understand population and evolutionary processes (Chapters 3 and 5); assess the effectiveness of such management actions as removals, sex-age class manipulations, and contraceptive treatments to reduce population growth rates (Chapter 4); provide important information for assigning values to parameters of population models (Chapter 6); determine whether AMLs are being maintained and meeting their objectives (Chapters 5 and 7); and inform all those who have an interest in free-ranging horses and burros (Chapter 8). This chapter responds to the BLM request for a review of free-ranging horse and burro population estimates, techniques to improve those estimates, and population growth rates.
In fiscal year 2011, BLM spent about $641,250 to estimate the abundance of horses and burros on HMAs; that is about 1 percent of the Wild Horse and Burro Program’s annual budget (BLM, 2011). However, maintaining a current, accurate, and robust inventory of
____________
1 “Excess animals” are ones that “must be removed from an area in order to preserve and maintain a thriving natural ecological balance and multiple-use relationship in that area” (P.L. 95-514). Chapter 7 discusses the concept of thriving natural ecological balance and the multiple-use mandate of the act.
horses and burros living on land under its jurisdiction has been a continuing struggle for BLM. Because accurate estimates of free-ranging horse and burro populations are the foundation of scientifically based management of these animals, third parties have paid considerable attention to assessments of BLM’s methods for inventorying horses and burros over the history of the program (NRC, 1980, 1982; GAO, 1990, 2008). The committee received unfavorable comments during the study process from many members of the public regarding BLM’s reports of equid population estimates and assumed or reported population growth rates.
This chapter focuses initially on estimation of free-ranging horse and burro populations. It first distinguishes the difference between counting animals and estimating population size and discusses why this methodological distinction is important for management and transparency. It then reviews several classes of population-survey methods and their strengths, weaknesses, and applicability to free-ranging horses and burros. The section that follows evaluates information available on the methods used by BLM to inventory equid populations and report the results to the public and Congress when this study was conducted. Recent initiatives to improve BLM’s inventory procedures are then described with recommendations for strengthening the scientific validity and accuracy of the inventory program and enhancing communication of these important statistics to stakeholders. The second topic addressed in the chapter deals with population growth rates. A number of data sources that provide insight into growth rates of horse and burro populations are reviewed, and the results critiqued and synthesized. The chapter ends with a summary of the committee’s conclusions regarding BLM’s horse and burro inventory and reporting procedures and an assessment of typical population growth rates realized on western rangelands. The conclusions are then interpreted in the context of the challenges faced in managing free-ranging equid populations in the future.
ESTIMATING THE SIZE OF FREE-RANGING EQUID POPULATIONS
Since the inception of the Wild Horse and Burro Program, BLM’s population inventory program has involved attempting to survey completely the fixed areas occupied by free-ranging equids, known as HMAs, and to count all the animals detected. Those inventory surveys are commonly referred to as censuses in BLM reports; however, a census involves the perfect enumeration of every animal that occupies a given area of interest; that is, every animal is detected and counted. That is ideal, but counting free-ranging animal populations is an imperfect exercise. Topography, the extent of survey areas, vegetation structure, weather, animal behavior and coat color, the size of areas used by individual animals, the performance of aircraft used by observers, the skill and condition of observers, sun angle, cloud cover, and wind speed are some of the major factors that can influence the detectability of animals, which in turn affects the accuracy, efficiency, and effectiveness of survey methods (MacKenzie et al., 2006). For any given set of survey conditions, those factors can result in observers’ failure to detect animals that are present in a survey area or their unknowing detection and counting of the same animals on multiple occasions. Although animals can be missed or double-counted during the same survey, a large body of scientific literature on techniques for inventorying large mammals has demonstrated that failure to detect animals is overwhelmingly more common (Caughley, 1974a; Pollock and Kendall, 1987; Samuel et al., 1987). The first studies of probabilities of detection of free-ranging horses on western rangelands reported that in typical surveys only 7 percent of horses were undetected in flat, treeless terrain, but 50-60 percent were undetected in more rugged terrain with tree cover (Frei et al., 1979; Siniff et al., 1982). More recent studies of inventory
techniques have reaffirmed those conclusions (Walter and Hone, 2003; Laake et al., 2008; Lubow and Ransom, 2009; Ransom, 2012a). Overcounting horses has only been reported for a relatively high-density population in New Zealand where the systematic flight pattern of the helicopter, with closely spaced flight lines and routinely low altitude above ground, resulted in bands of horses unknowingly being counted several times (Linklater and Cameron, 2002).
Thus, the animal counts (the total number of animals tallied in a given survey) derived from BLM’s typical inventory procedures do not reflect the true number of animals in an HMA but instead represent what is more appropriately termed a population estimate, that is, an approximation of the true population that is based on the data collected (the count). The counts themselves represent the minimum number of animals occupying the HMA, but how closely the counts approximate the true number of animals occupying a given HMA depends on the proportion of the animals that are undetected and thus are not counted. For example, if a BLM aerial survey counted 180 horses on an HMA and 90 percent of the animals were detected, the count was a reasonably accurate population estimate in that the true number of horses occupying the HMA was 200. However, if only 50 percent of the animals were detected, the count would represent a poor population estimate in that the true population size was actually 360 horses. There is a large body of methodological and statistical literature on the development and testing of techniques for obtaining accurate and precise estimates of animal abundance (Seber, 1982; Pollock et al., 1990; Lancia et al., 1996; Nichols and Conroy, 1996; Krebs, 1999; Williams et al., 2001; Mills, 2007; Conroy and Carroll, 2009). It provides insights on how to detect and count animals better, procedures for estimating detection probability, and techniques for “adjusting” or statistically extrapolating count data collected in various ways to produce more accurate population estimates and measures of the precision of estimates.
Population Survey and Detection Methods
Scientifically robust surveying techniques are essential for obtaining accurate estimates of the abundance of free-ranging horses and burros that are necessary for successful management of herds on BLM-managed rangelands. As detailed above, horses and burros are imperfectly detected for a number of reasons, but ground-based assessments, aerial surveys, remote-sensing imagery, genetic techniques, or some combination of these can be effective for locating animals and estimating the size of a population of equids in a target domain, such as an HMA or an HMA complex. This section reviews selected survey methods that were supported by scientific research and in use as of late 2012. It also describes methods that may have potential for detecting free-ranging equids in a logistically and fiscally feasible manner.
Ground-Based and Aerial Survey Methods
To prevent undercounting or double-counting of free-ranging ungulates, especially in heterogeneous or topographically complex landscapes, several techniques have been developed that allow explicit quantification of sampling uncertainty and detectability of animals. The following methods have been applied effectively to estimate detectability and uncertainty in estimating the abundance of free-ranging horses and burros.
Strip and Line Transects. A target domain is sampled by traveling along lines that are often placed systematically across relatively homogeneous landscapes and, in more heterogeneous
landscapes, may be distributed in more complex arrangements to ensure adequate coverage (Caughley, 1974a; Buckland and Turnock, 1992).
The lines, known as transects, are typically traveled by aircraft that carry one or more observers to record animals detected. In strip-transect surveys, the observer constrains recording of animals to a relatively narrow width of the transect to try to fulfill the assumption that all animals in the transect are detected. The resulting data are used to estimate a density of animals in the areas covered by the strip transects, and this density is extrapolated to the entire area that was sampled to obtain an estimate of the number of animals in the sampled area (Burnham et al., 1980; Marsh and Sinclair, 1989). In line-transect surveys, observers record all animals spotted while they traverse the transect, typically using distance sampling (Buckland et al., 2004, 2005), in which all groups of animals detected are recorded with their perpendicular distance from the transect. Such data aggregated across many transects are then used to estimate a detection probability function, which assumes that all groups of animals on the transect line are perfectly detected, and detectability declines for groups of animals at increasing distances from the transect line. The primary advantage of this technique for free-ranging horses is that distance sampling can accommodate large spatial areas of high topographic and vegetative heterogeneity (J. Ransom, National Park Service, personal communication, August 10, 2012), and detection probability is explicitly modeled and estimated. Assumptions of the approach are that lines are placed randomly with respect to the distribution of the objects (such as equids) sampled, that equids do not move because of the aircraft (that is, they are detected at their initial locations), that perpendicular distances from the transect line to each equid group are measured accurately, and that detections are statistically independent events. U.S. Geological Survey biologists have as yet been unable to find a distance-measuring device that works satisfactorily, but they were developing such a tool (J. Ransom, National Park Service, personal communication, August 10, 2012). Ransom et al. (2012) used distance sampling and minimally trained local observers in Mongolia to estimate the abundance of wild asses (Equus hemionus).
Mark-Recapture and Mark-Resight. In mark-recapture studies, animals are uniquely marked (or identified individually on the basis of unique markings or characteristics) and later recaptured (either physically or with visual recapture methods) so that a detection history of each marked animal can be compiled. Population size can be estimated by applying open-population or closed-population mark-recapture models to detection-history data (Schwarz and Arnason, 1996; Williams et al., 2001). Software packages, such as Program MARK (White and Burnham, 1999), provide a flexible framework for implementing closed-population and open-population models in estimating abundance and related parameters. Whereas conventional capture-recapture methods for estimating population size (e.g., Otis et al., 1978; Williams et al., 2001) generally require animals to be uniquely marked in such a way that a detection history for each marked animal can be compiled, more recent mark-resight approaches can also incorporate sightings of unmarked animals into the estimation framework (McClintock and White, 2009). Mark-resight efforts can often be less expensive and less invasive (Minta and Mangel, 1989; McClintock and White, 2007) than traditional mark-recapture methods (Otis et al., 1978). In particular, animals need to be captured only one time (capture is often the most hazardous, stressful, and expensive aspect of these estimation techniques); after initial marking periods, additional data can be collected with sighting surveys, which do not necessitate physical capture of animals and thus are less invasive (McClintock et al., 2009). However, mark-resight methods assume that animals are sampled and resighted in a closed population (that is, no immigration, emigration, births, or deaths occur) and that the number of marked animals available for
resighting is known exactly or can be reliably estimated (McClintock et al., 2009). Those assumptions can be approximated by conducting sighting surveys soon after the initial marking (to ensure a closed population), by using radio collars with mortality sensors on all captured animals (McClintock and White, 2007), or by using other mark-resight models that do not require that the number of marked animals be exactly known (Arnason et al., 1991; McClintock et al., 2009). McClintock et al. (2009) provided an estimation framework that addresses both constraints by using Poisson-log (PNE) and zero-truncated Poisson logit-normal (ZPNE) mixed-effects models. Various versions of mark-resight models are available in the freeware Program MARK (White and Burnham, 1999).
Mark-resight techniques using natural distinguishing characteristics of horses have been used in Australia and New Zealand (Linklater and Cameron, 2002; Dawson and Miller, 2008), and Lubow and Ransom (2009) used a photograph-based form of mark- resight methods for enumerating free-ranging horses in the western United States by identifying each group of horses (via such markings as blaze, socks, and coat color) and determining how many groups were resighted on later flights. Transects should be widely spaced so that an HMA can be completely covered multiple times with differently oriented transects (Lubow and Ransom, 2009). Lubow and Ransom (2009) reported that the advantages of the photographic mark-resight technique for free-ranging horses are that it can be performed with only one observer, it does not matter if horses are displaced by the aircraft or if a group is encountered repeatedly on the same survey, the technique works in areas with tree cover and complex terrain, and most covariate data are captured in each photograph, so the need to write them down is eliminated. Lubow and Ransom (2009) suggested that the method is likely to produce negatively biased (but quantified) estimates of abundance, and bias probably would increase as the visibility of the horses decreases (for example, more complex topography or more tree cover). Lubow and Ransom noted that it might take several visits to obtain reliable estimates; validation of photographic mark-resight data suggested that it would take six or more occasions in areas that have complex topography and heavy tree cover. According to data collected by Lubow and Ransom (2009) at McCullough Peaks, Little Owyhee, and Pryor Mountain HMAs, the approach provided consistent and reliable estimates of total horse numbers (within 3-9 percent of exact counts). The limitations are that helicopters (which are more expensive to use than fixed-wing aircraft) are usually needed to observe markings in photographs, a high-resolution digital camera with an image-stabilized lens must be used, and it may be difficult to separate horses that have similar coat colors or that are in HMAs that have large numbers of animals (J. Ransom, National Park Service, personal communication, August 10, 2012). This method will probably perform poorly for burros (J. Ransom, National Park Service, personal communication, August 10, 2012).
Simultaneous Double-Count. Two observers independently record the number of animals seen from a given location at the same time. Records are compared to inform population estimates by assessing how many animals or groups of animals are detected by both observers and how many are detected by only one observer or the other (Caughley, 1974a; Ransom, 2012b). The technique can also be used in combination with distance sampling (Kissling and Garton, 2006). It is assumed that observers do not communicate during the observations, that observations are recorded honestly (i.e., it is not a competition), and that transects traveled are uniform, are predetermined, and cover the entire area of interest. The advantages of this method for free-ranging horses are that it provides an estimate of abundance with quantified error and does not require any special equipment. The limitation is that, even with two observers, it is unlikely that it will be sufficient to overcome large biases due to high landscape heterogeneity.
Pre-Gather and Post-Gather Counts. The number of animals captured or removed from the land is used to inform population estimates. This technique can be used when a count has been conducted and is followed soon thereafter by a gather, in which a relatively large proportion of the horses are removed and the quantity is known. Another count is conducted soon after the gather. The difference between the two counts can be used to estimate the detection probability (Eberhardt, 1982).
All the methods except removals or captures can be conducted from the ground or from the air. In ground-based surveys, observers might traverse transects on foot, in vehicles, on horseback, or a combination of the three. Ground-based observers may be in prepositioned, stationary blinds to count animals with the mark-resight or double-observation methods. Cameras can be used to photograph animals at places of common congregation, such as watering holes (Cao et al., 2012; Petersen et al., 2012), and animals can be identified in a series of photographs over time by their markings; this procedure is typically used in a mark-resight analytical framework. Given the sizes of HMAs and their varied topography, it is usually practical and cost-effective to conduct surveys of horses and burros from the air. Helicopters and fixed-wing aircraft are the two aerial survey platforms typically used. In some cases, fixed-wing aerial surveys, which are less expensive than helicopter surveys, are adequate to locate and count animals, especially in areas dominated by sagebrush or other low-growing vegetation. In areas that have higher canopy and cover, however, helicopters may be needed for slower and more careful searching patterns. In aerial surveys, survey methods may be combined. For example, more than one observer may count animals as an aircraft follows a transect pattern by using distance sampling. Transect patterns can also be flown more than once during a survey to increase accuracy of population estimation, assuming that animals do not move substantially relative to flight paths between surveys.
Similarly, the Wild Horse Identification Management System (Osborn, 2004) was established in the Pryor Mountains to enumerate free-ranging horses by using unique coat-color markings and morphological characteristics in photographs. Lubow and Ransom (2009) used this approach in three HMAs (whose horse populations were of known size and were each smaller than 400) that were monitored weekly. Before correcting for detection probability, population size was biased (undercounted) by as much as 32 percent, but estimates accounting for heterogeneity of sighting probability (detection probability) were within 3-29 percent of the true number of animals known to be occupying the areas at the time of the surveys (Lubow and Ransom, 2009). The authors considered the cost of the more accurate models that quantified uncertainty in population-size estimates to be comparable with the costs of raw counts typically used by BLM (Lubow and Ransom, 2009), although the post-processing staff time required can be greater for this technique (Ransom, 2012b).
Remote-Sensing Methods
Remote-sensing technology can be used effectively to locate and count free-ranging horses and burros with a wide variety of sensors on satellites or manned and unmanned aircraft. The sensors can obtain high-resolution images at user-defined times and locations and can capture surface-reflectance characteristics at various spatial resolutions. Manned and unmanned aircraft can also take high-resolution videography that can be used to count horses and assess condition. New technology, including videography that detects movement patterns and measures speed of travel, can sense features with tremendous detail and accuracy. These methods will continue to be developed and improved and will allow even
higher-resolution information with decreased costs. The development of remote-sensing technology to be used with unmanned (drone) aircraft also reduces the risk associated with flying planes and helicopters.
High-resolution remote-sensing imagery can be used to observe unique coat patterns and to detect identifying marks or scars for horse identification. Aerial images taken from manned and unmanned aircraft can produce images with centimeter-level resolution. In addition to color or color-infrared imagery, forward-looking infrared (FLIR) cameras can detect body heat from more than one-fourth of a mile above the ground (Millette et al., 2011). Those cameras have the potential to distinguish horses from the surrounding environment and provide an accurate method for counting animals. Quickbird and Ikonos are satellite sensors that acquire data with resolution of 0.5 to 1 m. These midlevel resolution sensors may be effective for detecting horses and for monitoring change in population densities. Higher-resolution satellite images have been developed and in time will be more readily available.
There are limitations that should be considered when selecting the appropriate remote-sensing platform with respect to estimating populations of free-ranging horses and burros (Millette et al., 2011). First, the spatial resolution of the data must be fine enough to detect individual animals (especially when animals are moving or in a herd) and reduce misidentification with other animal species. Insufficient resolution can be a problem with many satellite-based sensors. Second, data acquisition may be untimely because some technologies rely on orbiting satellites that pass over a given landscape only at intervals of a few days to a few weeks. Third, many remote-sensing technologies are expensive. Fourth, some cameras have too small a field of view and may need to pan back and forth (such as FLIR and handheld cameras). Fifth, the detectability of animals may depend on weather, time of day, vegetation composition and structure, or local topography in a survey area, and quantification of detection probability can be difficult. For example, radiant heat from the earth’s surface (in particular during the daytime) can camouflage the heat produced from a horse or burro when FLIR sensors are used. Sixth, weather patterns, particularly cloud cover, can preclude data collection with many remote-sensing technologies and can add risk to aircraft operators. Finally, current Federal Aviation Administration restrictions limit the use of unmanned aerial vehicles.
Genetic Techniques
A number of studies have used molecular markers to identify animals in noninvasively collected samples to estimate population size. That approach is particularly effective for populations in which individuals are difficult to detect because of vegetative cover or elusive behavior. Traditionally, such populations were surveyed with indirect methods, or indexes, such as sign counts (e.g., feces and tracks), which were corrected for estimates of the rates at which the signs are deposited and decay. In many cases, however, those estimates have relatively large confidence intervals, which limit their usefulness in managing or monitoring populations (Barnes, 2002). For such populations, multilocus genotypes derived from noninvasively collected samples (e.g., feces, hair, and scent marks) have been used as genetic tags for individuals. With a capture-mark-recapture design, populations have been surveyed and the resulting data have been analyzed to estimate population sizes. Genetic tags have advantages over traditional tagging systems in that animals retain their genotypes throughout their lives (thus, tags cannot be lost), and there is no reason to believe that a noninvasively assigned tag will affect the ability to resample the animal (the animals cannot become trap-happy or trap-shy). For dangerous or difficult-to-observe
species—such as bears (Woods et al., 1999; Sawaya et al., 2012), mountain lions (Ernest et al., 2000), tigers (Sugimoto et al., 2012), wolves (Stenglein et al., 2010), coyotes (Kohn et al., 1999), and mountain gorillas (Guschanski et al., 2009)—genetic surveys have provided information about not only population sizes but sex ratios, levels of genetic diversity, and relatedness.
Although to the committee’s knowledge the genetic-tag method has not been used for free-ranging horses, the necessary preliminary work to develop methods of preserving and genotyping DNA from horse dung has been done. There was no need to estimate population size for the Assateague Island National Seashore herd because individual horses are carefully monitored by park management, but the National Park Service sought information about relatedness among individuals to assess and inform its management regime. In a collaborative study with scientists at the Smithsonian Institution, methods of preserving horse dung were tested, and a representative set of microsatellite loci was optimized (Eggert et al., 2010). Potential disadvantages of this method include the time needed for genotyping and data analysis and the difficulties that may be encountered in finding a laboratory willing to conduct the work at a reasonable cost.
Herd Management Area Survey Information Requested and Received by the Committee
The committee initially requested the most recent 12 years of records (2000-2011) on all HMAs so that it could evaluate the methods and procedures used by BLM to estimate sizes of free-ranging horse and burro populations at the time of its study. Because BLM publishes annual national statistics on the numbers of horses and burros on western public rangelands, the committee assumed that requested records would include an estimate of the population of each HMA for each year. Actual surveys of the number of animals occupying a given HMA are usually not conducted annually (BLM, 2010), so the committee expected only a subset of years for each HMA to include records of actual animals counted on the basis of some survey procedure and estimates for the intervening years to be based on previous inventories. For years when counts were conducted, the committee requested the approximate date of the count, the survey platform used (e.g., ground, fixed-wing aircraft, helicopter), and whether the inventory covered the entire HMA or used some sort of sampling regimen whereby a portion of the HMA was surveyed and the results were extrapolated to obtain a population estimate for the entire HMA.
Previous research on techniques for surveying free-ranging horses and burros (Frei et al., 1979; Siniff et al., 1982; Walter and Hone, 2003; Laake et al., 2008; Lubow and Ransom, 2009) and many other large mammal species (Caughley, 1974a; Pollock and Kendall, 1987; Samuel et al., 1987) has demonstrated that not all animals are detected on surveys. Thus, survey results require the estimation of detection probability and adjustment of the number of animals counted to account for the proportion of animals that were undetected. The committee also asked whether the number of animals counted was adjusted to produce the population estimate for a given year. The committee was informed that populations in years in which no counts were conducted were estimated by multiplying the previous year’s population estimate by some assumed population growth rate until another count was conducted (Box 2-1; BLM, personal communication, December 2011). If the HMA had experienced a gather and removal of horses in the intervening year, the number of animals removed was incorporated into the later year’s population estimate. Thus, for years in which no count was performed for the HMA, the committee requested that BLM report
BOX 2-1
Converting Counts to Population Estimates
BLM biologists obtain counts of free-ranging horses and burros to inform management decisions and to monitor equid populations. Counts can be reported directly as a “population estimate” of the animals occupying a given area, or they may be altered on the basis of other information in an attempt to make the estimate more accurate. Research has consistently shown that not all animals are detected and counted when biologists conduct surveys to count them, whether from the ground or with the use of aircraft. If an estimate of the percentage of animals detected is available, the count can be adjusted by that value to obtain an estimate that is a more accurate reflection of the number of animals in the population. For example, if 80 percent of the horses in an area are assumed to have been detected and counted in an aerial survey, this value can be converted into a proportion (0.80) and the count divided by the proportion to obtain a population estimate. The appropriate calculations for the 2 years depicted in Figure 2-1 in which counts were conducted would be
| Year | Count | Estimated Proportion of Animals Detected | Calculation | Population Estimate |
| 2001 | 422 | 0.80 | 422/0.80 | 528 |
| 2004 | 722 | 0.80 | 722/0.80 | 903 |
If a count is not conducted in a given year but a population estimate is still needed, an estimate can be obtained by multiplying the previous year’s population estimate by an estimate of the growth rate of the population. For example, if the horse population is assumed to be growing by 20 percent a year, this value can be converted into a λ value (finite population multiplier) of 1.20 and multiplied by the previous year’s population estimate to project the size of the population in the following year when a count was not conducted. The appropriate calculations for the 2 years depicted in Figure 2-1 in which a count was not conducted would be
| Year | Count | Estimated Proportion of Animals Detected | Calculation | Population Estimate |
| 2002 | 528 | 1.20 | (528)(1.20) | 634 |
| 2003 | 634 | 1.20 | (634)(1.20) | 761 |
If a detection probability or growth rate is used to adjust counts without empirically measuring either quantity, the values may simply be assumptions or “best guesses,” and the adjusted counts would be reported as population estimates with no associated measure of precision. The accuracy of such estimates depends on how closely the assumed detection probability and growth rate reflect the truth, which is probably unknown. There are, however, statistical procedures for obtaining quantitatively rigorous estimates of detection probability, population size, and growth rate on the basis of data, and there are measures of precision of each estimate. When values derived from such rigorous methods are used to adjust counts to obtain a population estimate, the precision of the population estimate can also be determined. Measures of precision are extremely valuable in interpreting estimates of population size and growth rate. A common way to convey precision of a population parameter (population size or growth rate) is to report a 90-percent confidence interval (CI) for the parameter such that there would be 90-percent probability that the real value of the parameter lies within the interval (Williams et al., 2001). For example, one population estimation method (method 1) may provide a population-size estimate of 700 horses with an associated 90-percent CI of 680–720 horses. A second method (method 2) may yield the same population-size estimate of 700 horses, with an associated 90-percent CI of 500–900 horses. In that hypothetical example, the estimate of population size obtained with method 1 is said to be more precise than that obtained with method 2 because method 1 provides a relatively narrow CI. Whenever possible, a population estimation method that provides a more precise estimate is desirable in that one can have more confidence that the population estimate is a better approximation of the true number of animals occupying the survey area than a less precise estimate.
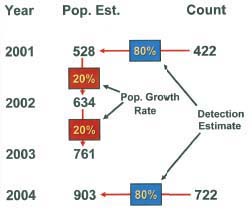
FIGURE 2-1 An example of how periodic counts of free-ranging horses on an individual Herd Management Area could be converted to estimates of population size by applying estimates of detection probabilities and the annual growth rate of the population. NOTE: In this fictitious example, aerial counts conducted in 2001 and 2004 were used to obtain population estimates on the basis of estimates of (or assumptions about) the detection probability (proportion of horses detected on the surveys) and the growth rate of the horse population. The example assumes no horse removals during the 4-year period. If a removal had occurred, the number of horses removed would be subtracted for the appropriate year to obtain the next year’s population estimate.
the growth rate that was applied to obtain the population estimate with the removal data provided in a separate master database.
The committee was informed by the national Wild Horse and Burro Program office that the HMA-specific data requested were not aggregated into a central database but were dispersed among the BLM field offices. It was suggested that a more manageable request for BLM personnel would be that the committee receive a sample of HMA data from a maximum of 40 HMAs. BLM provided a list of the 179 HMAs distributed among 10 western states, with associated data on AML and the current population estimate for each HMA, to aid the committee in selecting a sample of HMAs. The committee excluded HMAs for which the AML was zero, current population estimates were zero, or where reported numbers reflected a mix of burros and horses. To increase the uniformity of the data, HMAs that had burros (and no horses) were not included. The remaining 142 HMAs contained only horses and were ordered by the current population estimate, ranging from 5 to 1,355 (Figure 2-2; Appendix E, Table E-2). Of the 142 HMAs, the committee excluded the ones that had estimated populations of 50 or fewer because the small populations represent less than 3 percent of the horses on western rangelands. From the remaining HMA list, every third one was then selected to obtain a sample distributed evenly over the range of population sizes that occur on BLM-administered lands. That process resulted in a sample of 36 HMAs. The committee subjectively added four other HMAs that had been included
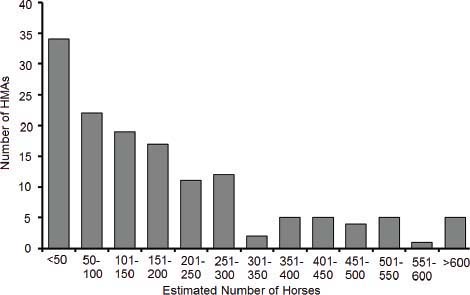
FIGURE 2-2 Distribution of 142 Herd Management Areas (HMAs) that contained free-ranging horse populations of various herd sizes.
NOTE: Population estimates were based on surerve recoreds available as of February 2011.
DATA SOURCE: Based on information provided to the committee by the Bureau of Land Management in December 2011.
in earlier research on population dynamics of free-ranging horses in the western United States (Eberhardt et al., 1982; Garrott et al., 1991b), and that brought the sample to 40 HMAs (Table 2-1; Appendix E, Table E-3). The committee received the data that it requested on all 40 HMAs. The assessment of methods used by BLM to obtain field counts of horses and estimates of population size is based information on the 40 HMAs provided to the committee by BLM. The committee sought to provide a synthetic overview of the horse inventory methods used by BLM; nonetheless, it recognized that its assessment, summarized in the following, may not accurately reflect how horses are counted or population sizes estimated on every HMA.
Assessment of Horse-Count Data for the Sample of Herd Management Areas
The frequency with which surveys were conducted to count horses in each HMA in the sample was highly variable. Among the 40 HMAs surveyed, four reported counting horses no more than once a decade, nine counted horses an average of every 3-4 years, five counted horses an average of 2 of every 3 years, 17 about every other year, and five every year. In HMAs in which horses were not counted every year, there was no discernible pattern in the interval between counts. Information on the methods used for each reported count was frequently unreported (Tables 2-2 to 2-4).
TABLE 2-1 Distribution of 142 Herd Management Areas (HMAs) among Western States That Contained Only Horses and were Actively Managed by the Bureau of Land Management and Distribution of the Sample of 40 HMAs Used by the Committee to Evaluate Wild Horse and Burro Program Methods for Surveying Horse Abundance and Estimating Population Sizes
| State | Number of HMAs Available | Number of HMAs in Sample |
| Arizona | 1 | 0 |
| California | 15 | 2 |
| Colorado | 4 | 3 |
| Idaho | 6 | 1 |
| Montana | 1 | 1 |
| Nevada | 63 | 21 |
| New Mexico | 2 | 0 |
| Oregon | 17 | 6 |
| Utah | 17 | 2 |
| Wyoming | 16 | 4 |
| Total | 142 | 40 |
TABLE 2-2 Example of Horse Inventory Data on Little Book Cliffs Wild Horse Range in Colorado, Showing Routine and Methodologically Consistent Annual Surveys
| Year | Annual Population Estimate | Population Count | Date of Count | Type of Craft | Percentage of Area Inventoried | Method | Adjustment of Count |
| 2000 | 153 | 153 | August | Vehicle/Horse | All | Visual | None |
| 2001 | 169 | 169 | August | Vehicle/Horse | All | Visual | None |
| 2002 | 195 | 195 | August | Vehicle/Horse | All | Visual | None |
| 2003 | 154 | 154 | August | Vehicle/Horse | All | Visual | None |
| 2004 | 178 | 178 | August | Vehicle/Horse | All | Visual | None |
| 2005 | 132 | 132 | August | Vehicle/Horse | All | Visual | None |
| 2006 | 144 | 144 | August | Vehicle/Horse | All | Visual | None |
| 2007 | 165 | 165 | August | Vehicle/Horse | All | Visual | None |
| 2008 | 122 | 122 | August | Vehicle/Horse | All | Visual | None |
| 2009 | 133 | 133 | August | Vehicle/Horse | All | Visual | None |
| 2010 | 138 | 138 | August | Vehicle/Horse | All | Visual | None |
| 2011 | 142 | 142 | August | Vehicle/Horse | All | Visual | None |
NOTE: The number of horses counted in complete surveys of the HMA is the same number reported for population estimates; thus, it is assumed that all animals were detected during the surveys.
SOURCE: Survey response from the Bureau of Land Management, February 2012.
Assuming that the reported data on the 40 sampled HMAs generally represent the procedures routinely used by BLM to enumerate horses on all HMAs, the committee made several generalizations about counts on all HMAs.2 Most counts are obtained with aerial surveys in which an entire area is surveyed in an effort to obtain a complete count of the horses occupying an HMA with no attempt to apply sampling methods or to estimate the proportion of animals that were undetected. A helicopter was the preferred aircraft, although fixed-wing aircraft were also frequently used. Some surveys were conducted from
____________
2 The data supplied by BLM for all 40 HMAs can be retrieved from the study’s public access file. To obtain the information, contact the National Research Council’s Public Access Records Office at paro@nas.edu.
TABLE 2-3 Example of Horse Inventory Data on Reveille Herd Management Area in Nevada, Showing Irregular and Inconsistent Survey Methods
| Year | Annual Population Estimate | Population Count | Date of Count | Type of Craft | Percentage of Area Inventoried | Method | Adjustment of Count |
| 2000 | 164 | 190 | November | Helicopter | 100 | Grid | None |
| 2001 | 187 | ||||||
| 2002 | 96 | ||||||
| 2003 | 111 | 9 | December 23 | Fixed-Wing Airplane | 70 | Grid | None |
| 2004 | 61 | 61 | October 15 | Helicopter | 100 | Grid | None |
| 2005 | 71 | ||||||
| 2006 | 135 | 119 | January 6 | Helicopter | 100 | Grid | None |
| 2007 | 57 | 79 | January 7 | Helicopter | 100 | Grid | None |
| 2008 | 66 | ||||||
| 2009 | 77 | 213 | September 9 | Helicopter | 100 | Grid | None |
| 2010 | 213 | 231 | February 10 | Helicopter | 100 | Grid | None |
| 2011 | 91 | ||||||
NOTE: These data provide an example of the difficulty of understanding how annual population estimates were derived from the survey count data.
SOURCE: Survey response from the Bureau of Land Management, March 2012.
TABLE 2-4 Example of Horse Inventory Data on Desatoya Herd Management Area in Nevada, Showing Irregular and Inconsistent Survey Methods and Incomplete Records
| Year | Annual Population Estimate | Population Count | Date of Count | Type of Craft | Percentage of Area Inventoried | Method | Adjustment of Count |
| 2000 | 304 | August | Jet Ranger | 100 | Direct | ||
| 2001 | 294 | December 1 | 80 | Direct | |||
| 2002 | |||||||
| 2003 | |||||||
| 2004 | February 4 | ||||||
| 2005 | |||||||
| 2006 | |||||||
| 2007 | 238 | April 7 | Jet Ranger | 100 | Direct | ||
| 2008 | |||||||
| 2009 | |||||||
| 2010 | 434 | April 10 | Jet Ranger | 100 | Direct | ||
| 2011 | 543 | July 11 | Jet Ranger | 100 | Direct | ||
NOTE: These data provide an example of the difficulty of understanding how annual population estimates were derived from the survey count data.
SOURCE: Survey response from the Bureau of Land Management, March 2012.
the ground on foot, in vehicles, on horseback, or with a combination of the three. Ground-based surveys appeared to be performed primarily in states that had relatively few HMAs and in which the total number of horses on an HMA was low (under 150). It was also common for reported counts to be attributed to gather operations. Explanations were not provided for the individual gather-based counts, but the committee’s best understanding of such counts was that a gather operation was conducted on an HMA for the purpose of removing horses from the rangeland. The gathers were assumed to have captured all horses on an HMA, and the reported count represented the number of captured horses that were
released back onto the rangelands. Although survey methods used for some HMAs in the sample that the committee examined appeared to be consistent with respect to time of year and survey platform, the timing of surveys on many of the reviewed HMAs were inconsistent; they were often distributed over 6-9 months of the year, and this led the committee to conclude that such practices are common. It was also relatively common in the sample of HMAs that the committee examined for survey methods to differ from count to count on a given HMA—some counts were performed from helicopters, others from fixed-wing aircraft, and others from gathers.
It was difficult for the committee to understand the rationale for the timing and distribution of counts for the sample of HMAs, but it did detect what appeared to be a pattern related to timing of gathers and horse removals (provided to the committee in separate files by the national office). A common pattern observed in the HMA records was a report of a complete count followed by a variable period of years in which no counts were performed and then a report of another count immediately before a major gather and horse removal. On some occasions, a follow-up count was reported immediately after a removal. On the basis of recommendations of the National Research Council Committee on Wild and Free-Roaming Horses and Burros (NRC, 1982), BLM-published procedures for surveying and counting free-ranging horses (BLM, 2010), correspondence with Wild Horse and Burro Program administrators, and a review of a sample of HMA environmental assessment documents prepared for horse removals, the committee interpreted that pattern as reflecting a need to have a recent count of horses on an HMA before a removal. Thus, the committee speculates that after a period of no counts, when a population was assumed to have increased, expertise of the local manager indicated that the horse population needed to be reduced, and a count was conducted to determine whether the population was over the AML. If the count was sufficiently higher than the established upper bound of the AML, an environmental assessment was prepared, and a gather and removal occurred. Post-removal counts were often recorded as gather counts; this suggests that the gather was assumed to have captured all horses on the HMA and that the count reflected the number of horses that were released back onto the HMA. Alternatively the post-removal counts may have reflected a combination of the number of horses released and some estimate of the number on the HMA that remained uncaptured. It was unclear from the data that the committee received which of those assumptions was made by BLM managers for individual records.
Relationship Between Direct Counts of Horses and Reported Herd Management Area Population Estimates
All annual population estimates for the 2000-2011 period requested were provided for 24 of the sample of 40 HMAs; no estimates were provided for five HMAs, and estimates for the other 11 were incomplete (generally, less than 50 percent of the estimates were provided). No reported population estimates included associated measures of precision. The committee assumed that all population estimates were derived in some fashion from survey count data (as described and illustrated in Figure 2-1), and the description of the process used to develop annual population estimates provided by the national Wild Horse and Burro Program office supports this assumption:
When the data [annual HMA population estimates] are updated for any given year the starting point is the previous year’s population estimate. These data are updated based on the following: 1) removals (gathers) that have been conducted since February 28th of the previous year, 2) new population surveys (census) that have been conducted since February 28th of the previous year and 3) when no population surveys were conducted
in the previous year, the previous year’s data are increased to account for the year’s foals based on historical experience regarding annual population increase typical of that HMA (normally about 20% if a gather/removal had not been conducted). When no population survey has been conducted consideration is also given to the estimated effects of any fertility control vaccines that have been previously administered. (BLM, email communication, May 2, 2012)
As mentioned, most population estimates reported in years when counts were conducted for the 40 sampled HMAs simply reported the number of animals counted without adjustment for the proportion of undetected animals or measures of precision. In the few instances in which a population estimate for a given year was higher than a count in the same year, there was little notation to indicate that the difference was due to application of a detection probability adjustment. In cases in which it seemed plausible that that occurred, the committee calculated the assumed detection probability by dividing the annual count by the population estimate; the resulting values of assumed detection probability generally ranged from 0.7 to 0.9. However, there were substantial records for the sampled HMAs of reported population estimates that were considerably smaller than the counts in those years when there were no records of horses being removed. There were also instances in which population estimates were much higher than reported counts but with no explanation for the differences.
The methods used to estimate population sizes in years in which no counts were conducted were seldom noted in the records provided to the committee. When records clearly stated that an assumed population growth rate was applied to the previous year’s population estimate, an annual growth rate of 20 percent was generally used. As with detection probability, when it seemed plausible that a population growth rate was used to project population estimates for years in which no counts were conducted, the committee calculated the assumed growth rate by dividing the second year’s annual population estimate by the previous year’s annual population estimate to obtain an estimate of λ, that is, the population growth rate. The resulting values (reported as percent growth) were variable, generally ranging from 3 to 38 percent; values of 15 to 25 percent were most common. A substantial proportion of the population estimates reported for years in which no counts or gathers were conducted, however, diverged enough from the estimates reported both immediately before and after that, without further explanations, the committee could not understand how such values were obtained. One plausible explanation for at least some of those cases is that horses were freely moving on and off HMAs.
Relationship Between Direct Counts of Horses and National Population Estimates
National statistics that provide estimates of the total number of free-ranging horses and burros on public rangelands are published annually in BLM reports and on the Wild Horse and Burro Program website. Those are important statistics because they are interpreted by various public constituencies to gauge the success of the program’s management, are used in formal government reviews of the program (NRC, 1980, 1982; GAO, 2008; OIG, 2010), and are foundational data for planning and budgetary documents, such as BLM’s Proposed Strategy: Details of the BLM’s Proposed Strategy for Future Management of America’s Wild Horses and Burros (BLM, 2011). The procedure used to generate the annual state and national estimates was described to the committee as follows:
Each year shortly after February 28th, field offices submit updated estimates for each HMA to the National Program Office. These field submissions are compiled into one national
report that lists new estimates for each HMA and that is organized by state. (BLM, email communication, May 2, 2012)
Given the incompleteness of the counts and population estimates that the committee received for the sample of 40 HMAs, which came from the field offices, it was not clear how the national statistics could be calculated. Therefore, the committee requested the series of HMA estimates that were reported to the national office from the field offices and used in generating the state and national estimates for the most recent 5-10 years. In response to its request, the committee was pointed to the national HMA-specific estimates for fiscal years 2005-2011 that were posted on the program’s website.3 The committee also received files with earlier national HMA estimates from fiscal years 2000-2004. However, the corresponding information that the national office received from the field offices to generate the published estimates was not provided to the committee. The committee was informed that that information was discarded after the annual national statistics were published (BLM, personal communication, May 2012). Thus, the committee received no documentation linking the national statistics to information reported from the field offices. It was not clear whether the information from the field offices was modified by some procedure at the national office before publication on the program’s website, but various correspondence with personnel at the national office suggested that some changes were made. That impression was reinforced when the committee compared the national HMA-specific population estimates with those reported by the field offices for the sample data on 40 HMAs provided to the committee. The committee found that a substantial proportion of the HMA estimates published by the national office did not correspond to the ones the committee received from the field offices; discrepancies ranged from modest to many hundreds of animals. In addition, all HMAs in the reported national statistics had a population estimate for all years, whereas a substantial proportion of the HMA records that the committee received from the field offices had no population estimates reported for some of the years.
Evaluation of Current Methods for Enumerating Free-Ranging Horse Populations
The sample of HMA records made available to the committee and examined with the evaluation of the national population statistics indicates that robust inventory procedures were adhered to on few HMAs during the most recent decade of population monitoring. The committee identified five primary weaknesses in inventory procedures: inconsistent methods, likely movement of horses among HMAs, little or no effort to quantify detection probability and apply corrections accordingly, no attempt to quantify precision of abundance estimates, and inadequate record-keeping and database management. It is reasonable to expect that different survey techniques may be optimal in inventorying animals depending on attributes of individual HMAs, such as the size of the equid population and of the area, accessibility, distinctiveness of individual horses, ruggedness of topography, and presence of tree cover. Once a survey method is determined for an HMA or HMA complex, however, it should be used consistently so that variation in the number of animals counted from one survey to the next can be reasonably attributed to population changes and is not confounded by the use of different techniques. The most prevalent problems that the committee identified in that regard were inconsistency in the timing of surveys and in the survey platform used (fixed-wing, helicopter, ground-based, or gathers). Movement
____________
3 Available online at http://www.blm.gov/wo/st/en/prog/whbprogram/herd_management/Data.html/. Accessed November 20, 2012.
of horses across HMA boundaries can seriously confound interpretation of changes in the numbers of animals counted from one survey to the next. Although there appear to be few data on this issue, field personnel recognize it as a common problem, and relatively large changes in the numbers of animals counted in consecutive surveys may be reasonably attributed to movement of animals on and off HMAs. It was not clear to the committee whether data on spatial distribution of animals are routinely collected during inventory surveys. Information on where animals are observed can provide important insights into habitat use and resource selection by free-ranging equids, which in turn would contribute to a better understanding of competition with livestock and wildlife and assist in decisions on forage allocation and other issues related to rangeland health (see Chapter 7).
It is also well documented that the types of survey methods used for counting free-ranging horses and burros are imperfect in that various proportions of animals will not be detected in any given survey and detection probability can vary over time and space. Evidence clearly indicates that, under some conditions that are common for rangelands occupied by free-ranging horses and burros, the proportion of animals missed can be substantial. Thus, the routine reporting of the uncorrected counts as population estimates results in inventory numbers that are systematically biased low. Finally, the apparent difficulty of meeting data requests from the committee, the incompleteness of many of the records provided to the committee, and the lack of data supporting the national population statistics indicate deficiencies in the routine documentation of survey efforts and results and in database management. Many of the same issues were also identified by the National Research Council Committee on Wild and Free-Roaming Horses and Burros, which reviewed similar records near the start of the Wild Horse and Burro Program over 30 years ago (NRC, 1980, 1982).
Initiatives to Improve Methods for Enumerating Free-Ranging Horse Populations
At the time this report was written, BLM had initiated a number of actions aimed at improving the rigor, reliability, and utility of the procedures used to estimate the abundance of free-ranging horses and burros. First, in its Wild Horses and Burros Management Handbook (BLM, 2010), BLM provided guidelines for survey techniques used to enumerate free-ranging horses and burros, stating
- The target interval for conducting population surveys is every 2 years, as recommended by the National Research Council Committee on Wild and Free-Roaming Horses and Burros (NRC, 1982).
- Techniques that provide sightability and detection corrections are to be used.
- Survey methods and timing are to be consistent.
- All details of each survey are to be recorded and permanently on file.
- Survey data are to be entered into a centralized database (the Wild Horse and Burro Program System).
The committee readily endorses those guidelines. Adherence to them will greatly improve the utility of equid population estimates.
Second, in response to the widely held perception that free movement of animals among adjacent HMAs confounds inventory procedures and reduces the ability to interpret counts, managers have subjectively assessed their knowledge of equid movements among adjoining HMAs and aggregated 93 of 179 HMAs into HMA “complexes.” Each complex is composed of two to six areas managed for equids; many HMAs are managed with adjacent
U.S. Forest Service Wild Horse (or Burro) Territories. The goal is to coordinate surveys, gathers, removals, and other management actions among HMAs within a designated complex and thus to manage all horses in a complex as a single biological population (BLM, 2010). The committee thinks that that procedural change has the potential to improve interpretation of counts substantially, although the degree of improvement hinges critically on how often and how many animals move across HMA boundaries. Conducting aerial inventories over large areas, however, has its own set of challenges. The committee had no knowledge of implementation at the field level with respect to the coordination of population surveys and management actions (removals) among HMAs within designated complexes.
Third, for over a decade, scientists from the U.S. Geological Survey (USGS) Fort Collins Science Center have conducted research and provided scientific support to the Wild Horse and Burro Program. In 2004, an Aerial Survey Work Plan was developed and field research was implemented to develop and test improved techniques for inventorying free- ranging horses from both helicopter and fixed-wing survey platforms. Several methods were evaluated including various mark-resight techniques, distance sampling (transects), and sightability models. The research reaffirmed that substantial proportions of horses are not detected in aerial surveys and that detection is poorer in more rugged and tree-covered terrain than in flatter, more open landscapes (Lubow and Ransom, 2009). Several of the mark-resight techniques evaluated were successful in accounting for varied detection probability and for providing estimates close to the known number of horses in the experimental populations. Less successful techniques that were evaluated included distance sampling, forward-looking infrared technology, and remote sensing. In addition, GPS mapping technologies were incorporated into all aerial survey procedures and provided data on animal distributions and patterns of resource selection. At the time this report was written, the USGS team had trained eight BLM personnel in the new survey methods, and they had started to conduct rigorous surveys on 35 HMAs in seven states. In August 2012, BLM hired a national aerial-survey coordinator to continue to implement reforms in the inventorying procedures (BLM, 2012).
The committee encourages BLM to continue such collaboration and reform of its procedures. Those actions and adherence to the survey guidelines in the 2010 handbook will improve the accuracy and defensibility of its population estimates. More robust and transparent data may also improve its relationship with stakeholders (see Chapters 7 and 8).
The change in abundance of a population over some period is generally known as a growth rate. Understanding growth rates is important for efficient and effective management of free-ranging equid populations. Knowing population growth rates gives managers the ability to project how quickly populations will increase and when management actions (such as removals or fertility treatments) need to be applied. They are also key information for determining the magnitude of fertility treatments needed to reduce population growth rate to some desired level and, after treatment, to evaluate whether the intervention had the expected effect. Growth rate estimates are used to estimate the size of populations in years in which counts will not be conducted (Figure 2-1), as estimates are needed for inventory, management, and planning purposes.
Like populations of most other terrestrial mammals in North America, free-ranging horse population dynamics have a seasonal cycle in which animals are added to the population by births during a relatively short interval in the spring and animals are removed from the population through deaths (and management removals) throughout the year. The
natural interval for estimating population growth rates is the year. Each species has an inherent maximum population growth rate that is dictated by its life-history characteristics, including how often animals can reproduce, the number of young produced per reproductive event, the age at which animals become reproductively mature, and the death rates for the various age classes of animals.
There was essentially no knowledge of free-ranging horse population dynamics and growth rates when the populations received federal protection in 1971. During the decade that followed, federal land-management agencies, primarily BLM and the U.S. Forest Service, began to inventory horse and burro populations, and a number of studies of horse demography were undertaken. Scientific demographic investigations of free-ranging horses, however, were limited to three 1- to 2-year studies of western herds (Feist and McCullough, 1975; Nelson, 1978; Boyd, 1980), two studies of herds on barrier islands on the Atlantic coast (Welsh, 1975; Keiper, 1979), and a study of ponies in Britain (Tyler, 1972). A review of the studies by the National Research Council Committee on Wild and Free-Roaming Horses and Burros (NRC, 1980) and novel analyses conducted by that committee revealed ambiguities. The committee noted that the 16- to 22-percent annual growth rate estimates derived from direct counts conducted by management agencies were notably higher (by up to 10 percent) than estimates obtained with population models (Conley, 1979; NRC, 1980, 1982; Wolfe, 1980) that used the best available survival and fecundity data on domestic horses and from the few studies of free-ranging horse populations. A good under standing of demographic processes can contribute substantially to the effectiveness of programs designed to manage wildlife populations; the 1980 National Research Council report recommended additional research on demography, and considerable progress has been made on horses. There have been a few studies of feral burro demography in Australia (Freeland and Choquenot, 1990; Choquenot, 1990, 1991), but little is known about the demography of free-ranging burros in the western United States. Because key aspects of burro life-history characteristics and their ecological niche differ from those of horses, this committee recommends separate studies on burro population growth rates.
Population Growth Rate Estimates Based on Counts
The most direct method for estimating growth rate of a population is to obtain counts or population estimates over multiple years. If the population is growing at a relatively constant rate over the period for which counts or estimates of abundance are available, the abundance values, when log-transformed, will be approximately linear. Linear-regression techniques can be used to fit a line to the data, and the estimated slope of the line provides an estimate of the instantaneous growth rate of the population, denoted by r (Caughley, 1977; Eberhardt, 1987). The procedure also provides an estimate of the precision of r. The slope estimate can be back-transformed (exponentiated) to obtain the finite population growth rate, denoted by λ. The λ value is also referred to as the population multiplier in that one can multiply a population estimate (or count) in a given year by λ to obtain an estimate of the number of animals in the population a year later (see Box 2-1 for an example). When λ is 1, the population is stable or unchanging; when λ is over 1.0, the population is increasing; and when λ is under 1.0 the population is decreasing. Thus, a λ of 1.03 indicates that a population is growing by 3 percent a year, and a λ of 1.20 indicates that a population is growing by 20 percent a year. For consistency in reporting growth rates, the committee used the convention used in BLM documents: reporting growth rates as annual percentages.
A number of studies have used log-linear regression of time series of counts of free-ranging horse populations in the western United States to estimate annual growth rates.
The National Research Council Committee on Wild and Free-Roaming Horses and Burros (NRC, 1980) calculated a weighted mean of 16 percent for aerial count data on 25 HMAs in five states. Wolfe (1980) used count data on 12 HMAs in six states and calculated values ranging from 8 to 30 percent and an unweighted mean of 22 percent but in a later publication suggested a typical growth rate of 15 percent for western U.S. herds (Wolfe, 1986). Counts of two Oregon horse herds were used by Eberhardt et al. (1982) to estimate growth rates ranging from 20 to 22 percent. Similarly, Garrott et al. (1991b) estimated growth rates ranging from 15 to 27 percent with a mean of 21 percent for 12 HMAs in four states. Since those studies were published, a number of additional analytical methods have been developed to estimate population growth rates, and associated measures of precision, on the basis of a time series of counts or abundance estimates that can provide enhanced insight into population processes (Dennis et al., 1991, 2006; Humbert et al., 2009). The techniques would be useful in future studies of Wild Horse and Burro Program inventory data.
The Pryor Mountain herd in Montana is perhaps the most well-studied free-ranging horse population in the western United States. The herd’s size (100-200) and the small and traversable geography of the HMA have been conducive to a number of estimates of this population’s growth rate over the last 3 decades. Nearly all animals have been individually identified in the population because of unique color and marking patterns and have been closely monitored each year, so reproduction, mortality, and total number of horses on the range have been known with considerable certainty, and this allows each annual growth increment to be approximated relatively precisely. Under those special conditions, it is reasonable to estimate an annual λ by dividing the count obtained in a given year by the count obtained in the preceding year. Estimating annual growth rates from counts conducted in two consecutive years is not reliable for most free-ranging equid populations because variation in the proportion of animals detected from one count to the next and movement of animals between adjacent HMAs can dramatically bias λ estimates either upward or downward. Those problems are not prevalent in the small, isolated, and intensively studied Pryor Mountain herd, in which annual estimates from consecutive counts can be considered reliable.
Garrott and Taylor (1990) reported an average annual growth rate of about 18 percent in 1977-1986 in the Pryor Mountain herd, and a similar growth rate was reported by Singer et al. (2000) in 1992-1997. More recently, Roelle et al. (2010) reported a temporary decline in the herd’s annual growth rate to about 11 percent. The lower growth rate was attributed at least partly to lower foal survival due to mountain lion predation and possibly the effects of contraceptive treatment of a modest number of mares, but growth had returned to higher rates near the end of their studies (2005-2007) coincident with hunters harvesting several mountain lions from the range. Similar individual-based studies of horse demography conducted in a number of populations occupying barrier islands along the Atlantic coast have documented annual growth rates of 4.3 percent in the Cumberland Island, Georgia, population (Goodloe et al., 2000), about 10 percent in the Assateague Island, Maryland, population (Keiper and Houpt, 1984), and 16 percent in the Shackleford Banks, North Carolina, population (Wood et al., 1987).
Population Growth Rate Estimates Based on Models
A more indirect method for investigating population growth rates of free-ranging horse populations is the construction of population models that use age-specific estimates of horse survival and fecundity rates obtained from field studies. Model-based approaches provide asymptotic or long-term population growth rate estimates that are based on input parameters as opposed to the abundance-based approaches discussed in the previous
section that provide estimates of realized growth rates. Such exercises were initially conducted about 3 decades ago when little demographic information was available to provide a basis for assigning values to parameters in such models (Conley, 1979; NRC, 1980, 1982; Wolfe, 1980). During the decade after those studies, additional information on survival and reproductive rates was published (Seal and Plotka, 1983; Keiper and Houpt, 1984; Berger, 1986; Siniff et al., 1986; Wolfe et al., 1989; Garrott and Taylor, 1990; Garrott, 1991b; Garrott et al., 1991b). Garrott et al. (1991a) used insights from those studies to parameterize the Lotka/Cole equation with a variety of age-specific fecundity and survival schedules to model western free-ranging horse population growth rates. The modeling exercise yielded growth rate estimates of 11-27 percent. Later published studies have provided additional estimates of the range of survival and fecundity rates in specific free-ranging and fenced-in horse populations on western U.S. rangelands (Greger and Romney, 1999; Turner and Morrison, 2001; Roelle et al., 2010) and Atlantic barrier islands (Goodloe et al., 2000) and herds in France (Monard et al., 1997; Cameron et al., 2000), New Zealand (Linklater et al., 2004), Argentina (Scorolli and Lopez Cazorla, 2010), and Australia (Dawson and Hone, 2012). Those studies generally reported survival and fecundity rates within the ranges of those used in earlier population modeling efforts.
When capture-recapture data collected on individually marked horses are available, Pradel’s temporal symmetry models can also be used to estimate realized population growth rate (Pradel, 1996; Williams et al., 2001). That approach allows the estimation of other useful demographic parameters (such as apparent survival and recruitment rates) and the modeling of these parameters as functions of covariates. However, application of the approach requires that horses be individually marked and recaptured (physically or visually) in such a way that the capture history of each animal can be compiled. BLM does not regularly mark horses, and the effort required to describe and catalog unique identifiable natural markings of individual horses in most situations is not practical. Data on several intensively studied horse populations on Atlantic barrier islands and in the western United States are being collected and can be used in those types of models (Goodloe et al., 2000; Turner and Kirkpatrick, 2002; Lubow and Ransom, 2009; Roelle et al., 2010).
Population Growth Rate Estimates Based on Horse Age-Structure Data
Another source of data that was available to the committee to help in gaining insight into the average growth rates of free-ranging horse populations was the age structure of the horses captured and removed from western rangelands. Those data are routinely collected on all horses captured and removed during management gathers; irruption and wear of teeth are used to estimate the age of each horse removed from public rangelands as it was processed before transfer to adoption or holding facilities. The age structure of a population is the result of many interacting population processes, and this complicates interpretation of age-ratio data on individual populations (Caughley, 1974b). However, as discussed earlier in this chapter, analysis of inventory data on free-ranging horse populations and population modeling approaches provided relatively consistent results with respect to the average growth rate of horses on western rangelands. Thus, it is reasonable to use the aggregate age-structure data on horses captured and removed from the rangelands, which are collected independently of the inventory data, in an attempt to corroborate horse population growth rates derived from inventory data.
The committee had access to age data on 167,927 horses captured and removed during 1989-2011; the number of animals captured and removed each year varied from 2,468
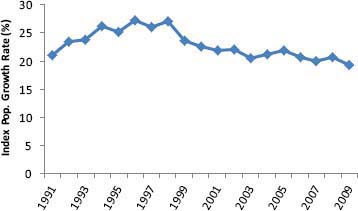
FIGURE 2-3 An index of population growth rate of free-ranging horses based on data on ages of 167,927 horses captured and removed from western rangelands in the United States to manage their abundance.
NOTE: Age-structure data were available for 1989-2011; the number of horses captured and removed each year ranged from 2,468 to 11,416. The index was calculated by dividing the number of young-of-the-year horses by the total number of horses 1 year of age and older in a sample of horses captured and removed from rangelands and then multiplying the result by 100 to obtain a percentage. A 5-year moving average was used to calculate a growth rate index; the annual values plotted in the graph were derived from the age data from a given year, the 2 preceding years, and the 2 following years.
to 11,416.4 A reasonable index of the average growth rate of horses on western rangelands can be calculated by dividing the number of young-of-the-year horses (that is, horses less than 1 year old) by the total number of horses 1 year of age and older in a captured-and-removed sample and multiplying the result by 100 to obtain a percentage. The committee used a 5-year moving average with the 1989-2011 dataset of ages of captured and removed horses when calculating the index to have a large sample of captured and removed horses that would be characteristic of the diverse ecological settings of western rangelands and to reduce variation due to the particular subset of horse populations gathered in any given year. The growth rate index generally was 20-25 percent with some indication of a modest increase during the 1990s; but during the most recent decade, the growth rate index was relatively stable or perhaps experienced a slight decline (Figure 2-3).
The age-structure data would need to have come from horses captured and removed immediately before the birth pulse for the index to reflect realized growth rates of the free-ranging horse populations accurately and thus to account for all deaths of horses over the year after the birth pulse. Gathers, however, occurred throughout the year and were most concentrated in August–February. The index therefore probably overestimates growth rates to some extent. It is difficult to estimate the magnitude of the bias, but, on the basis of the available literature on timing and extent of mortality of horses, the committee believes that the bias is modest.
____________
4 The data supplied by BLM for the removed animals can be retrieved from the study’s public access file. To obtain the information, contact the National Research Council’s Public Access Records Office at paro@nas.edu.
The index also assumes that the age distributions of horses captured and removed from rangelands were representative of the age structure of the free-ranging populations. A bias could have been introduced into the age-structure data of captured and removed horses if managers tended to remove the youngest horses, which were more easily adopted, and to return older, less adoptable horses to the rangelands. Such a practice, if widespread, would inflate the index and suggest that population growth rates were higher than what were actually realized. The committee had no way to evaluate such a potential bias directly. It did, however, review preliminary environmental assessments of a sample of recent HMA gather plans to gain some insight into the potential bias in the age-structure data. Age-selective removals were nearly always considered in the gather plans that the committee reviewed, but the preferred (proposed) actions often did not involve age-selective removals. The committee also noted that in a number of the environmental assessments that presented the history of gathers, usually no captured horses were returned to the rangelands. For gathers in which some captured horses were released, the number of horses returned to the rangeland generally constituted a small proportion of the total number of animals captured. In addition, diverse reasons for the selection of horses to be released were stated, including considerations of conformation, coat color and marking patterns, and mares that were treated with a contraceptive vaccine. It was also stated that horses were selected for release to “maintain a diverse age structure.” Thus, the committee found little evidence to suggest an overt and consistent bias in the age structure of horses that were removed from rangelands and concluded that the age-structure data can provide a reasonable assessment of the general growth rate of the free-ranging horse populations on public rangelands in the western United States.
The committee concludes that the population growth rate index derived from the age structure of captured and removed horses is generally consistent with the herd-specific population growth rates reported in the literature. That suggests that a mean annual population growth rate in the free-ranging western horse population approaching 20 percent is a reasonable approximation.
From its review of the information provided by BLM on population-survey methods, approaches to data collection and population estimation, and records on horse removals and the committee’s review of the relevant literature on estimating ungulate populations and population growth rate, the committee draws the following conclusions.
Estimating the Size of Free-Ranging Equid Populations
Management of the nation’s free-ranging horses and burros should be based on rigorous population-monitoring procedures that are consistently applied by all BLM field offices. The methods reviewed by the committee for monitoring animal numbers on a small subset of HMAs may be adequate, but all reviews of the procedures routinely used by BLM to survey free-ranging horses and burros since the inception of the Wild Horse and Burro Program have identified substantial methodological flaws. On the basis of the information that was reviewed, the committee concluded that many of the shortcomings identified in previous reviews have persisted. At the time that the committee completed its review, inventory methods and statistical tools common to modern wildlife management were used to count horses on only a few HMAs. In addition, survey methods used to obtain
sequential counts of horse populations on an HMA were often inconsistent and generally poorly documented.
Initiatives to improve population monitoring have, however, been implemented in recent years. Aggregating neighboring HMAs on which free movement of horses is known or likely into HMA complexes for the purposes of coordinating population surveys, removals, and other management actions is an important step that can improve data quality and interpretation and enhance population management. The committee commends the partnership between BLM and USGS to develop rigorous, practical, and cost-effective survey methods that account for imperfect detection of animals. The committee strongly encourages BLM to continue that collaborative research effort to identify and refine a suite of survey methods that are effective for the varied landscapes occupied by horses and burros. Transferring the resulting knowledge to those in the field offices responsible for routine monitoring of populations is essential if the reforms are to be institutionalized.
Once more rigorous survey methods are adopted, they need to be standardized and consistently used, as dictated in the Wild Horses and Burros Management Handbook (BLM, 2010). The committee reaffirms the recommendations of a previous National Research Council committee that annual population surveys are not required to adequately monitor and manage free-ranging horse and burro populations. BLM, however, should develop protocols for how frequently surveys are to be conducted and ensure that the resources are available to field personnel to maintain a standardized survey schedule. Consideration should also be given to identifying a subset of HMAs that typify the diverse ecological settings throughout western rangelands that can be used as sentinel populations in which detailed demographic studies are conducted annually to assess population dynamics and responses to changes in animal density, to management interventions, and to variation in seasonal weather and potential trends in climate. Record-keeping needs to be substantially improved; the committee recommends that the Wild Horse and Burro Program develop a uniform relational database—that is accessible to and used by all field offices—for recording all pertinent population survey data.
On the basis of the information provided to the committee, it cannot consider the national statistics scientifically rigorous. The data used in the national statistics are the HMA counts that the committee assumes are converted to population estimates for each year in which counts are conducted, and the counts are extrapolated to produce population estimates in later years in which counts are not conducted (Figure 2-1). The procedures used for developing annual HMA population estimates from counts are not standardized and often are not documented, but it seems clear that the national statistics are the product of many hundreds of subjective and probably independent judgments and assumptions by range managers and administrators about the proportions of horses counted in surveys, population growth rates, effects of management interventions, and potential animal movements between HMAs. Perhaps most important, the links between the national statistics and actual population-size surveys, which are the foundational data of all estimates (whether derived at the field-office or national level), are obscure. Thus, the procedures and processes used by the Wild Horse and Burro Program to generate the national statistics impart a large measure of uncertainty in the numbers and their interpretation. Development of a uniform and centralized relational database that captures all inventory and removal data generated at the level of the field offices and animal processing and holding facilities and that is used by the national office to generate annual program-wide statistics would provide a clear connection between the data collected and the reported statistics. The committee also suggests that the survey data at the level of the HMA and any procedures used to modify the survey data to generate population estimates be made
readily available to the public to improve the transparency of and public trust in the management program (see Chapters 7 and 8).
In addition to the methodological shortcomings of BLM’s current animal inventory and data-management procedures, it is the committee’s judgment that the reported annual population statistics are probably substantial underestimates of the actual number of horses occupying public lands inasmuch as most of the individual HMA population estimates are based on the assumption that all animals are detected and counted in population surveys—that is, perfect detection. A large body of scientific literature focused on inventory techniques for horses and many other large mammals clearly refutes that assumption and shows estimates of the proportion of animals missed on surveys ranging from 10 to 50 percent depending on terrain ruggedness and tree cover (Caughley, 1974a; Siniff et al., 1982; Pollock and Kendall, 1987; Garrott et al., 1991b; Walter and Hone, 2003; Lubow and Ransom, 2009). The committee has little knowledge of the distribution of HMAs with respect to terrain roughness and tree cover, but a reasonable approximation of the average proportion of horses undetected in surveys throughout western rangelands may be 0.20 to 0.30. If those proportions are applied to the 2012 population estimate of 31,453, the national statistic would need to be adjusted to 39,316–44,933. The conclusion by this committee that there are considerably more horses on public rangelands in the western United States than reported in the Wild Horse and Burro Program national statistics was also reached by an earlier National Research Council committee (NRC, 1980, 1982) and by the Government Accountability Office (2008).
Population Growth Rates
The earlier National Research Council committee questioned claims of population growth rates in free-ranging horses on western rangelands exceeding 5-10 percent (NRC, 1980), but adequate studies conducted since then have clearly demonstrated that growth rates approaching 20 percent or even higher are realized in many horse populations. That conclusion is corroborated by studies of survival and fecundity rates and reinforced by population models that integrated these estimates to project growth rates. It is more difficult to estimate the typical or average population growth rate in western horse populations inasmuch as such an assessment would require estimating growth rates in an adequate representative sample drawn from all horse populations managed by BLM. Although the literature provides a relatively large number of growth rate estimates, the studied populations constitute a sample of convenience in that they were selected simply because data for estimating growth rates were available or there was specific scientific or management interest in particular populations. Those studies collectively demonstrate that growth rates vary substantially from one population to another and may also vary from one period to another in the same population.
The age-structure data on animals removed from the range probably provide the most representative sample in that the data were collected over several decades, involved multiple management gathers from a large proportion of HMAs, and involved large numbers of animals. Those data also provided a relatively consistent estimate of the proportion of young-of-the-year animals in free-ranging populations that is consistent with the generally high growth rates documented for individual herds that were based on direct counts. It is also to be expected that most free-ranging horse population growth rates are close to the biological potential for the species, given the general management policy of periodically removing relatively large proportions of populations to meet AML goals, which, in turn, were established at least partially to ensure that horses were not routinely food-limited (see
Chapters 3 and 7). On the basis of the published literature and the additional management data reviewed by the committee, the committee concludes that it is likely that most free-ranging horse populations on public rangelands in the western United States are growing at an annual rate of 15-20 percent.
Consequences for Management
The committee’s conclusions that there are substantially more horses on public range-lands than reported and that horse populations generally are experiencing high population growth rates have important consequences for management. Population growth rates of 20 percent a year would result in populations doubling in about 4 years and tripling in about 6 years. Thus, if populations were not actively managed for even short periods, the abundance of horses on public rangelands would rapidly increase until animals became resource-limited (see Chapter 3). Resource-limited horse populations would affect forage and water resources for many other animals that share the rangelands with them and potentially conflict with the legislative mandate that BLM maintain a thriving natural ecological balance. They would also increase the possibility of conflict with the multiple-use policy of public rangelands (see Chapter 7). Thus, BLM should diligently monitor and manage free-ranging horse populations to meet the numerous congressional mandates in the Wild Free-Roaming Horses and Burros Act of 1971 and the Public Rangelands Improvement Act of 1978.
The larger the population of horses on public lands and the higher the growth rate of the populations, the larger the increment of new animals each year. BLM has been removing an average of about 8,000 horses from rangelands each year for the last decade in an effort to control horse populations and meet its legal obligations. Removing such a large number of horses each year has substantially exceeded the capacity of BLM to place horses into private ownership; a result is that many tens of thousands of unwanted horses are maintained in long-term holding facilities until they die. Despite the aggressive program to remove horses from public rangelands, BLM’s population-management program has not been able to reduce the free-ranging horse population to the targeted AML. For 2012, the maximum AML for horses was 23,622 (the maximum AML for burros was 2,923).
Additional management interventions in the form of various fertility-control agents have been pursued to enhance the efficacy of population management. The emerging technologies have the potential to reduce population growth rates and hence the increment of animals added to the national population each year (see Chapter 4); this might substantially increase the opportunity for the removal program to attain management goals. The potential impact of fertility control, however, is limited by the number and proportion of animals that must be effectively treated with the contraceptive agents, and it is likely to affect the genetic makeup of populations unless carefully monitored (see Chapter 5). All modeling studies exploring the potential impacts of contraceptive treatments on horse population growth rates have demonstrated that the higher the intrinsic growth rate of the population, the higher the proportion of horses that must be treated to reduce population growth rates to a prescribed level (Garrott, 1991a; Garrott and Siniff, 1992; Garrott et al., 1992; Coughenour, 1999, 2000, 2002; Gross, 2000; Bartholow, 2007; Ballou et al., 2008). Thus, the potential implementation of broad-scale fertility-control management to aid in curbing population growth rates will be confronted by the challenge of treating the large number of horses that will probably be required to have appreciable affects on horse population demography. Studies specific to burro population demography will be necessary to tailor similar management actions to that species.
Arnason, A.N., C.J. Schwarz, and J.M. Gerrard. 1991. Estimating closed population size and number of marked animals from sighting data. Journal of Wildlife Management 55:716-730.
Ballou, J.D., K. Traylor-Holzer, A.A. Turner, A.F. Malo, D. Powell, J. Maldonado, and L. Eggert. 2008. Simulation model for contraceptive management of the Assateague Island feral horse population using individual-based data. Wildlife Research 35:502-512.
Barnes, R.F.W. 2002. The problem of precision and trend detection posed by small elephant populations in West Africa. African Journal of Ecology 40:179-185.
Bartholow, J.M. 2007. Economic benefit of fertility control in wild horse populations. Journal of Wildlife Management 71:2811-2819.
Berger, J. 1986. Wild Horses of the Great Basin: Social Competition and Population Size. Chicago: University Chicago Press.
BLM (Bureau of Land Management). 2010. Wild Horses and Burros Management Handbook, H-4700-1. Washington, DC: U.S. Department of the Interior.
BLM (Bureau of Land Management). 2011. Proposed Strategy: Details of the BLM’s Proposed Strategy for Future Management of America’s Wild Horses and Burros. Washington, DC: U.S. Department of the Interior.
BLM (Bureau of Land Management). 2012. National Wild Horse and Burro Advisory Board Information for National Wild Horse and Burro Advisory Board Meeting, October 29-30, Salt Lake City, UT.
Boyd, L.E. 1980. Natality and Foal Survivorship of Feral Horses in Wyoming’s Red Desert. Ph.D. dissertation. University of Wyoming, Laramie.
Buckland, S.T. and B.J. Turnock. 1992. A robust line transect method. Biometrics 48:901-909.
Buckland, S.T., D.R. Anderson, K.P. Burnham, J.L. Laake, D.L. Borchers, and L. Thomas, eds. 2004. Advanced Distance Sampling: Estimating Abundance of Biological Populations. New York: Oxford University Press.
Buckland, S.T., D.R. Anderson, K.P. Burnham, and J.L. Laake. 2005. Distance sampling. Encyclopedia of Bio-statistics. Chichester, UK: John Wiley & Sons.
Burnham, K.P., D.R. Anderson, and J.L. Laake. 1980. Estimation of density from line transect sampling of biological populations. Wildlife Monographs 72:1-202.
Cameron, E.Z., W.L. Linklater, K.J. Stafford, and E.O. Minot. 2000. Aging and improving reproductive success in horses: Declining residual reproductive value or just older and wiser? Behavioral Ecology and Sociobiology 47:243-249.
Cao, Q., M. Songer, Y.J. Zhang, D.F. Hu, D.I. Rubenstein, and P. Leimgruber. 2012. Resource use and limitations for released Przewalski’s horses at Kalamaili Nature Reserve, Xinjiang, China. Presentation at the International Wild Equid Conference, September 18-22, Vienna, Austria.
Caughley, G. 1974a. Bias in aerial survey. Journal of Wildlife Management 38:921-933.
Caughley, G. 1974b. Interpretation of age ratios. Journal of Wildlife Management 38:557-562.
Caughley, G. 1977. Analysis of Vertebrate Populations. New York: Wiley-Interscience.
Choquenot, D. 1990. Rate of increase for populations of feral donkeys in northern Australia. Journal of Mammalogy 71:151-155.
Choquenot, D. 1991. Density-dependent growth, body condition, and demography in feral donkeys: Testing the food hypothesis. Ecology 72:805-813.
Conley, W. 1979. The potential for increase in horse and ass populations: A theoretical analysis. Paper presented at Symposium on Ecology and Behavior of Feral Equids, September 6-8, Laramie, WY.
Conroy, M.J. and J.P. Carroll. 2009. Capture–mark–recapture studies for estimating abundance and density. Pp. 135-159 in Quantitative Conservation of Vertebrates. Oxford: Wiley-Blackwell.
Coughenour, M.B. 1999. Ecosystem Modeling of the Pryor Mountain Wild Horse Range. Final Report to U.S. Geological Survey Biological Resources Division, U.S. National Park Service, and U.S. Bureau of Land Management. Fort Collins, CO: Natural Resource Ecology Laboratory.
Coughenour, M.B. 2000. Ecosystem modeling of the Pryor Mountain Wild Horse Range: Executive summary. Pp. 125-131 in Managers’ Summary—Ecological Studies of the Pryor Mountain Wild Horse Range, 1992-1997, F.J. Singer and K.A. Schoenecker, compilers. Fort Collins, CO: U.S. Geological Survey.
Coughenour, M.B. 2002. Ecosystem modeling in support of the conservation of wild equids: The example of the Pryor Mountain Wild Horse Range. Pp. 154-162 in Equids: Zebras, Asses and Horses: Status Survey and Conservation Action Plan, P.D. Moehlman, ed. Gland, Switzerland: International Union for Conservation of Nature.
Dawson, M.J. and C. Miller. 2008. Aerial mark-recapture estimates of wild horses using natural markings. Wildlife Research 35:365-370.
Dawson, M.J. and J. Hone. 2012. Demography and dynamics of three wild horse populations in the Australian Alps. Austral Ecology 37:97-109.
Dennis, B., P.L. Munholland, and J.M. Scott. 1991. Estimation of growth and extinction parameters for endangered species. Ecological Monographs 61:115-143.
Dennis, B., J.M. Ponciano, S.R. Lele, M.L. Taper, and D.F. Staples. 2006. Estimating density dependence, process noise, and observation error. Ecological Monographs 76:323-341.
Eberhardt, L.L. 1982. Calibrating an index by using removal data. Journal of Wildlife Management 46:734-740.
Eberhardt, L.L. 1987. Population projections from simple models. Journal of Applied Ecology 24:103-118.
Eberhardt, L.L., A.K. Majorowicz, and J.A. Wilcox. 1982. Apparent rates of increase for two feral horse herds. Journal of Wildlife Management 46:367-374.
Eggert, L.S., D.M. Powell, J.D. Ballou, A.F. Malo, A. Turner, J. Kumer, C. Zimmerman, R.C. Fleischer, and J.E. Maldonado. 2010. Pedigrees and the study of the wild horse population of Assateague Island National Seashore. Journal of Wildlife Management 74:963-973.
Ernest, H.B., M.C.T. Penedo, B.P. May, S.M. Syvanen, and W.M. Boyce. 2000. Molecular tracking of mountain lions in the Yosemite Valley region in California: Genetic analysis using microsatellites and faecal DNA. Molecular Ecology 9:433-441.
Freeland, W.J. and D. Choquenot. 1990. Determinants of herbivore carrying capacity: Plants, nutrients and Equus asinus in northern Australia. Ecology 71:589-597.
Feist, J.D., and D. McCullough. 1975. Reproduction in feral horses. Reproduction and Fertility (Supplement) 23:13-18.
Frei, M., J. Peterson, and I. Hall. 1979. Aerial census of wild horses in western Utah. Journal of Range Management 32:8-11.
GAO (U.S. General Accounting Office). 1990. Rangeland Management: Improvements Needed in Federal Wild Horse Program. Washington, DC: U.S. Government Accountability Office.
GAO (U.S. Government Accountability Office). 2008. Effective Long-Term Options Needed to Manage Unadoptable Wild Horses. Washington, DC: U.S. Government Accountability Office.
Garrott, R.A. 1991a. Feral horse fertility control: Potential and limitations. Wildlife Society Bulletin 19:52-58.
Garrott, R.A. 1991b. Sex ratios and differential survival of feral horses. Journal of Animal Ecology 60:929-936.
Garrott, R.A. and D.B. Siniff. 1992. Limitations of male-oriented contraception for controlling feral horse populations. Journal of Wildlife Management 56:456-464.
Garrott, R.A. and L. Taylor. 1990. Dynamics of a feral horse population in Montana. Journal of Wildlife Management 54:603-612.
Garrott, R.A., T.C. Eagle, and E.D. Plotka. 1991a. Age-specific reproduction in feral horses. Canadian Journal of Zoology 69:738-743.
Garrott, R.A., D.B. Siniff, and L.L. Eberhardt. 1991b. Growth rates of feral horse populations. Journal of Wildlife Management 55:641-648.
Garrott, R.A., D.B. Siniff, J.R. Tester, T.C. Eagle, and E.D. Plotka. 1992. A comparison of contraceptive technologies for feral horse management. Wildlife Society Bulletin 20:318-326.
Goodloe, R.B., R.J. Warren, D.A. Osborn, and C. Hall. 2000. Population characteristics of feral horses on Cumberland Island, Georgia and their management implications. Journal of Wildlife Management 64:114-121.
Greger, P.D. and E.M. Romney. 1999. High foal mortality limits growth of a desert feral horse population in Nevada. Western North American Naturalist 59:374-379.
Gross, J.E. 2000. A dynamic simulation model for evaluating effects of removal and contraception on genetic variation and demography of Pryor Mountain wild horses. Biological Conservation 96:319-330.
Guschanski, K., L. Vigilant, A. McNeilage, M. Gray, E. Kagoda, and M.M. Robbins. 2009. Counting elusive animals: Comparing field and genetic census of the entire mountain gorilla population of Bwindi Impenetrable National Park, Uganda. Biological Conservation 142:290-300.
Humbert, J., L.S. Mills, J.S. Horne, and B. Dennis. 2009. A better way to estimate population trends. Oikos 118: 1940-1946.
Keiper, R.R. 1979. Population dynamics of feral ponies. Pp. 175-183 in Proceedings of the Symposium on the Ecology and Behavior of Wild and Feral Equids, R.H. Denniston, ed. Laramie: University of Wyoming.
Keiper, R. and K. Houpt. 1984. Reproduction in feral horses: An eight year study. American Journal of Veterinary Research 45:991-995.
Kissling, M.L. and E.O. Garton. 2006. Estimating detection probability and density from point-count surveys: A combination of distance and double-observer sampling. Auk 123:735-752.
Kohn, M.H., E.C. York, D.A. Kamradt, G. Haught, R.M. Sauvajot, and R.K. Wayne. 1999. Estimating population size by genotyping faeces. Proceedings of the Royal Society of London Series B 266:657-663.
Krebs, C.J. 1999. Ecological Methodology. New York: Addison-Welsey.
Laake, J., M.J. Dawson, and J. Hone. 2008. Visibility bias in aerial survey: Mark-recapture, line transect or both? Wildlife Research 35:299-309.
Lancia, R.A., J.D. Nichols, and K.H. Pollock. 1996. Estimating the number of animals in wildlife populations. Pp. 215-253 in Research and Management Techniques for Wildlife and Habitat, T.A. Bookhout, ed. Bethesda, MD: The Wildlife Society.
Linklater, W.L. and E.Z. Cameron. 2002. Escape behaviour of feral horses during a helicopter count. Wildlife Research 29:221-224.
Linklater, W.L., E.Z. Cameron, E.O. Minot, and K.J. Stafford. 2004. Feral horse demography and population growth in the Kaimanawa Ranges, New Zealand. Wildlife Research 31:119-128.
Lubow, B.C. and J.I. Ransom. 2009. Validating aerial survey photographic mark-recapture for naturally marked feral horses. Journal of Wildlife Management 73:1420-1429.
MacKenzie, D.I., J.D. Nichols, J.A. Royle, K.H. Pollock, L.L. Bailey, and J.E. Hines. 2006. Occupancy Estimation and Modeling: Inferring Patterns and Dynamics of Species Occurrence. Burlington, MA: Academic Press.
Marsh, H. and D.F. Sinclair. 1989. Correcting for visibility bias in strip transect aerial surveys of aquatic fauna. Journal of Wildlife Management 53:1017-1024.
McClintock, B.T. and G.C. White. 2007. Bighorn sheep abundance following a suspected pneumonia epidemic in Rocky Mountain National Park. Journal of Wildlife Management 71:183-189.
McClintock, B.T. and G.C. White. 2009. A less field-intensive robust design for estimating demographic parameters with mark-resight data. Ecology 90:313-320.
McClintock, B.T., G.C. White, M.F. Antolin, and D.W. Tripp. 2009. Estimating abundance using mark-resight when sampling is with replacement or the number of marked individuals is unknown. Biometrics 65:237-246.
Millette, T.L., D. Slaymaker, E. Marcano, C. Alexander, and L. Richardson. 2011. AIMS-THERMAL—A thermal and high resolution color camera system integrated with GIS for aerial moose and deer census in northeastern Vermont. Alces 47:27-37.
Mills, L.S. 2007. Conservation of Wildlife Populations. Oxford: Blackwell.
Minta, S. and M. Mangel. 1989. A simple population estimate based on simulation for capture-recapture and capture-resight data. Ecology 70:1738-1751.
Monard, A.M., P. Duncan, H. Fritz, and C. Feh. 1997. Variations in the birth sex ratio and neonatal mortality in a natural herd of horses. Behavioral Ecology and Sociobiology 41:243-249.
NRC (National Research Council). 1980. Wild and Free-Roaming Horses and Burros: Current Knowledge and Recommended Research. Phase I Final Report. Washington, DC: National Academy Press.
NRC (National Research Council). 1982. Wild and Free-Roaming Horses and Burros. Final Report. Washington, DC: National Academy Press.
Nelson, K.J. 1978. On the Question of Male-Limited Population Growth in Feral Horses (Equus caballus). M.S. thesis. New Mexico State University, Las Cruces.
Nichols, J.D. and M.J. Conroy. 1996. Techniques for estimating abundance and species richness. Pp. 177-192 in Measuring and Monitoring Biological Diversity: Standard Methods for Mammals, D.E. Wilson, F.R. Cole, J.D. Nichols, R. Rudran, and M.S. Foster, eds. Washington, DC: Smithsonian Institution Press.
Osborn, R.G. 2004. WHIMS User’s Manual. Fort Collins, CO: U.S. Geological Survey.
OIG (Office of Inspector General). 2010. Bureau of Land Management Wild Horse and Burro Program. Report No. C-IS-BLM-0018-2010. Washington, DC: U.S. Department of the Interior.
Otis, D.L., K.P. Burnham, G.C. White, and D.R. Anderson. 1978. Statistical inference from capture data on closed animal populations. Wildlife Monographs 62:3-135.
Petersen, S.L., C.A. Carr, G.H. Collins, P.A. Clark, D.E. Johnson, C. Boyd, K. Davies, and A.J. Gooch. 2012. Free-roaming horse distribution and habitat use patterns within riparian and upland areas in western North America. Presentation at the International Wild Equid Conference, September 18-22, Vienna, Austria.
Pollock, K.H. and W.L. Kendall. 1987. Visibility bias in aerial surveys: A review of estimation procedures. Journal of Wildlife Management 51:502-510.
Pollock, K.H., J.D. Nichols, C. Brownie, and J.E. Hines. 1990. Statistical inference for capture-recapture experiments. Wildlife Monographs 107:1-97.
Pradel, R. 1996. Utilization of capture-mark-recapture for the study of recruitment and population growth rate. Biometrics 52:703-709.
Ransom, J.I. 2012a. Detection probability in aerial surveys of feral horses. Journal of Wildlife Management 76:299-307.
Ransom, J.I. 2012b. USGS Research on Wild Horses and Burros. Presentation to the National Academy of Sciences’ Committee to Review the Bureau of Land Management Wild Horse and Burro Management Program, January 27, Spokane, WA.
Ransom, J.I., P. Kaczensky, B.C. Lubow, O. Ganbaatar, and N. Altansukh. 2012. A collaborative approach for estimating terrestrial wildlife abundance. Biological Conservation 153:219-226.
Roelle, J.E., F.J. Singer, L.C. Zeigenfuss, J.I. Ransom, L. Coates-Markle, and K.A. Schoenecker. 2010. Demography of the Pryor Mountain Wild Horses, 1993-2007. U.S. Geological Survey Scientific Investigations Report 2010-5125. Reston, VA: U.S. Geological Survey.
Schwarz, C.J. and A.N. Arnason. 1996. A general methodology for the analysis of capture-recapture experiments in open populations. Biometrics 52:860-873.
Samuel, M.D., E.O. Garton, M.W. Schlegal, and R.G. Carson. 1987. Visibility bias during aerial surveys of elk in north-central Idaho. Journal of Wildlife Management 51:622-630.
Sawaya, M.A., J.B. Stetz, A.P. Clevenger, M.L. Gibeau, and S.T. Kalinowski. 2012. Estimating grizzly and black bear population abundance and trend in Banff National Park using noninvasive genetic sampling. PLoSOne 7(5):e34777.
Scorolli, A.L. and A.C. Lopez Cazorla. 2010. Demography of feral horses (Equus caballus): A long-term study in Tornquist Park, Argentina. Wildlife Research 37:207-214.
Seal, U.S. and E.D. Plotka. 1983. Age-specific pregnancy rates in feral horses. Journal of Wildlife Management 47:422-429.
Seber, G.A.F. 1982. The Estimation of Animal Abundance and Related Parameters. New York: Macmillan.
Singer, F.J., L. Zeigenfuss, L. Coates-Markle, and F. Schwieger. 2000. A demographic analysis, group dynamics, and genetic effective number in the Pryor Mountain wild horse population, 1992-1997. Pp. 73-89 in Managers’ Summary—Ecological Studies of the Pryor Mountain Wild Horse Range, 1992-1997, F.J. Singer and K.A. Schoenecker, compilers. Fort Collins, CO: U.S. Geological Survey.
Siniff, D.B., J.R. Tester, R.D. Cook, and G.L. McMahon. 1982. Census Methods for Wild Horses and Burros: Final Report. Minneapolis: University of Minnesota.
Siniff, D.B., J.R. Tester, and G.L. McMahon. 1986. Foaling rates and survival of feral horses in western Nevada. Journal of Range Management 39:296-297.
Stenglein, J.L., L.P. Waits, D.E. Ausband, P. Zager, and C.M. Mack. 2010. Efficient, noninvasive genetic sampling for monitoring reintroduced wolves. Journal of Wildlife Management 74:1050-1058.
Sugimoto, T., J. Nagata, V.V. Aramilev, and D.R. McCullough. 2012. Population size estimation of Amur tigers in Russian Far East using noninvasive genetic samples. Journal of Mammalogy 93:93-101.
Turner, A. and J.F. Kirkpatrick. 2002. Effects of immunocontraception on population, longevity and body condition in wild mares (Equus caballus). Reproduction Supplement 60:187-195.
Turner, J.W. and M.L. Morrison. 2001. Influence of predation by mountain lions on numbers and survivorship of a feral horse population. Southwestern Naturalist 46:183-190.
Tyler, S.J. 1972. The behavior and social organization of the New Forest ponies. Animal Behavior Monographs 5:85-196.
Walter, M.J. and J. Hone. 2003. A comparison of 3 aerial survey techniques to estimate wild horse abundance in the Australian Alps. Wildlife Society Bulletin 31:1138-1149.
Welsh, D.A. 1975. Population, Behavioral and Grazing Ecology of the Horse of Sable Island, Nova Scotia. Ph.D. dissertation. Dalhousie University, Halifax, Canada.
White, G.C. and K.P. Burnham. 1999. Program MARK: Survival estimation from populations of marked animals. Bird Study 46 Supplement:120-138.
Williams, B.K., J.D. Nichols, and M.J. Conroy. 2001. Analysis and Management of Animal Populations. San Diego: Academic Press.
Wolfe, M.L. 1980. Feral horse demography: A preliminary report. Journal of Range Management 33:354-360. Wolfe, M.L. 1986. Population dynamics of feral horses in the western North America. Journal of Equine Veterinary Science 6:231-235. Wolfe, M.L., L.C. Ellis, and R. MacMullen. 1989. Reproductive rates of feral horses and burros. Journal of Wildlife Management 53:916-924.
Wood, G.W., M.T. Mengak, and M. Murphy. 1987. Ecological importance of feral ungulates at Shackleford Banks, North Carolina. American Midland Naturalist 118:236-244.
Woods, J.G., D. Paetkau, D. Lewis, B.N. McLennan, M. Proctor, and C. Strobeck. 1999. Genetic tagging of free-ranging black and brown bears. Wildlife Society Bulletin 27:616-627.






























