
3
Biologic Markers For Immune-Mediated Disease
This chapter defines immune-mediated disease and the extent to which some chemicals in our environment cause hypersensitivity or immunopotentiation. The discussion focuses on three principal questions:
-
To what extent do chemicals in our environment have the potential to cause immune-mediated disease and thus pose a health threat in the United States today?
-
How can conventional and novel biologic markers be used to identify exposure and susceptibility to these agents and provide diagnostic information that is useful in the management of hypersensitivity disorders?
-
What are the critical issues, important questions, and kinds of studies that need to be considered to adequately define and manage this health problem?
This chapter is not a comprehensive treatise or catalogue of immune-mediated disorders and xenobiotics known to cause them. Rather, examples of the various disorders, chemicals that are suspected to cause them, and markers that indicate their presence are presented to review the scientific issues and suggest future directions.
DEFINITION OF THE PROBLEM
Hypersensitivity disorders are by far the most widely recognized manifestations of immunotoxicity. These disorders can result from exposure to environmental contaminants or chemicals in the workplace and have been widely reported and amply documented (Trizio et al., 1988). Hypersensitivity reactions are also the most common type of immunotoxicity associated with chemicals in the environment. Despite the relatively high morbidity and long history of these disorders (particularly in the workplace), much of the pathophysiology has only recently been established, and many questions remain to be answered. Progress in addressing this important health problem has been impaired by the slow development of appropriate animal models. Consequently, we have depended to a large extent on clinical research to establish mechanisms and effective modes of treatment.
It must be emphasized that hypersensitivity responses and susceptibility to autoimmune diseases are strongly influenced by genetics. In addition, variation from what is normal in the neurologic-hormonal balance
can significantly influence hypersensitivity responses.
Several features distinguish hypersensitivity from immunosuppression. First, hypersensitivity disorders are far more common in the general population than is immunosuppression, and hypersensitivity disorders are more readily defined and studied. Second, they usually entail a specific antibody, receptor, cell population, or target tissue. Third, because of this specificity, diagnostic and epidemiologic approaches tend to be problem oriented. Last, biologic markers of susceptibility, exposure, and effect are more often defined by suspected mechanisms, such as the presence of a specific antibody or the release of a particular mediator, rather than by the consequences of suppression of the immune system (by the presence of infection or tumors).
Consequently, the appropriateness, predictive value, and utility of laboratory tests tend to be more obvious for immune-mediated disease than for immunosuppression.
Although clinical signs of hypersensitivity generally have provided adequate markers of effect (the presence of a rash and bronchoconstriction), linkage to the offending chemical has presented a special challenge. The natural history, epidemiology, and clinicopathologic aspects pertinent to the consideration of biologic markers of these disorders are discussed in this chapter. Because exposure to environmental chemicals occurs principally through contact with the skin, the respiratory tract, and the gastrointestinal system, the discussion is organized around these routes of exposure to potentially sensitizing agents.
EXPOSURE THROUGH INHALATION (PULMONARY HYPERSENSITIVITY)
Pulmonary diseases attributed to hypersensitivity responses in the respiratory tract have been recognized for centuries. Three general categories of hypersensitivity can cause pulmonary disease. The most common category includes asthma or rhinitis that results from the production of specific IgE to inhaled allergens. The second category is hypersensitivity pneumonitis. Far fewer persons are affected than are those who have asthma or rhinitis, but because some individuals develop severe alveolitis after repeated exposure to inhaled organic dusts, this is a particularly serious liability. Hypersensitivity pneumonitis is primarily a disorder of nonatopic subjects and is associated with high levels of antibody. Both antibody and cellular immunity could be important in responses that lead to lung damage. The final category of hypersensitive lung disease includes disorders associated with cellular immunity, one example of which is chronic beryllium lung disease.
Occupational Asthma and Rhinitis
Asthma and rhinitis are common reactions to inhaled environmental plant and animal allergens. Inhalation of chemicals also can cause asthma. Asthma induced by inhaled chemicals is characterized by variable airway obstruction and hyperactivity of the airways caused by exposure to sensitizing concentrations of substances present in the environment (Chan-Yeung and Lam, 1986; Cotes and Steel, 1987), or workplace. It is estimated that 50,000 to 100,000 workers in the United States are potentially exposed to one such group of chemicals, diisocyanates (Musk et al., 1988).
Reactions to inhaled chemicals can be produced by a variety of mechanisms. The agent can act as a nonspecific irritant, it can act as an allergen that induces specific antibody production, or it can nonspecifically stimulate release of biologically active mediators. In many examples of occupational asthma, the actual mechanisms have not
been determined. Clinically, asthmatic responses to chemical haptens that induce IgE are difficult to distinguish from airway obstruction caused by irritants.
It is not clear how irritants cause asthmatic responses in the absence of an immune response. However, inflammation is an important component of asthmatic responses, and the induction of pulmonary inflammation by inhaled chemicals could be important in airway responses after inhalation of irritants. In late-phase asthmatic responses, eosinophils and neutrophils are increased in bronchoalveolar lavage fluids after antigen challenges (Metzger et al., 1985; Diaz et al., 1986). Although the exact role of inflammation in asthmatic responses is not clear, it is possible that inflammation induced by inhaled chemicals could be important in the induction of nonimmunologic asthmatic responses. The exposure of dogs to ozone results in increased airway responsiveness, suggesting that the neutrophil plays an important role in airway hyperreactivity (Holtzman et al., 1983). The production of a variety of mediators by inflammatory cells could play an important role in increasing nonspecific airway responses (Henderson et al., 1984; Cuss et al., 1986).
Many agents associated with occupational asthma are low-molecular-weight chemicals, such as nickel, platinum, palladium, toluene diisocyanate (TDI), and trimellitic anhydride (Pepys et al., 1972; McConnell et al., 1973; Zeiss et al., 1977). However, complex organic molecules, including bacterial enzymes used in detergents, also can cause this reaction (Newhouse et al., 1970). Thus with complex aeroallergens, there is an association between the presence of the disease and the atopic status of the workers.
Low-molecular-weight chemicals may act as haptens, binding to body macromolecules to form antigens that can induce haptenspecific antibodies. In some cases, antibodies also recognize a portion of the carrier molecule. Thus, immune-system responses to inhaled chemicals can be directed against a range of antigen specificities.
The initial sensitization, after exposure of the mucosa to a chemical, is thought to occur in lymph nodes that drain the lung or airways. After immunization, B cells that produce specific IgE can be found in mucosal tissues. The production of IgE in mucosal tissues and in lymph nodes that drain mucosal tissues results in the sensitization of circulating basophils and tissue-fixed mast cells throughout the body. Binding of IgE on these cells by allergens causes them to release mediators that contribute to allergic symptoms.
The incidence of occupational asthma has been estimated to be 2-4% (Salvaggio et al., 1986). Particularly potent sensitizing chemicals, including platinum salts, have been observed to sensitize more than 50% of exposed subjects (Roberts, 1951). Up to 20% of workers exposed to acid anhydrides and 5% exposed to diisocyanates have become sensitized to these agents (NIOSH, 1978; Bernstein, 1982).
Atopy is a predisposing factor for occupational asthma induced by high-molecular-weight sensitizers, such as enzymes, flour, and animal dander or venom (Newhouse et al., 1970). However, there has been no demonstrable correlation between atopy and response to low-molecular-weight sensitizers, such as TDI and trimellitic anhydrides. Other factors that could be important in the initiation of disease include altered adrenergic tone (Szentivanyi, 1968), recent or concurrent respiratory viral infection (Empey et al., 1976), and alterations in the integrity of the tight junctions of basal membranes (McFadden, 1984). Smoking does not appear to be a predisposing factor (Chan-Yeung and Lam, 1986; Cotes and Steel, 1987).
Although IgE specific for high-molecular-weight sensitizers usually can be demonstrated, they are less frequently detected for low-molecular-weight chemicals. Many individuals
diagnosed with clinical asthma associated with TDI do not demonstrate isocyanate-specific IgE in their sera (Salvaggio et al., 1986). Bronchial hyperresponsiveness to nonspecific provoking agents (histamine and methacholine) is present in most, but not all, subjects with occupational asthma. This appears to be an effect rather than a factor predisposing to sensitization (Chan-Yeung and Lam, 1986; Mapp et al., 1986a,b; Cotes and Steel, 1987). Also, some reports suggest that cell-mediated immunity may play a role in hypersensitivity reactions (Baur, 1983; Patterson et al., 1982).
Hypersensitivity Pneumonitis
Hypersensitivity pneumonitis is characterized by a spectrum of lymphocytic and granulomatous interstitial and alveolar-filling pulmonary disorders associated with intense and often prolonged exposure to any of a wide range of inhaled organic dusts and related occupational antigens. The list of organic materials that can induce hypersensitivity pneumonitis continues to grow, with disease classification based on the offending inhaled material.
The limited data available show that hypersensitivity pneumonitis can be induced by inhalation of chemicals. Diphenylmethane diisocyanate, TDI, trimellitic anhydride, and heated epoxy resin are examples of chemicals that can induce hypersensitivity pneumonitis (Blake et al., 1965; Charles et al., 1976; Salvaggio, 1987; Musk et al., 1988), but the frequency of this disorder is considerably less than the asthmatic type response to these chemicals.
Inhaled organic particulates must be deposited in the terminal airways and alveoli to induce an immune response leading to hypersensitivity pneumonitis (Salvaggio, 1987). Some of these antigens have adjuvant properties that could be important in the induction of these reactions (Bice et al., 1977). The induction of immunity to these antigens is not understood, but it seems likely that antigens deposited in the lower respiratory tract are phagocytized by neutrophils or alveolar macrophages and carried via the lymphatics to the lung-associated lymph nodes (Harmsen et al., 1985, 1987). Immune cells produced in these tissues are released into the circulatory system and recruited from the blood into the lung (Bice and Shopp, 1988). Inflammatory reactions to antigens deposited in the lung could be responsible for the recruitment of immune cells (Bice and Shopp, 1988).
Although antibody produced to inhaled antigens can be identified in the blood of most exposed persons, the levels do not correlate well with disease (Pepys, 1986). In most cases of hypersensitivity pneumonitis, pulmonary lesions are substantially different from immune-complex-mediated vasculitic lesions (Salvaggio, 1987). Nevertheless, some data suggest that antibody and immune complement in the lung could be important in the production of lung lesions (Warren et al., 1977; Pepys, 1986).
In provocation exposures, pulmonary reactions in individuals with hypersensitivity pneumonitis appear several hours after exposure. Moreover, many materials that induce hypersensitivity pneumonitis also activate immune complement by the alternative pathway. This mechanism could contribute to hypersensitivity pneumonitis (Warren et al., 1977). Other reports suggest that cellular immunity plays a role in the pathogenesis of pneumonitis (Salvaggio, 1987). The histologic features of lungs from such patients suggest cellular immune reactions. Sensitivity to a pulmonary challenge with some agents can be passively transferred using lymphocytes from immune animals (Bice et al., 1976; Schuyler et al., 1987, 1988).
An increased number of cells and lymphocytes appears in lungs of individuals with hypersensitivity pneumonitis when assayed by bronchoalveolar lavage. However, as observed with specific antibody and cellular
immunity, an increase in total cells and lymphocytes in the lung is not always directly associated with disease. In longitudinal studies, clinically normal individuals can have significantly elevated numbers of lymphocytes for extended periods (Cormier et al., 1984, 1986). Disease and the presence of antibody, the level of cellular immunity, and numbers of lymphocytes in the lung are poorly correlated. It should be considered that disregulation of immunity may be important in the pathogenesis of hypersensitivity pneumonitis.
EXPOSURE THROUGH INGESTION
Adverse reactions to the ingestion of foods have been documented (Metcalfe, 1989). When the pathogenesis of these reactions involves an immunologic response to a component of food, the reaction is called food hypersensitivity (Anderson, 1986). A nonimmunologic response to ingested food is called food intolerance, which can be pharmacologic, toxic, or metabolic. Food intolerance includes illnesses caused by foods that contain noxious substances, such as histamine, tyramine, and possibly other biogenic amines or lectins. Some foods also elicit the nonspecific release of chemical mediators (histamine, prostaglandins, leukotrienes, and lymphokines) from mast cells and perhaps from macro-phages and lymphocytes (Moneret-Vautrin, 1983). These nonspecific responses or a lack of tolerance to food additives or other chemicals can lead to responses that mimic allergic reactions (Schulz, 1983).
Food hypersensitivity includes several clinicopathologic entities: anaphylaxis and other immediate reactions, eosinophilic gastroenteritis, eczema, and protein-induced gastroenteropathy in infants and children (Metcalfe, 1989). Although antibody and cellular immunity have been associated with immunologic reactions to ingested materials, most reactions are IgE dependent; cellular immune reactions appear to account for few of these disorders (Moneret-Vautrin and André, 1983). As for all responses that appear to have an allergic cause, it is important to remember that a variety of mechanisms can lead to symptoms that resemble allergy symptoms. The prevalence of food hypersensitivity in the general population is unknown (Metcalfe, 1989). An important problem in estimating the number of persons with food hypersensitivity is the difficulty in distinguishing true allergic reactions from nonimmunologic adverse responses. One study of 480 children followed from birth to the age of three revealed that approximately 8% had adverse reactions to particular foods. One to two percent of all children have an immunologic basis for their reactions (Bock, 1987).
Allergic reactions to ingested materials appear to be more common in individuals with other allergic disorders. As many as 10% of children and 3% of adults with other allergies exhibit allergic responses to foods (Moneret-Vautrin, 1986). Early publications suggested that colorants, including azoic dyes (tartrazine, erythrosin, and amaranth) and derivatives of triphenylmethane, might produce hypersensitivity responses. In the past, sensitivity to tartrazine was thought to affect 50,000 to 100,000 persons in the United States (Lockey, 1959). More recent results, however, provide convincing evidence that tartrazine is not responsible for the chronic urticaria observed in some patients exposed to the dye (Stevenson et al., 1986).
Preservatives (antiseptics and antioxidants) also can be sensitizers. Citric acid added as an antiseptic preservative has been blamed for triggering buccal aphthous ulcers (Pradalier et al., 1984). Implicated antioxidants include tocopherols, lecithin, propyloctyl, dodecylgallates, butylhydroxytoluene, and butylhydroxyanisol. Those most often responsible for adverse reactions are sodium nitrite, sodium benzoate, sodium metabisulfite, butylhydroxytoluene, and butylhydroxyanisol (Sogn, 1984; Moneret-Vautrin, 1986).
Medications that contain these substances are another source of exposure (Koepke et al., 1984). The mechanisms responsible for many of these adverse reactions are not known.
Ingested sulfiting agents can cause urticaria, angioedema, intestinal dysfunction, vasomotor headaches, or airway constriction in sensitive individuals (Prenner and Stevens, 1976; Clayton and Busse, 1980; Stevenson and Simon, 1981; Schwartz, 1983; Bush et al., 1986; Moneret-Vautrin, 1986). Acute rhinitis, flushing, and even anaphylactoid reactions also have been reported. In some cases, increased asthmatic symptoms can occur after inhalation of aerosols from sulfite-containing foods (Werth, 1982). Although the extent of sulfite sensitivity in the general population is unknown, it appears that asthmatics are at the highest risk (Sogn, 1984). Respiratory effects of sulfite ingestion could be the greatest health threat, although some patients have reported severe abdominal distress as the principal adverse effect (Schwartz, 1983). Benzoic acid and its salts (Michaelsson and Juhlin, 1973), sodium glutamate (Allen and Baker, 1981; Swan, 1982), and nickel (Brun, 1979) are materials found in food that cause adverse reactions. The mechanisms of adverse reactions to ingested substances are not fully understood.
DERMAL EXPOSURE
Dermatitis can be caused by immune responses to antigens or haptens that come into contact with the skin or nonspecifically by contact with a substance that chemically damages the skin (irritant dermatitis). Most chemicals that cause dermatitis (70-80%) act as primary irritants, although some substances are both irritants and allergens. Only immunologically mediated dermatitis will be considered in this discussion. It is estimated that 20-25% of all cases of occupational dermatitis result from immune sensitization (Birmingham, 1988).
There are three types of immunologically mediated dermatitis. The first, allergic contact dermatitis, is the result of prior exposure of the skin to the material eliciting the dermatitis or to a chemical closely related to the substance (e.g., nickel sensitivity). The second variety, photocontact dermatitis, is similar to the first, but requires exposure to sunlight and allergen to produce a skin reaction. Allergic photocontact dermatitis is caused by antibacterial chemicals, chlorpromazine, and fragrances. In rare cases, shortwave ultraviolet light is required for the photoallergic response. Cell-mediated immunity is responsible for allergic contact dermatitis and photocontact dermatitis. The third form of allergic dermatitis is contact urticaria, a transient wheal-and-flare response caused by IgE antibody reactions. The patient's degree of sensitivity and the amount of antigen determine whether only localized urticaria occurs or if a generalized manifestation, such as angioedema, asthma, or anaphylaxis, will develop.
Contact Dermatitis
Contact dermatitis is widespread, and the substances responsible are usually low-molecular-weight chemicals that act as haptens to induce cell-mediated immunity (Fregert, 1978; Goh, 1988). Although skin changes can be seen as early as 3 hours after exposure, maximum responses usually occur 12 hours or later following exposure. Nickel sensitivity is commonly observed (5.8-8%), and women are more frequently sensitized than men, likely because of their exposure to jewelry and clothing items (Peltonen, 1979; Prystowsky et al., 1979). Chromium and cobalt also are common sensitizers. Other substances that can cause contact dermatitis include chemicals in detergents, shampoos, soaps, and perfumes, and rubber and rubber-based adhesives. Individuals exposed to epoxy glues and coatings, some amines, chromates, formaldehyde, and some antioxidants
also are at risk of becoming sensitized (Birmingham, 1988). Although these sensitivities are not usually life threatening, the large numbers of chemicals that can cause sensitivity indicates the degree of the problems.
The ability of a chemical to induce sensitization is related to its ability to traverse the skin and react covalently with host biomolecules (Eisen et al., 1952; Goh, 1988). The chemicals are carried to regional lymph nodes via lymphatic Ia-positive Langerhans cells (Stingl et al., 1978; Silberberg-Sinakin and Thorbecke, 1980; Anderson et al., 1987; Botham et al., 1987; goh, 1988). Immune cells produced in a primary immune response are distributed to all body tissues. Further exposure to the sensitizing chemical and localized immune recognition induces a mononuclear cell infiltrate and the production of a contact sensitivity reaction. The reaction peaks at 12-24 hours (Goh, 1988). Neutrophils generally are not present with contact dermatitis, while they are in inflammatory responses, infection, or irritation.
It is not known why some individuals become sensitized and others do not, but genetic differences are thought to be important. In addition, dose plays a major role in sensitization and in elicitation of response (Karol and Thorne, 1988). Increasing the concentration of sensitizer applied to the skin, increasing the skin area exposed, applying the sensitizer to inflamed skin, and repeated exposures all increase the chance that sensitization will occur.
Photocontact Dermatitis
Photocontact dermatitis is also believed to be mediated by cell-mediated immune responses. The clinical characteristics of photocontact dermatitis closely resemble those of allergic contact dermatitis. The major difference is that photocontact dermatitis requires sunlight in addition to skin exposure to a sensitizing chemical, for initial sensitization and for subsequent elicitation of skin responses. Phototoxic reactions that can be produced in the absence of immunization are not considered here.
Sensitivity to photoallergens appears to occur by the same mechanisms as that for allergic contact sensitivity. However, ultraviolet or visible radiation participates in the induction of immunity and in contact dermatitis by converting an immunologically inactive form of the photosensitizing compound into its active form (Elmets, 1986). The exact mechanisms are not known and could vary depending on the photoallergen involved (Morison and Kochevar, 1983).
Many drugs and chemicals cause photocontact dermatitis (Elmets, 1986). They frequently share structural similarities to chemicals that cause allergic contact dermatitis (Harber and Baer, 1972). Photoallergens are usually lipid-soluble, low-moleccular-weight substances that act as haptens and include sulfanilamide, halogenated salicylanilides, phenothiazines, benzophenones, and diphenhydramine. The most common photoallergens are topical agents, although some systemically administered drugs that absorb solar energy when they reach the skin can produce photoallergic responses.
Contact Urticaria
Contact urticaria is mediated by IgE to antibody allergens. As with other IgE reactions, contact urticaria may appear from a few minutes to a few hours after the eliciting agent comes into contact with the skin. The response usually disappears within 24 hours. In cases of strong hypersensitivity or large doses of allergen, symptoms can occur in other organs, resulting in rhinitis, conjunctivitis, asthmatic attack, or even anaphylactic shock. The term contact urticaria syndrome has been used for these systemic responses (Maibach and Johnson, 1975).
The substances that cause immunologic
contact urticaria include foods, chemicals in medications, industrial chemicals, cosmetics, and many chemically undefined agents (Lahti and Maibach, 1987). Some of the allergens that cause urticaria have high molecular weight (von Krogh and Maibach, 1985).
Exposure to allergens that cause contact urticaria can be through the skin or by other routes, including the respiratory and gastrointestinal tracts. The production of IgE may occur in draining lymph nodes. The specific IgE antibody binds to mast cells in the skin and elsewhere. Upon exposure, the allergens penetrate the epidermis and cause release of mediators from IgE-bound mast cells or basophils. The cutaneous symptoms, erythema and edema, are elicited by these vasoactive substances. In addition to histamine, other mediators of inflammation are believed to be prostaglandins, leukotrienes, and kinins.
NONSPECIFIC IMMUNE ENHANCEMENT
An immunologic adjuvant is a substance that enhances the immunogenicity of another substance. Examples include mineral suspensions, such as alum, on which the antigen is adsorbed, and a water and mineral-oil mixture in which the antigen is emulsified. Adding killed mycobacteria to the emulsion further increases antigenicity (Freund's complete adjuvant).
The demonstration that xenobiotics in the environment can similarly enhance antigenicity has led to the concept of environmental adjuvants, illustrated by a study in which guinea pigs were exposed to ovalbumin with and without sulfur dioxide (Riedel et al., 1988). Compared with the control group, the group exposed to sulfur dioxide developed more bronchial reactivity on subsequent challenge with ovalbumin, and more ovalbumin-specific antibodies were found in serum and bronchoalveolar lavage fluid at a sulfur dioxide exposure of 0.1 ppm for 8 hours a day for 5 consecutive days. Hence, the presence of sulfur dioxide potentiated the development of allergy to the protein ovalbumin in this study, and sulfur dioxide can be considered an environmental adjuvant.
Similar effects have been noted in monkeys exposed to a platinum salt in the presence of ozone. Findings included increased bronchial reactivity and a higher incidence of skin reactivity to subsequent platinum salt exposure in animals, which received ozone with the platinum exposure (at a concentration of 1 ppm) relative to animals exposed only to platinum (Biagini et al., 1986).
Brief exposures to high concentrations of ozone (> 5 ppm), nitrogen dioxide (70 ppm), and sulfur dioxide (330 ppm) have been shown to enhance sensitization of ovalbumin in guinea pigs (Matsumura, 1970a,b,c). Low concentrations of ozone (0.8 or 0.5 ppm), comparable to environmental exposures administered over 3- to 4-day periods, enhanced sensitization of mice to inhaled ovalbumin (Osebold et al., 1980).
Several studies have documented an increase in the incidence of deaths from asthma in this country and abroad. For example, the prevalence of asthma among Michigan Medicaid patients increased from 2.0/100 in 1981 to 2.8/100 in 1986 (Gertsman et al., 1989). Asthma deaths increased in the United States, New Zealand, Great Britain, and other countries from the 1970s to the mid-1980s (Jackson et al., 1988). Analysis of possible artifacts, such as changes in certification and coding, supports a real increase in asthma mortality. Attempts to understand the increased death rate have centered on underestimation of the severity of asthma and inadequate use of corticosteroids (Sears, 1988). An understanding of the increased incidence has centered on air pollution in general, and sulfur dioxide in particular. Exacerbation of asthma during pollution episodes have been reported. For example, in Donora, Pennsylvania, in 1948, 88% of asthmatics experienced exacerbation of their asthma during a pollution episode (Schrenk et al., 1949). Other epidemics have
been reviewed (Sheppard, 1988), and it is pointed out that the present Occupational Safety and Health Administration (OSHA) standard of 5 ppm of sulfur dioxide exposure over an 8-hour shift is one order of magnitude higher than the concentrations (0.4 ppm) known to potentiate acute bronchospasm in asthmatics during moderate to heavy exercise (Roger et al., 1985).
In Japan, allergic rhinitis to pollen changed from 1950 to 1980 from a rare disease to the most common allergic disease. A survey of 153 Japanese school children up to age 7 (Muranaka et al., 1986, and references therein) found a high prevalence of allergic rhinitis to pollen in districts polluted by automobile exhausts. A Japanese group was able to demonstrate that diesel-exhaust particulate matter was an effective adjuvant for inducing specific IgE production to Japanese cedar pollen, the most prevalent cause of allergic rhinitis in Japan (Muranaka et al., 1986). Thus, age would not seem to be an important factor in these increasing hypersensitivity reactions. The mechanisms by which inhaled pollutants apparently enhanced the immune recognition of allergens are not understood.
BIOLOGIC MARKERS OF HYPERSENSITIVITY
History and Clinical Signs
As with the diagnoses of many diseases, a good history and physical examination are often the most useful indicators for hypersensitivity disorders and are therefore useful in supporting the establishment of biologic markers. A thorough history should provide information about the extent and duration of the exposure and, perhaps, about the immunogen. Answers to questions about the home and workplace often reveal important clues. It is sometimes difficult to distinguish clinical signs of hypersensitivity disease from those of more common illnesses. For example, patients with hypersensitivity pneumonitis usually have a history of repeated bouts of dyspnea, fever, cough, malaise, and weight loss. Frequently, a presumed diagnosis of ''atypical" or "viral" pneumonia is made, and patients are treated with antibiotics, rather than removed from the environment with the offending chemical. One problem that complicates the diagnosis of chronic beryllium disease, for example, is the similarity of its clinical presentation to that of sarcoidosis, a systemic granulomatous disease of unknown etiology (Sprince et al., 1978).
Skin Tests
Results of direct skin tests (by intradermal infection, prick, or scratch) probably provide the most sensitive biologic markers of IgE-mediated hypersensitivity. They are most useful when the chemical induces antibody-or cell-mediated responses that result in skin reactions. Some antigens are also irritants or are biologically active and may produce nonspecific skin reactions that mask hypersensitivity responses.
Patch tests are frequently used to diagnose skin hypersensitivity. If a skin response is antibody mediated, vesiculation and a dermatitic reaction are observed shortly after application of the patch test. Urticaria with classic wheal-and-flare appearance tends to be relatively short lived, in contrast to cellular immune reactions, for which a positive response is frequently observed at 24-48 hours. The histologic changes of cellular immune reactions include spongiosis, vesiculation, upper dermal edema, and perivascular mononuclear infiltration. These effects are produced principally by combined actions of lymphokines released from activated lymphocytes in the skin.
Skin tests alone cannot be used to diagnose hypersensitivity (Metcalfe, 1989). Some individuals have positive skin tests but have no clinical manifestations of allergy.
Serum IgE Concentration
The antibodies responsible for immediate immune responses belong to the IgE subclass of immunoglobulin (Ishizaka et al., 1966; Johansson and Bennich, 1967), and total serum IgE is increased in young patients with asthma (Johansson and Bennich, 1967; Rowe and Wood, 1970). Based on these findings, investigators have attempted to distinguish extrinsic (allergic) from intrinsic (nonallergic) forms of asthma according to whether an elevated serum concentration of IgE can be demonstrated (Ostergaard, 1985). A large, general-population study has shown that, when test subjects are standardized for age and sex, the prevalence of asthma is closely related to serum IgE level (Burrows et al., 1989). The same study showed that allergic rhinitis, in contrast to asthma, could be indicated by skin test reactions to common aeroallergens, independently of the serum IgE level.
In Vitro Assays for Specific Antibody
In vitro assays can be used to identify antibodies (IgE or IgG) against specific antigens. Conjugates of low-molecular-weight chemical haptens with protein carriers are used most frequently to detect these antibodies. Albumin is often used as a carrier in assays for immune recognition to test chemicals conjugated with a hapten; such a conjugate could ellicit a negative response in a sensitized individual if it is not the same carrier-hapten conjugate to which the individual is sensitized. In vitro assays can be especially useful if the agent in question is toxic or an irritant.
Measurement of specific IgE antibody to low-molecular-weight chemicals is difficult. It must be remembered that the absence of specific IgE in the circulation does not mean that an individual has not been exposed to a chemical or is not sensitive to the agent. Although some investigators have implicated IgE in reactions to sulfites by passive-transfer testing, others have been unable to identify antisulfite IgE (Prenner and Stevens, 1976; Stevenson and Simon, 1981). It is possible that the clinical symptoms observed could result from an irritant effect of the sulfites rather than from production of IgE-specific antibody (Stevenson and Simon, 1981). Similar difficulties have been encountered with the measurement of isocyanate-specific IgE. Karol and Thorne (1988) discussed host-related problems associated with the measurement of isocyanate IgE.
The identification of specific antibody serves as a marker of exposure. Antibody produced to inhaled antigens can be identified in the blood of most individuals exposed to large quantities of antigens, but levels of specific antibody frequently do not correlate with disease (Pepys, 1986).
In Vitro Assays for Cellular Immunity
Lymphocyte stimulation and other in vitro assays for cellular immunity can be used as indicators of cellular immune-mediated reactions. Some antigens or chemicals, however, are either biologically active or are cytotoxic, making interpretation of lymphocyte stimulation assays difficult. Lymphocyte stimulation has been shown to be useful in the diagnosis of drug-induced occupational allergy (Stejskal et al., 1986). It is currently used to indicate sensitivity to beryllium and may be useful in the diagnosis of chronic beryllium disease. Antibodies to beryllium have also been detected in exposed workers (Clarke, 1991).
Provocation Challenges
Although specific inhalation challenges are not without risk and must be standardized, they provide a means to demonstrate sensitization in patients with occupational lung disease (Salvaggio et al., 1986; Salvaggio,
1987). The clinical responses vary and can be immediate or delayed in onset. Late asthmatic reactions are more frequent after exposure to low-molecular-weight sensitizers (Pepys and Hutchcroft, 1975; Mapp et al., 1986a). Late or delayed asthmatic reactions to provocation challenge are associated with a temporary increase of nonspecific bronchial hyperresponsiveness (Mapp et al., 1985, 1986a).
In addition to inhalation challenges in the laboratory, test exposures can be achieved by having patients return to the environment in question for observation of their response. Ingestion of a suspected toxic chemical with subsequent measurement of the clinical response also has been used as a diagnostic test (Bush et al., 1986; Sampson, 1986). In evaluating hypersensitivity, it is also best to use double-blind, placebo-controlled challenges (Metcalfe, 1989).
The use of environmental control units is a modification of a provocation challenge. An environmental control unit houses patients suspected of having sensitivities to the volatile organic compounds that are ubiquitous in our environment. The unit is constructed of building materials and is furnished to minimize outgassing of organic chemicals. There are no sources of combustion, such as gas or fuel oil furnaces, heaters, or appliances, inside the structure. The use of perfumes and colognes is restricted. The ventilation system is designed to reduce pollution levels in both intake and recycled air. Patients are housed in the unit and monitored for improvement. Challenges with chemicals or mixtures of chemicals are then performed to study the role these play in the patients' illnesses. Chemicals in foods and water are potential sources of sensitivity and should be considered simultaneously.
Bronchoalveolar Lavage
Bronchoalveolar lavage is often performed to provide lung cytology and biopsy samples for histopathology. The presence of increased numbers of neutrophils or eosinophils in bronchoalveolar lavage fluid is suggestive of occupational asthma (Fabbri et al., 1987; Lam et al., 1987). Bronchoalveolar lavage fluid from individuals with hypersensitivity pneumonitis generally has a large number of total cells and lymphocytes. The ratio of helper (CD4) to suppressor (CD8) T cells present in fluid from patients with hypersensitivity pneumonitis is <1, as opposed to 5:1 or more in that from patients with active sarcoidosis (Leatherman et al., 1984; Costabel et al., 1985). However, it is best to be cautious when basing diagnoses on cytologic observations. Positive findings represent exposure not disease. Individuals exposed to antigens known to cause hypersensitivity pneumonitis can have increased total cells and lymphocytes in their lungs even when there is no clinical disease.
ANIMAL MODELS FOR DETECTING CHEMICALLY MEDIATED HYPERSENSITIVITY
The development of animal models to detect and elucidate the mechanisms of chemically mediated hypersensitivity will improve our understanding of these important disorders. For animal models to be useful, the mechanisms of induction of sensitivity to chemicals and the response to exposure that leads to clinical and pathological changes should be similar to those observed in humans. Immune responses produced in target tissues (skin and lung) should have responses that are comparable to those in humans.
There are significant differences in the immunologic and inflammatory responses in various laboratory animals that have made it difficult to interpret data obtained from the variety of animals models used previously. Extrapolation of data from an animal model to humans is possible when the animal model has been carefully characterized regarding normal immune function and immunopathology.
The guinea pig has frequently been used to evaluate asthmatic responses to chemicals (Karol et al., 1981; Karol, 1983). Techniques using this species (including the monitoring of core temperature by telemetry) have been refined and validated as sensitive indicators of pulmonary hypersensitivity, and they show promise as predictors of these disorders in humans (Karol, 1988; Thorne et al., 1987b). Genetically hyperresponsive rats and dogs, sheep made hyperresponsive to ozone, guinea pigs sensitized to ovalbumin (Griffiths-Johnson and Karol, 1991) and mouse and rabbit models of late allergic reactions also have been used to study bronchial hyperresponsiveness (Tse et al., 1979; Woolcock, 1988). However, animal models usually require high doses of chemicals to induce asthmatic reactions. In addition, it is difficult to measure specific IgE in some models (Woolcock, 1988).
Animals have been used to study hypersensitivity pneumonitis induced by complex antigens or soluble proteins inhaled or instilled into the lung. Results of these studies have provided some understanding of pulmonary immunity. However, whether these reactions truly represent hypersensitivity is still in question. All test animals develop immunity after exposure of the lungs to some antigens. Rarely, however, do animals develop clinical disease comparable to that observed in the small percentage of humans with this disorder.
Rats, guinea pigs, and dogs have been used to evaluate the induction of immunity to instilled or inhaled beryllium, as well as the lung lesions induced by this agent (Schepers et al., 1957; Barna et al., 1981, 1984a,b). Dogs develop lung lesions similar to those seen in affected humans (Haley et al., 1989). Some strains of guinea pigs are sensitive and some insensitive to the induction of granulomas after instillation of beryllium. Some species require the instillation of relatively large doses of beryllium to induce immunity or lung lesions (Barna et al., 1981, 1984a,b), and in some animals, the lesions produced are histopathologically dissimilar to human lesions (Schepers et al., 1957).
Cellular responses in the lungs of various animals differ significantly after lung immunization (Bice and Shopp, 1988). Immune responses in the lungs of dogs appear to be similar to those of nonhuman primates, including chimpanzees. Dogs and nonhuman primates could prove useful as models to evaluate the effects of lung immunity on the development of pulmonary disease. Numerous species, including monkeys, mice, rabbits, dogs, and rats, have been used to investigate contact sensitivity. The guinea pig produces a reaction histologically and morphologically similar to that observed in humans (Mekori and Claman, 1986). Mice offer an advantage as a model, because there is considerable knowledge about the genetics and control of immunologic responses in this species. There are some differences in the duration of the sensitized state between mice and guinea pigs, with the response being relatively short-lived in mice (Karol, 1988). It has generally been difficult to sensitize experimental animals to orally administered chemicals reliably.
Techniques Used in Animal Studies to Determine the Potential of Chemicals to Cause Hypersensitivity
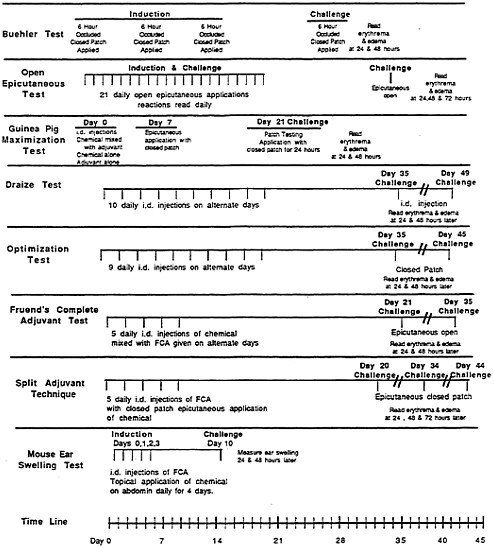
The major routes of exposure that elicit an immune reaction to a chemical are skin contact, pulmonary exposure (inhalation), and exposure by the oral route (gastrointestinal tract). The design and institution of an animal study depend on the anticipated route of exposure to the xenobiotic substance. Table 3-1 provides a summary of the tests that can be derived from toxicologic studies and special tests available for detection of chemicals that induce hypersensitivity. Animal studies used to predict the hypersensitivity responses originally described by Gell et al. (1975) are outlined below.
TABLE 3-1 Methods of Detecting Chemicals Producing Contact Hypersensitivity
|
Class |
Name |
Comments |
|
Epicutaneous methods |
Buehler Testa |
Use of occlusive technique for both inductions and elicitation. |
|
|
Open Epicutaneous Testb |
Screens many chemicals and finished formulations. Open epicutaneous for induction and closed patch for elicitation. |
|
|
Split Adjuvant Techniquec |
Increases sensitivity during induction phase. Epicutaneous induction with adjuvant injected i.d.; occlusive patch used for elicitation. |
|
Intradermal methods |
Draize Testd |
Intradermal induction and elicitation. |
|
|
Optimization Teste |
Intradermal induction and epicutaneous elicitation. |
|
|
Freund's Complete Adjuvant Testf |
Intradermal induction with and without Freund's Complete Adjuvant and epicutaneous elicitation. |
|
|
Epicutaneous induction with adjuvant on day 0 elicitation on ear and measure ear thickness. |
|
|
Combinations |
Guinea Pig Maximization Testi |
Intradermal induction with and without Freund's Complete Adjuvant and epicutaneous elicitation. |
|
|
Split-Adjuvant Techniquej |
Epicutaneous induction with adjuvant injected i.d.; occlusive patch used for elicitation. |
|
a Buehler (1964) b Klecak (1985) c Maguire (1972) d Draize (1955) e Maurer (1985) f Klecak (1985) g Gad et al. (1986) h Stern et al. (1989) i Magnusson and Kligman (1969) j Maguire (1972) |
||
IgE-and IgG-Mediated Immediate Reactions
An immediate hypersensitivity response usually can be identified in routine toxicologic studies. Continuous daily exposure of guinea pigs, dogs, or primates provides an opportunity for a chemical to induce an immune response, allowing for adverse effects to be identified. Urticaria, asthmatype reactions, and cardiovascular collapse are indicators of immediate hypersensitivity. Because the guinea pig is considered a sensitive animal for hypersensitivity responses, special studies often use this model, with erythema, edema, urticaria, pulmonary distress, and other clinical signs of anaphylactic shock as indicators of a hypersensitivity reaction (Karol, 1988). In some species, adjuvants are used to increase the probability
of producing an immediate hypersensitivity response, but they are unnecessary with guinea pigs.
Cytotoxic Reactions
As with the anaphylactic reactions, production of major antibody to an antigenic determinant on a cell often will be manifested during subchronic or chronic toxicologic studies. Reductions in erythrocytes or platelets, for example, can indicate such a response, and special studies can be performed to investigate this possibility. Such studies are often carried out to determine whether antibodies bind to target cells and can in turn agglutinate or lyse them in the presence of the complement. Ligand-binding and histochemical assays that have been used to measure drug-induced antibodies bound to cell membranes include the use of fluorescent antibody and radiolabeled staphylococcal protein A; monoclonal-antibody techniques also have been used. These tests, as well as more traditional assays, such as the Coombs test for detecting red-cell agglutinins, are increasingly used in toxicologic research.
Antigen-Antibody Complexes
Examples of hypersensitivity disorders in which antigen-antibody complexes are deposited on cell membranes include immune-complex glomerulonephritis, pneumonitis, and vasculitis. Such disorders would usually be detected over the course of subchronic or chronic toxicity studies. The conditions can be detected clinically during the live phase of these studies, but they are more often diagnosed by histologic and cytologic examination, employing some of the techniques listed above, to detect immune complexes deposited in tissues.
Contact Hypersensitivity
Contact-hypersensitivity response is mediated by T cells and is classically demonstrated 24-48 hours after challenge. Although this type of response has been best described and investigated in the skin, it is believed to occur in other organs, such as the lung. Contact-hypersensitivity responses to chemicals have characteristics similar to the Jones-Mote response, in that the reaction decreases after 48 hours and is transferable with T cells. Like the tuberculin response, contact hypersensitivity lasts up to 72 hours.
Many skin-sensitization procedures are available for examining xenobiotic-induced contact hypersensitivity. Guinea pigs have been used in risk assessment for more than 20 years. Table 3-1 shows approaches available to demonstrate contact hypersensitivity. Guinea pig skin is used most widely for these studies, but the mouse-ear-swelling test (MEST) and the lymphocyte proliferation assay have shown promise as methods for detecting and investigating the mechanism of hypersensitivity reactions (Gad et al., 1986). The ability of the MEST to demonstrate dose-response sensitization is an important concept for biologic monitoring (Thorne et al., 1987a).
The methods used to test for the skin contact-hypersensitivity response can be organized into three classes, and details of the guinea pig methods are described by Klecak et al. (1977), who also describe several variables that are important in using the guinea pig to detect contact sensitizers. Figure 3-1 illustrates the major features necessary for a chemical to produce an initial sensitization and the elicitation of hypersensitivity response in the skin. Two phases, an induction phase and an elicitation (challenge) phase, are illustrated. Figure 3-2 shows and compares selected methods for detecting chemical contact sensitizers.
The aims of these testing procedures are
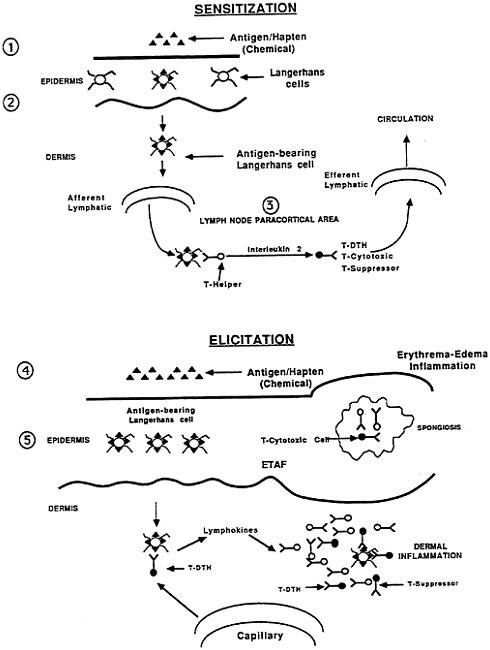
twofold: first, to optimize the penetration of the chemicals through the skin to immunocompetent cells; and second, to amplify any immunologic response that occurs but is not manifested to a level that overcomes "background noise." The Buehler test (Buehler, 1964) maximizes penetration by a topical closed-patched technique that resembles the human patch test. In the optimization test (Maurer, 1985), the chemical is injected intradermally. The guinea pig maximization test described by Magnusson and Kligman (1969) uses both intradermal and epicutaneous exposure. Because there is concern about using intradermal injections for predicting contact hypersensitivity, Jordan (1982) reported the use of dermabrasion as a means of enhancing penetration. Amplification of the response is also accomplished with adjuvants. The Freund's complete adjuvant test and guinea pig maximization test are two examples of assays that employ adjuvants (Klecak, 1985).
Guinea pig hypersensitivity assays, although sensitive and reproducible, are limiting, in that they often require subjective evaluation. Irritating chemicals or colored compounds are difficult to score. For the most part, to differentiate immune from irritating reaction, nonirritating concentrations are used to challenge the animal. This raises concern that not enough chemical has been made available to elicit the immune memory response. Erythema and edema are the end points for the immune response in all the tests described.
Efforts are under way in many laboratories to develop and validate more quantitative "immunologically based" assays for species other than the guinea pig. In vivo murine assays being examined include the ear swelling, cellular influx, and local lymph node proliferation and infiltration tests. Gad et al. (1986) showed the potential usefulness of the mouse as an experimental animal. Recently, Gad reported on the comparison of four methods (the Buehler test, the guinea pig maximization test, the epicutaneous maximization test, and the MEST) with human data to show that all four tests can generate data that are useful for a relative-hazard classification process (Gad, 1988). Kammüller et al. (1989) reported the usefulness of a popliteal lymph node (proliferation) assay in mice.
Stern et al. (1989), by radiolabeling the immune cells of mice before challenge, measured immune-cell localization in the challenge site. Kimber and Weisenberger (1989) have used a node proliferation assay in the mouse to differentiate between immune-system-mediated effects and irritation. Some of the in vitro tests under study include monitoring of lymphocyte blastogenesis, cytokines, and acute-phase proteins. In addition, several assays use cytologic or histologic markers, such as ICAM-1 (intercellular adhesion molecule-1) in mice and cutaneous basophil hypersensitivity in guinea pigs. The in vitro assays, coupled with cell identification with histologic markers, are clarifying the mechanisms of chemical hypersensitivity.
SUMMARY
Hypersensitivity disease resulting from inhalation of, dermal exposure to, or ingestion of chemicals contaminating the workplace and the environment has become an important health problem in industrialized societies. The ability of a chemical, particularly a low-molecular-weight chemical, to induce sensitization is generally related to its ability to couple covalently to body proteins. Sensitization, however, does not necessarily lead to disease.
Inhalation of a variety of chemicals can sensitize individuals and result in asthma, rhinitis, pneumonitis, or chronic granulomatous pulmonary disorder. The increasing prevalence of and mortality from asthma is a serious health problem that needs urgent attention. Animal models suggest that sulfur dioxide and ozone exposures can intensify
allergic asthma. Current standards, for the general environment as well as the workplace, particularly for sulfur dioxide, might be adequate to protect healthy individuals from acute bronchospasm, but may be inadequate to protect individuals who have asthma or who may be disposed to develop asthma.
Chemicals ingested in food can cause food hypersensitivity or food intolerance, a nonimmunologic response. Food hypersensitivity can be associated with a wide range of clinical symptoms that often are difficult to distinguish from food intolerance.
Similarly, exposure of skin to some chemicals can result in allergic contact dermatitis. Chemicals can also act as primary irritants or as agents that induce a nonspecific release of inflammatory mediators. Distinguishing among these conditions can present a diagnostic challenge.
Animal models used to predict the likelihood of chemicals to produce hypersensitivity may be limited in predicting the incidence of response in humans because of hereditary and psychologic factors that influence these reactions. Some models currently under development show promise as research tools to determine mechanisms of sensitization and to establish the toxic potential of xenobiotic substances.
Established and validated biologic markers of hypersensitivity include skin tests, in vitro assays for specific antibodies (IgE, IgG) or cellular immunity (lymphocyte stimulation), provocation challenges, and the cytologic evaluation of bronchoalveolar lavage fluid. These most frequently serve as markers of exposure or susceptibility. Our ability to develop biologic markers for hypersensitivity disease is impaired by our limited understanding of disease mechanisms, predisposing factors, and epidemiology of these disorders, as well as by species differences in response to chemical exposure.
RECOMMENDATIONS FOR FUTURE RESEARCH
IgE Sensitivity
Immunologically Mediated Adverse Reactions
It is difficult to differentiate between IgE-specific responses to chemical haptens and adverse reactions that cause clinical symptoms by nonspecific mechanisms (e.g., the release of mediators). Data are needed about the mechanisms of nonimmunologically mediated sensitization, including identification of the cells and mediators involved. The role of pulmonary inflammation produced by inhaled chemicals in the induction of asthma should be investigated.
Local Versus Systemic Immunity
The role of locally produced IgE, as opposed to systemic IgE, in the induction of clinical symptoms should be addressed, since clinical responses do not always correlate with the levels of systemic IgE. The severity of disease may be controlled by the level of IgE in local mucosal tissues.
Sensitizers
Many chemical sensitizers have been identified. The mechanisms of hapten binding, the translocation to lymphoid tissues, and the establishment of localized immunity in mucosal tissues and skin need to be examined. Studies are needed to determine whether there are differences in the mechanisms of sensitivity to high-versus low-molecular-weight allergens. These studies should include comparisons of initial induction of immunity in lymphoid tissues and the accumulation of immune cells in mucosal tissues and skin that influence localized immune responses.
Individual Differences in Sensitivity
We know that allergic diseases exist, but there is little information available to determine why some individuals become sensitized or have adverse reactions to chemicals and others do not. The differences between individuals need to be identified, and markers useful in identifying susceptible persons need to be developed.
Development of Self-Antigens
It is possible that altered self-antigens result from reactions between low-molecular-weight haptens and tissues. The response to altered self-antigens might be responsible for the production of disease in some individuals. The possible existence of self-antigens after exposure to chemicals should be examined. Such products (adducts) would also serve as markers of chemical exposure.
Effects of Exposure Conditions
Animal data indicate that the incidence and severity of pulmonary and dermal hypersensitivity reactions are related to the amount of exposure. More studies are needed on the influence of dose and dose schedule on sensitization and its clinical manifestation.
Epidemiology
Although information on hypersensitivity and adverse reactions to chemicals is frequently updated, better data are needed to define the magnitude of this problem. This is especially true for adverse reactions to ingested chemicals. There are few epidemiologic studies to indicate the incidence of hypersensitivity or intolerance to food additives.
IGE AND CELLULAR IMMUNITY
Cellular and Antibody Immunity
Cellular and antibody responses have been identified in hypersensitivity disease, such as in hypersensitivity pneumonitis. However, data are still needed to clarify the roles played by antibody and cellular immunity in the production and maintenance of disease.
Individual Variability
Most exposed individuals develop an immune response to inhaled antigens. The reasons that only a few individuals develop lung disease are not known, but it seems likely that disease is produced selectively because of disorders in immune regulation in the lung. More data are needed to differentiate the exposures and responses that lead to immune-mediated disease from those that produce immunologic changes but not disease. For example, many exposed individuals have large numbers of lymphocytes in their lungs, yet are asymptomatic. What is the function of these immune cells? Is it possible that they provide immune protection by assisting in the recognition and elimination of offending antigens?
Local Versus Systemic Immunity
Systemic immune responses can be measured in patients with hypersensitivity disorders. However, the role of systemic responses, compared with the induction of local immunity, is not understood. Local immune responses in tissues could be considerably more important than systemic immunity in the production of clinical symptoms.
Organic Versus Chemical Hypersensitivity
Many biologic materials have been identified
that can induce hypersensitivity pneumonitis. Relatively few chemicals, however, have been associated with this disease. Studies are needed to determine whether immune-system responses to chemical haptens are as likely to induce hypersensitivity pneumonitis as are immune-system responses to inhaled organic antigens.
Effects of Exposure
It appears that peak exposure is more important than time-weighted exposure in the induction of hypersensitivity to chemicals. Data are needed to determine the nature of exposures most likely to lead to induction of sensitivity to chemical haptens. Clarification is needed of both induction of sensitivity and development of disease after brief high-level exposures and the effect of repeated exposure over defined periods.
Particle Composition
Many antigens that lead to hypersensitivity are complex (high-molecular-weight) materials. Data are needed to illuminate the effects of particle composition on the induction of sensitivity and clinical disease.
Chemical Haptens
Additional data are needed on the mechanisms involved in the induction of immunity to chemical haptens, such as TDI and beryllium. In most instances, the biomolecules bound by the hapten have not been identified.




















