
2
Biologic Basis of Neurotoxicity
The nervous system is the communication and information-storage system of the body. In addition to coordinating functions that are commonly associated with the brain, such as thinking, the nervous system plays a major role in physiologic regulation. Virtually all physiologic functions are influenced or controlled by the nervous system. Other systems, such as the circulatory and reproductive systems, provide information to the nervous system, which in turn controls their functioning. The nervous system also integrates the functions of the various organ systems. Thus, neurologic dysfunction has a major impact that goes beyond the nervous system itself. Conversely, abnormal functioning in other systems can modify neurologic functioning.
Our abilities to act, to feel, to learn, and to remember all depend on the complex anatomic arrangements and proper integrative functioning of the nervous system, of which we have only a rudimentary understanding. The ultimate goal of neuroscience is to understand how this system functions normally and how subtle perturbations can lead to disease and dysfunction. Observation of the response of experimentally exposed animals or inadvertently exposed humans to neurotoxic chemicals advances our understanding of neurobiology while generating information about the consequences of exposure to environmental pollutants.
The first two sections of this chapter summarize our current understanding of the nervous system. The next section discusses several aspects of the nervous system that make it especially vulnerable to chemically induced damage. The fourth section describes examples of neurotoxicant-induced damage of several types. A final section summarizes the problems arising from the nature of neurobiology that must be addressed when a substance is tested for neurotoxicity, the subject of Chapter 4.
CELLULAR ANATOMY AND PHYSIOLOGY
The nervous system is composed of two general types of cells, nerve cells (neurons) and glial cells, which are present in approximately equal numbers. Neurons are responsible for the reception, integration, transmission, and storage of information. They can be classified as interneurons and motor, sensory, and neuroendocrine neurons. Motor neurons innervate and elicit contraction of muscle fibers. Sensory neu-
rons monitor the organism's external and internal environments. Specialized receptors on sensory nerves convey information regarding light, sound, muscle position, taste, and smell to the central nervous system. In many cases, sensory neurons form the input (afferent) side of information loops in the central nervous system; the output (efferent) side is mediated by motor fibers or by the endocrine system. Neuroendocrine neurons elicit secretions from glandular cells. Interneurons facilitate or modulate interactions between other neurons, often in complex feedback loops. Glial cells differ from neurons in their ability to divide throughout the organism's life, particularly in response to injury, and in their functions (Table 2-1), which are less well understood than those of neurons, but are believed to be largely restricted to structuring and regulating the environment surrounding the neurons. Glial cells also play a major role in myelination.
Structure of the Neuron
A neuron (Figure 2-1) has a more or less round cell body (soma), which contains its nucleus and organelles for protein synthesis. The diameter of the cell body of a vertebrate animal is about 10–100 µm (the largest would be roughly the size of the period at the end of this sentence). The neuron gets its distinctive appearance from its dendrites and axon, elongated processes that emanate from the cell body. The length and branching patterns of the dendrites vary from one type of neuron to another. Dendrites and their branches greatly increase a neuron's surface area, and this increases the number of messages that the neuron can receive and the number of sources from which they can come. The arrangement is analogous to the branching of a tree, which increases its reception of sunlight by increasing its total leaf surface area.
In most neurons, the axon is a thin, particularly elongated cellular process emanating from the neuronal soma and is specialized for the conduction of nerve signals away from the cell body and toward its synapse with other cells (other neurons, muscle cells, or gland cells). One neuron communicates with another by releasing chemical signals onto a small gap, or the synapse, between the tip of an axon and the dendrites or soma of its target cell. Axons are shorter or longer, depending on the type of neuron, but an axon's length is generally tens or thousands of times greater than the diameter of the cell body. An extreme example is the axon of a motor neuron that innervates the
TABLE 2-1 Nonneuronal (Glial) Cells of the Nervous System and their Function
|
Glial Cell Type |
|
Functions |
|
Astrocyte |
|
Repair, structural support, transport |
|
Oligodendrocyte |
|
|
|
|
Satellite |
Neuronal maintenance |
|
|
Intermediate |
Potential for maintenance or myelination |
|
|
Intrafasicular |
Myelination |
|
Microglia |
|
Phagocytosis |
|
Ependymal cells |
|
Service buffer between brain and cerebrospinal fluid |
|
Schwann cells |
|
Myelination of peripheral nerves |
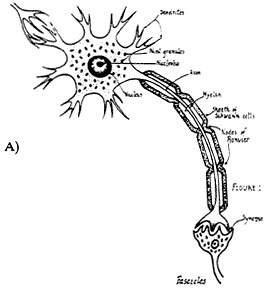
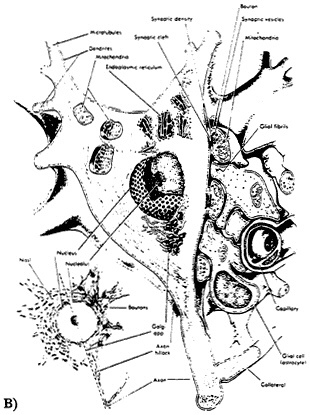
Figure 2-1 Diagrammatic representation of neuronal structure. A) The drawing illustrates the gross structure of a neuron with its many dendrites, myelin sheath, and synaptic contact with another neuron. Source: Liebman, (1991). B) The drawing shows structures visible at the electron microscope level. The nucleus, nucleolus, chromatin, and nuclear pores are represented. In the cytoplasm are mitochondria, rough endoplasmic reticulum, the Golgi apparatus, neurofilaments, and microtubles. Along the surface membrane are such associated structures as synaptic endings, astrocytic processes, and a capillary containing a red blood corpuscle.
Source: Willis and Grossman, (1973).
muscles in the human toe. Its cell body (a fairly large one) lies in the spinal cord at the level of the middle of the back; its axon is more than a meter long. Near its target, an axon divides into many small branches called telodendria. Telodendria end in small enlargements called synaptic boutons, which contain the chemical messengers that will be released to the target cell. A single axon can end in thousands or hundreds of thousands of synaptic boutons, by which a nerve can communicate with a few or many thousands of target cells.
Many axons are insulated by myelin, the extended and modified plasma membrane of specialized glial cells. During early development, myelin is deposited in a spiral fashion to form a sheath around the axon. Myelin is richer in lipid than are other membranes, and several unusual lipids are found in the myelin sheath; some well-characterized proteins are peculiar to myelin. Myelin of the central nervous system is made by oligodendrocytes and differs from the myelin of the peripheral nervous system, which is made by Schwann cells.
The myelin sheath formed by one oligodendrocyte or Schwann cell covers only a short length of the axon; adjacent lengths of axon are ensheathed by adjacent myelinating cells. Very short lengths of bare axon between myelinated areas are called nodes of Ranvier. Within myelinated axons, a nerve impulse travels by jumping from one node to the next. Compared with conduction in axons that are not myelinated, this mode of travel, called saltation, increases the speed of impulse conduction by a factor of up to 100. In a poorly understood metabolic dependence between a myelinated axon and its ensheathing cells, some types of injury to one cause dysfunction in the other.
The Nerve Impulse
Two processes are involved in communication by neurons: electric (discussed here) and chemical (discussed in the next section). Within a neuron, transmission is electric and analogous to that in a battery. The axonal membrane is semipermeable to the positively and negatively charged ions (chiefly, potassium, sodium, and chloride) that are in solution inside and outside the cell. In the absence of active and directed metabolic transport, those ions would be distributed equally on each side of the axonal membrane, and the inner and outer electrochemical potentials would be the same. However, several enzyme systems normally transport ions actively from the inside of the axon to the outside or from the outside to the inside. As a result, the axonal membrane separates areas of unequal electrochemical potential; i.e., it is polarized and generates a resting potential (Davies, 1968). The situation is analogous to a flashlight battery, where an impermeable membrane (a strip of cardboard) separates two fluids (pastes in a dry-cell battery) that contain ions of positive charge on one side and negative charge on the other. The neuronal "battery" must be maintained by continuous active transport of ions across the membrane.
The action potential is generated as a result of opening and closing of various ion channels in the nerve membrane (Hille, 1984). Several types of ion channels are known, but the most common are sodium and potassium channels, which are tiny pores that allow sodium and potassium ions, respectively, to pass through. The action potential is propagated regeneratively along the nerve fiber without loss of amplitude. In the resting state without stimulation, the potassium channels open and close occasionally, and the sodium channels remain mostly closed. Therefore, the resting nerve membrane is permeable mainly to potassium, but only sparingly to sodium. That results in the resting potential, with the cell's interior being negative with respect to the outside by 70 mV or so. On electric stimulation, however, many sodium channels start opening,
and that makes the membrane permeable predominantly to sodium. Electric stimulation thus generates an action potential with a peak some 50 mV more positive on the interior than the exterior of the nerve. Soon the sodium channels start closing and the potassium channels start opening at a high frequency; this brings the membrane potential back to the resting level. During the action potential, sodium ions enter the cell through open sodium channels and potassium ions leave the cell through open potassium channels; the result is a slight imbalance of the internal sodium and potassium concentrations, which is corrected quickly by a metabolic pump that extrudes extra sodium and absorbs potassium. Thus, the cell is capable of generating many action potentials in rapid succession.
The ion channels are the primary target sites of some toxicants and therapeutic drugs. For example, tetrodotoxin (a puffer fish poison) and local anesthetics block the sodium channel, thereby impairing the impulse conduction. The insecticidal pyrethroids and DDT keep the sodium channels open for unusually long periods, making the nerve generate impulses in a hyperactive fashion.
Synaptic Transmission
When a nerve impulse reaches a terminal branch of an axon, it depolarizes the synaptic boutons. That initiates the second type of transmission by means of the cellular release of special messenger chemicals (neurotransmitters) that are stored in the boutons. The amount of secretion is related to the degree of depolarization, which, in turn, is a function of the number of nerve impulses that reach a synaptic bouton. A very narrow gap, the synapse, separates the bouton from its target cell (Figure 2-2). The released chemicals diffuse across the synapse and attach themselves to specific receptor molecules embedded in the membrane of the target cell. Attachment (binding) initiates one of several responses, usually involving changes in ion flux, depending on the type of target cell and the type of messenger-receptor pair involved. For example, when the target cell is a muscle cell, the neurotransmitter-receptor interaction leads to contraction of the muscle; when the target is a gland cell, the interaction leads to secretion.
Synaptic transmission between neurons is slightly more complicated. Messenger molecules from the bouton of a presynaptic (releasing) cell bind to receptors on the surface of the postsynaptic (receiving) cell; that can lead to either excitation or inhibition of the target neuron. Neuronal cell bodies and dendrites are polarized in the same way as the axon. At an excitatory synapse, the neurotransmitter-receptor interaction leads to a momentary opening in ion-specific channels that produces depolarization (excitatory postsynaptic potential). At an inhibitory synapse, a different neurotransmitter-receptor interaction leads to a momentary opening of different ion-specific channels that produces an increase in polarization (hyperpolarization) or a small depolarization (inhibitory postsynaptic potential). Unlike the membrane of the axon, the membrane of the cell body and dendrites does not generate nerve impulses. Instead, the sum of all the excitatory and inhibitory postsynaptic potentials determines a moment-by-moment average polarization that is monitored by the portion of the cell's axon adjacent to the cell body. When the running average exceeds a threshold depolarization at the axon's initial segment, a nerve impulse is generated and begins its electrochemical journey down the axon.
The duration of neurotransmitter action is a function primarily of the time that it remains in the synapse. Under normal circumstances, that is a very short time (a few thousandths of a second), because specialized enzymes quickly degrade the neurotransmitter, and, in some cases, metabolically driven re-uptake systems rapidly scavenge
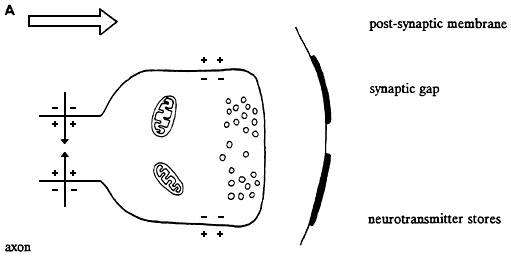
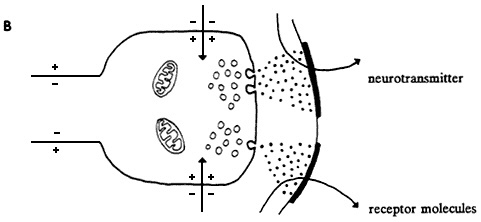
Figure 2-2 Events in chemical synaptic transmission. A, action potential shown approaching axon terminal containing transmitter stored in synaptic boutons. B, arrival of nerve impulse triggers release of transmitter into synaptic gap. Transmitter diffuses across synapse and reacts with receptor molecules associated with postsynaptic membrane. Resulting change in membrane permeability to ions, such as sodium and potassium, causes flow of synaptic current. In this case, current flows inward through postsynaptic membrane and results in depolarization called excitatory postsynaptic potential. Later transmitter action is terminated by enzymatic destruction, diffusion, or re-uptake. Source: Willis and Grossman (1973).
the synapse for neurotransmitter and transport it back into the synaptic bouton of the releasing cell.
Synaptic Messengers
Several dozen neurotransmitters are known, and more are discovered every year. (A partial list is given in Table 2-2.) A given neuron can synthesize and release a single neurotransmitter or, in the case of peptidergic neurons, several neurotransmitters. Similarly, a neuron can contain receptors for and be influenced by several neurotransmitters.
TABLE 2-2 Neuron Type Classified by Neurochemical Released for Synaptic Transmission
|
Neuron Type |
Neurotransmitter |
|
Cholinergic |
Acetylcholine |
|
Excitatory amino acid |
Glutamate, aspartate |
|
Inhibitory amino add |
γ-Aminobutyric acid (GABA), glycine |
|
Catecholaminergic |
Dopamine, epinephrine, norepinephrine |
|
Sertoninergic |
Sertonin |
|
Peptidergic |
Substance P, enkephalins, endorphins, neurotensin, carnosine, angiotensin II, pituitary hormones and their hypothalmic controlling factors |
|
Other |
Histamine, taurine, proline, N-acetyl-L-aspartate, serine, octop amine, p-tyramine, N-acetyl serotonin, tryptamine, purine, 5-methoxytryptamine, pyrimidine derivatives |
Research on the functions of the multitude of synaptic messengers is one of the most active fields in neuroscience today. Although not all the details are known, one conclusion is clear: neurons communicate by a complex code that involves a large number of chemicals and exquisitely specific chemical interactions.
GENERAL ASPECTS OF NERVOUS SYSTEM STRUCTURE AND FUNCTION
The nervous system has two parts: the central nervous system (CNS) and the peripheral nervous system (PNS). The CNS comprises the brain and the spinal cord. The PNS is composed of nerve cells and their processes that lie outside the skull and vertebral column. The nerve cells of the PNS are generally found in aggregates called ganglia. The neurons of the CNS are often segregated into aggregates called nuclei. Various nuclei are functionally related to one another to constitute levels of organization called systems; thus, we speak of the motor system, the visual system, and the limbic system.
The cells of the nervous system are organized in networks of increasing evolutionary complexity. In anatomic terms, the most primitive level is the spinal cord, wherein responses to sensory signals are virtually instantaneous; such responses are called reflexes. Spinal-cord functions tend to have been conserved in evolution, so the system in lower vertebrates resembles that in humans. The oldest regions of the brain regulate involuntary activities of the body, such as breathing and blood circulation, and exert much of their influence via the reflexes of the spinal-cord. The brain stem—which comprises the medulla, pons, and mid-brain—has functions related primarily to motor and sensory activities of the head and neck, including hearing, balance, and taste. The diencephalon is composed of the thalamus and the hypothalamus. The thalamus processes most information from the rest of the nervous system on its way to the cerebral cortex, and the hypothalamus regulates autonomic, endocrine, and visceral integration. The basal ganglia assist the cerebral cortex
with motor functions. The cerebral cortex, the most highly evolved level, is a vast information-storage and-retrieval area.
In addition to its role in such functions as perception and learning, the nervous system has a critical role in controlling the body's endocrine glands. That control is concentrated in several small nuclei at the base of the brain that are known collectively as the hypothalamus. Nerve cells in the hypothalamus secrete chemical messengers into a short loop of blood vessels that carries them to the pituitary, the body's master gland. The pituitary, in turn, releases chemical messengers into the general circulation. The pituitary messengers regulate other glands, such as the thyroid, the adrenals, and the gonads. The entire system maintains homeostasis, i.e., a regulated state of optimal physiologic functioning or regular cycling throughout the body.
Metabolism
The CNS has a very high rate of metabolism. In an average adult human, the weight of the brain (about 3 lb) is approximately 2% of total body weight. But its energy demands are so great that it receives about 14% of the heart's output and consumes about 18% of the oxygen absorbed by the lungs (Milnor, 1968). Unlike other organs, the brain depends almost entirely on glucose as a source of energy and as a raw material for the synthesis of other molecules (Damstra and Bondy, 1980). The CNS depends critically on an uninterrupted supply of oxygen and on the proper functioning of enzymes that metabolize glucose. Other tissues (muscle, for example) have alternative sources of energy and can survive for a relatively long time when the circulation is compromised, but nerve cells usually begin to die within minutes.
Blood-Brain Barrier
With few exceptions (e.g., proteins), substances that are carried in the bloodstream can pass from capillaries to surrounding extracellular fluid (in nearly all regions of the body). That is not true in the central nervous system (Betz et al., 1989). The cells that form the walls of capillaries in the CNS abut one another with unusually tight junctions. Those tight junctions, and perhaps other cells that line the outside of the capillaries, form a barrier that prevents the free passage of most bloodborne substances and thereby helps to create fine control of the extracellular environment of nerve cells. (Peripheral nerves and the retina have a similar barrier.) Without this blood-brain barrier, the brain would be even more vulnerable to chemical damage.
Chemical access to cells varies in different regions of the nervous system and at different stages of development (Klaassen, 1986). The area postrema and circumventricular areas have no blood-brain barrier. In developing animals, monosodium glutamate selectively kills cells in the arcuate nucleus and the retina, where the barrier is insufficient to exclude it. Other areas have incomplete barriers; e.g., the dorsal root ganglion is vulnerable to penetration and resulting cell-killing by chemicals (such as doxorubicin) that are excluded from the nearby spinal cord by a more complete barrier (Cho et al., 1980), and the olfactory nerve directly connects the outside environment via the nose to the frontal cortex (Broadwell, 1989).
Most nuclei in the nervous system, particularly in the CNS, have relatively intact barriers and thus are protected from some classes of chemicals. Although those barriers exclude many chemicals from the brain, some neurotoxic chemicals can gain access to the CNS via normal uptake mechanisms, if they are structurally similar to endogenous substances normally taken into the brain.
As will be discussed in more detail in Chapter 3, the selectivity of the blood-brain barrier results in fairly specific biologic markers that correspond to particular substances. Because the CNS is enclosed in a fairly isolated compartment defined by the blood-brain barrier, monitoring of more readily sampled tissues, such as blood, is not usually sufficient for detecting biologic markers of exposure or effect in the CNS itself; for that purpose, it might be necessary to sample spinal fluid.
Development
The organizational complexity of the adult human nervous system is staggering. Indeed, the complexity is so great that we have only a rough idea of the number of neurons in the brain. The most widely cited estimate is 30 billion. It is reasonable to assume that an average neuron receives input from about 1,000 other neurons and, in turn, sends its messages to another 1,000 neurons.
Details of the development of the human nervous system are poorly understood, but the broad outline is known. It begins early in embryonic development and is not complete until about the time of puberty. It proceeds as a carefully timed multistage process guided by hormones and other chemical messengers. Some of the initial organization acts as scaffolding, i.e, a temporary framework for later development. The temporal and spatial organization of the developmental process is precise and complex, and the deleterious effects of a relatively minor disturbance—one that perturbs developmental interactions among just a few cells and for only a brief time—might be amplified enormously through the cascade of developmental steps. Developmental stage is important in determining the limits of plasticity, i.e., the relocation of cellular resources to overcome the effects of injury; this ability to adapt to damage is thought to be greatest early in development. For example, a lesion that might be compensated for in a developing nervous system might produce irreversible deficits in an adult. In contrast, interference with a particular cell-cell interaction can have qualitatively different effects in developing and mature nervous systems. For example, dopamine receptors respond to a decrease in dopamine input during development with a permanent decrease in their number, but in adulthood with an increase in their number (Rosengarten and Friedhoff, 1979; Miller and Friedhoff, 1986). Thus, the nervous system's developmental period is a time of both vulnerability and ability to compensate.
Disuse of a particular synaptic connection can result in its inactivation and eventual dissolution. When more than a critical number of synaptic connections on a particular neuron are inactivated, the neuron itself might become superfluous and be deleted. That phenomenon occurs extensively during development: excess neurons and synaptic connections are established, many of which apparently disappear during the modeling of the mature nervous system. The converse also happens: synaptic connections used repeatedly become fixed, and transmission at these locations is facilitated through long-lasting changes in how ion channels or receptors are linked to second messenger systems. It appears that the shape of a given synapse is plastic and that the arrangement of structural proteins within a synapse can be modified to alter the efficiency of information transmission.
Neuron production does not cease at birth in the human (or in any common laboratory species), but continues through most of the first year of postnatal life. However, neurogenesis for most regions is completed during embryonic stages or fetal stages. Thereafter,
neurons that die are not replaced, although supporting cells continue to divide. Later development, maturation, or repair of the nervous system involves extension of the population of neuronal processes (but not production of new neurons) and modification of interconnections. That is in marked contrast with most other tissues, in which cell replacement is continuous. The absence of neurogenesis in the adult places severe constraints on the nervous system's ability to recover from damage.
Plasticity
Although the general pattern of interconnections in the nervous system is fixed according to the genetic program that guides development, the fine structure of the pattern of connections in at least some regions probably responds to exogenous factors and retains a considerable degree of adaptability throughout life. This characteristic of plasticity involves rearrangement of the details of synaptic connections. Plasticity confers on the CNS the capacity to respond to environmental signals by learning and memory formation. Damage to the nervous system can also evoke rearrangement of connections among neurons that survive the insult; rearrangement permits a limited, but important, functional adjustment to compensate for neuronal damage.
The clearest example of neuronal plasticity is seen after damage to axons in peripheral nerves. Neurons supplying peripheral nerves usually survive when their axons have been damaged. Such a neuron is able to regenerate its axon, which then locates its original target. That is the basis, for example, of the eventual partial return of sensation and muscle control in a surgically reattached limb. Neurons in the CNS are also able to regenerate damaged axons, but it is far more difficult for them to reacquire their targets, partly because of the barrier formed by proliferating glia, partly because of the inherently more complex interconnections of the CNS, and partly because the CNS lacks a pathway for regenerating axons to follow, such as is formed in the PNS by basal laminae and collagen and reticular fibers of the endoneurium (the tissue that separates the fibers in a peripheral nerve).
Trophic Interactions
The functioning of individual neurons is determined partly by a continuous supply of trophic factors, chemicals that originate in the cell's targets. For example, if the axons of motor neurons are severed, both the motor neuron and the muscle cells atrophy because of the lack of the chemicals that they normally exchange. A target provides trophic feedback to the neuron that innervates it, so compromise of neuronal function can be secondary to injury to nonneuronal cells. Such interdependence is also seen among neurons. Some neurons atrophy when the neurons that normally communicate with them die (Kelly, 1985). Thus, it is possible that damage can spread from a small number of inactivated neurons.
VULNERABILITY OF THE NERVOUS SYSTEM TO CHEMICAL TOXICANTS
In general terms, neurotoxic injury involves adverse functional or structural change. But the different vulnerabilities of various cells to injury by a given toxicant and the normal roles of susceptible cells determine the biologic markers of effect or clinical manifestations of exposure to a particular neurotoxic chemical. For example, myelin degeneration after exposure to triethyltin or hexachlorophene is manifested as spasticity; effects on neurons in the hippocampus due to exposure to trimethyltin, triethyllead, or methylmercury are associated with learning and memory deficits; degeneration of dopa-
mine neurons in the substantia nigra due to MPTP exposure leads to the tremor and rigidity of parkinsonism; and axonal degeneration in the PNS due to γ-diketones or acrylamide is reflected in muscular weakness and sensory deficits. Such effects can often be detected in either exposed humans or corresponding experimental animal models with clinical examination or other diagnostic means, such as electrophysiology, computed axial tomography and positron-emission tomography (CAT and PET scanning), and magnetic resonance imaging. In contrast, some of the acute effects of toxic chemicals are manifested as dysfunction of the nervous system without any observable structural changes; examples are intoxication by insecticide pyrethroids, DDT, and malathion. Several factors render the nervous system especially vulnerable to chemical assault; they are described briefly in the next few pages.
Complexity of Structural and Functional Integration
The proper functioning of the nervous system depends critically on complex interactions among different cell types in many anatomic locations that communicate via a variety of electric and chemical signals. The sheer complexity of the system makes it vulnerable, just as the functioning of a computer is more vulnerable to damage than is the functioning of an abacus. Other organs also are complex and have many cells, but most have a high degree of functional redundancy. For example, the loss of a few liver cells creates little disturbance in the organ's function. In contrast, although the nervous system is able to compensate extensively after injury, its functional redundancy is nonetheless circumscribed. A small lesion might initially result in a functional deficit (for example, a small stroke might impair speech or motor coordination), but eventually be compensated for as new synaptic pathways are established. If a lesion is widespread, the corresponding functional capacity might be lost; for example, a relatively small loss of neurons that use acetylcholine as their neurotransmitter might produce a disturbance of memory, whereas a relatively minor insult concentrated in a subsystem that relies on dopamine as its neurotransmitter might have no effect, and a larger insult might impair motor coordination.
Limitations on Repair
To understand neurotoxicity, one must remember that adults have no neurogenesis. Nerve cells that are destroyed by chemical insult are not replaced; the lesion is permanent. However, the nervous system has remarkable compensatory mechanisms. Such compensation is evident, for example, in the development of tolerance to morphine and alcohol, the long-term potentiation phenomenon documented in experimental animals, functional recovery after stroke or brain surgery, and neuronal sprouting after axonotomy. However, the scantiness of functional redundancy and the lack of neurogenesis limit (although they do not preclude) compensatory ability, particularly after extensive neuronal death. And the limits are exacerbated by normal aging processes: Neurons die throughout adult life, and additional cell loss due to exposure to toxicants may cause a further deficit in the functional reserve that is ordinarily drawn on in aging.
Accessibility to Lipid-Soluble Toxicants
All cell membranes are composed chiefly of lipid molecules. However, the lipid content of the nervous system is especially high—the myelin in axonal sheaths, for example, constitutes approximately 25% of the brain's dry weight. Many classes of toxicants, such as chlorinated hydrocarbon
pesticides and industrial solvents, dissolve readily in lipids. Molecules dissolved in membrane lipids are relatively protected from enzymatic degradation and sheltered from the general circulation. Thus, lipids act as depots where various toxicants can accumulate and reach high local concentrations. Toxicants can gradually leach out of lipid depots, such as liver and adipose tissue, to such an extent that the exposure of the nervous system itself to a toxic chemical lasts much longer than the external exposure of the organism (Boylan et al., 1978). Toxicant concentrations in adipose tissue can reflect exposure of other lipid-rich tissue, such as is found in the nervous system, but determination of the temporal course of exposure or of tissue doses in the CNS is more problematic.
Dependence on Glucose
The nervous system depends almost exclusively on glucose for energy and as a starting material for the synthesis of other molecules (Sokoloff, 1989). Its energy requirements, e.g., for maintaining electric potentials for meeting the bioenergetic demands of neurotransmitter reuptake, and for performing axonal transport are high. The system is therefore particularly vulnerable to chemicals that inhibit the enzymes that metabolize glucose.
Axonal Transport
All types of cells must transport proteins and other molecular components from their site of production near the nucleus to other sites in the cell. Intracellular transport is relatively simple in most cells, because the greatest distance to be traversed is only a small fraction of a millimeter. Neurons are unique among cells, in that the cell body not only must maintain the functions normally associated with its own metabolic support, but also must provide support to dendrites and axons, often over relatively vast distances. The axon and synaptic boutons, have little ability to synthesize the materials needed to sustain their structure and function, so they depend on delivery of the materials from their parent-cell body (Hammerschlag and Brady, 1989). Delivery of such substances by intracellular transport down the axon (axonal transport) is highly vulnerable to interruption by toxic chemicals. In addition, the functional integrity of the neuronal cell body often depends on a reciprocal supply of trophic factors from the cells that it innervates, and these trophic factors must also be supplied via axonal transport over long distances with very high energy requirements.
Axonal transport has several forms. Fast anterograde (forward) transport carries macromolecular assemblies of primarily membrane-associated glycosylated or sulfated proteins along structural tracks of microtubules from cell body to distal axon at a rate of about 400 mm/day. From the cell body, slow anterograde transport carries soluble enzymes involved in metabolism and neurotransmission at 1–2 mm/day in conjunction with microtubules and neurofilaments or at 5–10 mm/day in association with microfilaments. Fast retrograde (backward) transport carries a variety of materials up the axon to the cell body at about 250 mm/day; its cargo might be endogenous molecules (largely glycoproteins) destined for recycling, trophic signals, or exogenous materials (e.g., tetanus toxin, lead, and doxorubucin, which might circumvent the blood-brain barrier by traveling up the axon from the PNS).
It has been hypothesized that the great demand of axonal transport for energy derived from oxidative metabolism is a point of neural vulnerability to some toxicants. Toxicants that interfere with that metabolism or that disrupt the spatial arrangement or manufacture of neurofilaments block axonal transport. Many chemicals that produce distal axonopathies probably act by such mechanisms. For example, γ-diketones
(metabolites of n-hexane and methyl n-butyl ketone) appear to exert their toxic effect via the mechanism of neurofilament cross-linking (St. Clair et al., 1989), whereas several biochemical mechanisms (e.g., reaction of sulfhydryl moieties and altered glycolysis) might contribute to acrylamide's impairment of axonal transport (Miller and Spencer, 1985).
Synaptic Transmission
Neurons communicate with one another and with muscle and gland cells via a large number of chemical messengers that interact in precisely defined ways with specialized receptor molecules. Chemically mediated communication is vulnerable to disruption by exogenous chemicals.
Disruption can occur in several ways. For example, it is functionally important for synaptically released neurotransmitters to have brief effects. Some neurotoxicants, such as the organophosphate pesticides, inhibit the enzyme that terminates the effect of the neurotransmitter acetylcholine on its target. The result is an injurious overstimulation of the target cell. Other neurotoxicants, particularly toxins and their chemical relatives, mimic the action of a neurotransmitter by interacting with its receptor molecule. For example, LSD (lysergic acid diethylamide), a synthetic relative of a natural toxin (ergot) produced by a fungus that infects cereal grain, interacts with the receptor for the neurotransmitter serotonin (Aghajanian, 1972).
Other neurotoxic chemicals act by deranging other aspects of chemical neurotransmission. Some chemicals interfere with the synthesis of particular neurotransmitters, and others block a neurotransmitter's access to its receptor molecule (Marwaha and Anderson, 1984). Neuroactive pharmaceuticals commonly have those types of mechanisms of action. Some neurotoxicants are metabolized by neuronal enzymes and produce damaging metabolites. For example, 6-hydroxydopamine is taken up by the reuptake system at synapses that use dopamine as a neurotransmitter. The metabolism of 6-hydroxydopamine within the synaptic bouton yields hydrogen peroxide and other toxic species, which kill the cell (Graham et al., 1978).
Ion Channels
A nerve impulse depends on the proper functioning of ion-specific channels in the membrane. During the rising phase of the action potential, sodium ions enter the neurons through open sodium channels. During the falling phase, potassium ions leave neurons through open potassium channels. Thus, the internal sodium concentration increases and the internal potassium concentration decreases during an action potential. Although the resulting ionic imbalance is small (around 0.1% for an axon 1 µm in diameter), it must be restored. That is accomplished by the sodium-potassium pump, which extrudes sodium and absorbs potassium. The ion pump is operated by metabolic energy, so substances that inhibit metabolic enzymes cause sodium to accumulate in the neurons and potassium to be lost; that leads to membrane depolarization and, eventually, the loss of excitability. Several kinds of neurotoxic chemicals—including such local anesthetics as lidocaine and such natural toxins as tetrodotoxin and saxitoxin—block the sodium and potassium channels and thereby halt the electric conduction of nerve impulses.
EXAMPLES OF NEUROTOXIC MECHANISMS
In some kinds of neurotoxicity, understanding of the pathogenetic progression from molecular lesion to clinical manifestation is well developed. Pyrethroids cause acute, but reversible, neurotoxic effects. Chronic, but not acute, exposure to n-hexane results in
degeneration of motor and sensory axons. MPTP, in sufficiently high doses, causes acute degeneration of dopamine neurons, which results in a severe parkinsonian syndrome; however, it is suspected that the progressive reduction in dopamine neurons that occurs with age will result in additional cases of parkinsonism in people exposed to smaller amounts of MPTP (Calne et al., 1985). The possibility of long latency between exposure to a toxicant and clinical manifestation of disease has profound implications both for demonstrating a toxic etiology in degenerative CNS disease and for testing chemicals for neurotoxicity.
Pyrethroids
Pyrethroids are synthetic analogues of pyrethrins, the active substances in the flowers of Chrysanthemum cinerariaefolium. The pyrethrum insecticides (made from the dried flowers) had been used extensively until the end of World War II, but a variety of potent synthetic insecticides developed after the war—such as DDT, lindane, dieldrin, parathion, and malathion—made pyrethrum almost obsolete. The pendulum swung in the 1960s, when the persistence of pesticides became a major issue in their use. Many pyrethrin derivatives were then synthesized and tested as insecticides, and some of them have proved extremely useful and safe. The pyrethroid insecticides now in use are characterized generally by potent insecticidal activity, relatively low acute mammalian toxicity, little known chronic mammalian toxicity (including mutagenicity and carcinogenicity, although information is incomplete), and—what is very important—environmental biodegradability.
Among insecticides, pyrethroids and organophosphate-carbamate anticholinesterases have been studied most thoroughly for their mechanisms of action. Pyrethroids exert primarily acute toxicity, so studies have focused on how the signs of poisoning are produced. The major aim of the studies has been to generate information for improving pyrethroids as insecticides.
Pyrethroids can be classified into two large groups (Figure 2-3). Type I pyrethroids do not contain a cyano group in their molecules and include allethrin, tetramethrin, permethrin, and phenothrin. Type II pyrethroids contain a cyano group at the a-carbon position and include newer compounds, such as deltamethrin, cyphenothrin, cypermethrin, and fenvalerate. The two types of pyrethroids cause somewhat different symptoms of mammalian poisoning. Poisoning with type I pyrethroids is characterized by hyperexcitation, ataxia, convulsions, and eventual paralysis; poisoning with type II pyrethroids, by hypersensitivity, choreoathetosis, tremors, and paralysis. Despite differences in the symptoms, both types of pyrethroids have the same major target site: the sodium channel of nerve membrane, i.e., the channel directly responsible for generating action potentials.
Pyrethroids cause essentially similar signs
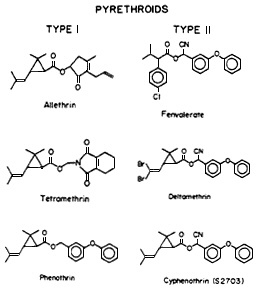
Figure 2-3 Structures of type I and type II pyrethroids. Source: Narahashi (1985).
of poisoning in mammals and invertebrate animals. In vitro studies of their mechanism of action on the nervous system have used nerve preparations isolated from insects, crayfish, and squid and cultured mouse neuroblastoma cells (Narahashi, 1971, 1985, 1989; Ruigt, 1984; Wouters and van den Bercken, 1978).
Pyrethroid poisoning is associated with an increase in electric activity of the central and peripheral nervous systems: a single presynaptic stimulation in the presence of pyrethroids causes repetitive excitation of presynaptic nerve and nerve terminals, thereby evoking repetitive postsynaptic discharges. This can be demonstrated by exposing isolated nerve fiber preparations, such as crayfish giant axons and frog myelinated nerve fibers, to low concentrations of pyrethroids (especially type I pyrethroids). Intracellular potential recording experiments reveal that the depolarizing after-potential is gradually increased in amplitude and prolonged after application of pyrethroids, until it finally generates repetitive after-discharges (Lund and Narahashi, 1983).
How the depolarizing after-potential is increased by pyrethroids is best studied by voltage-clamp techniques, whereby membrane ionic currents can be recorded. The sodium current that generates action potentials is markedly prolonged after the preparation is exposed to pyrethroids, and that causes a sustained depolarization after an action potential (Narahashi and Lund, 1980). However, the sodium current thus recorded represents an algebraic sum of currents passing through a large number of open sodium channels. The activity of individual ion channels can be studied by patch-clamp techniques, which allow measurements of ionic currents flowing through individual open channels. Pyrethroids have been found usually long periods (Yamamoto et al., 1983). A sodium channel normally is kept open during a depolarizing step for only a few milliseconds, whereas a sodium channel exposed to pyrethroids can remain open much longer—even up to several seconds, depending on the pyrethroid in question. Type I and type II pyrethroids have similar effects, but the increase in open time is more pronounced with type II pyrethroids.
Pyrethroids exert another effect on sodium channels. A pyrethroid-exposed sodium channel can be opened at negative membrane potentials near the resting potential. Prolonged opening of many sodium channels near the resting membrane potential leads to the nerve-membrane depolarization observed in the presence of pyrethroids. Membrane depolarization produces several changes in nervous function: depolarization in sensory neurons sends massive discharges to the central nervous system, causing hypersensitivity to external stimuli and paresthesia or a tingling sensation in the facial skin; depolarization of presynaptic terminals increases transmitter release, thereby disturbing synaptic transmission; and depolarization beyond some magnitude blocks nerve conduction and results in paralysis. In addition to the modified electric behavior of the sodium channels, all the more readily detected changes are biologic markers of exposure to pyrethroids.
During the last several years, it has been hypothesized that the γ-aminobutyric acid (GABA) receptor-channel complex, rather than the sodium channels, might be the target site of type II pyrethroids. That was based on several observations, including pyrethroid inhibition of ligand binding to the GABA receptor channel (Lawrence and Casida, 1983). However, recent patch-clamp experiments with rat dorsal root ganglion neurons in culture have unequivocally demonstrated that, whereas sodium channel current undergoes drastic and characteristic prolongation during exposure to the type II pyrethroid deltamethrin, the GABA-induced chloride channel current remains unaffected (Ogata et al., 1988). The GABA receptor-channel system thus plays a negligible role in poisoning with type II pyrethroids.
Pyrethroids thus seem to modify the
gating kinetics of sodium channels, causing prolonged opening of individual channels, which leads to a wide range of toxic signs. Only a very small fraction of sodium channels need to be modified by pyrethroids to cause severe signs of poisoning (Lund and Narahashi, 1982). That is why pyrethroids are such potent insecticides.
Although the mechanism of action on nerve membrane sodium channels is generally the same in both mammals and insects, pyrethroids are less toxic to mammals than to insects. The selective toxicity to insects might be due to at least three major factors: pyrethroids are more effective on nerves at low temperature (Narahashi, 1971), so insects at room temperature would be affected more severely than mammals at 37°C; pyrethroids are detoxified by enzymes, and enzymatic reactions are slower at low temperature; and, because insects are smaller than mammals, pyrethroid molecules are likely to reach the target site before being inactivated metabolically.
In summary, pyrethroids primarily modify gating kinetics of sodium channels, and this action can account for the various symptoms of poisoning in animals. Future studies should be directed toward the molecular mechanisms whereby the pyrethroid molecule interacts with sodium channels, including the identification of the subunit of a channel to which pyrethroids bind.
γ-Diketones
At high concentrations, the simple alkane n-hexane (a glue solvent) can be acutely toxic; e.g., it can cause dizziness and eye, throat, and skin irritations, all of which are nonspecific biologic markers of exposure and effect. Those manifestations, however, are reversible. The progressive weakness and sensory loss that arise from chronic exposure to n-hexane are the major human health hazards that motivated its regulation.
After it was reported that workers exposed to n-hexane at high concentrations developed a motor-sensory peripheral neuropathy (Yamamura, 1969; Herskowitz et al., 1971), Schaumberg and Spencer (1976) showed that rats develop an identical neuropathy after inhaling n-hexane. The finding that humans exposed to a solvent mixture containing methyl butyl butane developed peripheral neuropathy identical with that found after n-hexane exposure (Billmaier et al., 1974) demonstrated the peripheral neuropathy as a biologic marker characteristic of the two chemicals.
Biopsies of humans and careful morphologic studies of exposed rats showed that exposure to n-hexane and exposure to methyl n-butyl ketone each resulted in large neurofilament-filled swellings of axons, with axonal degeneration distal to the swellings developing over time. Thus, a biologic marker was established that was useful across species, as well as for the chemical class. Cavanagh and Bennetts (1981) showed that the neurofilament-filled swellings in the CNS and the PNS uniformly developed along the distal portions of long axons; however, accompanying axonal degeneration was much less common in the CNS (Griffin and Price, 1980). Spencer and Schaumberg (1977a,b) classified the neuropathy as a central-peripheral distal axonopathy and observed that myelinated axons were more vulnerable than nonmyelinated axons. Swelling and distal degeneration were most prominent in myelinated axons of the largest diameter.
The common morphologic findings after n-hexane and methyl n-butyl ketone exposure suggested a metabolic relationship between the two toxicants, which was elucidated by Krasavage et al. (1980) and Couri and Nachtman (1979). Studies disclosed a common metabolite, 2,5-hexanedione (HD), a biologic marker of exposure that itself caused an identical neuropathy. The Krasavage group coined the term ''γ-diketone''
neuropathy to emphasize the importance of the gamma spacing of the two ketone groups (separated by two carbons; sec Figure 2-4). γ-Diketones and γ-diketone precursors produce a specific neurotoxic response not shared by diketones with α, β, or γ spacing (with zero, one, or three intervening carbons, respectively) (Spencer et al., 1978; Katz et al., 1980). However, 2,4-pentanedione, an α-diketone, produces a different form of neurotoxicity (Misumi and Nagano, 1984). The seminal observation regarding the significance of gamma spacing began the process of understanding the neuropathy at the molecular level.
The search for the fundamental mechanism focused on two concepts: inhibition of energy metabolism (Spencer et al., 1979) and neurofilament cross-linking (Graham, 1980). The key observation, however, was that lysyl amino groups of proteins are the primary target of γ-diketones and that pyrroles are the product of the reaction (DeCaprio et al., 1982; Graham et al., 1982).
A number of studies have tested the hypothesis that pyrrole formation is the initial step in the sequence that leads to neurofilament accumulation. Anthony et al. (1983a,b) showed that 3,4-dimethyl-2,5-hexanedione (DMHD) was a γ-diketone that
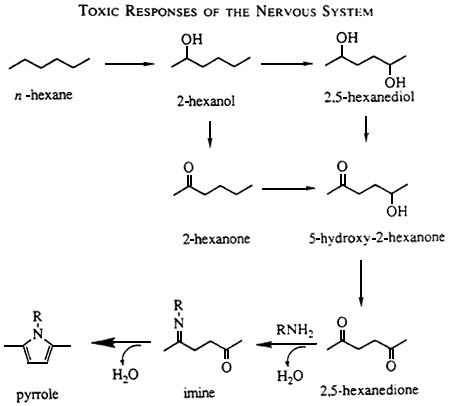
Figure 2-4 Metabolism of hexane. Both n-hexane and 2-hexanone (methyl n-butyl ketone) are neurotoxic, and both are activated through ω-1 oxidation to the ultimate toxic metabolite, 2,5-hexanedione. The toxicity of g-diketones derives from the ability of these diketones to react with protein amino groups (RNH2) to form an amine in an initial reversible step, and then to cyclize irreversibly to form a pyrrole. Source: Anthony and Graham (1991). Reprinted with permission from Amdur et al. (1991).
formed pyrroles more rapidly than HD and was correspondingly more neurotoxic. Larger substituents—such as ethyl, isopropyl, and phenyl groups on the 3 and 4 carbons—impeded pyrrole formation by steric hindrance (Szakal-Quin et al., 1986) and were not neurotoxic (Genter et al., 1987). Thus, although the gamma spacing is critical, not all γ-diketones are neurotoxic. Genter et al. (1987) clarified the role of pyrrole formation with the observation that the dl diastereomer of DMHD forms pyrroles more readily and is more toxic than the meso diastereomer. Given that dl-[14C]DMHD and meso-[14C]DMHD disperse comparably in the nervous system, as shown by Rosenberg et al. (1987), and that the tetramethylpyrroles formed are the same, the conclusion that pyrrole formation is the first step in the pathogenetic sequence was virtually ensured. The conclusion was strongly supported by the finding that 3,3-dimethyl-2,5-hexanedione, which cannot form a pyrrole, is not neurotoxic (Sayre et al., 1986) and by the reduced rate of pyrrole formation and neurotoxicity of deuterium-substituted HD (DeCaprio et al., 1988).
The significance of pyrrole formation for neurofilament function, however, has been a point of controversy. DeCaprio (1985) and Sayre et al. (1985) have proposed that conversion of the hydrophilic lysyl amino groups to hydrophobic pyrrole derivatives alters neurofilament transport through impairment of interactions between neurofilaments and other cytoplasmic components in the axon. The alternative hypothesis (Graham et al., 1982) has been that pyrrole autoxidation leads to covalent cross-linking of neurofilaments. Progressive cross-linking of neurofilaments during chronic intoxication would lead eventually to the formation of masses of neurofilaments too large to pass through the constrictions in axonal diameter that occur at every node of Ranvier. Indeed, the greater vulnerability of large-diameter myelinated axons is apparently explained by the greater proportional reduction in diameter at their nodes. Thus, continued transport up to the point of occlusion could account for the large neurofilament-filled swellings of the axon that most often occur proximal to nodes of Ranvier (Spencer et al., 1977a; Jones and Cavanaugh, 1983).
To test whether pyrrole derivatization itself is sufficient to account for the neurofilament-filled swelling or whether neurofilament cross-linking is necessary, St. Clair et al. (1988) used another γ-diketone, 3-acetyl-2,5-hexanedione (AcHD). The acetyl group of AcHD is small enough not to hinder pyrrole formation; in fact, the rate is nearly as great as that for dl-DMHD and greater than that for meso-DMHD. However, whereas the electron-donating methyl groups of the tetramethylpyrrole derived from DMHD result in a reduced oxidation potential (362 mV), compared with that of the dimethylpyrrole derived from HD (620 mV), the oxidation potential of the pyrrole formed by AcHD is much greater (975 mV), because of the electron-withdrawing properties of the acetyl group. Thus, the pyrrole formed from AcHD is relatively resistant to oxidation and so provides a probe for discriminating between the competing hypotheses. If pyrrole derivatization itself is sufficient, AcHD should be highly neurotoxic. Conversely, if pyrrole autoxidation and neurofilament cross-linking are required, AcHD would not result in neurotoxicity. When rats were given AcHD, massive pyrrole formation occurred, but neither clinical neurotoxicity nor neurofilament accumulation was observed; the results point strongly to the role of neurofilament cross-linking in the pathogenesis of n-hexane neurotoxicity.
Thus, in n-hexane neuropathy, it appears established that the γ-diketone HD is the toxic metabolite. Reaction with amino groups yields pyrrole adducts, which undergo autoxidation that results in covalent cross-linking of proteins. It is the extreme stability of the neurofilaments that results in the localization of toxic injury in the nervous
system, and it is the particular anatomy of the myelinated axon at nodes of Ranvier that makes these axons most vulnerable.
MPTP
A particularly unfortunate byproduct of the so-called designer drugs produced in the 1980s was 1-methyl-4-phenyl-1,2,3,6-tetrahydropyridine (MPTP) (Figure 2-5). Instead of the desired opiate-like characteristics of the intended meperidine derivative (which was to be sold illicitly as "synthetic heroin"), a synthetic drug that contained MPTP caused a profound, acute-onset neurologic disorder with extreme muscular rigidity and tremor that was indistinguishable from idiopathic Parkinson's disease (Langston et al., 1983). Animal studies showed that MPTP itself is not neurotoxic, but requires metabolism by monoamine oxidase B, a process that appears to take place in astrocytes, a type of glial cell (Jenner, 1989). The product of MPTP oxidation is the pyridinium ion MPP+ (Figure 2-5); this putative toxic metabolite of MPTP leaves the astrocytes and is preferentially concentrated by catecholaminergic neurons via their own catecholamine uptake system. Whether the mechanism of MPP+ cytotoxicity is related to oxidation-reduction cycling, interference with mitochondrial energy production, or some other process, the end result is cell death, particularly death of the dopaminergic neurons of the substantia nigra (Langston and Irwin, 1986). The marked depletion of dopamine seen in the brain of MPTP-treated
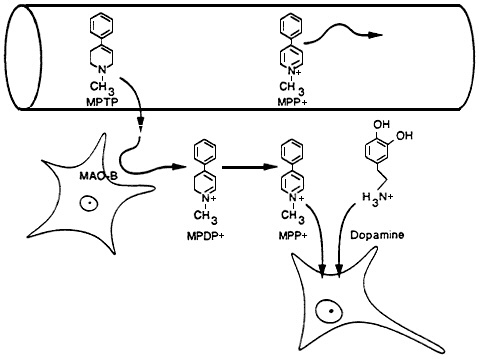
Figure 2-5 Diagram of MPTP toxicity. MPP+, either formed elsewhere in the body following exposure to MPTP or injected directly, is unable to cross the blood-brain barrier. In contrast, MPTP gains access and is oxidized in situ to MPDP+ and MPP+. The same transport system that carries dopamine into the dopaminergic neurons also transports the cytotoxic MPP+. Source: Anthony and Graham (1991). Reprinted with permission from Amdur et al. (1991).
animals is also the central neuropathologic feature of Parkinson's disease in humans.
Not all people exposed to MPTP have developed parkinsonism, ostensibly because the dose administered was sometimes insufficient to cause death of a large enough proportion of dopaminergic neurons to produce the 80% or greater depletion of dopamine in the brain that is required to produce a clinical syndrome (Hornykiewicz, 1986). Recently, a number of exposed people have been studied with positron-emission tomographic (PET) scanning (Calne et al., 1985). Through the use of positron-emitting fluorodopa, which is sequestered in the striatum (where the axonal projections of the substantia nigra form synapses), a deficiency of striatal dopamine was demonstrated. Although the exposed people were asymptomatic, the loss of dopaminergic neurons with normal aging (McGeer et al., 1977) is likely to reduce the population of these cells, and thus brain dopamine concentrations, below the 80% required for normal function; at that time, the signs and symptoms of Parkinson's disease would be manifested (Calne and Langston, 1983).
This is a critically important concept: An acute toxic injury can be separated from its clinical manifestations by years. If MPTP-exposed people who are now asymptomatic (and there might be over 500 such cases) eventually develop parkinsonism, it will open up the possibility that some patients who develop progressive degenerative CNS disease later in life (such as Parkinson's disease, Alzheimer's disease, and amyotrophic lateral sclerosis) might have suffered neuronal injury from a toxicant years earlier. If that turns out to have been the case, tests for neurotoxicity will have to take into account the possibility that effects develop after a long latency.
SUMMARY
The nervous system has several unique aspects, e.g., poor regenerative capacity, unusual anatomy, specialized metabolic requirements and subcellular systems, and a wide variety of neurotransmitter and neuromodulator systems. As the discussions of pyrethroids, γ-diketones, and MPTP illustrate, the complexity of the nervous system and its tremendous structural and functional heterogeneity might make it particularly susceptible to disruption by toxic agents, and neuronal dysfunction can have impacts on a number of organ systems. Even minor changes in the structure and function of the nervous system can have profound effects on neurologic, behavioral, and other body functions.
Many foreign substances can alter the normal activity of the nervous system via a variety of mechanisms. Some produce immediate short-term effects; others cause neurotoxic effects only after long-term, repeated exposures. Some substances can cause permanent damage after a single exposure; others can cause subtle and reversible damage. This diversity in nervous system response increases the challenge to toxicologists to develop biologically appropriate and empirically reliable test systems.
Considerable progress has been made in recent decades in understanding the nervous system on an integrated and comprehensive basis. Some mechanisms underlying behavior, such as sensorimotor function and information-processing, are being elucidated from the translation of molecular and cellular activities into organ-level events and from the translation of organ-level events into whole-organism behavior. Only by understanding the basic mechanisms involved in neurobiologic processes can we develop adequate neurotoxicity test methods. Our present understanding of some expressions of nervous system function permits such an integrated approach—e.g., much is known about the roles of molecules, cells, organs, and the whole organism in visual processing and proprioceptive control of movement. For most other neurologic functions, al-
though gaps in our knowledge still restrict integrative analyses, we can examine their components at specific levels of integration-with in vivo and in vitro approaches. An understanding of the nervous system guides the search for useful biologic markers, as is discussed in Chapter 3. This information is developed further in Chapter 4 to assess available in vivo and in vitro testing methods and address the need for efficient means for identifying neurotoxic substances.






















