
Engineering 3D Tissue Systems to
Better Mimic Human Biology
MATTHEW GEVAERT
KIYATEC Inc.
The scientific method—hypothesis-driven design and execution of an experiment—is great … except when it could kill you (or me). That’s why, for example, there are extensive legal requirements to investigate new pharmaceutical agents using proxies before testing drug toxicity in a human clinical trial. The US Food and Drug Administration requires a combination of nonliving techniques (biochemical assays and in silico analysis), in vitro models (i.e., cell culture), and animal studies before new compounds may be administered to humans.
Although pharmaceutical and regulatory industries are doing the best they can in the current paradigm, to be blunt it’s not going very well. According to recent publications, of all drugs that enter clinical trials, only 12% are eventually approved for use in humans (Paul et al. 2010). In other words, despite best efforts to predict those drug candidates’ efficacy and toxicity during preclinical testing, 88% of them fail—usually in terms of their lack of efficacy or unacceptable toxicity—when put to the test in humans.
A new paradigm is needed! And the biggest opportunity lies in cell culture, which typically is still done in a Petri dish (or its derivative, the multiwell plate). This ubiquitous scientific container, first described well over a hundred years ago in the late 19th century (Petri 1887), was already commonplace when cells were first widely cultured in the mid-20th century and remains the standard of cell culture today.
The vast majority of human cell types are adhesion dependent, and after fluid transfer to a Petri dish or well plate they attach to the bottom. Once attached, they normally proliferate and cover the entire bottom surface without stacking, forming a confluent, flat monolayer (shorthanded as “2D” cell culture). As evidenced by usage patterns, normal limitations of this experimental mode (the environment
is static, diffusion is passive, constant evaporation alters solute concentrations, frequent media changes are necessary, cell numbers plateau at confluence, the cell experiences stimuli largely unrelated to those it experiences in vivo) are viewed as less important than benefits (cells grow well, the approach is cost effective, 2D planes are easily imaged with inexpensive microscopes, existing body of data is 2D, granting agencies still fund it, and the method enables high throughput).
Yet, as I put it in a recent talk to a group of high school STEM whiz kids, “Your Petri Dish Is So 1887.”
SIGNIFICANCE
Few people ask (and fewer answer) these basic questions: Do the results of Petri dish—type cell culture experimentation mean anything? Are they at all relevant to the intent of the experiment, which in most cases is to model a process that occurs in the human body? Although the assumption is “yes,” in an increasing number of demonstrated cases the answer to these important questions is actually “no.”
A quantitative way to measure the “behavior” of a cell in culture is its gene expression. In a beautiful demonstration that answers the questions above, a comparison was made of key gene expression profiles of primary human cancers with comparative immortalized epithelial cells in 2D (Ridky et al. 2010). Tellingly, the correlation coefficient between the two datasets was 0.0. But there are much easier and cheaper ways to obtain datasets with exactly zero correlation to the behavior one is trying to characterize than to conduct 2D cell culture experiments!
The tremendous opportunity for improvement lies in the fact that cells are living organisms and can respond dynamically to local stimuli provided by and in their environment. The solution is to provide a different environment with more of the “right” physical, mechanical, and biochemical stimuli. Developments that address this challenge will affect much more than in vitro modeling of in vivo physiology. Aside from the desire to model human beings and the need to minimize the very serious consequences of the scientific method for certain kinds of questions, better in vitro systems have enormous implications as both manufacturing methods for implants (e.g., in tissue engineering and regenerative medicine) and as process steps for cell therapy.
ENGINEERING CELL SHAPE THROUGH MATERIAL INTERACTION
As a living entity, each cell has the potential to sense and respond to physical stimulus at each point in all its transecting planes—i.e., its entire surface in three dimensions.
When an adhesion-dependent cell is presented with a flat surface to which it can favorably attach, it tends to maximize its adhesion and adopts a primarily flat morphology. Cells in a 2D paradigm tie up approximately 50% of this interaction capacity with the bottom surface of the well plate, approximately 50% with the
liquid environment above the flat cell, and a very small amount in lateral cell-cell interactions.
The fundamental value proposition of “3D” cell culture is to provide a microenvironment in which the potential for physical interaction is distributed in a biologically relevant fashion across the entire surface of the cell. This is normally achieved by culturing cells in a scaffold or matrix material, which can span gels, fibers, or porous solids, among others. Cells in a 3D culture matrix adopt a more complex morphology (e.g., roughly ellipsoid) that is typically much closer to their morphology in their native state—that of a cell in tissue in a living organism.
Does this matter? Again relying on gene expression as a way to measure cell behavior, researchers have documented significant changes in gene expression profiles (recently genomewide) of multiple cell types as a result of 3D relative to 2D cell culture conditions. These changes have been shown to be associated with key biological processes such as tissue development, cell adhesion, immune system activation, and defense response (e.g., Zschenker et al. 2012). Thus, cell morphology is fundamentally deterministic of some important aspects such as cell behavior, signal transduction, protein-protein interaction, and responsiveness to external stimuli. Gene expression profiles in 3D are also shown to have much more relevance to those measured in vivo (Birgersdotter et al. 2005; Martin et al. 2008).
In addition to the value of a 3D microenvironment that more effectively models in vivo realities, this form-function relationship is also subject to manipulation toward less “natural” ends. Stem cells’ differentiation pathway has historically been controlled by soluble factor interactions, either from a second “feeder” layer cell type or as a result of soluble factors added to the cells’ media. Surprisingly, forcing a cell into a particular shape (e.g., the stars and flowers shown in Figure 1) by physical confinement can also affect its differentiation pathway even in the absence of soluble factor manipulation (Kilian et al. 2010).
Unfortunately, effectively engineering the 3D microenvironment is not as simple as providing physical interactions in three dimensions. Topography and mechanical stiffness are among biophysical cues in a 3D context that affect cell function. This is proven via either the addition of 3D topography (e.g., grooves, pillars, posts, pyramids, pits) to an otherwise flat surface via microfabrication techniques (wherein the cell is cultured on the material) or the incorporation of controlled topography internally and culturing of the cell in the material (Nikkhah et al. 2012). Topography can also induce effects that determine stem cell differentiation pathways (Kumar et al. 2012).
Mechanical stiffness affects cell behavior and function, as exemplified by the presence of an “edge effect” in 3D gel scaffolds. Fraley and colleagues (2011) characterized focal adhesions of cells embedded in a 3D collagen gel and reported that tension in the gel decreased with increasing distance from the container surface. Cellular focal adhesions, associated with each cell’s cytoskeletal structure, decreased as well. As shown in Figure 1, the authors were able to loosely qualify 2D (cell on surface), 2.5D (cell partially on surface), “3D near” (cell within
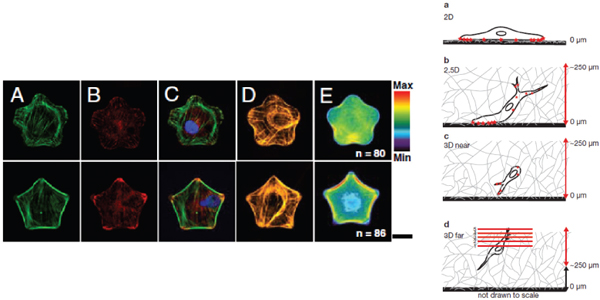
FIGURE 1 (Left) Immunofluorescent images and fluorescent heatmaps of cells in flower (top) and star (bottom) shapes, demonstrating differential cell response to nuanced physical constraints that influence the differentiation pathway. Reprinted with permission from Kilian et al. (2010). (Right) Schematic representation of focal adhesion visualization (stars on cell surface) in live HT-1080 cells cultured at increasing distances (a-d) from the dish bottom, characterizing edge effect in 3D matrix. Reprinted with permission from Fraley et al. (2011). This figure appears in color in the online posting of this article at www.nap.edu/catalog.php?record_id=18185.
250 μm of surface), and “3D far” regions based on the number of focal adhesions per cell.
Just how “3D” an environment is has very important implications for applications other than modeling. In a recent paper with profound ramifications for cell therapy, investigators demonstrated that a complex response (immunomodulation, e.g., the recruitment of monocytes to an inflamed endothelial monolayer) of cells in 3D was reduced fivefold compared to the same cells on 2D surfaces (Indolfi et al. 2012). The authors observed that the 3D cells had markedly altered cytoskeletal structure with rearranged focal adhesion proteins.
ENGINEERING THE SOLUBLE ENVIRONMENT
In addition to the interaction of a given cell with the materials and other cells surrounding it, the soluble environment has a considerable effect on cell behavior. At the most basic level, it is through the soluble environment that cells receive nutrients and perform basic functions such as respiration and waste elimination. Interference with these basic needs over time compromises the viability of the cell culture. Cells cultured on a 2D surface have nearly 50% of their surface area interacting with the soluble environment and simple, passive diffusion is usually more than sufficient to enable these processes. With frequent media changes to compensate for evaporation, depletion of nutrients, and generation of wastes, compromised viability of 2D cell cultures due to insufficient soluble environment interaction is rarely a concern.
However, inherent in soluble environment interactions and the frequent replacement of cell media is a cyclic change in the media pH and a “feast to famine” dynamic with respect to nutrient access. Media pH in typical 2D cell culture decreases over time (Wu and Kuo 2011) and differing pH levels have been shown to affect cell function (Wu et al. 2007). The removal of “spent” media, containing relatively fewer nutrients and more waste, also removes nonwaste excretions (e.g., proteins), an environmental change that may be directly related to the observable phenomenon that confluent cells in 2D culture do not typically stack but occur as monolayers. In one experiment, researchers began with cells in a typical 2D culture environment and, using a specialized bioreactor that allowed nutrient and waste exchange but preserved insoluble extracellular matrix secretions, created mineralizing, collagenous tissue up to 150 μm thick with as many as 6 cell layers (Dhurjati et al. 2006).
Depending on the density of both the 3D matrix and other cells, a particular cell’s soluble environment interactions can be severely compromised and result in muted function or eventually cell necrosis, particularly in the middle of the construct. The window for effective density management is significantly smaller if the in vitro model relies only on the passive diffusion that occurs with use of 3D scaffolds in static multiwell plates. The use of perfusion culture systems or bioreactors can minimize or alleviate the deleterious effects and stabilize the soluble
environment by avoiding feast-to-famine changes in nutrient availability and maintaining pH. Compared with static conditions, perfusion cell culture has been shown to affect culture morphology and organization (Tomei et al. 2009), increase key enzyme activity (Goldstein et al. 2001), increase mineral deposition and production of protein and cytokines (Gomes et al. 2003; Mercille and Massie 1999), increase cell penetration into and distribution throughout the scaffold (Cimetta et al. 2007; Goldstein et al. 2001; Gomes et al. 2003), increase cell viability especially at the center of cell-scaffold constructs (Cimetta et al. 2007; Mercille et al. 1999), and thus extend the effective duration of the culture experiment.
INCORPORATING BIOLOGICAL SYSTEMS EFFECTS WITH MULTIPLE CELL TYPES
Consideration of a particular cell’s interactions with other cells is essential to increasing the correlation of its in vitro functions and behavior to an in vivo organism. These interactions can take the form of direct cell-to-cell contact or of soluble factor interactions mediated by the environment. Human biology relies on both modes of interaction. Culture-based intercellular interactions among cells of the same type (monocultures) in 3D have been implicitly included in the discussions above, and are at least partially responsible for the morphological changes and functional benefits described.
A second type of intercellular interaction can be modeled via coculture of different types of cells, which can occur in the same culture chamber and create direct cell-to-cell contacts (a mixed coculture) or in multiple, separate chambers with a connected soluble environment through the exchange of soluble factors (a segregated coculture). A coculture and the multicellular biological feedback loop it represents are necessary to reproduce many complex in vivo effects, which is not surprising given the many interacting physiological systems that combine to result in complex human biology.
Coculture provides yet another opportunity to engineer greater relevance into an in vitro model. As previously described, stem cell differentiation pathways are one of the best known multiple cell type interactions, whereby the differentiation of stem cell “A” is directed (or suppressed) by the presence of soluble factors from cell “B.” Rivaling and perhaps surpassing stem cell cocultures for scientific activity are cancer cocultures, particularly cancer-stroma cocultures: it is increasingly being demonstrated that the incorporation of a second cell type materially affects cancer cells in culture (Khodarev et al. 2003) and boosts their relevance to the in vivo pathology (Chung et al. 2005; Mahadevan and Von Hoff 2007).
LAYERING COMPLEXITY
Incorporation of any of these themes—3D matrix microenvironment, actively stabilized soluble environment, mixed and segregated cocultures—in an in vitro
system represents an increase in complexity compared to standard 2D cell culture. Increased complexity is often associated with increased cost and time and decreased efficiency (often measured by throughput). These negative consequences are perhaps the largest reason these promising innovations have not achieved the wide use and rapid commercial uptake initially expected. Yet, in the absence of their purposed integration into drug delivery processes, correlation of in vitro models to in vivo results is poor, 88% of drug candidates fail in clinical trials, and each successful drug costs approximately $1 billion to develop and launch (Deloitte 2011).
There are nonetheless strong indications of progress toward a new era of in vitro models. With biologically derived gel matrices having led the way, there are now many commercially available scaffolds specifically marketed for 3D cell culture. Commonly used in multiwell plates to maintain throughput (but simultaneously limited by their static format), these scaffolds are primed for layering the additional complexities of a stabilized, actively perfused soluble environment and for the clever use of coculture, potentially with multiple matrices matched to cell type.
Basic segregated coculture has become widespread through the use of inserts fitted into the wells of multiwell plates and more recently through mixed but spatially controlled cocultures made possible by 2D microfabrication techniques. Limitations of the first iterations of these innovations include the static nature of multiwell plate culture and (for inserts) a limited range of materials suitable as membranes, but they have established important baselines that will be expanded with the integration of more and better 3D physical and soluble microenvironments.
Finally, early bioreactor systems have demonstrated the clear benefits of perfusion, but their adoption is hampered by high costs per experiment, a requirement for atypical cell culture equipment, and low throughput.
Microfluidic and Mesofluidic Approaches
These first examples of successfully integrating a single innovation theme that acceptably increases complexity (i.e., is worth the tradeoff) have laid the foundation for “layered complexity” approaches that may break new ground in adoption and use. In the United States, recent National Institutes of Health (NIH) and Defense Advanced Research Projects Agency (DARPA) grant solicitations themed around modeling 3 and 10 (respectively) interacting physiological systems were awarded to microfluidics “lab-on-a-chip” submissions (Figure 2, left).
The microfluidics approach embraces perfusion systems at a micrometer scale (the scale of the cells themselves), while layering the complexity of cocultures at various points in the fluidic channels. Benefits include a smaller footprint for the culture chamber device, reduced flow circuit volumes, and the use of micromanufacturing techniques for device manufacturing. This approach has most

FIGURE 2 Examples of microfluidics(left, printed with permission from the Wyss Institute) and mesofluidics(right, KIYATEC Inc.) “layered complexity” in vitro systems. Both are perfusion based and incorporate 3D microenvironments and coculture interactions, but they differ in scale, manufacturing techniques, potential applications for cell-scaffold constructs, cost, and throughput.
effectively been demonstrated when modeling flat biological barrier models (e.g., gut, lung luminal interfaces) where perfusion takes the form of laminar-type flow over a dense, flat, cell-membrane construct. Technical challenges can include the inability to load and recover scaffolds, “edge effects” of soft matrices, management of cell/matrix density over time in gel matrices, and successful maintenance of constant flow in channels with small dimensions.
Another approach that successfully achieves the desired layered complexity may be thought of as “mesofluidic,” with culture chamber dimensions on the scale of millimeters rather than micrometers (Figure 2, right). In contrast to microfluidics, this approach has focused on modeling tissues rather than barriers, and perfusion can take the form of interstitial-type flow through a 3D cell-scaffold construct.
The mesofluidic approach inherits the benefits of more traditional bioreactors, including the highest cell viability over time, best potential to model complexity, and broadest incorporation and recovery of diverse 3D scaffold materials, the latter being an important bridge to biomanufacturing applications such as regenerative medicine, tissue engineering, and some forms of cell therapy. Layered complexity is achieved through inherent accommodation of both mixed and segregated cocultures and more controlled management of the soluble environment through active perfusion. Although cost may be mitigated to the extent that these smaller bioreactor systems can leverage the existing cost structure for 2D cell culture processes,1 lower throughput in mesofluidic systems remains a tradeoff.
Impacts of Economic and Social Factors
Nontechnical factors are aligning with the emergence of layered-complexity technological approaches. The economic and political environments have changed such that there is an increased focus on the societal value derived from the expenditure of granting agencies’ (and ultimately the public’s) research monies. It is becoming less acceptable to fund or conduct research that can be demonstrated to have a low, or zero, correlation to the biology being modeled when alternatives with higher correlation exist, even though they are more complex. Funding agencies are increasingly supportive of initiatives that mandate the incorporation of layered complexities.
Contractions in the global pharmaceutical industry have resulted in emphasis on new approaches that both drive down development costs and point toward new understanding of complex biology (and new targets, mechanisms, and pathways). Successful regenerative medicine and cell therapy business models have emerged, heightening the demand for improved in vitro manufacturing and quality control processes compliant with Current Good Manufacturing Processes (cGMP). And
__________
1This cost structure encompasses the costs of commoditized supplies, equipment, instruments, and general infrastructure of traditional cell culture methods.
finally, increasing societal interest (particularly in the European Union) is also driving broader and faster adoption of more complex in vitro models and techniques that show promise for refining, reducing, and replacing the use of animals.
CONCLUSIONS
Perception of the value of in vitro models is slowly changing to both embrace the need for more relevance and accept the tradeoffs of lower throughput and increased complexity (Table 1). This change is driven by multiple, related dynamics: (1) scientific literature demonstrating the increased relevance of more complex (e.g., 3D, perfused, coculture) cell cultures to in vivo biology, especially that of humans, compared to the low relevance of 2D monolayer cultures; (2) the increasing adoption of approaches incorporating single-factor complexity (e.g., 3D environment only), albeit for a limited number of applications; (3) the emergence of “layered complexity”–type approaches whose combined dynamics have begun to enable the modeling of organism-level interactions, with potentially broad application; (4) the unfavorable failure rate and high costs of clinical trials for the pharmaceutical industry, especially given the dearth of new blockbuster drugs; (5) the emergence of viable business models in related industries (regen-
TABLE 1 Evolving Paradigm Through Which the Value of In Vitro Models Is Perceived
| Traditional Paradigm | New Paradigm | ||
| 2D static monolayer cell culture | More complex cell culture: 3D or perfusion or coculture | 2D static monolayer cell culture | Layered complexity cell culture (e.g., 3D perfused coculture) |
|
• High throughput |
• Lower throughput |
• Higher throughput but lower relevance |
• Higher relevance but lower throughput |
|
• Acute cost minimization |
• Costs more than 2D |
• Cost and value of data both matter |
• Overall cost reduction potential |
|
• Synch with past data |
• Past data disconnect |
• Oversimplified |
• Managed complexity |
|
• Convenience |
• Interesting but impractical |
• Use when can get away with |
• Use when value justifies cost |
| NET EFFECT: Very heavy reliance on 2D static monolayer methods with emphasis on throughput and acute cost minimization. | NET EFFECT: Balanced approach that recognizes throughput/relevance tradeoffs and integrates both options. Ultimately reduces overall costs by decreasing late-stage failures (drugs) and/or increasing performance (cell therapy, regenerative medicine). | ||
erative medicine, cell therapy) that require and can coopt cell culture innovation for cell maturation and processing; and (6) aligned nontechnical trends, including increased emphasis on funding clearly relevant research and on further refining, reducing, and replacing the use of animals.
The combination of these factors results in unprecedented opportunity and provides the required foundation to usher in a new era of better in vitro models. As they are implemented, these models will significantly advance understanding of human physiology while simultaneously translating to substantial health and cost benefits.
FINANCIAL DISCLOSURE
Dr. Gevaert is the CEO of, and owns stock in, KIYATEC Inc., a company focused on in vitro models with higher correlation to in vivo results via convenient and cost-effective perfused 3D cell-based assays.
REFERENCES
Birgersdotter A, Sandberg R, Ernberg I. 2005. Gene expression perturbation in vitro: A growing case for three-dimensional (3D) culture systems. Seminars in Cancer Biology 15:405–412.
Chung LWK, Baseman A, Assikis V, Zhau HE. 2005. Molecular insights into prostate cancer progression: The missing link of tumor microenvironment. Journal of Urology 173:10–20.
Cimetta E, Flaibani M, Mella M, Serena E, Boldrin L, De Coppi P, Elvassore N. 2007. Enhancement of viability of muscle precursor cells on 3D scaffold in a perfusion bioreactor. International Journal of Artificial Organs 30:415–428.
Deloitte. 2011. Measuring the return from innovation: Is R&D earning its investment? Deloitte Annual Review of How the Life Sciences Industry Is Performing in Generating Value from R&D:5.
Dhurjati R, Liu X, Gay CV, Mastro AM, Vogler EA. 2006. Extended-term culture of bone cells in a compartmentalized bioreactor. Tissue Engineering 12:3045–3054.
Fraley SI, Feng Y, Wirtz D, Longmore GD. 2011. Reply: Reducing background fluorescence reveals adhesions in 3D matrices. Nature Cell Biology 13(1):5–7.
Goldstein AS, Juarez TM, Helmke CD, Gustin MC, Mikos AG. 2001. Effect of convection on osteoblastic cell growth and function in biodegradable polymer foam scaffolds. Biomaterials 22:1279–1288.
Gomes ME, Sikavitsas VI, Behravesh E, Reis RL, Mikos AG. 2003. Effect of flow perfusion on the osteogenic differentiation of bone marrow stromal cells cultured on starch-based three-dimensional scaffolds. Journal of Biomedical Materials Research Part A 67A:87–95.
Indolfi L, Baker AB, Edelman ER. 2012. The role of scaffold microarchitecture in engineering endo-thelial cell immunomodulation. Biomaterials 33:7019–7027.
Khodarev NN, Yu JQ, Labay E, Darga T, Brown CK, Mauceri HJ, Yassari R, Gupta N, Weichselbaum RR. 2003. Tumour-endothelium interactions in co-culture: Coordinated changes of gene expression profiles and phenotypic properties of endothelial cells. Journal of Cell Science 116:1013–1022.
Kilian KA, Bugarija B, Lahn BT, Mrksich M. 2010. Geometric cues for directing the differentiation of mesenchymal stem cells. Proceedings of the National Academy of Sciences of the United States of America 107:4872–4877.
Kumar G, Waters MS, Farooque TM, Young MF, Simon CG Jr. 2012. Freeform fabricated scaffolds with roughened struts that enhance both stem cell proliferation and differentiation by controlling cell shape. Biomaterials 33:4022–4030.
Mahadevan D, Von Hoff DD. 2007. Tumor-stroma interactions in pancreatic ductal adenocarcinoma. Molecular Cancer Therapeutics 6:1186–1197.
Martin KJ, Patrick DR, Bissell MJ, Fournier MV. 2008. Prognostic breast cancer signature identified from 3D culture model accurately predicts clinical outcome across independent datasets. PLoS ONE 3(8):e2994.
Mercille S, Massie B. 1999. Apoptosis-resistant E1B-19K-expressing NS/0 myeloma cells exhibit increased viability and chimeric antibody productivity under perfusion culture conditions. Biotechnology and Bioengineering 63:529–543.
Nikkhah M, Edalat F, Manoucheri S, Khademhosseini A. 2012. Engineering microscale topographies to control the cell-substrate interface. Biomaterials 33:5230–5246.
Paul SM, Mytelka DS, Dunwiddie CT, Persinger CC, Munos BH, Lindborg SR, Schacht AL. 2010. How to improve R&D productivity: The pharmaceutical industry’s grand challenge. Nature Reviews Drug Discovery 9:203–214.
Petri RJ. 1887. Eine kleine modification des koch’schen plattenverfahrens. Centralblatt für Bacteriologie und Parasitenkunde 1:279–280.
Ridky TW, Chow JM, Wong DJ, Khavari PA. 2010. Invasive three-dimensional organotypic neoplasia from multiple normal human epithelia. Nature Medicine 16:1450–1455.
Tomei AA, Siegert S, Britschgi MR, Luther SA, Swartz MA. 2009. Fluid flow regulates stromal cell organization and CCL21 expression in a tissue-engineered lymph node microenvironment. Journal of Immunology 183:4273–4283.
Wu M-H, Kuo C-Y. 2011. Application of high throughput perfusion micro 3-D cell culture platform for the precise study of cellular responses to extracellular conditions: Effect of serum concentrations on the physiology of articular chondrocytes. Biomedical Microdevices 13:131–141.
Wu M-H, Urban JPG, Cui ZF, Cui Z, Xu X. 2007. Effect of extracellular pH on matrix synthesis by chondrocytes in 3D agarose gel. Biotechnology Progress 23:430–434.
Zschenker O, Streichert T, Hehlgans S, Cordes N. 2012. Genome-wide gene expression analysis in cancer cells reveals 3D growth to affect ECM and processes associated with cell adhesion but not DNA repair. PLoS ONE 7:e34279.












