
Vijay-Kumar, M., Sanders, C.J., Taylor, R.T., Kumar, A., Aitken, J.D., Sitaraman, S.V., Neish, A.S., Uematsu, S., Akira, S., Williams, I.R., and Gewirtz, A.T. (2007). Deletion of TLR5 results in spontaneous colitis in mice. J. Clin. Invest. 117, 3909–3921.
Vijay-Kumar, M., Aitken, J.D., Carvalho, F.A., Cullender, T.C., Mwangi, S., Srinivasan, S., Sitaraman, S.V., Knight, R., Ley, R.E., and Gewirtz, A.T. (2010). Metabolic syndrome and altered gut microbiota in mice lacking Toll-like receptor 5. Science 328, 228–231.
Wong, J.M.W., de Souza, R., Kendall, C.W.C., Emam, A., and Jenkins, D.J.A. (2006). Colonic health: fermentation and short chain fatty acids. J. Clin. Gastroenterol. 40, 235–243.
Wright, J., Kennedy-Stephenson, J., Wang, C., McDowell, M., and Johnson, C. (2004). Trends in intake of energy and macronutrients—United States, 1971–2000. JAMA 291, 1193–1194.
Yatsunenko, T., Rey, F.E., Manary, M.J., Trehan, I., Dominguez-Bello, M.G., Contreras, M., Magris, M., Hidalgo, G., Baldassano, R.N., Anokhin, A.P., et al. (2012). Human gut microbiome viewed across age and geography. Nature 486, 222–227.
A4
MICROBIAL EXPOSURE DURING EARLY LIFE HAS PERSISTENT EFFECTS ON NATURAL KILLER T CELL FUNCTION9
Torsten Olszak,10,*Dingding An,11,*Sebastian Zeissig,12Miguel Penilla Vera,13Julia Richter,14Andre Franke,15Jonathan N. Glickman,16Reiner Siebert,14Rebecca M. Barron,13Dennis L. Kasper,11,†‡and Richard S. Blumberg10,†‡
Abstract
Exposure to microbes during early childhood is associated with protection from immune-mediated diseases such as inflammatory bowel disease (IBD) and asthma. Here, we show that in germ-free (GF) mice, invariant natural killer T (iNKT) cells accumulate in the colonic lamina propria and
________________
9 From “Microbial exposure during early life has persistent effects on natural killer T cell function,” Science 336(6080):489-493, 2012. Reprinted with permission from the AAAS.
10 Division of Gastroenterology, Hepatology, and Endoscopy, Brigham and Women’s Hospital, Harvard Medical School, Boston, MA 02115, USA.
11 Channing Laboratory, Brigham and Women’s Hospital and Department of Microbiology and Immunobiology, Harvard Medical School, Boston, MA 02115, USA.
12 Department of Internal Medicine I, University Medical Center Schleswig-Holstein, Kiel 24105, Germany.
13 Division of Pulmonary and Critical Care Medicine, Brigham and Women’s Hospital, Harvard Medical School, Boston, MA 02115, USA.
14 Institute of Human Genetics, Christian-Albrechts University Kiel and University Medical Center Schleswig-Holstein, Kiel 24105, Germany.
15 Institute of Clinical Molecular Biology, Christian-Albrechts University, Kiel 24105, Germany.
16 GI Pathology, Caris Diagnostics, Caris Life Sciences, Newton, MA 02464, USA.
* These authors contributed equally to this work.
† These authors contributed equally to this work.
‡ To whom correspondence should be addressed. E-mail: rblumberg@partners.org (R.S.B.); dennis_kasper@hms.harvard.edu (D.L.K.).
lung, resulting in increased morbidity in models of IBD and allergic asthma as compared with that of specific pathogen-free mice. This was associated with increased intestinal and pulmonary expression of the chemokine ligand CXCL16, which was associated with increased mucosal iNKT cells. Colonization of neonatal—but not adult—GF mice with a conventional microbiota protected the animals from mucosal iNKT accumulation and related pathology. These results indicate that age-sensitive contact with commensal microbes is critical for establishing mucosal iNKT cell tolerance to later environmental exposures.
The mammalian host immune system is broadly stimulated with the first exposure to microorganisms during neonatal life (Kelly et al., 2007). The inner surfaces of the gastrointestinal tract and lungs are particularly affected because they are predominant sites of microbial contact (Renz et al., 2012). Epidemiologic observations further suggest that the immune effects of early-life microbial exposure are durable and persist into later life because they can be associated with prevention of diseases such as inflammatory bowel disease (IBD) and asthma (Ege et al., 2011; López-Serrano et al., 2011; von Mutius, 2007).
Invariant natural killer T (iNKT) cells probably play an important role in the pathogenesis of ulcerative colitis (UC)—a major form of IBD—and in asthma (Akbari et al., 2003; Fuss et al., 2004). Such cells recognize endogenous and exogenous lipid antigens presented by the nonpolymorphic major histocompatibility complex (MHC) class I–like protein CD1d and secrete abundant amounts of proinflammatory cytokines such as interleukin-4 (IL-4) and IL-13 upon activation (Cohen et al., 2009; Kronenberg, 2005). We therefore investigated age-dependent regulation of iNKT cells by use of microbes in mouse models of IBD and asthma.
We first examined the appearance of iNKT cells in tissues of 8-week-old germ-free (GF) and specific pathogen-free (SPF) Swiss-Webster (SW) mice. Relative and absolute numbers of iNKT cells were increased in GF mice in colonic lamina propria (LP) (Figure A4-1, A to C). These differences in colonic iNKT cell numbers between GF and SPF mice were detectable after weaning and stable for life, suggesting early and persistent effects of the microbiota (Figure A4-1D). iNKT cells were not increased in the ileal LP (ileum) of GF mice, and the liver, spleen, and thymus contained even fewer iNKT cells under GF relative to SPF conditions (fig. S1), which is consistent with a recent report (Wei et al., 2010). GF C57BL/6 mice (B6) exhibited similar increases of iNKT cells in the colonic LP as well as the liver, in contrast to GF SW mice (fig. S2). Although increased in number, the iNKT cell expression of several activation and memory markers was unaltered in GF SW, relative to SPF, mice (fig. S3).
To examine the relevance of these findings, we investigated the susceptibility of GF and SPF mice to oxazolone-induced colitis, a model of intestinal inflammation that possesses features of UC and is dependent on IL-13 production by
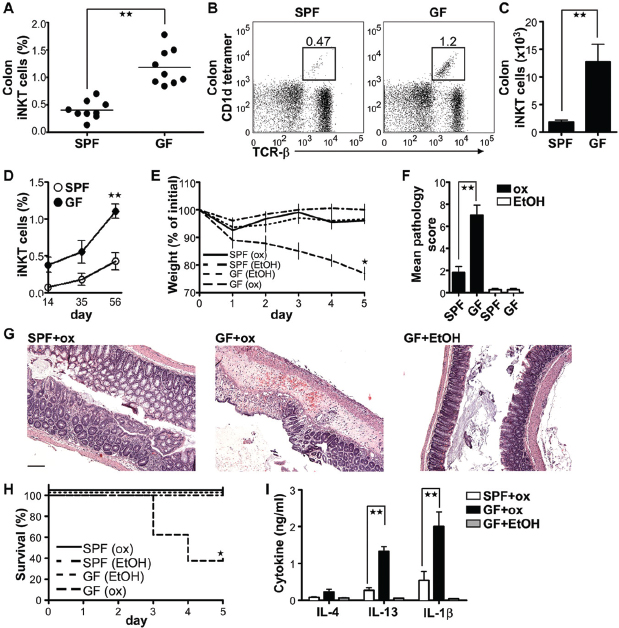
FIGURE A4-1 Intestinal bacteria-dependent accumulation of colonic iNKT cells in GF mice leads to high mortality in oxazolone-induced colitis. (A to C) The percentage of CD1d tetramer–positive cells (iNKT cells) within the live lymphocytes from the lamina propria (LP) of gender-matched GF and SPF SW mice (7 to 8 weeks old) was analyzed by means of flow cytometry (A). Representative dot-plots are shown in (B), and the absolute number of iNKT cells is shown in (C). Each circle in (A) represents an individual mouse. (D) Percentages of colonic iNKT cells of live LP lymphocytes in gender-matched SPF and GF mice were analyzed at different ages by means of flow cytometry (n = 4 mice per group). (E to H) Eight-week-old GF and SPF mice were monitored and scored after rectal challenge with 1% oxazolone (ox) or 50% ethanol for survival and body weight loss for 5 days. On day 5, the colons were collected and dissected for histological analysis (n = 5 mice per group). (G) Scale bar, 50 μm. (I) The concentration of IL-4, IL-13, and IL-1β in the supernatant of 24 hours–colon organ explant cultures determined by means of enzyme-linked immunosorbent assay (ELISA) on day five (n = 5 mice per group). All data were obtained from three independent experiments with similar results. In all panels, error bars represent the SD. *P < 0.05, **P < 0.01, unpaired t test and *P < 0.05, log-rank test in (H).
CD1d-restricted iNKT cells (Heller et al., 2002; Schiechl et al., 2011). Although GF or germ-reduced mice exhibit exacerbated inflammation in an innate mouse model of colitis (Maslowski et al., 2009; Rakoff-Nahoum et al., 2004), colitis is typically prevented under GF conditions in models dependent on an adaptive immune response (Strober et al., 2002). Surprisingly, GF mice were more sensitive to oxazolone-induced colitis, as revealed by severe weight loss, pathology, and a high mortality rate in contrast to SPF mice (Figure A4-1, E to H). This was not due to overexpression of cell-surface CD1d expression on intestinal epithelial (fig. S4A) and hematopoietic cells (fig. S4B). Consistent with previous studies (Heller et al., 2002), the colitis in GF mice was characterized by a marked increase in production of IL-13 and IL-1β in comparison with SPF mice (Figure A4-1I).
To confirm the CD1d-restriction of this colitis in GF mice, we investigated the effects of CD1d blockade. Adult (8- to 9-week-old) GF and SPF mice were treated with a blocking monoclonal antibody (Ab) specific for CD1d (19G11) or an isotype control Ab (Roark et al., 1998). Treatment of GF mice with 19G11, but not the isotype control, protected against colitis-induced mortality and associated pathology (Figure A4-2, A to C, and fig. S4, C and D). Furthermore, CD1d blockade of GF or SPF mice did not lead to significant functional changes in dendritic cells (Yue et al., 2005) or B cells (Mizoguchi and Bhan, 2006) as demonstrated by stable IL-12 or IL-10 production (fig. S4, E and F), respectively.
We next examined whether reestablishing microbiota in adult GF mice would normalize iNKT cell levels in the colon. Quite surprisingly, exposure of adult GF mice to SPF conditions (GF/a) did not restore iNKT cells in the colon to the levels observed in SPF mice or reverse the mortality and severe pathology after oxazolone administration (Figure A4-2, D to F). We therefore considered whether the ability to normalize iNKT cell levels and function in the colon was dependent on the age at which microbial contact occurred. Indeed, when we colonized pregnant GF female mice just before delivery and therefore exposed neonatal GF mice to SPF conditions on their first day of life (GF/n), we observed a complete normalization of iNKT cell levels that persisted even two months after colonization (Figure A4-2D). Consistent with this, GF/n exhibited reduced susceptibility to oxazolone-induced colitis (relative to that of GF mice) two months after colonization, with a degree of severity identical to that observed in SPF animals, as shown by an analysis of mortality, weight loss, pathology, and cytokine production (Figure A4-2, E and F, and fig. S5).
To verify that the early life events associated with the absence of normal microbial colonization on iNKT cell homeostasis were CD1d-dependent, we treated GF mice with 19G11 Ab for their first 6 weeks of life. 19G11, but not control Ab, treatment blocked accumulation of iNKT cells in all tissues examined and susceptibility to oxazolone-induced colitis in later life (fig. S6). Similar to GF mice, SPF mice (B6) born under germ-reduced conditions exhibited an increase in colonic iNKT cells and excessive oxazolone-induced colitis at 4 weeks of life
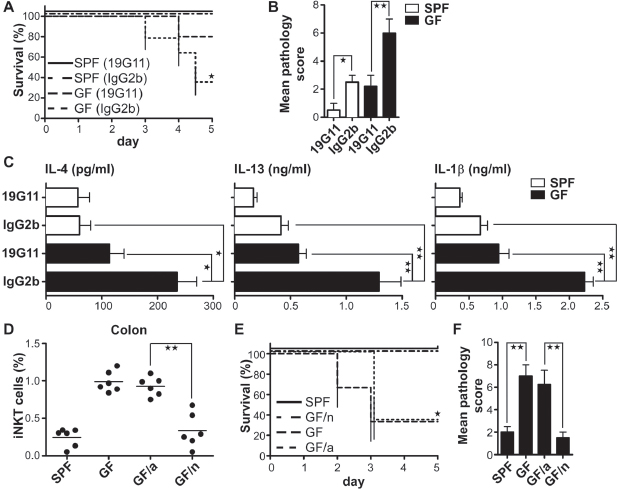
FIGURE A4-2 Microbial colonization during early life prevents the CD1d/iNKT cell-dependent high mortality in oxazolone colitis in GF mice. (A to C) Eight-week-old GF and SPF mice were treated once with 1 mg of 19G11 or isotype control IgG2b antibody before presensitization and rectal oxazolone challenge. (A) Survival after oxazalone challenge is shown. (B) On day 5, the colons were collected for histological analysis, and (C) the cytokine concentration in the supernatant of 24 hours-colon organ explant cultures was measured with ELISA (n ≥ 4 mice per group). (D) Colonic LP lymphocytes were analyzed for iNKT cells by flow cytometry. (GF/a) mice were GF mice that were exposed to SPF environmental conditions at the age of 5 weeks and maintained for 4 more weeks. (GF/n) mice were pups exposed to SPF conditions on their first day of life and maintained under SPF environmental conditions for 8 to 9 weeks. Each circle represents a mouse. (E to F) Colonized mice were treated as above with oxazolone (n = 5 mice per group). All data were obtained from more than two independent experiments with similar results. Error bars indicate SD. *P < 0.05, **P < 0.01, unpaired t test and *P < 0.05, log-rank test in (A) and (E).
(fig. S7). As predicted (Heller et al., 2002), Cd1d-/- and Jα18-/- mice, both of which lack iNKT cells, were not susceptible to oxazolone-induced colitis either under SPF or germ-reduced conditions (fig. S7).
Animal models of asthma demonstrate an important role for iNKT cells (Meyer et al., 2008; Wingender et al., 2011), although the contribution to human
disease remains controversial (Iwamura and Nakayama, 2010). We found that the lungs of GF SW and B6 mice contained significantly higher relative and absolute numbers of iNKT cells in comparison with that of the lungs of the respective SPF mice (Figure A4-3, A to C, and fig. S8). Because commensal microbes may affect the induction of experimental asthma (Herbst et al., 2011), we tested an ovalbumin (OVA)–driven allergic-asthma mouse model in GF mice. We observed that similar to the colon, GF mice developed an allergic airway response to OVA significantly greater than that observed in SPF mice, as demonstrated by increases in airway resistance, total bronchial alveolar lavage fluid (BALF) cell numbers, BALF eosinophilia, serum immunoglobulin E (IgE) levels, proinflammatory cytokine production in the BALF, and lung tissue eosinophil infiltration (Figure A4- 3, D to F, and fig. S9). The asthma in GF mice was CD1d- and antigen-dependent because elimination of the asthmatic response was only observed with 19G11, but not control, Ab treatment (Figure A4-3, D to F, and fig. S9) of OVA-sensitized mice but not mice exposed to phosphate-buffered saline (PBS), which did not induce allergic asthma (fig. S10).
We therefore next investigated whether early-life exposure to a conventional microbiota also protects animals from CD1d-restricted inflammation in the lungs. As observed in the intestinal mucosa, neonatal (GF/n)—but not adult-life (GF/a)—exposure of GF mice to a conventional microbiota abrogated the increased accumulation of iNKT cells in the lungs and protected adult mice in the allergic asthma model from the pathology (Figure A4-3, G to I, and fig. S11).
The chemokine receptor CXCR6 on iNKT cells (Germanov et al., 2008) and its ligand CXCL16, which is expressed at high levels by human epithelial cells (Day et al., 2009) and increased in inflammation (Diegelmann et al., 2010), plays an important role in iNKT cell homeostasis. Therefore, we examined the serum of GF and GF/a mice for the presence of CXCL16 and observed significantly higher levels than that observed in SPF and GF/n mice (Figure A4-4A). This was due to significant increases of Cxcl16 mRNA expression levels in the colon and lung—but not ileum, liver, and thymus—of GF and GF/a relative to SPF and GF/n mice (Figure A4-4B and fig. S12A), and mainly immunolocalized to the epithelium (fig. S12B). Although Cxcl16 mRNA levels in the colon, ileum, and lung were similar immediately after birth in the GF and SPF animals at the beginning of colonization, they increased significantly and specifically only in the colon and lungs of the GF mice during later life (Figure A4-4C). These results suggest that microbial exposure provides signals that determine Cxcl16 gene expression in specific tissues. GF and SPF mice, however, exhibited similar levels of Cxcr6 mRNA expression in tissues and CXCR6 expression on the cell surface of iNKT cells (fig. S12, C and D).
CXCL16 was responsible for the accumulation of iNKT cells because treatment of newborn SPF and GF mice with a CXCL16-neutralizing Ab caused a decrease in iNKT cells in the colon and lung (Figure A4-4D) but had no effect on iNKT cell levels in the ileum or thymus (fig. S13A). However, it was associated
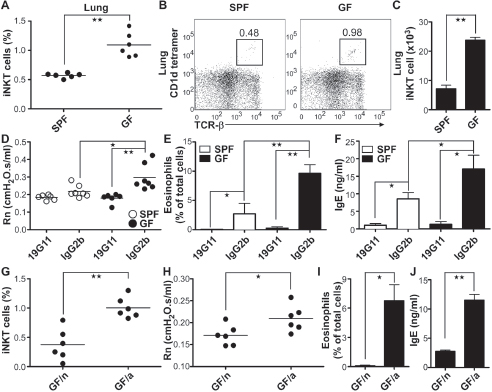
FIGURE A4-3 The increased CD1d/iNKT cell-mediated allergic response sensitivity of GF mice is dependent on age of colonization. (A to C) Lymphocytes from lungs of 8-week-old mice in each group were analyzed for iNKT cells by means of flow cytometry (A). Representative dot-plot is shown in (B), and the absolute number of iNKT cells is shown in (C) (n = 6 mice per group). (D to F) Age-matched mice from each group were treated once before OVA presensitization with 1 mg of 19G11 or IgG2b isotype control antibody and with 0.5 mg of 19G11 or IgG2b antibody before the first and the third serial OVA exposure. Twenty-four hours after the last challenge with 5% aerosolized OVA, the mice were analyzed for (D) airway resistance (Rn), (E) percentage of eosinophils of total BALF cells, and (F) IgE (n = 4 mice per group). (G) Lung lymphocytes were analyzed for iNKT cells by means of flow cytometry of age- and gender-matched GF/n and GF/a mice. (H to J) Mice were presensitized with OVA and analyzed after the last serial OVA challenge as described above (n ≥ 4 mice per group). Each circle in (A), (D), (G), and (H) represents an individual mouse. All data were obtained from more than two independent experiments with similar results. Error bars indicate the SD. *P ≤ 0.05, **P < 0.01, unpaired t test.
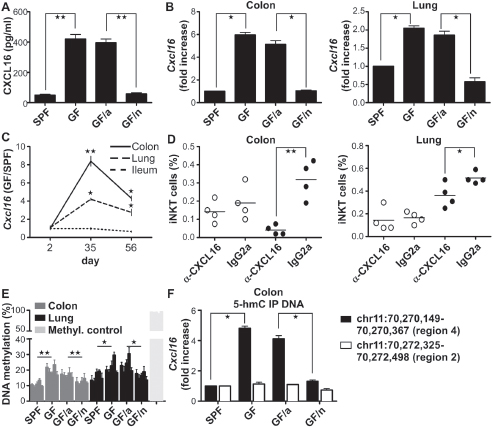
FIGURE A4-4 Microbiota affects tissue specific iNKT cell accumulation by genetic modifications of Cxcl16. (A) The CXCL16 concentration in the serum of each group was measured with ELISA in SPF, GF, GF/a, and GF/n as described above (n = 4 mice per group). (B) Colon and lung tissue samples were harvested from age-matched mice from each group and analyzed for Cxl16expression (n = 4 mice per group). (C) Colon, ileum, and lung tissue samples were analyzed from age-matched SPF and GF mice at different time points for Cxcl16 expression (n = 5 mice per group). (D) SPF (open circles) and GF (closed circles) newborn mice were treated three times a week intraperitoneal with 25 μg of a neutralizing CXCL16 antibody (αCXCL16) or its isotype control IgG2a antibody and analyzed at the age of 2 weeks for iNKT cell percentages by means of flow cytometry. Each circle represents a mouse. (E) Analysis of DNA methylation of five CpG sites of Cxcl16 by bisulfite pyrosequencing. For each group, the mean of DNA methylation of 5 CpG sites is shown as a percentage according to the methylated control (Methyl. control) (n ≥ 3 mice per group). (F) Cxcl16 qPCR analysis of colonic DNA after performing a 5-hydroxymethylated DNA immunoprecipitation (5-hmC IP DNA) (n ≥ 3 mice per group). All data were obtained from at least two independent experiments with similar results. Error bars indicate the SD. *P ≤ 0.05, **P < 0.01, unpaired t test.
with an increased accumulation of iNKT cells in the liver (fig. S13A), suggesting CXCL16 blockade caused a redirection of iNKT cells to this organ. Consequently, Ab-to-CXCL16– but not isotype-treated GF mice were protected from oxazolone colitis–induced mortality and pathology (fig. S13, B to D).
We next examined the epigenetic content of the Cxcl16 gene of GF and SPF mice. A region 5′ of the Cxcl16gene of SW mice that contains five potential CpG sites was determined by means of bisulfite pyrosequencing to be hypermethylated in the colon and lungs (Figure A4-4E)—but not in other tissues such as the spleen and liver (fig. S14)—under GF conditions as compared with SPF conditions. Colonization of neonatal —but not adult—GF mice with a conventional microbiota decreased the hypermethylation of the Cxcl16 gene to SPF levels (Figure A4-4E). Gene activation due to hypermethylation may occur as a consequence of specific types of environmental exposure (Rishi et al., 2010). To confirm this, we treated SPF neonates on their first day of life by oral gavage with high doses of folinic acid as previously described (Pu) so as to force Cxcl16 methylation. Compared with PBS-treated control mice, folinic acid administration resulted in elevated methylation of the Cxcl16 gene in the colon, ileum, and lung; at least a three- to fourfold increase of Cxcl16 mRNA expression in the same tissues; increased CXCL16 serum levels; and accumulation of iNKT cells in these tissues as observed in GF mice (fig. S15). Alternatively, we investigated whether elimination of Cxcl16 methylation in GF mice would reverse the elevation of CXCL16 expression by treating GF newborn littermates with the DNA methyltransferase inhibitor 5-Azacytidine (5-Aza). When examined at 2 weeks of life, 5-Aza treatment inhibited methylation and mRNA expression of Cxcl16 in the colon, ileum, and lung (fig. S16, A and B), diminished the levels of CXCL16 in the serum, and reduced iNKT cells in these tissues (fig. S16, C and D). 5-Aza treatment had no effect on other cell populations such as CD45+CD11c+ double positive cells (fig. S17).
Epigenetic marks provided by 5-hydroxymethylcytosine (5-hmC) incorporation into DNA have been suggested to be different from those provided by 5-methylcytosine (5-mC) and a distinct signature for elevated levels of transcription (Song et al., 2011). We therefore performed DNA immunoprecipitation with a monoclonal 5-hmC Ab followed by quantitative polymerase chain reaction (PCR) analysis for Cxcl16 in seven different regions of Cxcl16 in DNA samples obtained from the colons of SPF, GF, GF/a, and GF/n mice. We observed that Cxcl16 was highly increased in three out of the seven investigated regions (with region 4 as a representative) within the 5-hmC–modified DNA of GF and GF/a mice but not in the DNA acquired from SPF and GF/n mice (Figure A4-4F). No differences were observed in the other four regions (with region 2 as a representative) and in the DNA samples obtained from the ileum demonstrating the colonic specificity for 5-hmC modification of Cxcl16 (fig. S18). A map summarizing this information is provided in fig. S19.
Our studies indicate that CXCL16 is an age- and organ-dependent microbially regulated factor that modulates the quantities and function of iNKT cells in the colon and lungs and, consequently, susceptibility to tissue inflammation. The exact mechanism by which the microbiota regulates CXCL16 expression and thus iNKT cell accumulation in these organs is unknown, although it is independent of the Toll-like receptor adapter protein MyD88 (fig. S20). These observations are in accordance with previous epidemiological studies collectively known as the “hygiene hypothesis,” which proposes that early-life exposure to specific microbe-enriched environments decreases susceptibility to diseases such as IBD and asthma (Ege et al., 2011; Neish, 2009; Strachan, 1989; von Mutius, 2007), whereas its absence, as in antibiotic treatment during childhood, may have the opposite effect (Goksör et al., 2011; Shaw et al., 2010). Our results suggest that CD1d-restricted iNKT cell pathways and their relationship to microbes play a central role in these aforementioned processes. Early-life microbial exposure elicits long-lasting effects on iNKT cells, and in their absence, later-life exposure to factors that stimulate these cells may induce an autoinflammatory response. These findings are predicted to extrapolate to humans, given the extensive similarities between the mouse and human CD1d and iNKT cell systems (Sullivan and Kronenberg, 2007).
Acknowledgements
We thank E. Troy and S. Edwards for handling of the GF mice, J. Cusick for technical assistance, E. Nieuwenhuis and K. Schneeberger for assistance with epigenetics studies, A. Bellacosa and K. Baker for helpful discussions and manuscript preparation, and the KinderKrebsInitiative Buchholz/Holm-Seppensen for providing pyrosequencing infrastructure. The work was supported by NIH grants DK44319, DK51362, DK53056, and DK88199 (R.S.B.) and AI090102 (D.L.K.); Crohn’s Colitis Foundation of America Senior Research Award (D.L.K.) and Crohn’s and Colitis Foundation of America Postdoctoral Fellowship Award (D.A. and S.Z.); the Harvard Digestive Diseases Center (DK034854); the Medizinausschuss Schleswig-Holstein, German Ministry of Education Research through the National Genome Research Network (A.F.); The Medical Faculty, Kiel (R.S.) and the Deutsche Forschungsgemeinschaft (DFG) (OL 324/1-1, SZ 814/1-1, 814/4-1); as well as DFG Excellence Cluster “Inflammation at Interfaces” (A.F. and S.Z.). The data reported in this manuscript are tabulated in the main paper and in the supplementary materials.
References
D. Kelly, T. King, R. Aminov, Importance of microbial colonization of the gut in early life to the development of immunity. Mutat. Res. 622, 58 (2007).
H. Renz et al., The impact of perinatal immune development on mucosal homeostasis and chronic inflammation. Nat. Rev. Immunol. 12, 9 (2012).
M. J. Ege, et al., GABRIELA Transregio 22 Study Group, Exposure to environmental microorganisms and childhood asthma. N. Engl. J. Med. 364, 701 (2011).
P. López-Serrano et al., Environmental risk factors in inflammatory bowel diseases. Investigating the hygiene hypothesis: A Spanish case-control study. Scand. J. Gastroenterol. 45, 1464 (2010).
E. von Mutius, Allergies, infections and the hygiene hypothesis—The epidemiological evidence. Immunobiology 212, 433 (2007).
O. Akbariet al., Essential role of NKT cells producing IL-4 and IL-13 in the development of allergen-induced airway hyperreactivity. Nat. Med. 9, 582 (2003).
I. J. Fusset al., Nonclassical CD1d-restricted NK T cells that produce IL-13 characterize an atypical Th2 response in ulcerative colitis. J. Clin. Invest. 113, 1490 (2004).
N. R. Cohen, S. Garg, M. B. Brenner, Antigen Presentation by CD1 Lipids, T Cells, and NKT Cells in Microbial Immunity. Adv. Immunol. 102, 1 (2009).
M. Kronenberg, Toward an understanding of NKT cell biology: Progress and paradoxes. Annu. Rev. Immunol. 26, 877 (2005).
B. Wei et al., Commensal microbiota and CD8+ T cells shape the formation of invariant NKT cells. J. Immunol. 184, 1218 (2010).
F. Heller, I. J. Fuss, E. E. Nieuwenhuis, R. S. Blumberg, W. Strober, Oxazolone colitis, a Th2 colitis model resembling ulcerative colitis, is mediated by IL-13-producing NK-T cells. Immunity 17, 629 (2002).
G. Schiechl et al., Tumor development in murine ulcerative colitis depends on MyD88 signaling of colonic F4/80+CD11b(high)Gr1(low) macrophages. J. Clin. Invest. 121, 1692 (2011).
K. M. Maslowskiet al., Regulation of inflammatory responses by gut microbiota and chemoattractant receptor GPR43. Nature 461, 1282 (2009).
S. Rakoff-Nahoum, J. Paglino, F. Eslami-Varzaneh, S. Edberg, R. Medzhitov, Recognition of commensal microflora by toll-like receptors is required for intestinal homeostasis. Cell 118, 229 (2004).
W. Strober, I. J. Fuss, R. S. Blumberg, The immunology of mucosal models of inflammation. Annu. Rev. Immunol. 20, 495 (2002).
J. H. Roark et al., CD1.1 expression by mouse antigen-presenting cells and marginal zone B cells. J. Immunol. 160, 3121 (1998).
S. C. Yue, A. Shaulov, R. Wang, S. P. Balk, M. A. Exley, CD1d ligation on human monocytes directly signals rapid NF-κB activation and production of bioactive IL-12. Proc. Natl. Acad. Sci. U.S.A. 102, 11811 (2005).
A. Mizoguchi, A. K. Bhan, A case for regulatory B cells. J. Immunol. 176, 705 (2006).
E. H. Meyer, R. H. DeKruyff, D. T. Umetsu, T cells and NKT cells in the pathogenesis of asthma. Annu. Rev. Med. 59, 281 (2008).
G. Wingender et al., Invariant NKT cells are required for airway inflammation induced by environmental antigens. J. Exp. Med. 208, 1151 (2011).
C. Iwamura, T. Nakayama, Role of NKT cells in allergic asthma. Curr. Opin. Immunol. 22, 807 (2010).
T. Herbst et al., Dysregulation of allergic airway inflammation in the absence of microbial colonization. Am. J. Respir. Crit. Care Med. 184, 198 (2011).
E. Germanov et al., Critical role for the chemokine receptor CXCR6 in homeostasis and activation of CD1d-restricted NKT cells. J. Immunol. 181, 81 (2008).
C. Day, R. Patel, C. Guillen, A. J. Wardlaw, The chemokine CXCL16 is highly and constitutively expressed by human bronchial epithelial cells. Exp. Lung Res. 35, 272 (2009).
J. Diegelmann et al., Expression and regulation of the chemokine CXCL16 in Crohn’s disease and models of intestinal inflammation. Inflamm. Bowel Dis. 16, 1871 (2010).
V. Rishi et al., CpG methylation of half-CRE sequences creates C/EBPα binding sites that activate some tissue-specific genes. Proc. Natl. Acad. Sci. U.S.A. 107, 20311 (2010).
M. Pufulete et al., Effect of folic acid supplementation on genomic DNA methylation in patients with colorectal adenoma. Gut 54, 648 (2005).











