
C. X. Song et al., Selective chemical labeling reveals the genome-wide distribution of 5-hydroxymethylcytosine. Nat. Biotechnol. 29, 68 (2011).
A. S. Neish, Microbes in gastrointestinal health and disease. Gastroenterology 136, 65 (2009).
D. P. Strachan, Hay fever, hygiene, and household size. BMJ 299, 1259 (1989).
S. Y. Shaw et al., Am. J. Gastro. 105, 2687 (2010).
E. Goksör et al., Preschool wheeze—Impact of early fish introduction and neonatal antibiotics. Acta Paediatr. 100, 1561 (2011).
B. A. Sullivan, M. Kronenberg, TCR-mediated recognition of glycolipid CD1 complexes. Curr. Top. Microbiol. Immunol. 314, 165 (2007).
K. A. Sauer, P. Scholtes, R. Karwot, S. Finotto, Isolation of CD4+ T cells from murine lungs: A method to analyze ongoing immune responses in the lung. Nat. Protoc. 1, 2870 (2006).
J. L. Matsuda et al., Tracking the response of natural killer T cells to a glycolipid antigen using CD1d tetramers. J. Exp. Med. 192, 741 (2000).
R. M. Baronet al., Nitric oxide synthase-2 down-regulates surfactant protein-B expression and enhances endotoxin-induced lung injury in mice. FASEB J. 18, 1276 (2004).
B. Lamprecht et al., Derepression of an endogenous long terminal repeat activates the CSF1R protooncogene in human lymphoma. Nat. Med. 16, 571, 1p, 579 (2010).
A5
THE APPLICATION OF ECOLOGICAL THEORY TOWARD AN UNDERSTANDING OF THE HUMAN MICROBIOME17
Elizabeth K. Costello,18Keaton Stagaman,19Les Dethlefsen,18,20Brendan J. M. Bohannan,19David A. Relman18,20,21,*
Abstract
The human-microbial ecosystem plays a variety of important roles in human health and disease. Each person can be viewed as an island-like “patch” of habitat occupied by microbial assemblages formed by the fundamental processes of community ecology: dispersal, local diversification, environmental selection, and ecological drift. Community assembly theory, and metacommunity theory in particular, provides a framework for understanding the ecological dynamics of the human microbiome, such as compositional
________________
17 From “The application of ecological theory toward an understanding of the human microbiome,” Science 336(6086):1255-1262, 2012. Reprinted with Permission from the AAAS.
18 Department of Microbiology and Immunology, Stanford University School of Medicine, Stanford, CA 94305, USA.
19 Institute of Ecology and Evolution, University of Oregon, Eugene, OR 97403, USA.
20 Department of Medicine, Stanford University School of Medicine, Stanford, CA 94305, USA.
21 Veterans Affairs Palo Alto Health Care System, Palo Alto, CA 94304, USA.
* To whom correspondence should be addressed. E-mail: relman@stanford.edu
variability within and between hosts. We explore three core scenarios of human microbiome assembly: development in infants, representing assembly in previously unoccupied habitats; recovery from antibiotics, representing assembly after disturbance; and invasion by pathogens, representing assembly in the context of invasive species. Judicious application of ecological theory may lead to improved strategies for restoring and maintaining the microbiota and the crucial health-associated ecosystem services that it provides.
Each person is an assemblage of not only human cells but also many symbiotic species. The abundant and diverse microbial members of the assemblage play critical roles in the maintenance of human health by liberating nutrients and/or energy from otherwise inaccessible dietary substrates, promoting differentiation of host tissues, stimulating the immune system, and protecting the host from invasion by pathogens. A number of clinical disorders are associated with alterations in host-associated microbial communities (the “microbiota”), including obesity, malnutrition, and a variety of inflammatory diseases of the skin, mouth, and intestinal tract. Thus, the human body can be viewed as an ecosystem, and human health can be construed as a product of ecosystem services delivered in part by the microbiota.
There is growing interest in the use of theoretical methods to study microbial community ecology in general and host-associated microbiota in particular (Mihaljevic, 2012; Prosser et al., 2007). Recent discoveries of unexpected variation in the composition of the microbiome of healthy individuals (Palmer et al., 2007; Ravel et al., 2011; Wu et al., 2011) highlight the importance of identifying the processes that could possibly give rise to such variation. Ecological theory seeks to explain and predict observable phenomena, such as temporal and spatial patterns of diversity. Here, we explore how community assembly theory can be used to understand the human-associated microbiota and its role in health and disease.
Ecological Processes Within Humans
The essential building blocks of community assembly theory encompass the processes that create and shape diversity in local assemblages: dispersal, diversification, environmental selection, and ecological drift (Vellend, 2010). In addition, coevolution provides another lens through which to view the human-microbial ecosystem (Dethlefsen et al., 2007), although in this review we focus on shorter-term dynamics at the level of individual hosts.
Dispersal, or the movement of organisms across space, is a fundamental process by which diversity accumulates in local microbial communities. The concept put forth in the late 19th and early 20th centuries that “everything is everywhere, but the environment selects” had a powerful impact on thinking about community
assembly (O’Malley, 2007), but a more recent appreciation of microbial dispersal limitation suggests that this conceptualization was overly simplistic. Thinking in terms of dispersal leads to a view of the human body as an “island,” a patch of habitat that is continually sampling the pool of available colonists. The list of available colonists may be influenced by microbial traits—those affecting dispersal efficiency, transmission routes, and “ex-host” survivability—and by patterns of host contact and carriage, among other factors. Control of infectious disease transmission depends on accurate models of host-to-host microbial dispersal (Koopman, 2004), and these could guide investigations into the dissemination of the human microbiome. Selection favors efficient dispersal in pathogens, but perhaps less so among beneficial bacteria, because the host is harmed by the first and not by the second; for beneficial microbes, transmission routes such as direct or close contact may be more important. The density and spatial arrangement of host habitat patches has been highly dynamic throughout human history, as a result of human migration, urbanization, changes in family living structure, air travel, and other factors.
A second process operating in microbial communities is local diversification. Unlike in most plant and animal communities, this process can take place over short ecological time scales for microbes. Large microbial population sizes, high growth rates, and strong selective regimes, all of which can be found in the human body, result in rapid microbial adaptation via mutation or recombination. Recombination via horizontal gene transfer may be especially common among members of the human microbiota, especially those sharing the same ecological niche (e.g., body site) (Smillie et al., 2011). Microbial diversification may also be driven by interactions with phage in the human body. The processes of dispersal and diversification may interact (Urban et al., 2008); for example, immigration may suppress adaptive radiations (Fukami et al., 2007).
Relative abundances in local communities are shaped over time by a third process: environmental selection. When considering environmental selection, or niche-based interactions, the human body can be viewed in two ways. First, it can be viewed as a “habitat filter,” a collection of resources and conditions that allow the growth of some microbes but not others, resulting in the selection of microbial traits that permit survival and growth in the host. In this view, the host shapes the microbiota, but not the other way around. Body temperature is an example of such filtering, because microbes alter body temperature (causing fever) only when they transgress host anatomic boundaries. Second, the human body and its symbionts can be viewed as a community of interacting cells. This view differs from the habitat filter view in that it assumes strong feedbacks between hosts and microbes as well as among microbes. This view assumes that the host shapes the microbiota, and vice versa. Interactions between the host immune system and the microbiota might be best represented by this view (see Hooper et al., 2012). The overall patterns that arise from dispersal and environmental selection can vary as a function of the spatial scale over which these processes occur (Kerr et al., 2002).
In addition to selection-driven changes, species abundances may fluctuate as the result of a fourth process, known as ecological drift or demographic stochasticity. As a result of this process, low-abundance species (e.g., recent immigrants, strains nearly eliminated by antibiotics, or strains occupying niches with low carrying capacity) are more likely to proceed toward local extinction and become lost from the system, unless they have (or can gain) a competitive advantage, access a different niche, or become replenished by dispersal from outside the community. Thus, dispersal can effectively “rescue” species from the brink of local extinction.
Finally, the human habitat can be understood as a host-symbiont “holobiont”— an ecological system under selection to minimize conflict between individual members. This view emphasizes a dominant role of coevolution in the assembly and dynamics of the human ecosystem and reminds us that long- and short-term selective pressures on the human microbiota are not necessarily aligned. Any mutualistic trait that imposes a cost on the microbes that express it—such as producing dedicated molecules to interfere with pathogens or modulate host immune activity—represents a trade-off between the immediate selection against that cost and the long-term selection in favor of mutualism (Dethlefsen et al., 2007).
In summary, different views of humans as microbial habitats make different assumptions about the processes most important to the assembly and dynamics of the human microbiome. Community assembly can be conceptualized as being niche-based, dispersal-limited, historically contingent, or random, depending on the relative contributions of habitat conditions, colonist availability, arrival order (and timing), or chance-driven events, respectively, in shaping observed patterns (Figure A5-1). Metacommunity theory integrates the four processes described above and provides a useful framework for considering community assembly in the human body (Vellend, 2010).
Metacommunity Theory and the Human Microbiome
Various theoretical frameworks are used to study community assembly; one key framework is neutral theory (Hubbell, 2001), in which it is assumed that dispersal, diversification, and ecological drift are purely chance-driven processes. It is a neutral model because it invokes neither environmental selection nor inherent differences in species’ ability to disperse or diversify. Although neutral theory on its own is quite valuable in testing this null hypothesis, a broader framework for the assembly of the human microbiome might accommodate alternative theories and combine the strengths of transmission dynamic models (e.g., inclusion of host contact and carriage dynamics) with those of community ecology (e.g., focus on communities rather than individual pathogens). One such approach is metacommunity theory (Leibold et al., 2004), which could be especially useful for modeling host-associated communities.
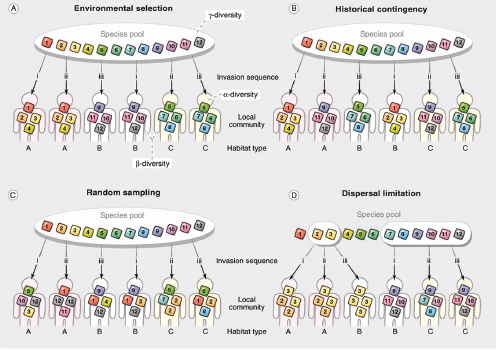
FIGURE A5-1 Alternative community assembly scenarios could give rise to the compositional variations observed in the human microbiota. Each panel shows the assembly of local communities in different habitat types from a pool of available species. (A to C) Each local community has access to all available colonists, but the order of invasion varies. In (A), local species composition is determined primarily by environmental selection: Regardless of invasion order, habitats with initially similar conditions select for similar assemblages. In (B), the opposite is true: Regardless of initial habitat conditions, historical contingencies (i.e., differences in the timing and order of species invasions) determine assemblage composition. In (C), neither habitat nor history matter: Local communities assemble via random draws from the species pool. (D) Dispersal barriers result in local communities that assemble from different species pools. For each of the pools, local communities may assemble as in (A), (B), or (C). The meaning of three different diversity measures is shown in (A): γ-diversity refers to the “regional” species pool (i.e., the total diversity of the local communities connected via dispersal); β-diversity refers to the differences between local communities (species turnover); and α-diversity refers to the diversity within a local community. Although multiple scenarios are likely to apply to any real-world setting, one may dominate. For example, differences between body habitats may be best explained by environmental selection, differences between siblings for the same habitat may be best explained by historical contingency, differences between monozygotic twins prior to weaning highlight the role of stochasticity, and differences between neonates born by cesarean section versus vaginal delivery are likely to be explained by dispersal limitation. [Adapted from (Chase, 2003; Fukami, 2010)].
Metacommunity theory views the world as a collection of patches—spatially distinct areas of suitable habitat surrounded by a matrix of unsuitable habitat. These patches each contain a community of organisms, and these spatially distinct communities are connected together to form a metacommunity by the dispersal of organisms from patch to patch. Human populations can be viewed likewise, with host-to-host dispersal linking microbial communities. Metacommunity theory is especially helpful for understanding the relative importance of dispersal and environmental selection in shaping host-associated communities (Mihaljevic, 2012), an issue that has received relatively little attention in studies of the human microbiome.
The predictions of metacommunity theory depend on the frequency and extent of dispersal, differences in the traits of individual organisms, and the degree to which patches vary in their environmental conditions (Ellis et al., 2006; Leibold et al., 2004). Dispersal can be infrequent and localized, or widespread and frequent, as discussed above. In some metacommunity models, patches are assumed to be environmentally identical, with movement among patches determining variation in community membership. Such models might be especially appropriate for populations of closely related hosts. Other models assume that patches vary strongly in their available niches, and that variation in community membership results at least in part from environmental selection (e.g., underpinned by host genetics or diet).
Metacommunity theory enables one to predict the conditions under which community dynamics within a patch are driven by immigration from outside versus local adaptation (Urban et al., 2008). Low dispersal rates favor adaptation within a patch, whereas high dispersal favors immigration. This concept could be useful, for example, in understanding responses to antibiotic use. If acquisition of antibiotic resistance were primarily a result of immigration, then interventions focused on quarantine and hygiene would be more effective than those focused on altering antibiotic duration or dose (see below).
Metacommunity theory has been used to elucidate the drivers of non–hostassociated microbial community membership and dynamics (Logue et al., 2011; Ofiteru et al., 2010; Van der Gucht et al., 2007), but it has rarely been used to study host-associated communities (Hovatter et al., 2011; Sloan et al., 2006)—for example, to explore the stringency of host selection and its dependence on the microbial group or the age of the host. Ultimately this information will result in a better understanding of how microbes are “filtered” by the host and, conversely, how microbes evade this filtering. This information is crucial if clinicians are to directly manipulate host-associated communities—for example, by designing probiotics that can evade host filtering and establish within a host.
The effective application of metacommunity theory (and assembly theory in general) to the human microbiome requires a preliminary understanding of how the microbiome varies across hosts and over time. We next review our current understanding of this variation, focusing on the dynamics of communities in
newly created habitats (e.g., neonatal colonization), after disturbance (e.g., after antibiotic treatment), and after invasion (e.g., by a pathogen). We chose these scenarios because they represent and reveal the fundamental types of assembly relevant to human health.
Postnatal Acquisition and Development of the Human Microbiome
Babies are born essentially sterile and acquire their microbiome from their surroundings. The postnatal assembly of the human microbiota plays an important role in infant health, providing resistance to pathogen invasion, immune stimulation, and other important developmental cues early in life (Mackie et al., 1999). Acute and chronic disorders, such as necrotizing enterocolitis, antibiotic-associated diarrhea, malnutrition, inflammatory bowel disease, and asthma have been linked to inadequate, inappropriate, or disrupted postnatal microbiome acquisition and development (Murgas and Neu, 2011). Mechanisms controlling the appearance of bacteria in healthy infants have been studied for well over a century (Escherich, 1988), with microbiome development having been likened to ecological succession (Mackie et al., 1999; Savage, 1977). The view of succession as a mode of community assembly has largely emphasized niche-based processes, but the importance of stochastic and/or historical events has also long been recognized.
In the absence of microbial invasion of the amniotic cavity, which is thought to be a rare, pathologic condition, rupture of membranes signals the moment when microbes, most likely of maternal vaginal origin, first gain access to the neonate. Vaginally delivered infants clearly receive a strong input of vaginal and possibly other urogenital or fecal microbiota as they pass through and exit the birth canal (Dominguez-Bello et al., 2010; Mändar and Mikelsaar, 1996). Vaginal microbiome composition in nonpregnant, reproductive-age women is highly dynamic and is characterized by at least five compositional classes delineated by different, dominant Lactobacillus species or a lack of Lactobacillus dominance. There is frequent class switching over time, including switching to and from compositions indicative of bacterial vaginosis, even in the absence of symptoms (Brotman et al., 2010; Gajer et al., 2012; Ravel et al., 2011). Whether these dynamics occur similarly in pregnant and postpartum women has important implications for the initial colonization of vaginally delivered infants; if they do, infant-to-infant variation in the composition of initial colonists may be imposed in some cases by maternal vaginal microbiome class at the time of delivery. Likewise, maternal gut microbiome types (Arumugam et al., 2011; Wu et al., 2011) may also determine the pool of colonists available to vaginally delivered infants at birth. Thus, variation among neonate microbiomes may reflect variation in maternal microbiomes, but this has not been widely tested for maternal habitats other than the vagina. At the time of delivery, microbiomes do not differ consistently among infant body sites (Dominguez-Bello et al., 2010), which implies
that sampling is driving initial community assembly, with minimal filtering by the infant host.
Delivery mode also determines microbial exposure at the time of birth. For example, infants delivered by cesarean section do not receive contributions from the vaginal microbiota, and instead are exposed initially to what appears to be ambient skin microbiota (Dominguez-Bello et al., 2010). Incidental exposures to maternal (or other) gut or vaginal microbiota may occur later in cesarean section infants, at low density or low frequency, but may be inadequate for outcompeting already established strains. For example, cesarean section infants display reduced abundances and/or incidences of colonization by the genera Bacteroides and Bifidobacterium early in development relative to infants born by vaginal delivery (Bennet and Nord, 1987; Penders et al., 2006). The effects of delivery mode can persist for months and may have consequences for infant health; cesarean section infants have a higher risk for some immune-mediated diseases (Kuitunen et al., 2009; van Nimwegen et al., 2011). The ambient environment may also play a role in colonization at delivery; infants delivered at home versus the hospital were colonized differently at 1 month of age (Penders et al., 2006). Thus, dispersal limitation imposed by certain medical interventions may contribute to interindividual variation early in life.
Over the first few months—roughly up until the first solid foods are introduced—a well-constrained range of stereotypical bacteria appear in the feces (distal gut), diversity generally increases, and aerobes are thought to be supplanted by facultative and then strict anaerobes (Mackie et al., 1999). Exclusive breast-feeding selects for increased abundance of particular Bifidobacterium species whose genome sequences reflect specialized use of human milk oligosaccharides and similar host-derived substrates (Sela et al., 2008), or for other bacteria such as Bacteroides that could compete for the same ecological niche (Marcobal et al., 2010). Strikingly, during this early phase, microbiota composition is highly dynamic within and between infants (Favier et al., 2002; Koenig et al., 2011; Mackie et al., 1999; Palmer et al., 2007; Trosvik et al., 2010), with temporal variation characterized by periods of relative stability (for varying lengths of time) punctuated by abrupt shifts in composition and structure. In some cases, these shifts can be linked with life events that likely impose environmental selection, such as fever, formula feeding, or antibiotic therapy (Koenig et al., 2011; Palmer et al., 2007; Savino et al., 2011). Extraordinarily parallel transitions observed in a pair of dizygotic twins suggest that stochastic exposures (shared exposures in their case) can also play an important role during this phase, driving within- and between-infant variation (Palmer et al., 2007). This finding underscores the need to better understand how infants sample their environment over time (e.g., whether outside-of-host environmental reservoirs, or host exchange scenarios—either direct or indirect, such as via fomites—prevail) and with regard to the frequency and extent of dispersal (as discussed above). Abrupt shifts might reflect opportunistic invasions by better-adapted species or changes in filtering by
the host. An infant’s unique developmental path through this early unstable phase may have longer-term health implications. For example, recent work has shown that colonization during the neonatal period has a particularly important effect on mucosal immune development (Olszak et al., 2012).
The introduction of solid foods and weaning are associated with the onset of a transition toward an adult-like gut microbiome. Differences attributable to early exposures such as delivery mode fade as microbiota compositions become more canalized. Life events such as illness, diet modification, and antibiotic therapy can still impose disturbances, although specific compositions appear to recover. Taxa characteristic of the adult eventually establish, but the process of microbial community assembly appears to extend past the first year of life and into childhood (Palmer et al., 2007; Koenig et al., 2011). If there is an imprint of microbial flow from parents to children, it is difficult to detect at early ages and/or emerges gradually later in life. In one study, fecal patterns of bacterial taxonomic diversity in 1-year-olds were not found to be more similar to those of their parents than to those of unrelated adults (Palmer et al., 2007); but in another study, patterns of microbial diversity in adult twins were slightly more similar to those of their mother (Turnbaugh et al., 2009). These findings suggest that we acquire microbes from sources other than, or in addition to, our family members. Further, there may be strong selection for an individualized microbiota. Describing the adult state as “stable” may not suffice when stability is defined as the permanent coexistence of locally occurring species, because even adult gut composition appears to change over time (Caporaso et al., 2011). In summary, microbiome assembly in newly created habitats likely involves a gradual shift from conditions under the strong influence of dispersal limitation (as well as stochastic and/or historical factors) toward conditions increasingly influenced by environmental selection (e.g., by diet), with weaning as a strong catalyst, and with development toward adult-like composition continuing into childhood.
Community Assembly After Disturbance: Antibiotics as a Paradigm
The assembly of human-associated microbial communities does not, in general, proceed smoothly to a stable climax state, which then resists further changes in composition. Disturbances often remove or kill some fraction of the community, providing an opportunity for remaining community members or new colonists to increase in abundance. For example, personal oral hygiene removes bacterial biofilm from teeth, and antibiotics affect not only the targeted pathogen but also members of the normal microbiota (Figure A5-2A). The former case represents a deliberate attempt to interrupt the development of microbial communities that might be associated with periodontitis; the latter case is an inadvertent consequence of medical intervention. In addition to directly inducing a shift in the community state, disturbance may also involve a change in the community’s habitat—for example, as the result of a change in host diet (Figure A5-2B). In
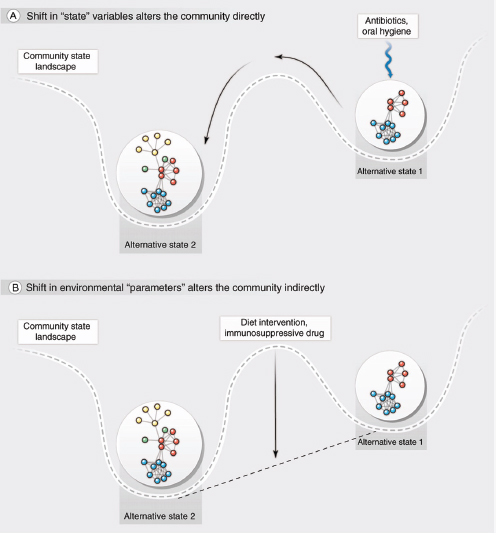
FIGURE A5-2 Disturbance can be illustrated using a stability landscape schematic. The ball represents the community; the changing horizontal position of the ball represents the changing community state. The depth of a basin indicates the likelihood of a community remaining in that basin despite frequent “buffeting” by minor disturbance (Walker et al., 2004) and hence the relative stability of the community state. Disturbance can alter the community directly (A) by changing its composition or activity, or indirectly (B) by changing the environmental parameters. In either case, the community can shift to an alternative state. In reality, continuous feedback between the community and its environment means that they change together. See (Lemon et al., 2012) for applications to therapy.
many cases, a crucial unknown is resilience—that is, the degree to which the post-disturbance community returns to its former state (Walker et al., 2004). Although most work on resilience has considered resilience in terms of taxonomic composition, assessment of function and ecosystem services may be even more important.
The effect of antibiotics on the gut microbiota serves as a paradigm for disturbances in human-associated communities. Antibiotics are now one of the most common and important forms of disturbance of the human microbiota; on any given day, approximately 1 to 3% of people in the developed world are exposed to pharmacologic doses of antibiotics (Goossens et al., 2005). Over the past several decades there has been increasing concern about the spread of antibiotic resistance among pathogens, as well as growing concern that antibiotic use may disrupt the host-microbe interactions that contribute to human health.
Antibiotic therapy is meant to achieve a sufficient concentration of the drug for a sufficient duration in a particular body site so that the targeted pathogen is eliminated. Even if this aim were always attained, the antibiotic will be found at a range of concentrations at many locations in the body, depending on the mode of administration and its pharmacodynamic properties. Where members of the indigenous microbiota are exposed to antibiotics that affect their growth without killing them, there is selection for resistance. Human gut and oral communities are recognized as reservoirs for the evolution and horizontal transfer of antibiotic resistance determinants, including to pathogens (Roberts and Mullany, 2010; Salyers et al., 2004; Smillie et al., 2011). However, antibiotic resistance among the microbiota is one of several mechanisms that may act to enhance the resilience of indigenous communities, hence preserving their beneficial ecosystem services. Others may include population-level resistance via stress-response signaling (Lee et al., 2010) and the existence of dormant persister cells (Vega et al., 2012) or refugium-like locations (e.g., mucus layer).
Only a handful of studies have used cultivation-independent surveys to examine the consequences of therapeutic doses of antibiotics on the human gut microbiota (e.g., Dethlefsen et al., 2008; Dethlefsen and Relman, 2011; Jernberg et al., 2007; Jakobsson et al., 2010; Vega et al., 2012; Young and Schmidt, 2004). These studies—although they examined different antibiotics by means of various sampling strategies, treatment durations, and analytical approaches—all have found that antibiotics alter the composition of the gut microbiota, and that the abundance of most taxa begins to return to prior levels within several weeks. However, the studies are also consistent in showing that various taxa recover to different extents and that some do not recover over the duration of the study. The antibiotic effect is greater than the routine temporal variability of community composition (Dethlefsen et al., 2008; Dethlefsen and Relman, 2011; Jernberg et al., 2007). Some studies have revealed that the composition of strains within a taxon is sometimes altered, even if the relative abundance of the taxon as a whole has returned to pre-antibiotic levels. In both of the studies that involved measurements of the prevalence of antibiotic-resistant strains, elevated levels of
resistance persisted to the end of the study (Jernberg et al., 2007; Jakobsson et al., 2010).
Overall, research suggests that the human gut microbiota of generally healthy adults is largely, but not entirely, resilient to short courses of antibiotic therapy, whereas clinical evidence indicates that extended or repeated courses are more likely to result in serious complications such as the invasion and bloom of Clostridium difficile (Owens et al., 2008). Perhaps over short courses of antibiotics, a sufficient, although possibly quite small, number of residual cells from most of the large, preexisting populations survives to recolonize the gut. An increasing number of these residual cells may be lost with longer or repeated courses of antibiotics. Thus, reassembly of the microbial community after extended antibiotic treatment may require colonization from outside the host—a process that would likely be more variable and require a longer period of time than reassembly via the filtering of existing populations in the host. In addition, the microbiome may be highly vulnerable to invasion by (and/or blooms of) pathogens during recovery after disturbance, because resources are in high abundance and resident populations are diminished. The longer recovery time required after extended antibiotic treatment could lead to a higher probability of invasion by pathogenic strains. One can envision a more enlightened strategy for the clinical use of antibiotics that includes pretreatment estimates of a patient’s microbial community resilience, based on the use of a standardized disturbance and monitoring of key community products, mapping of the community stability landscape, and assessment of the likelihood for community displacement and adoption of a disadvantageous, altered state. Assessments of elevated risk, or of loss of resilience, might then prompt efforts at restoration (see Lemon et al., 2012).
Little is known regarding the response of the microbiome to frequent antibiotic use. When disturbances take place with a magnitude or frequency beyond what a community has had an opportunity to adapt to, ecological surprises may occur (Paine et al., 1998). Such frequent disturbances may allow the persistence of microbial taxa that are inferior competitors within a given host but are nonetheless maintained across hosts because they have traits that result in widespread and frequent dispersal (i.e., “fugitive” taxa). Such a scenario is analogous to the patch dynamics paradigm of metacommunity theory (Leibold et al., 2004).
Assembly of the Human Microbiome in the Context of Invaders (Pathogens)
It is naïve to consider only host and pathogen when predicting the likelihood of microbial disease. Host-associated microbial communities play an important role in preventing disease, and although outside the scope of this discussion, they may also promote pathology, as in the case of inflammatory bowel disease (Elinav et al., 2011). It may be useful to view some pathogens as invasive species, and to view the consequences of invasion as a special case of community assembly. As
with invasive species in more traditionally studied settings, the ability of a species to invade a particular community depends largely on niche opportunities— that is, the filters imposed by the abiotic environment and the resistance of the community to colonization by an exotic species (Shea and Chesson, 2002). A successful invasion involves dispersal of an invader to a new community, initial colonization, and proliferation; these steps are influenced by the same processes as in community assembly more generally.
The environment created by the host determines the number of potential niche opportunities. The nature of this environment is influenced by a number of conditions, including “abiotic” factors (such as oxygen levels, pH, and temperature) as well as the abundance and types of available resources (such as the composition of the host’s diet) (Turnbaugh et al., 2006) and carbon sources provided directly by the host (such as mucosal poly- and oligosaccharides) (Sonnenburg et al., 2004). In addition, the host immune system acts as an important environmental filter to limit the spatial extent of the microbiota’s available niches. The main functions of the mucosal immune system are to create an inhospitable buffer zone between the microbiota and the host epithelium, and to minimize the incidence of systemic inflammation that would normally be induced in the face of so many bacterial products (Duerkop et al., 2009; Hooper et al., 2012; Macpherson et al., 2012). The immune system performs these functions through three general mechanisms: (i) physical barriers such as the inner mucus layer of the colon and stomach, which is generally impenetrable to bacterial cells (Johansson et al., 2008); (ii) antimicrobial peptides and mucosal antibodies in the mucus layer that further hinder bacterial colonization of the epithelium (Duerkop et al., 2009); and (iii) innate and adaptive immune responses within the regional lymphatic tissues (Macpherson et al., 2012). These three mechanisms, in most healthy hosts, select for bacterial species that do well at or near mucosal surfaces or strong barriers such as the skin. However, host filtering is not the only factor influencing the ability of pathogens to invade the host-microbiota community.
One of the most important roles of the microbiota in mediating host-pathogen interactions is protection of the host from pathogen invasion, or “colonization resistance” (Duerkop et al., 2009; Sekirov and Finlay, 2009). Protection is achieved through induction of the innate and adaptive branches of the immune system, creating an environment that is unfavorable to pathogens (as illustrated by the observation that axenic mice [Macpherson et al., 2004] and zebrafish [Kanther and Rawls, 2010] have diminished immune responses and impaired barriers to infection), and through direct competition (or community filtering, e.g., by lowering vaginal pH). In the latter case, pathogens are kept at bay by competition with the microbiota for space and resources. This protective effect is demonstrated by the increased susceptibility to infection of hosts that have had their microbiota altered by antibiotics, a phenomenon well documented by Miller and Bohnhoff in the early 1960s with Salmonella invasion of mice pretreated with antibiotics (Bohnhoff and Miller, 1962). The ability of certain anaerobes to limit the invasion
and growth of Clostridium perfringens in a diet-dependent manner is an example of competition for resources (Yurdusev et al., 1989). Bifidobacterium breve produces an exopolysaccharide that protects it from the immune response; this allows it to compete for space and colonize the mouse gut at high loads both in the lumen and at the epithelial surface without inducing inflammation (Fanning et al., 2012). Even if invaders do gain a foothold, the indigenous microbiota can block lethality: In mice, some B. longum strains can protect against enterohemorrhagic E. coli–mediated death by inhibiting translocation of Shiga toxin from lumen to blood (Fukuda et al., 2011).
By viewing pathogens as invasive species, we see that the contexts in which they are able to cause disease are the same as those required for any other species that invades and proliferates in a community. Niche opportunities can result from exploiting novel or excess resources (from the host’s food), outcompeting a commensal species for the same resource, or, perhaps most important, exploiting niches left open after a disturbance. The importance of exploiting disturbance is well illustrated by the increasing number of cases of disease caused by C. difficile (Kelly and LaMont, 2008), which is a “weedy” exotic (or native) species that can rapidly fill niches once they are vacant but, in most cases, is eventually removed or kept at low abundance in the absence of disturbance. Salmonella enterica serovar Typhimurium is an example of an exotic invasive that exploits disturbance, but in this case, it also causes the disturbance it exploits. S. Typhimurium expresses many virulence factors that induce inflammation in the mammalian intestine. A mutant S. Typhimurium strain lacking these virulence factors is unable to invade the gut community and cause disease; however, if inflammation is provided by some other mechanism, avirulent strains are able to invade the host communities (Stecher et al., 2007). Inflammation likely reduces the abundances of other bacteria that would compete with pro-inflammatory pathogens for space. As one possible mechanism, inflammation causes the intestine to produce tetrathionate, which S. Typhimurium uses as an electron acceptor for respiring ethanolamine, a carbon source that cannot be exploited by other bacteria, thus avoiding competition for nutrients (Thiennimitr et al., 2011). By causing acute inflammation, the pathogen is able to alter the native microbiota and effectively colonize and proliferate.
In contrast to the above examples, a reduction in disturbance frequency can also promote invasion by pathogens. Patients with cystic fibrosis produce thickened mucus, which inhibits the ability of the cilia to remove foreign material from normally sterile lung airways. This lack of constant removal (i.e., impaired innate immune host-filtering mechanism), among other factors, allows for the establishment of bacterial communities that would normally not be able to persist at that site (Klepac-Ceraj et al., 2010).
In summary, predicting the success and outcome of infection by pathogens can be aided by framing the issue as an ecological problem of community assembly. Invasion ecology highlights the importance of niche opportunities as
determinants of success of invasion, and the manipulation of which might help in pathogen control and disease prevention. Experimental models using gnotobiotic organisms such as mice and zebrafish will be helpful in understanding the role of community diversity, as well as the role of particular community members in conferring colonization resistance through indirect inhibition and resource competition. In addition, the frequency and magnitude of disturbance plays a crucial role in facilitating colonization by exotic invasives as well as the expansion of native species. Finding ways, through prebiotics, probiotics, or pharmabiotics, to alter pathogen or other bacterial species abundances (or to inhibit their detrimental effects on the host) in a specific manner without causing additional disturbance to the community will be very important for preventing and treating disease caused by invasive species.
Translating Ecological Understanding into Clinical Practice
An improved understanding, informed by ecological theory, of how microbiomes assemble could alter clinical practice by changing the perspective clinicians bring to the treatment of infectious disease and chronic inflammatory disorders. The traditional perspective has been to think of the human body as a battleground on which physicians attack pathogens with increasing force, occasionally having to resort to a scorched-earth approach to rid a body of disease. Although this perspective has been very successful for several diseases, it has come at a great cost. Even for those diseases for which it has worked, the collateral damage can be severe. As we have discussed, antibiotics often kill more than the target organisms (Dethlefsen and Relman, 2011) and can increase the chance of invasion by unwanted organisms (such as C. difficile [Kelly and LaMont, 2008]).
The body-as-battleground approach ignores the community context of infectious disease and chronic inflammation, and does not take into account our increasing knowledge regarding the assembly of the human microbiome. We suggest that it is time for clinicians to abandon the war metaphor (Lederberg, 2000). Given the ecological parallels between assembly of the human microbiome and assembly of other ecological communities, we suggest that human medicine has more in common with park management than it does with battlefield strategy. To effectively manage a plant or animal community requires a multipronged approach of habitat restoration, promotion of native species, and targeted removal of invasives. We describe below some examples of how such a human-as-habitat approach might alter clinical practice.
An Ecological Approach to Managing Invasions
An understanding of the dominant mechanisms of community assembly could directly alter how clinicians treat disease. Consider, for example, the rise of drug-resistant pathogens during the course of drug treatment. We can consider
this a “special case” of community assembly, much as we did invasion by a pathogen (see above). We can ask: What is the relative importance of dispersal, diversification, environmental selection, and ecological drift in the successful invasion by this drug-resistant strain? If the source of these strains is primarily through random sampling of the external environment, then the most effective preventive strategy may be quarantine and enhanced hygiene. In contrast, if such strains arise primarily through diversification of resident taxa, then multidrug treatment may be more effective (to make successful evolution more difficult). If the drug-resistant strains are already present at the outset of treatment and increase in abundance via environmental selection, then drug cycling may be the most effective treatment to reduce the overall competitive advantage of the resistant strains. If drug-resistant strains establish primarily through ecological drift, then disturbance may be crucial to their establishment (freeing up ecological “space” for their invasion). In this case, reducing disturbance of the resident microbiota may be most effective. In this way, a detailed understanding of the relative importance of different community assembly processes can be used to tailor the treatment of disease.
Health as a Product of Ecosystem Services
Humans benefit from a variety of processes supplied by natural ecosystems. Collectively these benefits are known as ecosystem services (Daily et al., 1997). There is growing evidence that human health is a collective property of the human body and its associated microbiome, and thus could be considered a net effect of ecosystem services. We envision clinical medicine focused on managing the human body and its associated microbiome to preserve these ecosystem services. How might this be accomplished? In general ecology, the management of an ecosystem service requires four basic steps (Allan and Stankey, 2009): (i) identification of ecosystem service providers (ESPs; taxa that provide specific ecosystem services) and characterization of their functional roles; (ii) determination of how community context influences the function of these providers; (iii) assessment of key environmental factors influencing the provision of services; and (iv) measurement of the spatial and temporal scales at which these providers and their functions operate. This general framework would work equally well for human health-associated ecosystem services. If studies of the human microbiome were structured around these four priorities, the development of an ecological approach to medicine could be accelerated. Progress has been made in identifying ESPs (“biomarkers”; see Lemon et al., 2012); for example, declines in Faecalibacterium prausnitzii are associated with inflammatory bowel disease, and this organism may be an ESP for health in the human gut (Sokol et al., 2008).
Adaptive Management of the Human Body
Transitioning clinical practice from the body-as-battleground to the human-as-habitat perspective will require rethinking how one manages the human body. In the management of plant and animal communities, a system-level approach known as “adaptive management” has become popular. This approach is a structured, iterative process of decision making, one that uses system monitoring to continually update management decisions (Allan and Stankey, 2009). It has been successfully used to manage biodiversity in a variety of habitats, including communities in highly disturbed environments affected by overfishing and by climate change (Allan and Stankey, 2009). For the human body, we envision that this approach would involve monitoring of the microbiome during health, to establish a healthy baseline, with more intensive monitoring during disease and treatment. This will require the development of new diagnostic tools that are both accurate and sufficiently rapid to inform decisions regarding therapeutics [see Lemon et al., 2012). Such diagnostics are not yet feasible, but given recent advances in our ability to survey the human microbiome, this possibility is not far in the future, especially if we are able to identify particular components of the human microbiome that contribute disproportionately to the maintenance of human health. An adaptive management approach to clinical medicine provides a wonderful example of personalized medicine, with treatments tailored to individuals on the basis of diagnostic changes in an individual’s microbiome, and continually adjusted through regular monitoring. Such an information-intensive approach, guided by ecological theory, has the potential to revolutionize the treatment of disease.
Acknowledgments
We thank members of the Relman and Bohannan laboratories, and in particular E. Bik, N. Qvit-Raz, G. Schmid, and K. Shelef (Stanford), and K. Guillemin, J. Green, Z. Stephens, and A. Burns (Oregon) for valuable ideas and critique. E.K.C. is a Walter V. and Idun Berry Postdoctoral Fellow. B.J.M.B. is supported by NIH grant R01GM095385. D.A.R. is supported by a Distinguished Clinical Scientist Award from the Doris Duke Charitable Trust, NIH Pioneer Award DP1OD000964, and the Thomas C. and Joan M. Merigan Endowment at Stanford University. There are no conflicts of interest.
References
J. R. Mihaljevic, Linking metacommunity theory and symbiont evolutionary ecology. Trends Ecol. Evol. 10.1016/j.tree.2012.01.011 (2012). 10.1038/nrmicro1643
J. I. Prosser et al., The role of ecological theory in microbial ecology. Nat. Rev. Microbiol. 5, 384 (2007).
C. Palmer, E. M. Bik, D. B. DiGiulio, D. A. Relman, P. O. Brown, Development of the human infant intestinal microbiota. PLoS Biol. 5, e177 (2007).
J. Ravel et al., Vaginal microbiome of reproductive-age women. Proc. Natl. Acad. Sci. U.S.A. 108 (suppl. 1), 4680 (2011).
G. D. Wu et al., Linking long-term dietary patterns with gut microbial enterotypes. Science 334, 105 (2011).
M. Vellend, Conceptual synthesis in community ecology. Q. Rev. Biol. 85, 183 (2010).
L. Dethlefsen, M. McFall-Ngai, D. A. Relman, An ecological and evolutionary perspective on human-microbe mutualism and disease. Nature 449, 811 (2007).
M. A. O’Malley, The nineteenth century roots of ‘everything is everywhere’. Nat. Rev. Microbiol. 5, 647 (2007).
J. Koopman, Modeling infection transmission. Annu. Rev. Public Health 25, 303 (2004).
C. S. Smillie et al., Ecology drives a global network of gene exchange connecting the human microbiome. Nature 480, 241 (2011).
M. C. Urban et al., The evolutionary ecology of metacommunities. Trends Ecol. Evol. 23, 311 (2008).
T. Fukami, H. J. Beaumont, X. X. Zhang, P. B. Rainey, Immigration history controls diversification in experimental adaptive radiation. Nature 446, 436 (2007).
L. V. Hooper, D. R. Littman, A. J. Macpherson, Interactions between the microbiota and the immune system. Science 336, 1268 (2012). 10.1038/nature00823
B. Kerr, M. A. Riley, M. W. Feldman, B. J. Bohannan, Local dispersal promotes biodiversity in a real-life game of rock-paper-scissors. Nature 418, 171 (2002).
S. P. Hubbell, The Unified Neutral Theory of Biodiversity and Biogeography (Princeton Univ. Press, Princeton, NJ, 2001).
M. A. Leibold et al., The metacommunity concept: A framework for multi-scale community ecology. Ecol. Lett. 7, 601 (2004). 10.1111/j.1461
A. M. Ellis, L. P. Lounibos, M. Holyoak, Evaluating the long-term metacommunity dynamics of tree hole mosquitoes. Ecology 87, 2582 (2006). 10.1890/0012-9658(2006)87[2582
J. B. Logue, N. Mouquet, H. Peter, H. Hillebrand, Metacommunity Working Group, Empirical approaches to metacommunities: A review and comparison with theory. Trends Ecol. Evol. 26, 482 (2011).
I. D. Ofiteru et al., Combined niche and neutral effects in a microbial wastewater treatment community. Proc. Natl. Acad. Sci. U.S.A. 107, 15345 (2010).
K. Van der Gucht et al., The power of species sorting: Local factors drive bacterial community composition over a wide range of spatial scales. Proc. Natl. Acad. Sci. U.S.A. 104, 20404 (2007).
S. R. Hovatter, C. Dejelo, A. L. Case, C. B. Blackwood, Metacommunity organization of soil microorganisms depends on habitat defined by presence of Lobelia siphilitica plants. Ecology 92, 57 (2011). 10.1890/10
W. T. Sloan et al., Quantifying the roles of immigration and chance in shaping prokaryote community structure. Environ. Microbiol. 8, 732 (2006). 10.1111/j.1462
R. I. Mackie, A. Sghir, H. R. Gaskins, Developmental microbial ecology of the neonatal gastrointestinal tract. Am. J. Clin. Nutr. 69, 1035S (1999). 10232646
R. Murgas Torrazza, J. Neu, The developing intestinal microbiome and its relationship to health and disease in the neonate. J. Perinatol. 31 (suppl. 1), S29 (2011).
Th. Escherich, The intestinal bacteria of the neonate and breast-fed infant. Rev. Infect. Dis. 10, 1220 (1988).
D. C. Savage, Microbial ecology of the gastrointestinal tract. Annu. Rev. Microbiol. 31, 107 (1977).
M. G. Dominguez-Bello et al., Delivery mode shapes the acquisition and structure of the initial microbiota across multiple body habitats in newborns. Proc. Natl. Acad. Sci. U.S.A. 107, 11971 (2010).
R. Mändar, M. Mikelsaar, Transmission of mother’s microflora to the newborn at birth. Biol. Neonate 69, 30 (1996).
R. M. Brotman, J. Ravel, R. A. Cone, J. M. Zenilman, Rapid fluctuation of the vaginal microbiota measured by Gram stain analysis. Sex. Transm. Infect. 86, 297 (2010).
P. Gajer et al., Temporal dynamics of the human vaginal microbiota. Sci. Transl. Med. 4, 132ra52 (2012). 10.1136/sti.2009.040592
M. Arumugam et al., MetaHIT Consortium, Enterotypes of the human gut microbiome. Nature 473, 174 (2011).
R. Bennet, C. E. Nord, Development of the faecal anaerobic microflora after caesarean section and treatment with antibiotics in newborn infants. Infection 15, 332 (1987).
J. Penders et al., Factors influencing the composition of the intestinal microbiota in early infancy. Pediatrics 118, 511 (2006). 10.1542/peds.2005
M. Kuitunen et al., Probiotics prevent IgE-associated allergy until age 5 years in cesarean-delivered children but not in the total cohort. J. Allergy Clin. Immunol. 123, 335 (2009).
F. A. van Nimwegen et al., Mode and place of delivery, gastrointestinal microbiota, and their influence on asthma and atopy. J. Allergy Clin. Immunol. 128, 948, e1 (2011).
D. A. Sela et al., The genome sequence of Bifidobacterium longum subsp. infantis reveals adaptations for milk utilization within the infant microbiome. Proc. Natl. Acad. Sci. U.S.A. 105, 18964 (2008).
A. Marcobal et al., Consumption of human milk oligosaccharides by gut-related microbes. J. Agric. Food Chem. 58, 5334 (2010).
C. F. Favier, E. E. Vaughan, W. M. De Vos, A. D. Akkermans, Molecular monitoring of succession of bacterial communities in human neonates. Appl. Environ. Microbiol. 68, 219 (2002). 10.1128/AEM.68.1.219
J. E. Koenig et al., Succession of microbial consortia in the developing infant gut microbiome. Proc. Natl. Acad. Sci. U.S.A. 108 (suppl. 1), 4578 (2011).
P. Trosvik, N. C. Stenseth, K. Rudi, Convergent temporal dynamics of the human infant gut microbiota. ISME J. 4, 151 (2010).
F. Savino et al., Faecal microbiota in breast-fed infants after antibiotic therapy. Acta Paediatr. 100, 75 (2011). 10.1111/j.1651
T. Olszak et al., Microbial exposure during early life has persistent effects on natural killer T cell function. Science 336, 489 (2012).
P. J. Turnbaugh et al., A core gut microbiome in obese and lean twins. Nature 457, 480 (2009).
J. G. Caporaso et al., Moving pictures of the human microbiome. Genome Biol. 12, R50 (2011). 10.1186/gb-2011-12-5
B. Walker, C. S. Holling, S. R. Carpenter, A. Kinzig, Resilience, adaptability and transformability in social-ecological systems. Ecol. Soc. 9, 5 (2004). 10.1186/gb-2011-12-5
H. Goossens, M. Ferech, R. Vander Stichele, M. Elseviers, ESAC Project Group, Outpatient antibiotic use in Europe and association with resistance: A cross-national database study. Lancet 365, 579 (2005). 15708101
A. P. Roberts, P. Mullany, Oral biofilms: A reservoir of transferable, bacterial, antimicrobial resistance. Expert Rev. Anti Infect. Ther. 8, 1441 (2010).
A. A. Salyers, A. Gupta, Y. Wang, Human intestinal bacteria as reservoirs for antibiotic resistance genes. Trends Microbiol. 12, 412 (2004).
H. H. Lee, M. N. Molla, C. R. Cantor, J. J. Collins, Bacterial charity work leads to population-wide resistance. Nature 467, 82 (2010).
N. M. Vega, K. R. Allison, A. S. Khalil, J. J. Collins, Signaling-mediated bacterial persister formation. Nat. Chem. Biol. 8, 431 (2012).
L. Dethlefsen, S. Huse, M. L. Sogin, D. A. Relman, The pervasive effects of an antibiotic on the human gut microbiota, as revealed by deep 16S rRNA sequencing. PLoS Biol. 6, e280 (2008).
L. Dethlefsen, D. A. Relman, Incomplete recovery and individualized responses of the human distal gut microbiota to repeated antibiotic perturbation. Proc. Natl. Acad. Sci. U.S.A. 108 (suppl. 1), 4554 (2011).
H. E. Jakobssonet al., Short-term antibiotic treatment has differing long-term impacts on the human throat and gut microbiome. PLoS ONE 5, e9836 (2010).
C. Jernberg, S. Löfmark, C. Edlund, J. K. Jansson, Long-term ecological impacts of antibiotic administration on the human intestinal microbiota. ISME J. 1, 56 (2007).
V. B. Young, T. M. Schmidt, Antibiotic-associated diarrhea accompanied by large-scale alterations in the composition of the fecal microbiota. J. Clin. Microbiol. 42, 1203 (2004). 10.1128/JCM.42.3.1203
R. C. Owens Jr., C. J. Donskey, R. P. Gaynes, V. G. Loo, C. A. Muto, Antimicrobial-associated risk factors for Clostridium difficile infection. Clin. Infect. Dis. 46 (suppl. 1), S19 (2008).
K. P. Lemon, G. C. Armitage, M. A. Fischbach, Microbiome-targeted therapies: An ecological perspective. Sci. Transl. Med. 4, 137rv5 (2012). 10.1126/science.1208344
R. T. Paine, M. J. Tegner, E. A. Johnson, Compounded perturbations yield ecological surprises. Ecosystems 1, 535 (1998).
E. Elinavet al., NLRP6 inflammasome regulates colonic microbial ecology and risk for colitis. Cell 145, 745 (2011). 10.1016/S0169-5347(02)02495
K. Shea, P. Chesson, Community ecology theory as a framework for biological invasions. Trends Ecol. Evol. 17, 170 (2002). 10.1016/S0169-5347(02)02495
P. J. Turnbaugh et al., An obesity-associated gut microbiome with increased capacity for energy harvest. Nature 444, 1027 (2006).
J. L. Sonnenburg, L. T. Angenent, J. I. Gordon, Getting a grip on things: How do communities of bacterial symbionts become established in our intestine? Nat. Immunol. 5, 569 (2004).
B. A. Duerkop, S. Vaishnava, L. V. Hooper, Immune responses to the microbiota at the intestinal mucosal surface. Immunity 31, 368 (2009).
A. J. Macpherson, M. B. Geuking, E. Slack, S. Hapfelmeier, K. D. McCoy, The habitat, double life, citizenship, and forgetfulness of IgA. Immunol. Rev. 245, 132 (2012). 10.1111/j.1600
M. E. Johansson et al., The inner of the two Muc2 mucin-dependent mucus layers in colon is devoid of bacteria. Proc. Natl. Acad. Sci. U.S.A. 105, 15064 (2008).
I. Sekirov, B. B. Finlay, The role of the intestinal microbiota in enteric infection. J. Physiol. 587, 4159 (2009).
A. J. Macpherson, N. L. Harris, Interactions between commensal intestinal bacteria and the immune system. Nat. Rev. Immunol. 4, 478 (2004).
M. Kanther, J. F. Rawls, Host-microbe interactions in the developing zebrafish. Curr. Opin. Immunol. 22, 10 (2010).
M. Bohnhoff, C. P. Miller, Enhanced susceptibility to Salmonella infection in streptomycin-treated mice. J. Infect. Dis. 111, 117 (1962).
N. Yurdusev, M. Ladire, R. Ducluzeau, P. Raibaud, Antagonism exerted by an association of a Bacteroides thetaiotaomicron strain and a Fusobacterium necrogenes strain against Clostridium perfringens in gnotobiotic mice and in fecal suspensions incubated in vitro. Infect. Immun. 57, 724 (1989). 2537255
S. Fanning et al., Bifidobacterial surface-exopolysaccharide facilitates commensal-host interaction through immune modulation and pathogen protection. Proc. Natl. Acad. Sci. U.S.A. 109, 2108 (2012).
S. Fukuda et al., Bifidobacteria can protect from enteropathogenic infection through production of acetate. Nature 469, 543 (2011).
C. P. Kelly, J. T. LaMont, Clostridium difficile—more difficult than ever. N. Engl. J. Med. 359, 1932 (2008).
B. Stecher et al., Salmonella enterica serovar typhimurium exploits inflammation to compete with the intestinal microbiota. PLoS Biol. 5, e244 (2007).
P. Thiennimitr et al., Intestinal inflammation allows Salmonella to use ethanolamine to compete with the microbiota. Proc. Natl. Acad. Sci. U.S.A. 108, 17480 (2011).
V. Klepac-Ceraj et al., Relationship between cystic fibrosis respiratory tract bacterial communities and age, genotype, antibiotics and Pseudomonas aeruginosa. Environ. Microbiol. 12, 1293 (2010). 10.1111/j.1462
J. Lederberg, Infectious history. Science 288, 287 (2000).




















