
LESSONS FROM STUDYING INSECT SYMBIOSES28
As in mammals, insect health is strongly influenced by the composition and activities of resident microorganisms. However, the microbiota of insects is generally less diverse than that of mammals, allowing microbial function in insects to be coupled to individual, identified microbial species. This trait of insect symbioses facilitates our understanding of the mechanisms that promote insect-microbial coexistence and the processes by which the microbiota affect insect well-being. As a result, insects are potentially ideal models to study various aspects of interactions between the host and its resident microorganisms that would be impractical or unfeasible in mammals and to generate hypotheses for subsequent testing in mammalian models.
Introduction
The common condition for animals is to be chronically infected by microorganisms, most of which are benign or beneficial. The influence of these resident microorganisms on their animal host is profound and operates at two levels: in physiological time, such that the physiology and well-being of the animal is influenced by the composition, density, and activities of colonizing microorganisms; and in evolutionary time, through selection on the magnitude and pattern of the animal response to the infecting microorganisms. We can only understand animal-microbial interactions by combining these physiological and evolutionary perspectives.
There is immense variation in the detail of the interactions between animals and their resident microbiota. A proper grasp of the nature of interactions between humans and their microbiota can only come from an awareness of the parallels and contrasts with other animals. Insects represent a superb system for comparison because, as a group, they display far greater diversity than mammals in their interactions, including some associations of remarkable morphological intimacy and molecular integration. Research on insect-microbial interactions has been invigorated in recent years by the technical advances that make it possible to identify and study unculturable microorganisms. The recent gains in mechanistic understanding of the relationships in molecular terms have immediate relevance to research on the impact of resident microbes on the physiology and health of humans.
________________
28 Reprinted from Cell Host & Microbe, 10(4), A. E. Douglas, Lessons from studying insect symbioses, Pp. 359-367, Copyright (2011), with permission from Elsevier.
29 Department of Entomology, Comstock Hall, Cornell University, Ithaca, NY 14853, USA.
This review summarizes these recent advances by focusing on four topics: the mechanisms that promote coexistence of insect host and microbes; the contribution of microbes to insect nutrition and defense; insect dependence on their resident microbiota; and coevolutionary interactions between the partners. The impact of insect research on our understanding of associations in humans is also briefly considered.
Diversity of Insect-Microbial Symbioses
Insects and mammals share the common feature that they are inhabited by microorganisms. Even so, there are two important differences in the host-microbial relationship between the two animal groups: (1) the diversity of the microbiota tends to be an order of magnitude greater in mammals than in insects, and (2) many insects, but no known mammals, have beneficial intracellular microorganisms. To address these differences, the gut microbiota and intracellular symbioses of insects are considered in turn.
The gut lumen is very densely colonized by microbes in most mammals and insects. The animal gut can be considered a portion of the external environment in which the conditions and resources are controlled largely by the animal. Microorganisms associated with food have unrestricted access to the gut; and those that can tolerate, modulate or evade the digestive processes and immune function of the animal gut gain access to a nutrient-rich environment and a vehicle for dispersal via the feces. Nevertheless, the habitats in the insect and mammalian guts cannot be equivalent because the patterns in the composition and diversity of the gut microbiota are markedly different. The identification of bacterial operational taxonomic units (OTU) by 97% sequence identity of 16S rRNA gene sequences revealed that the gut of most insects bears < 20–30 taxa (Dillon and Dillon, 2004; Robinson et al., 2010; and Wong et al., 2011), and that of mammals (or their feces) yield 500–1000 taxa (Dethlefsen et al., 2007 and Nemergut et al., 2011). The reasons for this difference are unclear. As with many other low diversity habitats (Connell, 1978), insect guts tend to be transient and have extreme disturbance regimes. Compounding the short life span of many insects, microbes associated with the foregut and hindgut are eliminated at every insect molt (when the cuticle lining these gut regions is shed), and all microbes are shed when the larval gut is broken down and the adult gut develops during metamorphosis of higher (“holometabolous”) insects, including the true flies, butterflies, and beetles. It has also been suggested that the adaptive immune system of mammals may promote microbial diversity through its greater capacity than the innate immune system to discriminate among different microorganisms, enabling the mammalian host to maintain complex multispecies consortia (McFall-Ngai, 2007).
The greater microbial diversity in individual mammals as opposed to insects is overlaid by a greater diversity across all insects relative to all mammals. Thus, the dominant microbes in all mammals are Bacteroidetes and Firmicutes, but the
gut microbiota of insects vary widely among different taxa, including Proteobacteria, Firmicutes, and Protists (Brugerolle and Radek, 2006; Dillon and Dillon, 2004; Morales-Jiménez et al., 2009; and Robinson et al., 2010). This difference could be a consequence of the greater variation of diets and gut physiology in insects as opposed to mammals, which is linked to the greater phylogenetic diversity of insects. (The class Mammalia comprises 5000 species, and the class Insecta has 900,000 known—and 2–30 million predicted—species.) For example, all mammals have an extremely acidic stomach, but an equivalent region is rare among insects and apparently restricted to the higher Diptera, including Drosophila (Shanbhag and Tripathi, 2009). The pH in many insects lies within the range of 6–8 units, and some, notably lepidopteran caterpillars, have a very basic midgut region, at 11–12 pH units (Wieczorek et al., 2009).
The second dominant habitat in insects utilized by microorganisms is cells. An estimated 10%–20% of insect species bear intracellular symbionts that are localized to specialized cells, known as bacteriocytes, whose sole function appears to be to house and maintain their symbionts. These associations are widespread or universal in several groups, notably cockroaches, hemipterans, ten families of beetles, and lice, and they also occur in some flies and ants (see Table 1 in Douglas, 2007). Although they have evolved independently multiple times, the microorganisms are invariably transmitted vertically (from mother to offspring), usually via the eggs in the female ovaries. The resultant perfect congruence between the phylogenies of the microbial symbiont and their insect hosts, in some insect groups over more than 100–200 million years (Dale and Moran, 2006; and Moran et al., 2005), has no parallel in mammals. Correlated with the long history and intimacy of the association, both the insect and microbial partners are dependent on the relationship, such that insects experimentally deprived of their bacteria fail to grow and reproduce (Douglas, 2007), and the symbionts generally have much-reduced genomes (<1 Mb) and are unculturable (Dale and Moran, 2006).
Bacteriocytes are not the only insect cell type that contains microorganisms. Across all insect groups, microorganisms have been reported in cells of various organs, including the fat body, gut epithelium, and gonads. Some of these taxa (e.g., Wolbachia, Hamiltonella) can occupy multiple compartments, within and between the cells of insect organs and in the blood, for example, although the location of the microorganisms may vary with host and symbiont genotype as well as the age and physiological condition of the host (Oliver et al., 2010; and Werren et al., 2008). This trait of broad and variable tissue distribution is facilitated by the open circulatory system of insects, meaning that blood is not restricted to closed vessels, but is in direct contact with various organs and moves relatively sluggishly around the body.
Bacteriocyte symbioses are unknown among mammals, and virtually all microorganisms that adopt an intracellular phase in mammals mediate chronic or acute pathogenic infections. There is no entirely satisfactory explanation for this
difference. It has been suggested repeatedly that the adaptive immune system of vertebrates poses a very high barrier to the evolution of intracellular microorganisms, which has been overcome predominantly by pathogens. Consistent with this argument, intracellular symbionts are commonplace among many invertebrates, but extremely rare across all vertebrates, with the alga Oophila in the embryos of a salamander (Kerney et al., 2011) as the sole documented example.
Mechanisms for Coexistence
Much more is known about the patterns than the mechanisms that promote coexistence of insects and their resident microbes. The key feature of these patterns is a relative uniformity in the location and numbers of microorganisms, suggesting that the host exerts tight controls over its symbionts. A rich literature has demonstrated that symbionts of insects are restricted to specific anatomical sites or cell types, and their abundance varies predictably with developmental age and sex of the insect host and environmental conditions. For microbes that can be either mutualistic or deleterious depending on environmental circumstance or host genotype, a key feature of the deleterious phenotype is high proliferation rates and abundance, often accompanied by an expanded distribution within the insect body. For example, the negative effect of the popcorn strain of Wolbachia on the longevity of Drosophila is strongly correlated with high bacterial numbers (McGraw et al., 2002); and the γ-proteobacterium Hamiltonella is harmful to its aphid host only on certain rearing plants, correlated with a 10-fold increase in bacterial abundance (Chandler et al., 2008).
Several studies indicate that the immune system plays a central role in the persistence of the microbiota. One such study concerns the gut microbiota of Drosophila melanogaster, dominated by Acetobacter and relatives, and Lactobacillus. The epithelial cells of the insect midgut bear receptors for the immune deficiency (IMD) signaling pathway, which is also active in other tissues, including the fat body, where it mediates the expression of various antimicrobial peptides (AMPs) against bacteria (Lemaitre and Hoffmann, 2007). The resident gut bacteria induce IMD signaling in the gut epithelial cells, resulting in localization of the NF-κB transcription factor Relish to the epithelial cell nucleus, but, contrary to expectation, the expression of AMPs is not induced (Ryu et al., 2008). The inhibition of AMP production is dependent on expression of the homeobox gene caudal (Figure A7-1A); when caudal expression is reduced by RNAi, AMP gene expression is upregulated. Importantly, the consequence of AMP production in the gut is not elimination of the gut microbiota, but a change in the microbial composition. Specifically, the relative abundance of two bacteria is dramatically altered; in the untreated fly, a bacterium of the Acetobacteriaceae (strain A911, known as Commensalibacter intestini) is dominant, and the abundance of Gluconobacter morbifer (G707) is very low. When the AMPs are expressed, the ratio of A911:G707 shifts from nearly 200:1 (favoring A911) to 1:20 (favoring
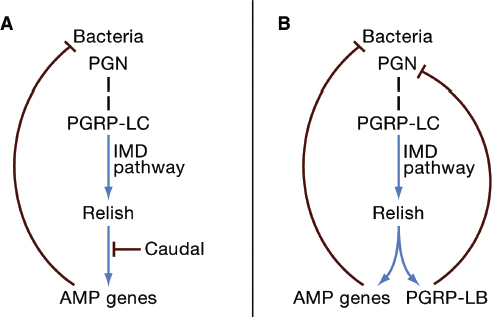
FIGURE A7-1 The insect IMD pathway and persistence of resident microorganisms. The IMD (Immune Deficiency) pathway is triggered by the binding of bacterial peptidoglycan (PGN) to the peptidoglycan recognition protein PGRP-LC (dashed vertical line). The resultant activation of the NF-κB transcription factor Relish leads to the upregulated expression of antimicrobial peptide (AMP) genes. Positive interactions (blue), negative interactions (red). (A) Resident microorganisms in the Drosophila gut activate the IMD pathway, but expression of AMPs is repressed by the transcription factor Caudal. (B) The IMD pathway in bacteriocytes (insect cells bearing symbiotic bacteria) of the Glossina tsetse fly and weevil Sitophilus is activated, but the immunoreactivity of bacteriocytes is dampened by the very high expression of the IMD-responsive gene PGRP-LB. PGRP-LB is a PGN-amidase that degrades the PGN ligand, thereby downregulating the IMD pathway.
EW707) (Roh et al., 2008; and Ryu et al., 2008). G. morbifer is deleterious to the host, resulting in severely depressed life spans of the flies with a G. morbifer-dominated gut community.
The implications are two-fold. First, AMPs are not exclusively antagonistic to microorganisms, but can also act to manage and regulate the microbial community. This conclusion has been reached independently for AMPs in other invertebrate animals (Fraune et al., 2010), and there is also evidence that other immune-related molecules can function in immunological management. For example, Toll-like receptor-2 (TLR2), which contributes to pathogen recognition in mammals, also suppresses the host inflammatory response to a major resident gut microorganism, Bacteroides fragilis; specifically, a product of B. fragilis (polysaccharide A) signals through TLR2 expressed by regulatory T cells to suppress
proinflammatory TH17 cells, thereby enabling B. fragilis to associate closely with the intestinal mucosa without inducing a host inflammatory response (Round et al., 2011). Second, although the great flexibility of the vertebrate adaptive immune system has been suggested to facilitate immunological management of the resident microbiota (McFall-Ngai, 2007), research on Drosophila demonstrates that the innate immune system can also function to regulate and manage the microbiota.
The study of Ryu et al. (2008) on the interaction between the gut microbiota and immune effectors of Drosophila focused exclusively on the immune responses localized to the gut. Data on Anopheles mosquitoes suggest that the gut microbiota induces a systemic immunological response that limits the abundance and distribution of the microorganisms. RNAi-mediated silencing of AMPs and immune signaling pathways has been shown to result in increased proliferation of the gut microbiota, including Pseudomonas and Novosphingobium species, and their localization to the hemolymph (Dong et al., 2009; and Garver et al., 2008). The effect of the gut microbiota on the systemic immune response of other insects has not been investigated.
Further evidence that the innate immune system can serve to promote coexistence between insects and their resident microbiota comes from several studies on intracellular bacteria in bacteriocytes. Although they have much-reduced genomes, these bacteria are predicted to possess the molecular patterns that are recognized by the insect immune system. The host immune response is attenuated in the bacteriocytes of both the weevil Sitophilus and tsetse fly Glossina. In both cases, PGRP-LB (a peptidoglycan recognition protein with amidase activity) is strongly expressed in the bacteriocytes. Its expression is dependent on activation of the IMD pathway, and it functions to remove the peptidoglycan ligand that triggers the IMD pathway (Figure A7-1B). In this way, the responsiveness of the IMD pathway to symbiont peptidoglycan fragments is dampened, promoting persistence of the bacteria. The symbioses in Sitophilus and Glossina have separate evolutionary origins, indicating that the central role of PGRP-LB in these symbioses has arisen independently. PGRP-LB is not, however, the universal arbiter of symbiont persistence in bacteriocytes. Aphids, for example, lack both PGRPs and a functional IMD pathway, and it has been suggested that the selection pressure to accommodate their bacteriocyte symbionts (Buchnera) may have led to the evolutionary loss of this portion of the immune system (Gerardo et al., 2010). Taken together, these data illustrate how selection by resident microorganisms can influence the immune responsiveness of specific organs and cell types and, potentially, of the entire animal. Although parasites have been invoked repeatedly as a selection pressure for diversification of the animal immune system (Rolff, 2007; and Sackton et al., 2007), beneficial symbioses can also be important.
A recent study on Wolbachia in the mosquito Aedes aegypti has implicated insect miRNAs (18- to 25-mer nonprotein-coding RNAs) in the regulation of symbiont numbers (Hussain et al., 2011). Specifically, when either insects or
cultured insect cells are infected by Wolbachia, the titer of one specific miRNA (aae-miR-2940) is elevated, resulting in the increased expression of an insect metalloprotease gene. When either the miRNA was inhibited or the metalloprotease gene expression silenced, Wolbachia numbers in the insect cells were reduced. Further research is required to establish the generality of miRNA-mediated promotion of insect-microbial coexistence and to determine how these mechanisms interact with the humoral immune system.
Overlaying these mechanisms acting in physiological time are selection pressures for coexistence operating in evolutionary time. In this regard, mode of transmission of the microorganisms plays a key role (Ewald, 1994). The selective interest of microorganisms that require a live host, and especially depend on a live host for transmission, overlaps with that of the host, and these microorganisms tend to be more benign that those that are transmitted efficiently from dead or dying hosts. Obligate vertical transmission generates an especially great overlap in selective interest between the microbial and host partners because the fitness of the microorganism is critically dependent on the fecundity of the host. Obligate vertical transmission is the norm in bacteriocyte symbioses of insects and is assured by behavioral mechanisms for various gut associations. The transmission of the γ-proteobacterium Ishikawaella in the distal midgut of various stinkbugs provides a particularly vivid example. The female insect deposits a fecal pellet containing Ishikawaella alongside an egg, the offspring feed on the pellet immediately after emerging from the egg, and the larval gut then undergoes dramatic changes involving the effective separation of the proximal blind-ended gut region for digestion and the distal symbiotic organ in the distal region (Fukatsu and Hosokawa, 2002; and Hosokawa et al., 2007).
Benefits of the Resident Microbiota to the Host
The contribution of the resident microbiota to the well-being of an animal, and the underlying mechanisms, can most readily be investigated by comparing animals experimentally deprived of their microbiota with untreated animals bearing their natural microbial complement. Useful supplementary approaches to identifying the contribution of specific microorganisms and the underlying mechanisms include the analysis of animals with modified microbial contents, e.g., associations with single taxa, microorganisms with known genetic mutations or microbiota of different host genotypes or species. These manipulations come under the umbrella term of “gnotobiotics,” meaning the rearing of animals under germ-free conditions, either continuously through life or with experimental administration of specific microorganisms. Gnotobiotics are especially important for investigating potentially important functions of microbial taxa that are at low abundance in unmanipulated associations. Various insect associations are very amenable to gnotobiotics because the treatments can be administered easily and cheaply to hundreds to thousands of individual insects. It is generally
straightforward to attribute function to individual microbial taxa because many insects bear a microbiota of low diversity (see above). The equivalent experiments using gnotobiotic mammals are technically demanding and, in some instances, extremely difficult to interpret because many microorganisms are members of complex, interdependent consortia with much functional redundancy, such that one host may bear multiple taxa with similar traits and the dominant taxa mediating a function of interest may vary among different host individuals.
Because of their experimental tractability, insect symbioses can provide valuable lessons about the services that resident microbes provide to animal hosts. Analyses of both gut and intracellular symbioses in many insects have revealed two core microbial functions: nutrition and defense. In many instances, it has been straightforward to link function to individual microbial taxa.
Particular insights are being obtained from studying insects that feed through the life cycle on diets of extremely unbalanced composition or low overall nutritional value, e.g., vertebrate blood (deficient in B vitamins), plant sap (for which essential amino acids are in short supply), and sound wood (grossly deficient in nitrogen and various essential nutrients). These insects all possess symbiotic microorganisms that provide specific nutrients in short dietary supply (Douglas, 2009). Strong indications that blood-feeding insects, including tsetse flies, lice, and bedbugs, derive B vitamins from their symbionts come from experiments comparing the performance of insects bearing and lacking their symbionts on blood diets that are untreated or supplemented with B vitamins. The same experimental approach, together with complementary metabolic analysis, has been applied to plant sap feeders, revealing that these symbionts provide essential amino acids.
The most detailed information is available for the association between the plant phloem sap-feeding pea aphid Acyrthosiphon pisum and its bacteriocyte symbiont Buchnera, for which both genomes are sequenced and annotated (International Aphid Genomics Consortium, 2010; and Shigenobu et al., 2000). Up to 50% of the essential amino acids synthesized by Buchnera cells are released to the surrounding host cell contents (Akman Gündüz and Douglas, 2009). The genome of this bacterium has a dearth of recognizable regulatory sequences, its gene expression is remarkably unresponsive to dietary perturbation (Reymond et al., 2006; and Wilson et al., 2006), and variation in aphid requirements for essential amino acids cannot generally be attributed to SNPs or other sequence variants in the Buchnera genome of the different aphids (MacDonald et al., 2011; and Vogel and Moran, 2011). Taken together, these data suggest that the insect host plays a major role in shaping the composition and quantity of essential amino acids released from Buchnera. Analysis of both the transcriptome and proteome of bacteriocytes has demonstrated that the bacteriocytes are enriched in enzymes mediating the synthesis of precursors required for Buchnera-mediated essential amino acid synthesis (Hansen and Moran, 2011; and Poliakov et al., 2011). Although the size and turnover of the precursor pools in the host cell
remain to be established, these data raise the possibility that Buchnera metabolism is poised for maximal production of essential amino acids and the realized synthesis rate is determined by precursor supply from the host cell.
Most research on the contribution of microorganisms resident in the gut to insect nutrition has focused on termites, which feed on various diets rich in plant fiber (wood, soil, humus). Historically—and erroneously—these associations were described as “miniature cows,” in which cellulose-rich plant material was degraded exclusively by the hindgut microbiota to short-chain fatty acids (SCFAs) that were utilized by the termite. It is now realized that the termites are more complex and diverse than originally envisaged. The guts of termites (unlike mammals) have considerable intrinsic cellulase activity, which is partly or entirely responsible for cellulose degradation, varying among termite groups (Watanabe and Tokuda, 2010). Members of the gut microbiota additionally contribute to the nitrogen economy of the insect by recycling insect waste nitrogen or fixing atmospheric nitrogen, again varying among termite taxa (Burnum et al., 2011; and Warnecke et al., 2007).
As the examples above illustrate, most research on the nutrition of insect-microbial symbioses has focused on animals with diets that are extremely nutrient-poor or nutritionally unbalanced and are not utilized generally by mammals. Even so, the nutritional significance of the resident gut microbiota in a few insects utilizing less extreme diets has been studied. Notably, elimination of the microbiota from Drosophila melanogaster has been reported to extend development time and shorten life span (Bakula, 1969; and Brummel et al., 2004). The indication that the latter effect appears to be diet-dependent (Ren et al., 2007) suggests that the microbiota may have a nutritional role in this insect, but this has not been demonstrated definitively.
There is unambiguous evidence that specific resident microorganisms promote the resistance of insects to certain natural enemies, including viruses, bacteria, fungi, nematodes, and parasitic wasps. Examples include: the α-proteobacterium Wolbachia, which protects Drosophila melanogaster against various viruses (Hedges et al., 2008); Spiroplasma bacteria, which confer resistance in Drosophila neotestacea against the nematode parasite Howardula aoronymphium (Jaenike et al., 2010) and in Drosophila hydei against the parasitic wasp Leptopilina heterotoma (Xie et al., 2010); and the γ-proteobacterium Regiella insecticola, which reduces the mortality of pea aphids infected with entomopathogenic fungi (Scarborough et al., 2005). In these and many other interactions, the underlying mechanisms are not known, but for a few instances an outline understanding of the mechanisms has been obtained. The following three examples illustrate the diversity of interactions mediated by defensive microbes.
The first example concerns actinobacteria with protective function. Beewolf digger wasps house cultures of actinobacteria “Candidatus Streptomyces philanthi” in antennal glands and smear the bacteria onto cocoons deposited in their humid, microbe-rich burrows (Kaltenpoth et al., 2005). The complex mix of
antibiotics synthesized by the bacteria confers a generalized protection against microbial attack (Kroiss et al., 2010). The thorax of leafcutting ants is enveloped in a multispecies actinobacterial mat (Mueller et al., 2008) that produces antibiotics with activity against fungi, including a major fungal pathogen, Escovopsis (Oh et al., 2009). Actinobacteria in a similar relationship with Dendronoctus beetles provide analogous protection (Scott et al., 2008). In each of these systems, the low diversity of the actinobacterial partners has facilitated the identification of the actinobacterial partner, its function, and the chemical nature of its antimicrobials.
The second example relates to parasitic wasps, many of which exploit various insects by depositing an egg in the body cavity of their victim. The resultant wasp larva consumes tissues of the living insect over some days, and following the death of the insect, the adult wasp emerges from the dead insect “mummy.” The resistance of pea aphids to the parasitic wasp Aphidius ervi is heightened by the presence of the γ-proteobacterium Hamiltonella defensa, which occurs in the body cavity of some pea aphids (the prevalence of H. defensa varies widely among different pea aphid populations). Importantly, this defensive role is evident only for H. defensa isolates that contain the virus APSE bearing the genes for toxins, including homologs of cytolethal toxins (Oliver et al., 2009). The reasonable inference is that resistance against the parasitic wasp is mediated by these toxins, but neither this interpretation nor how the aphid tissues avoid toxin-incurred damage has been demonstrated.
The resident microbiota in Anopheles mosquitoes reduces infection of the mosquitoes by the malaria parasite Plasmodium ingested with the blood meal. Specifically, the invasion of the midgut epithelium by the Plasmodium ookinetes is inhibited. This effect is mediated by bacteria established in the gut, and cofeeding Anopheles with bacteria and Plasmodium or coinjecting bacteria into the insect body cavity with Plasmodium feeding mediates these effects, indicating that it is not caused by a direct interaction between the bacteria and Plasmodium (Dong et al., 2009). Instead, the insect immune system is involved, including immune effectors that are active against both bacteria and Plasmodium. The precise mechanism of protection is not fully established (Cirimotich et al., 2010). After feeding on blood, the mosquito may mount a vigorous immune response against its resident bacteria, which proliferate rapidly when blood is ingested, or against hemozoin, which is an immunogenic breakdown product of the blood with an indirect effect on the Plasmodium. Alternatively or additionally, the mechanical disruption of the midgut epithelium by the invading Plasmodium ookinetes may introduce bacteria into the insect blood, triggering a systemic immune response. There is evidence for some specificity in this effect, with gram-negative bacteria providing greater protection than gram-positives (Cirimotich et al., 2010).
Dependence and Addiction
An important lesson from insect symbioses comes from the parasitic wasp Asobara tabida. In nature, every individual of this insect bears the bacterium Wolbachia, which is transmitted vertically via the eggs of the wasp. When Wolbachia is eliminated by antibiotic treatment, A. tabida is reproductively sterile with egg production halted by massive apoptosis of the nurse cells in the ovary. A limited incidence of apoptosis occurs during normal oogenesis in A. tabida bearing Wolbachia. A reasonable inference from these data is that Wolbachia inhibits apoptosis, and there is strong correlative evidence that Wolbachia perturbs iron metabolism in its host, resulting in increased oxidative stress that disrupts cellular physiology, including apoptosis (Kremer et al., 2009). This interaction has been argued to select for increased apoptotic signaling in the host to compensate for the otherwise harmful effects of Wolbachiaon host physiology (Pannebakker et al., 2007). The putative compensatory response of A. tabida is constitutive, such that apoptosis is excessive in the absence of the inhibitory signal from Wolbachia. In this way, A. tabida has become dependent on Wolbachia without deriving any discernible benefit, a condition that is defined as addiction (Aanen and Hoekstra, 2007).
Dependence without benefit (i.e., addiction) has received little attention, partly because dependence is often interpreted as evidence for benefit, even in the absence of evidence for any service (e.g., nutrient provisioning, defensive role), but also because interactions between animal hosts and their resident microbiota are often investigated without considering their evolutionary context. Even so, there are candidate instances of dependence without benefit in various symbioses, including those between mammals and their resident microbiota. As one specific example, the capillary network in the small intestine of the mouse develops normally only in the presence of the gut microbiota (Stappenbeck et al., 2002). When mice are reared under aseptic conditions, the small intestine is poorly vascularized and the mice have reduced capacity to assimilate nutrients from the gut. Capillary growth is not induced by the bacteria directly, but by antibacterial peptides produced by the Paneth cells of the intestine in response to the presence of the microorganisms. In this way, the bacteria have become integrated into the animal signaling pathway required for normal development of the intestinal capillary bed, even though the capillaries function independently of the microbiota. There is no selection on the host to be independent of the microbiota because they are invariably infected by gut microorganisms soon after birth. Further candidate instances of dependence without benefit come from the central role of the gut microbiota in the development of the gut-associated lymph tissue and various populations of intestinal immune cells (Duan et al., 2010; Falk et al., 1998; Niess et al., 2008; and Pollard and Sharon, 1970).
How can dependence without benefit evolve? One possibility is that these many instances are animal compensatory responses to microbial perturbations of the host immune system or metabolism. As in A. tabida, the host responses may
be constitutive because there is no direct selection for function in the absence of the microbiota (because the microbiota are never absent under natural conditions). An alternative scenario is that certain signaling networks are truly symbiotic: that the resident microbiota in the ancestors of modern animals may have provided elements of the signaling networks regulating developmental processes, including angiogenesis and immune function, and that these elements have been retained in modern animals (Lee and Mazmanian, 2010; and McFall-Ngai, 2002). Whatever the evolutionary origin of dependence, the consequent addiction of animals for their resident microbiota is evident only under the unnatural experimental conditions of axenic (germ-free) rearings or large-scale perturbation of the microbiota, as in various immunological diseases or grossly unsuitable diets.
Coevolution Between Insects and Their Microbiota
Insect symbioses offer spectacular examples of coevolution with major consequences for the health and well-being of the animal host. For example, coevolutionary changes may include increased microbial production of nutrients valuable to the host and correlated changes in host metabolism, transporters, etc., that promote the host processing of microbial nutrients.
Coevolutionary interactions have been inferred from genome sequencing projects on plant sap-feeding insects and their microbial symbionts, including several instances of perfect complementarity in amino acid metabolism through differential gene loss in the partners in the association. For example, the genomes of both the pea aphid Acyrthosiphon pisum and its Buchnera symbiont have been sequenced (International Aphid Genomics Consortium, 2010; and Shigenobu et al., 2000), revealing a large-scale loss in the capacity of Buchnera to synthesize nonessential amino acids (which can be synthesized by the aphid host) and loss of the capacity to synthesize one amino acid, arginine, in the aphid host (Figure A7-2A). Unlike aphids, which have a single obligate symbiont, plant sap-feeding insects of a related suborder (the Auchenorrhyncha) possess dual symbioses, comprising a Bacteroidetes Sulcia muelleri and a second auxiliary bacterium, the identity of which varies with the insect group (Figure A7-2B). Remarkably, Sulcia has the capacity to synthesize 7 or 8 of the 10 essential amino acids (varying with insect group), and the auxiliary symbiont can synthesize the remaining essential amino acids (McCutcheon et al., 2009; McCutcheon and Moran, 2007; and McCutcheon and Moran, 2010). As a consequence, each symbiont supplies the required nutrients for both the other bacterium and the host (Figure A7-2B).
The “perfect” metabolic complementarity in these insect symbioses is a product of the reciprocal coevolutionary changes in the host and symbiont(s), accompanied by genome reduction of the symbionts. Both of these latter traits are facilitated by the persistent obligate vertical transmission of the symbionts via the egg, for example, over an estimated 160 million years for Buchnera and
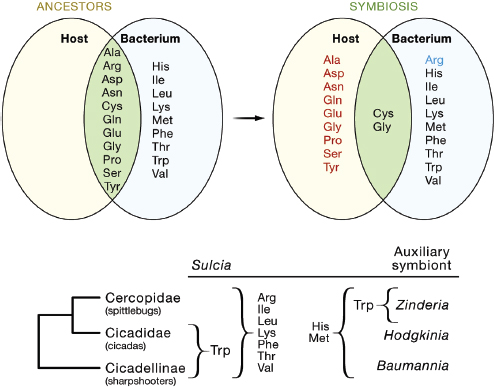
FIGURE A7-2 Coevolution of amino acid biosynthesis in plant sap–feeding insects and their bacterial symbionts. (A) In the symbiosis between the pea aphid and Buchnera bacteria, the Buchnera (but not aphid) has lost the capacity to synthesize eight amino acids (red), and the aphid (but not Buchnera) has lost the capacity to synthesize arginine (blue) relative to the ancestral condition (E. coli, a relative of Buchnera, and a generalized insect, respectively). The one-reaction synthesis of glycine from serine is retained by both partners. Cysteine is synthesized by different pathways in the two partners (from methionine in the aphid, from serine in Buchnera). (B) Spittlebugs, cicadas, and sharpshooters bear two bacterial symbionts, Sulcia and an auxiliary symbiont with complementary amino acid biosynthetic capabilities. The amino acids produced by each bacterium are made available to both the insect host and the alternative bacterium. Both bacteria require the ten nonessential amino acids from the host.
Note: ala: alanine; arg: arginine; asn: asparagine; asp: aspartate; cys: cysteine; gln: glutamaine; glu: glutamate; gly: glycine; his: histidine; ile: isoleucine; leu: leucine; lys: lysine; met: methionine; phe: phenylalanine; pro: proline; ser: serine; thr: threonine; trp: tryptophan; tyr: tyrosine; val: valine.
260 million years for Sulcia (Moran et al., 1993; and Moran et al., 2005). The continuous interaction between individual lineages of host and symbiont provides the basis for both genome reduction of the symbiont (by relaxed selection and genome deterioration) and strict reciprocal coevolution.
These associations provide an important lesson in the power of coevolutionary interactions in shaping the functional traits of animals and their resident
microbes. Nevertheless, we should not necessarily anticipate precisely equivalent relationships in mammals because no resident microorganisms of mammals are known to be obligately vertically transmitted and the coevolutionary interactions are more diffuse, i.e., likely to involve guilds of microbes (Oh et al., 2010).
Current and Future Lessons from Insect Symbioses
Insect symbioses offer various clear-cut exemplars of processes underlying interactions between animals and their resident microbiota. Most of these interactions are unlikely to be replicated precisely in mammalian systems, simply because symbioses in mammals and insects are different. Their principal value to researchers investigating mammalian systems is conceptual, demonstrating, for example, how the persistence of microorganisms can be shaped by modulation of the innate immune system (Figure A7-1) or how the metabolic capabilities of hosts and multiple microorganisms can coevolve to perfect complementarity (Figure A7-2). The low diversity of microorganisms in most insects, compared to mammals, is important for the clarity of these exemplars. For insects, it is generally possible to couple microbial function to one (or several) well-defined microbial taxa, and functional redundancy is minimal.
The key traits of low diversity and minimal functional redundancy in the microbiota of many insects provide the opportunity to exploit insect systems to investigate fundamental problems in animal-microbial interactions that are impractical or unfeasible to investigate in mammalian systems. From insect-based research, specific hypotheses can be generated for subsequent analysis in biomedical rodent models or in human trials. The opportunities relate especially to the link between the gut microbiota and various human diseases, including metabolic syndrome (obesity, type 2 diabetes, and cardiovascular disease), Crohn’s disease, and related immunological dysfunction (Lee and Mazmanian, 2010; and Wang et al., 2011). The causal networks underpinning these diseases are difficult to disentangle because the mammalian microbiota is complex and variable. Various insect systems have potential value. Drosophila melanogaster offers a particularly strong model because the low diversity of its gut microbiota (Cox and Gilmore, 2007; Roh et al., 2008; and Wong et al., 2011) can be harnessed with the unparalleled set of Drosophila genetic resources and tools. Insect symbioses can certainly provide many valuable lessons to be learned, but their greatest value is not yet realized. We should anticipate a shift in the curriculum toward the explicit exploitation of insect symbioses as models for biomedical research on the role of the resident microbiota in health and disease.
Acknowledgments
This work was supported by NIH grant 1R01GM095372-01, NSF grant IOS-0919765, and the Sarkaria Institute for Insect Physiology and Toxicology.
References
Aanen, D.K., and Hoekstra, R.F. (2007). The evolution of obligate mutualism: if you can’t beat ‘em, join ’em. Trends Ecol. Evol. (Amst.) 22, 506–509.
Akman Gündüz, E., and Douglas, A.E. (2009). Symbiotic bacteria enable insect to use a nutritionally inadequate diet. Proc. Biol. Sci. 276, 987–991.
Bakula, M. (1969). The persistence of a microbial flora during postembryogenesis of Drosophila melanogaster. J. Invertebr. Pathol. 14, 365–374.
Brugerolle, G., and Radek, R. (2006). Symbiotic protozoa of termites. In Intestinal Microorganisms of Soil Invertebrates, H. König and A. Varma, eds. (Berlin: Springer-Verlag), pp. 243–269.
Brummel, T., Ching, A., Seroude, L., Simon, A.F., and Benzer, S. (2004). Drosophila lifespan enhancement by exogenous bacteria. Proc. Natl. Acad. Sci. USA 101, 12974–12979.
Burnum, K.E., Callister, S.J., Nicora, C.D., Purvine, S.O., Hugenholtz, P., Warnecke, F., Scheffrahn, R.H., Smith, R.D., and Lipton, M.S. (2011). Proteome insights into the symbiotic relationship between a captive colony of Nasutitermes corniger and its hindgut microbiome. ISME J. 5, 161–164.
Chandler, S.M., Wilkinson, T.L., and Douglas, A.E. (2008). Impact of plant nutrients on the relationship between a herbivorous insect and its symbiotic bacteria. Proc. Biol. Sci. 275, 565–570.
Cirimotich, C.M., Dong, Y., Garver, L.S., Sim, S., and Dimopoulos, G. (2010). Mosquito immune defenses against Plasmodium infection. Dev. Comp. Immunol. 34, 387–395.
Connell, J.H. (1978). Diversity in tropical rain forests and coral reefs. Science 199, 1302–1310.
Cox, C.R., and Gilmore, M.S. (2007). Native microbial colonization of Drosophila melanogaster and its use as a model of Enterococcus faecalis pathogenesis. Infect. Immun. 75, 1565–1576.
Dale, C., and Moran, N.A. (2006). Molecular interactions between bacterial symbionts and their hosts. Cell 126, 453–465.
Dethlefsen, L., McFall-Ngai, M., and Relman, D.A. (2007). An ecological and evolutionary perspective on human-microbe mutualism and disease. Nature 449, 811–818.
Dillon, R.J., and Dillon, V.M. (2004). The gut bacteria of insects: nonpathogenic interactions. Annu. Rev. Entomol. 49, 71–92.
Dong, Y., Manfredini, F., and Dimopoulos, G. (2009). Implication of the mosquito midgut microbiota in the defense against malaria parasites. PLoS Pathog. 5, e1000423.
Douglas, A.E. (2007). Symbiotic microorganisms: untapped resources for insect pest control. Trends Biotechnol. 25, 338–342.
Douglas, A.E. (2009). The microbial dimension in insect nutritional ecology. Funct. Ecol. 23, 38–47.
Duan, J., Chung, H., Troy, E., and Kasper, D.L. (2010). Microbial colonization drives expansion of IL-1 receptor 1-expressing and IL-17-producing gamma/delta T cells. Cell Host Microbe 7, 140–150.
Ewald, P.W. (1994). Evolution of Infectious Diseases (Oxford, UK: Oxford University Press).
Falk, P.G., Hooper, L.V., Midtvedt, T., and Gordon, J.I. (1998). Creating and maintaining the gastrointestinal ecosystem: what we know and need to know from gnotobiology. Microbiol. Mol. Biol. Rev. 62, 1157–1170.
Fraune, S., Augustin, R., Anton-Erxleben, F., Wittlieb, J., Gelhaus, C., Klimovich, V.B., Samoilovich, M.P., and Bosch, T.C. (2010). In an early branching metazoan, bacterial colonization of the embryo is controlled by maternal antimicrobial peptides. Proc. Natl. Acad. Sci. USA 107, 18067–18072.
Fukatsu, T., and Hosokawa, T. (2002). Capsule-transmitted gut symbiotic bacterium of the Japanese common plataspid stinkbug, Megacopta punctatissima. Appl. Environ. Microbiol. 68, 389–396.
Garver, L.S., Xi, Z., and Dimopoulos, G. (2008). Immunoglobulin superfamily members play an important role in the mosquito immune system. Dev. Comp. Immunol. 32, 519–531.
Gerardo, N.M., Altincicek, B., Anselme, C., Atamian, H., Barribeau, S.M., de Vos, M., Duncan, E.J., Evans, J.D., Gabaldón, T., Ghanim, M., et al. (2010). Immunity and other defenses in pea aphids, Acyrthosiphon pisum. Genome Biol. 11, R21.
Hansen, A.K., and Moran, N.A. (2011). Aphid genome expression reveals hostsymbiont cooperation in the production of amino acids. Proc. Natl. Acad. Sci. USA 108, 2849–2854.
Hedges, L.M., Brownlie, J.C., O’Neill, S.L., and Johnson, K.N. (2008). Wolbachia and virus protection in insects. Science 322, 702.
Hosokawa, T., Kikuchi, Y., and Fukatsu, T. (2007). How many symbionts are provided by mothers, acquired by offspring, and needed for successful vertical transmission in an obligate insect-bacterium mutualism? Mol. Ecol. 16, 5316–5325.
Hussain, M., Frentiu, F.D., Moreira, L.A., O’Neill, S.L., and Asgari, S. (2011). Wolbachia uses host microRNAs to manipulate host gene expression and facilitate colonization of the dengue vector Aedes aegypti. Proc. Natl. Acad. Sci. USA 108, 9250–9255.
International Aphid Genomics Consortium. (2010). Genome sequence of the pea aphid Acyrthosiphon pisum. PLoS Biol. 8, e1000313.
Jaenike, J., Unckless, R., Cockburn, S.N., Boelio, L.M., and Perlman, S.J. (2010). Adaptation via symbiosis: recent spread of a Drosophila defensive symbiont. Science 329, 212–215.
Kaltenpoth, M., Göttler, W., Herzner, G., and Strohm, E. (2005). Symbiotic bacteria protect wasp larvae from fungal infestation. Curr. Biol. 15, 475–479.
Kerney, R., Kim, E., Hangarter, R.P., Heiss, A.A., Bishop, C.D., and Hall, B.K. (2011). Intracellular invasion of green algae in a salamander host. Proc. Natl.Acad. Sci. USA 108, 6497–6502.
Kremer, N., Voronin, D., Charif, D., Mavingui, P., Mollereau, B., and Vavre, F. (2009). Wolbachia interferes with ferritin expression and iron metabolism ininsects. PLoS Pathog. 5, e1000630.
Kroiss, J., Kaltenpoth, M., Schneider, B., Schwinger, M.G., Hertweck, C., Maddula, R.K., Strohm, E., and Svatos, A. (2010). Symbiotic Streptomycetes provide antibiotic combination prophylaxis for wasp offspring. Nat. Chem.Biol. 6, 261–263.
Lee, Y.K., and Mazmanian, S.K. (2010). Has the microbiota played a critical role in the evolution of the adaptive immune system? Science 330, 1768–1773.
Lemaitre, B., and Hoffmann, J. (2007). The host defense of Drosophila melanogaster. Annu. Rev. Immunol. 25, 697–743.
MacDonald, S.J., Thomas, G.H., and Douglas, A.E. (2011). Genetic and metabolic determinants of nutritional phenotype in an insect-bacterial symbiosis. Mol. Ecol. 20, 2073–2084.
McCutcheon, J.P., and Moran, N.A. (2007). Parallel genomic evolution and metabolic interdependence in an ancient symbiosis. Proc. Natl. Acad. Sci. USA 104, 19392–19397.
McCutcheon, J.P., and Moran, N.A. (2010). Functional convergence in reduced genomes of bacterial symbionts spanning 200 My of evolution. Genome Biol Evol 2, 708–718.
McCutcheon, J.P., McDonald, B.R., and Moran, N.A. (2009). Convergent evolution of metabolic roles in bacterial co-symbionts of insects. Proc. Natl. Acad. Sci. USA 106, 15394–15399.
McFall-Ngai, M.J. (2002). Unseen forces: the influence of bacteria on animal development. Dev. Biol. 242, 1–14.
McFall-Ngai, M. (2007). Adaptive immunity: care for the community. Nature 445, 153.
McGraw, E.A., Merritt, D.J., Droller, J.N., and O’Neill, S.L. (2002). Wolbachia density and virulence attenuation after transfer into a novel host. Proc. Natl. Acad. Sci. USA 99, 2918–2923.
Morales-Jiménez, J., Zùñiga, G., Villa-Tanaca, L., and Hernández-Rodríguez, C. (2009). Bacterial community and nitrogen fixation in the red turpentine beetle, Dendroctonus valens LeConte (Coleoptera: Curculionidae: Scolytinae). Microb. Ecol. 58, 879–891.
Moran, N.A., Munson, M.A., Baumann, P., and Ishikawa, H. (1993). A molecular clock in endosymbiotic bacteria is calibrated using the insect hosts. Proc. Biol. Sci. 253, 167–171.
Moran, N.A., Tran, P., and Gerardo, N.M. (2005). Symbiosis and insect diversification: an ancient symbiont of sap-feeding insects from the bacterial phylum Bacteroidetes. Appl. Environ. Microbiol. 71, 8802–8810.
Mueller, U.G., Dash, D., Rabeling, C., and Rodrigues, A. (2008). Coevolution between attine ants and actinomycete bacteria: a reevaluation. Evolution 62, 2894–2912.
Nemergut, D.R., Costello, E.K., Hamady, M., Lozupone, C., Jiang, L., Schmidt, S.K., Fierer, N., Townsend, A.R., Cleveland, C.C., Stanish, L., and Knight, R. (2011). Global patterns in the biogeography of bacterial taxa. Environ. Microbiol. 13, 135–144.
Niess, J.H., Leithäuser, F., Adler, G., and Reimann, J. (2008). Commensal gut flora drives the expansion of proinflammatory CD4 T cells in the colonic lamina propria under normal and inflammatory conditions. J. Immunol. 180, 559–568.
Oh, D.C., Poulsen, M., Currie, C.R., and Clardy, J. (2009). Dentigerumycin: a bacterial mediator of an ant-fungus symbiosis. Nat. Chem. Biol. 5, 391–393.
Oh, P.L., Benson, A.K., Peterson, D.A., Patil, P.B., Moriyama, E.N., Roos, S., and Walter, J. (2010). Diversification of the gut symbiont Lactobacillus reuteri as a result of host-driven evolution. ISME J. 4, 377–387.
Oliver, K.M., Degnan, P.H., Hunter, M.S., and Moran, N.A. (2009). Bacteriophages encode factors required for protection in a symbiotic mutualism. Science 325, 992–994.
Oliver, K.M., Degnan, P.H., Burke, G.R., and Moran, N.A. (2010). Facultative symbionts in aphids and the horizontal transfer of ecologically important traits. Annu. Rev. Entomol. 55, 247–266.
Pannebakker, B.A., Loppin, B., Elemans, C.P., Humblot, L., and Vavre, F. (2007). Parasitic inhibition of cell death facilitates symbiosis. Proc. Natl. Acad. Sci. USA 104, 213–215.
Poliakov, A., Russell, C.W., Ponnala, L., Hoops, H.J., Sun, Q., Douglas, A.E., and van Wijk, K.J. (2011). Large-scale label-free quantitative proteomics of the pea aphid-Buchnera symbiosis. Mol. Cell Proteomics 10, M110.007039.
Pollard, M., and Sharon, N. (1970). Responses of the Peyer’s Patches in Germ-Free Mice to Antigenic Stimulation. Infect. Immun. 2, 96–100.
Ren, C., Webster, P., Finkel, S.E., and Tower, J. (2007). Increased internal and external bacterial load during Drosophila aging without life-span trade-off. Cell Metab. 6, 144–152.
Reymond, N., Calevro, F., Viñuelas, J., Morin, N., Rahbé, Y., Febvay, G., Laugier, C., Douglas, A.E., Fayard, J.-M., and Charles, H. (2006). Different levels of transcriptional regulation due to trophic constraints in the reduced genome of Buchnera aphidicola APS. Appl. Environ. Microbiol. 72, 7760–7766.
Robinson, C.J., Schloss, P., Ramos, Y., Raffa, K., and Handelsman, J. (2010). Robustness of the bacterial community in the cabbage white butterfly larval midgut. Microb. Ecol. 59, 199–211.
Roh, S.W., Nam, Y.D., Chang, H.W., Kim, K.H., Kim, M.S., Ryu, J.H., Kim, S.H., Lee, W.J., and Bae, J.W. (2008). Phylogenetic characterization of two novel commensal bacteria involved with innate immune homeostasis in Drosophila melanogaster. Appl. Environ. Microbiol. 74, 6171–6177.
Rolff, J. (2007). Why did the acquired immune system of vertebrates evolve? Dev. Comp. Immunol. 31, 476–482.
Round, J.L., Lee, S.M., Li, J., Tran, G., Jabri, B., Chatila, T.A., and Mazmanian, S.K. (2011). The Toll-like receptor 2 pathway establishes colonization by a commensal of the human microbiota. Science 332, 974–977.
Ryu, J.H., Kim, S.H., Lee, H.Y., Bai, J.Y., Nam, Y.D., Bae, J.W., Lee, D.G., Shin, S.C., Ha, E.M., and Lee, W.J. (2008). Innate immune homeostasis by the homeobox gene caudal and commensal-gut mutualism in Drosophila. Science 319, 777–782.
Sackton, T.B., Lazzaro, B.P., Schlenke, T.A., Evans, J.D., Hultmark, D., and Clark, A.G. (2007). Dynamic evolution of the innate immune system in Drosophila. Nat. Genet. 39, 1461–1468.
Scarborough, C.L., Ferrari, J., and Godfray, H.C. (2005). Aphid protected from pathogen by endosymbiont. Science 310, 1781.
Scott, J.J., Oh, D.C., Yuceer, M.C., Klepzig, K.D., Clardy, J., and Currie, C.R. (2008). Bacterial protection of beetle-fungus mutualism. Science 322, 63.
Shanbhag, S., and Tripathi, S. (2009). Epithelial ultrastructure and cellular mechanisms of acid and base transport in the Drosophila midgut. J. Exp. Biol. 212, 1731–1744.
Shigenobu, S., Watanabe, H., Hattori, M., Sakaki, Y., and Ishikawa, H. (2000). Genome sequence of the endocellular bacterial symbiont of aphids Buchnera sp. APS. Nature 407, 81–86.

















