
11
Justice- and Fairness-Related Behaviors in Nonhuman Primates
SARAH F. BROSNAN
A distinctive feature across human societies is our interest in justice and fairness. People will sometimes invest in extremely costly behavior to achieve fair outcomes for themselves and others. Why do people care so much about justice? One way to address this is comparatively, exploring behaviors related to justice and fairness in other species. In this chapter, I review work exploring responses to inequity, prosocial behavior, and other relevant behaviors in nonhuman primates in an effort to understand both the potential evolutionary function of these behaviors and the social and ecological reasons for the individual differences in behavior. I also consider how these behaviors relate to human behavior, particularly in the case of experimental studies using games derived from experimental economics to compare nonhuman primates’ responses to those of humans in similar experimental conditions. These results emphasize the importance of a comparative approach to better understand the function and diversity of human behavior.
Afeature of humans that apparently sets us apart as a species is our focus on others’ well-being, as well as our own (Eisenberg and Mussen 1989; Henrich et al. 2004). We will sometimes at
_____________
Georgia State University, Departments of Psychology and Philosophy, Neuroscience Institute, Atlanta, GA 30302. E-mail: sbrosnan@gsu.edu.
great personal cost, protest outcomes that we consider unfair, whether they personally affect us or have no direct impact on our lives. What is it that causes people to behave in this way? One possibility is that it is attributable to culture. In fact, only English has a word for the concept of “fairness”; other languages have adopted the English word, indicating a culturally broad interest in the concept but potentially implying that the concept is not ubiquitous (Wilson, 2012). Another possibility is that we have evolved to be interested in the well-being of others. Although there are clear fitness benefits to considering one’s own well-being, there are also (more delayed) benefits to considering others’ well-being. There may be costs to helping others in the short term, but if there are long-term benefits, either because of continued interactions based on reciprocity or, more indirectly, because of gaining a reputation as a “fair” person, then the behavior could be selected (Frank, 1988). If this behavior has evolved, we may find similar behaviors, or precursor behaviors, in other species. Other species will not show these behaviors in the same way that humans do. Among other reasons for this, many behaviors related to fairness and justice, such as the ability to share information about a third party’s previous behavior, require language. Nonetheless, an important question is the degree to which fairness and justice have evolved from similar or related behaviors in other species. This evolutionary approach provides insight into understanding our own behavior.
In humans, the concept of fairness is closely related to that of justice. Distinguishing the two, fairness involves voluntary interactions with other individuals, whereas justice is meted out by an impartial third party (Wilson, 2012). Based on these definitions, most of the experimental work discussed below relates to fairness, rather than justice, because the subjects being studied are involved in the interactions and outcomes in question. Outside of the experimental context, observations suggest impartial interventions in others’ interactions, indicating that these species may also be sensitive to justice concerns. There are many different aspects of justice and fairness, including distributive, procedural, retributive, restorative, etc. Most experimental studies involving nonhuman species measure subjects’ responses to different distributions of food rewards and, hence, concern distributive justice or fairness. In humans, two common distribution mechanisms are egalitarianism, in which all individuals get the same outcome (e.g., equality) and equity, in which individuals’ benefits are proportional to their inputs. Both of these are amenable to study in other species and are the feature most commonly manipulated in the aforementioned experimental studies (e.g., by altering subjects’ payoffs compared with their partner’s). Outside of the experimental realm, observational studies indicate that nonhuman primates are also sensitive to nondistributional aspects of justice and fairness, such as judging the appropriateness
of another’s behavior, providing a broader context in which to consider justice or fairness in these species.
Studying other species’ behaviors in these situations allows us to elucidate the evolutionary function of justice and fairness by exploring behaviors that are related, either because the behavior itself is a component of justice or fairness or because it represents a precursor to human behavior. This comparative approach is informative in two complementary ways, through homology and convergence. Homologies occur when a behavior exists in multiple species because the behavior existed in a shared common ancestor. This pinpoints when in the taxon’s phylogenetic history the trait evolved. Convergences occur when a behavior evolved in multiple species because of common selective pressures but without shared descent from a common ancestor. Convergences may be particularly informative when studying the function of a behavior because, through comparison across species, they may indicate which traits co-occur and the social or ecological pressures that may have selected for the behavior.
My goal in this review is to bring together data regarding behaviors related to justice and fairness in nonhuman primate species (hereafter, primates). Observations have highlighted situations in which primates act as if their behavior is guided by an interest in the outcomes of others (de Waal, 1991). A decade ago, the first experimental studies emerged, focusing on one aspect of justice and fairness: how individuals responded to getting less than a partner (Brosnan and de Waal, 2003; Brosnan et al., 2005). These studies used methods derived from game theory and showed that, in experimental settings in which inequity between two individuals was introduced, subjects responded negatively to inequity not in their favor. Since that time, this task has been extended to 14 species, including 10 primate species, allowing for a broad phylogenetic view that provides insight into the evolutionary function of inequity (Brosnan, 2011). Additionally, responses to inequity have been explored in more species-specific contexts and situations (de Waal and Berger, 2000; Brosnan et al., 2006). These results highlight the correlation between inequity and cooperation, as well as emphasizing the socioecological factors that influenced the emergence of justice- and fairness-related behavior. This comparative analysis is beginning to shed light on the factors that may have led to the evolution of justice- and fairness-related behaviors in humans and other primates.
JUSTICE- AND FAIRNESS-RELATED BEHAVIOR IN OTHER SPECIES
One hypothesis for the evolution of fairness is that recognizing inequities helped individuals ascertain the value of their partners in cooperative
interactions (Fehr and Schmidt, 1999; Brosnan, 2011). Individuals should not continue to work with others if they do not, on average, benefit from the relationship. Whether outcomes are equitable may be used as a proxy; if one’s outcomes are routinely wanting compared with one’s partner’s, then it is likely that an interaction with another partner, even one chosen at random, will lead to a more beneficial outcome. This leads to a partner choice mechanism in which individuals sample other partners when they are experiencing high levels of inequity with the current partner or partners (Brosnan, 2006). Note that individuals thus recognize and respond to inequity without any understanding of why they are doing so. Additionally, this provides a benefit even if the individual’s outcomes in the less advantageous interaction are net positive, because the currency of natural selection is relative, rather than absolute, gains.
Although this hypothesis linking inequity and cooperation emerged in the context of humans, other species also cooperate with one another, so behaviors related to inequity may occur in other cooperative species as well. Comparative research presents challenges. First, the manifestations of justice and fairness may be different in different species, making it difficult for human experimenters to recognize relevant behaviors. Second, nonhuman species are nonverbal, with the dual implications that we cannot ask them about their motivations for their behavior, nor can we ask them about outcomes that did not result in an overt change in their behavior. Thus, we will miss any response that does not have a behavioral manifestation. As a result, experiments focus on whether other species act in a certain way, not whether they mean to act in that way. This is different from the measure used in many human studies, and ideally results should only be compared if the variable of interest is subjects’ behavioral reactions, not subjects’ reported motivations or reactions. Finally, the limitation to measuring behavioral changes means that the methods used in humans typically differ substantially from those used in other primates, which can complicate comparative analyses.
There have been two approaches to comparative work on justice and fairness. The first is to adapt game theory methods to explore decision making in other species. Game theory is a powerful approach that allows complex decision making to be modeled experimentally as a series of carefully controlled choices, which can be experimentally manipulated to rigorously explore the behavior of interest (Smith, 1987). The advantage of game theory for comparative research is the focus on very simple choices to uncover complex behavior, making these techniques amenable to use in a wide variety of other species. Procedures can be designed that require no verbal instruction whatsoever and that can be used in the same form in humans and other species, allowing for direct comparisons and subsequent explorations of shared outcomes or cognitive mechanisms
(Brosnan et al., 2013). Although we cannot assume that different species (or different individuals within the same species) interpret even identical procedures in the same way, if procedures differ between species, then we are never able to disentangle whether any resulting differences in behavior are attributable to species differences or differences in the procedure. One procedural challenge to such experimental approaches is that humans interact with an experimenter of the same species, whereas other species interact with an experimenter of a different species (a human). Although this is a practical necessity, other species did not evolve to interact with humans, and so careful controls are needed to determine that the nonhuman subjects were evaluating their outcomes relative to their conspecific partner, not the experimenter or another aspect of the environment. Additionally, experimental approaches are artificial, and so we may miss situations in which justice-related behavior occurs. Despite these drawbacks, game theoretic approaches have the advantage of being comparable to the human literature and so are indispensable for comparative research.
A second approach is to explore behaviors that may be related to justice and fairness in species-specific contexts. This includes observational studies, in which behavior is observed during unconstrained interactions. This approach has the advantage of being the more natural, which leads to likely candidate behaviors and a better understanding of how justice or fairness manifest in the species’ typical interactions. Moreover, because these studies are not reliant on manipulating outcomes and measuring subjects’ subsequent behavior, there is greater opportunity to study justice outside of the context of food distributions. The disadvantages are that these results are less comparable across species, particularly if methods or the contexts in which individuals are attentive to inequity differ, and the underlying motivations for subjects’ behavior are not known. The ideal is to combine insights from both approaches.
USING GAME THEORY TO EXPLORE JUSTICE-RELATED BEHAVIOR
The Ultimatum Game
In 1982, Werner Guth presented the Ultimatum Game (UG), designed to explore fairness in bargaining behavior in humans (Guth et al., 1982) (Fig. 11.1). In this game, one individual, a proposer, is given a set sum and told to allocate it between himself or herself and a partner. If the partner, called the responder, accepts the allocation, both parties receive the money as allocated, but if the responder rejects it, neither party receives anything. Hundreds of experiments across the last three decades have shown that modern Western humans typically allocate about 40 percent of the sum
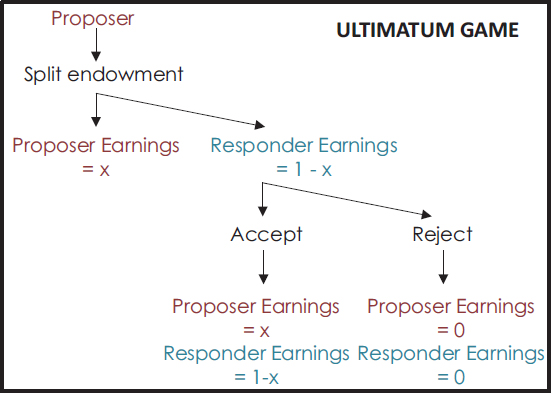
FIGURE 11.1 Schematic diagram of the UG.
to the responder, and responders typically reject any offer under about 20 percent of the allocation (Camerer, 2003). Allocations are higher than those seen in a related game, the Dictator Game (DG), in which the responder has no recourse to the proposer’s allocation, indicating that the proposer alters his or her allocations dependent upon whether the responder can influence the outcome.
The UG is often considered to measure fairness preferences; thus, it is particularly relevant to considerations of fairness and justice in other species. There have been two studies in chimpanzees, both relying on a limited-form version of the game in which proposers make a choice from between two preset distributions. In neither case did chimpanzee responders refuse allocations; however, proposers’ behavior differed between the studies. In the first, proposers chose between the two trays, each of which contained foods for them and their partner (neither chimpanzee could reach their partner’s food). After the proposer chose one option, by pulling the corresponding tray within reach of the responder, the responder could choose whether or not to accept the offer by pulling the tray in such a way that both the proposer and the responder could access their rewards. In this study, proposers consistently chose the larger outcome for themselves and responders accepted virtually any offer (Jensen et al., 2007). However, responders accepted more than half of zero offers (i.e., they received nothing themselves), indicating that they may not have understood the task. A subsequent study showed that human responders rarely refused in a similar situation, possibly because of the difficulty of inhibitory responses
(Smith and Silberberg, 2010), indicating that the deviation from behavior typically seen in the UG could have been attributable to the experimental design rather than a species difference.
In a more recent study (Proctor et al., 2013), subjects were trained that one token represented an equal 3/3 split of six food items, whereas the other represented a 5/1 split of the foods in favor of the proposer. Proposers could choose a token and pass it to the responder, who could then trade it out to the experimenter (accepting the proposal) or not (refusing it). Proposers’ choices in the UG were compared with those in a DG-like condition in which the responder was present but had no recourse (proposers passed the chosen token directly back to the experimenter). The procedure was repeated using children paired with another familiar individual from their daycare class to ensure that humans would respond similarly to chimpanzees in this procedure.
Despite responders never refusing offers, all chimpanzee proposers’ choices shifted from preferring the more beneficial 5/1 token in the DG-like condition to preferring the equal 3/3 split in the UG condition. Validating the procedure, children responded in the same way. These results differ from typical UG results because neither the children nor the chimpanzees ever refused the low offer. However, in typical human UGs, responders are interacting with anonymous partners in a one-shot situation, and their only recourse is to refuse the offer. In this study, responders were interacting with individuals from their social group and had options besides refusal, such as punishment or the threat of punishment, which are less costly means of protest than refusing a positive, if relatively smaller, outcome (Milinski, 2013). Similarly, in typical UGs, proposers’ refusals decrease when they are given other options, even in one-shot games [e.g., the responder can write a note to the proposer (Xiao and Houser, 2005)]. These results do not necessarily mean that the chimpanzees showed a sense of fairness, because if this were the case, they should have preferred the equal token in both the DG and the UG conditions. This also does not mean that proposers had prosocial motivations, because they may have changed their preference to avoid refusals by the responder. Nonetheless, both humans and chimpanzees were apparently sensitive to how their choices influenced their partner’s decisions and changed their behavior accordingly.
The Impunity Game
Closely related to the UG is the Impunity Game (IG) (Fig. 11.2). The IG is similar to the UG, but if the responder refuses the proposer’s offer, the responder gets nothing, whereas the proposer receives their portion of the allocation as indicated by the proposer (Bolton et al., 1998). This game
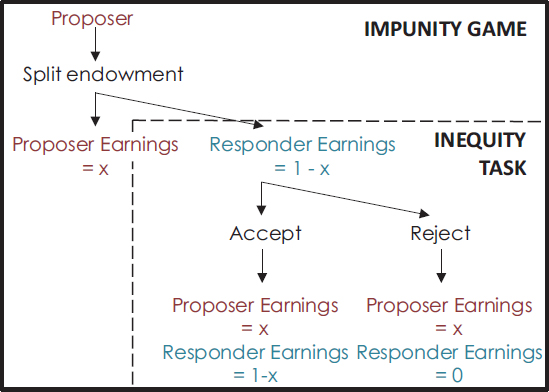
FIGURE 11.2 Schematic of the IG. The full IG has not yet been explored with other species; however, a typical inequity procedure is reminiscent of the responder’s choice in the IG (indicated by the dashed line).
has received very little attention in the human literature. In part, this is due to the assumption that no rational responder would ever refuse an offer, because this action leaves them with both absolutely and relatively less than the proposer (in the UG, a refusal leaves the responder with absolutely less but relatively the same as the proposer). Nonetheless, in a study that used the same procedure for both games, human subjects refused in the IG about half as often as in the UG for a given distribution (Yamagishi et al., 2009).
Much as in the UG, the challenge of the IG with primates is assuring that the responders understand that their choice was constrained by the actions of the proposer. Most studies of primates to date have explored only responses to unequal distributions, to learn how individuals respond to inequity when they cannot alter their partner’s outcomes [reviewed in Price and Brosnan (2012)]. In a typical experiment, paired subjects alternate completing a task (such as exchanging a token) with a human experimenter. The experimenter rewards subjects with the same rewards (Equity condition) or gives one a more preferred reward (Inequity condition). To rule out the possibility of contrast effects (Reynolds, 1961; Roma et al., 2006), in which subjects compare their outcomes to those present in the environment without attending to the social component, pairs are given a Contrast condition, in which both partners are shown the preferred reward but, after completing the task, receive the less-preferred one (Brosnan et al., 2010b; Talbot et al., 2011). These responses are referred
to as Inequity and Equity rather than Inequality and Equality because, despite the fact that the outcomes are identical (or not), we cannot know whether they are perceived identically by subjects because of differences in rank, hunger level, etc.
Initial studies confirmed that both capuchin monkeys (Brosnan and de Waal, 2003; van Wolkenten et al., 2007; Fletcher, 2008) and chimpanzees (Brosnan et al., 2005, 2010b) were more likely to refuse to complete the interaction (e.g., return the token or accept the subsequent food reward) in the Inequity condition compared with either the Equity condition or the Contrast condition, although there was variation within and between studies [discussed below in more detail (Brosnan et al., 2005; Bräuer et al., 2009; Silberberg et al., 2009)]. Because humans refuse more frequently in situations in which they do not like the outcome (e.g., in the UG game), this similar behavior in other species has been interpreted as responding negatively to inequitable outcomes. The response is also sensitive to context, with refusals occurring only after subjects have completed a task but not when rewards are simply handed out for “free” (Bräuer et al., 2006; Roma et al., 2006; Dindo and de Waal, 2007; Neiworth et al., 2009; Silberberg et al., 2009). Despite this, subjects do not change their rate of refusal when they have to work harder than a partner to achieve their reward [Fontenot et al. (2007), but see Takimoto and Fujita (2011) for a response to effort using a different paradigm]. This indicates that although subjects are sensitive to inequity in the context of joint action, it is the difference in outcome rather than effort that is important for their decision making. This may indicate that subjects are more sensitive to distributional inequities than procedural ones.
Demographic variables also influence individuals’ responses, but not always in consistent ways. For instance, not all studies find a dominance effect, although for those that do, dominant individuals always respond more strongly than do subordinates, presumably because they are more accustomed to receiving higher-valued rewards (Bräuer et al., 2009; Brosnan et al., 2010b). There are also no consistent sex differences (Brosnan et al., 2005, 2010b), possibly because of relatively small sample sizes (although several dozen chimpanzees have been tested across three facilities, a large sample for captive primate studies, this is a very small sample size for untangling individual variation). The degree to which individual differences influence behavior—both in nonhuman and human primates—is a topic that requires additional exploration.
Finally, not all primate species refuse in conditions of inequity. The same procedure to elicit inequity has now been used to test seven primate species, and similar procedures have been used with several others (Table 11.1). Comparing these different species’ responses allows us to consider which social or environmental factors may have played a role in shap-
TABLE 11.1 Summary of Inequity Studies by Species, Including Whether Inequity Aversion Has Been Found with Different Methods and the Influence of Effort and Social Context
| Species | Respond to Inequity? | Task Used/Results | Effects of … | ||||||
| Exchange | Other Task | No Task | Effort? | Social Context? | |||||
| Chimpanzee | Yes | Yes (Brosnan et al., 2005, 2010b; Bräuer et al., 2009) | — | No (Bräuer et al., 2006) | No (Brosnan et al., 2010b) | Social closeness (Brosnan et al., 2005) Sex (Brosnan et al., 2010b) Rank (Bräuer et al., 2006; Brosnan SF et al. 2010b) | |||
| Bonobo | Possibly | Trend toward yes (Bräuer et al., 2009) | — | No (Bräuer et al., 2006) | — | , — | |||
| Gorilla | Yes | — | — | No (Bräuer et al., 2006) | — | Social play (van Leeuwen et al., 2011) | |||
| Orangutan | No | No (Bräuer et al., 2009; Brosnan et al., 2011) | — | No (Bräuer et al., 2006) | — | — | |||
| Macaque | Yes | Yes (Hopper et al., 2013) | Yes (Massen et al., 2012) | — | Yes (Massen et al., 2012) | Rank (Massen et al., 2012) Social relationship (Massen et al., 2012) Ontogeny (Hopper et al., 2013) | |||
| Capuchin | Yes | Yes (Brosnan and de Waal, 2003; van Wolkenten et al., 2007) No (Fontenot et al., 2007; Silberberg et al., 2009) | Yes (Fletcher, 2008; Brosnan et al., 2010a; Takimoto et al., 2010) | No (Roma et al., 2006; Dindo and de Waal, 2007) | Yes (Takimoto and Fujita, 2011) No (Fontenot et al., 2007; van Wolkenten et al., 2007) | Rank (Takimoto et al., 2010) | |||
| Squirrel | No | No (Talbot | — | — | — | — | |||
| monkey | et al., 2011) | ||||||||
| Tamarin | No | No (Neiworth et al., 2009) | — | — | — | — | |||
| Marmoset | No | No (Freeman et al., 2013) | — | — | — | — | |||
| Owl monkey | No | No (Freeman et al., 2013) | — | — | — | — | |||
NOTE: Dashes indicate that no study has yet been done on the topic.
SOURCE: Adapted from Price and Brosnan (2012) with permission from Springer Science+Business Media B.V.
ing the evolution of the response. Although more species will need to be tested to verify these conclusions, patterns are emerging. For instance, responding negatively to inequity is not a homology within the primates. Among the great apes, bonobos may respond similarly to chimpanzees and humans, indicating that the last common ancestor of Pan and Homo shared this behavior. Only five bonobos have been tested, making statistical analyses difficult; however, their refusal rates doubled from 10 to 20 percent in the Inequity compared with the Equity condition (Bräuer et al., 2009). On the other hand, orangutans showed no response to inequity (Bräuer et al., 2009; Brosnan SF et al., 2011), eliminating the possibility of a homology within the great apes. Additionally, the behavior is not a homology within the New World monkeys, because of the five New World monkeys tested, only capuchins responded differently to the Inequity condition. The orangutan’s behavior also rules out the possibility that this is related to increased brain size or advanced cognitive capabilities, because orangutans have a brain-to-body ratio and cognitive skills similar to the other great apes.
Thus far, negative response to inequity correlates positively with whether a species cooperates routinely with nonkin [reviewed in Brosnan (2011)] (Fig. 11.3). Chimpanzees, bonobos, and capuchin monkeys cooperate with nonkin, for instance in coalitions and alliances, and all three species responded to inequity in these experiments. Macaques, too, both responded negatively to inequity (Massen et al., 2012; Hopper et al., 2013) and showed cooperation in the form of coalitions and alliances (Maestripieri, 2007). However, in squirrel monkeys, a species confamilial (and sympatric) with capuchins but lacking extensive social cooperation, males responded more strongly to the Contrast condition than either the Inequity or Equity conditions, indicating that their reactions were more strongly influenced by their previous offers than by rewards received by another monkey (Talbot et al., 2011). Again considering orangutans, one way in which they differ from chimpanzees and bonobos is their relative lack of social cooperation. A potential challeng to this hypothesis arises with the species that show biparental care. These species cooperate extensively, working together to raise offspring, but thus far there is no indication that such species respond negatively to inequity in this experimental paradigm (Neiworth et al., 2009; Freeman et al., 2013). However, the proposed function of responding to inequity is to judge the value of cooperative partners to determine whether to search for a new partner. It is very costly to find a new pair mate, so it is possible that even if individuals in biparental care species do notice inequities, they have evolved to respond only to very costly inequities, which do not include receiving a less-preferred reward than their mated partner.
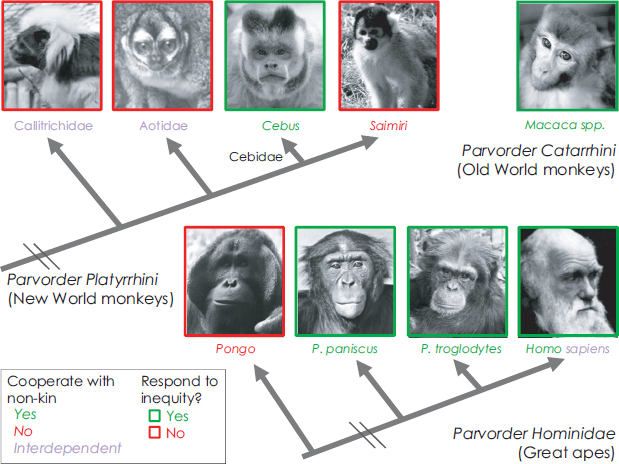
FIGURE 11.3 Schematic diagram indicating the responses to inequity of species that have been tested using the typical inequity procedure in which subjects exchange tokens for rewards while sitting next to a partner. The photos representing species that show a negative response to inequity in this task are outlined in green, and those that do not are outlined in red. The names of species that show extensive social cooperation, such as coalitions and alliances, are in green, those that do not are in red, and interdependent species (e.g., those that show biparental care) are in violet. Humans show both extensive social cooperation and biparental care. Note that this diagram excludes gorillas, for whom too little information is known. The photographs of the macaque and bonobo are by F. B. M. de Waal. The photograph of the orangutan is by C. Talbot. The photograph of Darwin is from the public domain. All other photographs are by the author. [NOTE: Figure can be viewed in color in the PDF version of this volume on the National Academies Press website, www.nap.edu/catalog.php?record_id18573.]
Related to this, subjects should be more sensitive to inequity when forming a relationship, which is a time when the costs of finding a new partner are less, than after the relationship is established and there is joint investment in offspring (Brosnan, 2011). One drawback to our understanding of human behavior is that most studies to date have involved unrelated individuals, mostly strangers, virtually always in conditions of anonymity, and often without repeated interactions, which removes the
possibility of reputations forming. Although these conditions are informative for exploring the degree to which humans will cooperate without either a relationship or social knowledge, this is not as informative for telling us about typical human cooperative behaviors, which primarily take place within established relationships with known partners. Additional experiments that explore differences in behavior in the context of different social relationships and with different degrees of anonymity will be essential to understand how inequity functions in the context of the relationship.
Prosocial Games
Despite children’s calls of “that’s not fair” when they receive a less-preferred outcome than another child, responding negatively to receiving less than one’s partner is only one part of a sense of fairness. A sense of fairness or justice requires responding not only to inequities that affect oneself but also to those that affect others (Hatfield et al., 1978; Fehr and Schmidt, 1999). In other species, this has most commoly been experimentally explored using procedures reminiscent of the Dictator Game. In the typical experimental game with primates, the subject has the option to provide rewards, or not, to a social partner. Importantly, the subject’s behavior when the subject is with a social partner is compared with their behavior when alone, ruling out the possibility that they are choosing the prosocial option because there are a greater number of rewards visible than in the nonprosocial option. The critical difference between these games and the DG is that choosing to reward the partner does not reduce the subject’s own payoffs in these prosocial games; thus, there are no disincentives for prosocial behavior.
Although reward structures have varied across experiments, making direct comparisons difficult, a sufficient number of these studies exist that patterns are emerging. Chimpanzees choose to provide rewards to their partners in some experiments (Horner et al., 2011) but not others (Silk et al., 2005; Jensen et al., 2006), and it is not yet clear which features lead to prosocial behavior in this ape. It is also not clear that these results are reflective of chimpanzees’ prosocial tendencies. Results in the inequity studies described above imply that chimpanzees may recognize when they receive a better outcome than a partner even when they chose not to act to change the situation. In one study, subjects sometimes refused preferred rewards when their partners got less-preferred ones and did so at greater rates than when both received preferred rewards (Brosnan et al., 2010b). This does not mean that the subject’s motivations were prosocial, because the subject may have feared the partner’s reaction to receiving a
less-preferred reward than the subject, but these results verify that chimpanzees recognized when they received better outcomes than a partner.
Among the monkeys, capuchin monkeys preferentially chose the prosocial option (de Waal et al., 2008; Lakshminarayanan and Santos, 2008) and were more likely to reward kin than either nonkin group mates or nongroup mates (de Waal et al., 2008). Callithrichids, one of the taxa that show biparental care, also chose the prosocial option that rewarded a group mate (Burkart et al., 2007; Cronin et al., 2010), although like the chimpanzees, they did not do so in all experiments (Cronin et al., 2009). Finally, long-tailed macaques also showed prosocial behavior, but in this study, dominants chose the prosocial option, whereas subordinates did not (Massen et al., 2010). Again, these studies measured behavior, not motivations, so they cannot shed light on why the monkeys behaved prosocially (i.e., they cannot disentangle whether subjects were prosocially motivated to help their partners or were selfishly motivated to avoid repercussions for not helping their partners). Nonetheless, although there is still debate about the contexts under which each of these species shows prosocial behavior and the selective pressures underpinning it, in some circumstances, all of the primates tested thus far actively chose an outcome that benefited a partner as well as themselves.
Species-Specific Approaches to Justice and Fairness Inequity in the Context of Cooperation
If responding to inequity is a mechanism for identifying good cooperative partners, then inequity should influence behavior when cooperation is required. This can be experimentally explored by introducing inequity into an experimental cooperation paradigm. Capuchin monkeys have been studied extensively in the realm of cooperation, and there is evidence that they understand the contingencies of cooperative tasks and are sensitive to their partner’s behavior [reviewed in Brosnan (2010)]. In the first study to explore cooperation in a situation of inequity, capuchin monkeys could work together to pull in a heavy tray to obtain food, but only one monkey got the food. Monkeys worked together, but only if the individual who received the food shared with the individual who did not. Perhaps anticipating this, the monkey who got the food was more likely to share in this condition than in a control condition in which they could acquire the food without their partner’s help. This indicates a sensitivity to their partner’s reaction and an anticipation of their need for their partner’s help (de Waal and Berger, 2000).
In a study more explicitly exploring inequity and cooperation, monkeys were tested in a similar paradigm in which both monkeys always received food, but these foods differed in the degree to which the monkeys
preferred them. Thus, sometimes both monkeys received the (same) preferred food, sometimes they received the (same) less-preferred food, and sometimes one received the preferred food, whereas the other received the less-preferred food. The monkeys were not separated nor were their roles predetermined by the experimenter, so in all cases, the monkeys had to negotiate between themselves who would pull from which location, which also determined who got which food item. The pair’s pulling success was not dependent upon the value of the rewards; pairs pulled in the tray just as often for two identical rewards as for two different ones. Instead, the monkeys were sensitive to their partner’s behavior. In some partnerships, individuals shared access to the more preferred reward when the rewards differed, each receiving it approximately half of the time. In these partnerships, cooperative pulling success was high (~70 percent) in all three conditions. On the other hand, for partnerships in which one member dominated the preferred reward in the different-reward condition, they succeeded in pulling in the tray only about 30 percent of the time across all three conditions (Brosnan et al., 2006).
These results show two important things. First, subjects in the inequitable partnerships apparently reacted against their partner, not the reward distribution. That is, when one member dominated the preferred reward in the different-reward condition, pulling rates dropped across all conditions, even the two conditions in which both monkeys got the same reward. This supports the hypothesis that recognizing inequity helps to identify cooperative partnerships, because these subjects refused to work with a specific partner rather than refusing a certain outcome. Second, in the partnerships in which neither individual dominated the preferred rewards, subjects were, by definition, receiving the less-preferred reward on half of all different-reward trials, and yet cooperation was maintained. The reality of cooperation is that it is unlikely that every individual will receive the same reward on every trial, and these results indicate that capuchins are somehow able to extrapolate across multiple trials and therefore reap the gains of cooperation despite variation in outcome.
Inequity in Interactions with the Experimenter
Although the above data relate to primates’ rewards in comparison with each other, how do primates react toward the experimenter who caused the inequity? A recent study explored which of two novel experimenters the monkeys preferred to interact with in a task in which the monkey returned a token to an experimenter to get a food reward. Subjects first gained experience with both experimenters, one of whom consistently provided the offered rewards and one of whom failed to provide the offered reward (or any reward) on up to 50 percent of trials.
When the monkeys then had free choice as to which experimenter they would return the token to, subjects were equally likely to interact with both experimenters (Brosnan and de Waal, 2009). Although it may be that capuchin monkeys do not form preferences for experimenters based on reliability [they do show preferences between experimenters in other situations (Paukner et al., 2009)], it is also possible that the relationship between humans and captive nonhuman primates influenced the results. Despite the fact that subjects in Brosnan and de Waal (2009) had never interacted with the experimenters who participated in the study (novel experimenters were used for each condition), they did have extensive experience with experimenters in general, as well as with trials in which rewards were not given (e.g., because of incorrect responses). Thus, they may have responded to this situation by trying to “correct” their response, rather than recognizing the experimenter’s unreliability. Pet dogs, who may have a different relationship with humans, do discriminate between human experimenters in a similar task (Horowitz, 2012).
Inequity in Social Interactions
Some of the best evidence in favor of a sense of justice or a sense of fairness comes from observational studies of nonhuman primates’ interactions during their natural behavior. These interactions also offer the opportunity to look beyond distributional equity. Unfortunately, in many cases, these events are extremely rare, making it difficult to make strong statements about these behaviors. Moreover, observational work yields correlations, which cannot tell us causation without controlled studies that alter one variable at a time. Nonetheless, these situations may be the best window we have into primates’ sense of fairness and justice because they are natural interactions, unconstrained by interactions with humans or experimental contingencies.
Chimpanzees display a sense of “social regularity” that has been linked to the human sense of justice (de Waal, 1991). Chimpanzees appear to have expectations about others’ behavior and are known to express frustration, sometimes aggressively, when these expectations are violated. Moreover, such responses are not limited to the apes. Apes and monkeys both engage in policing behavior in which individuals, typically the highest-ranking males, intervene in fights to maintain group stability. Some of the best evidence for the utility of policing behavior comes from studies done with macaques. In these studies, the group’s behavior was compared on typical days and those in which the highest ranking males were removed. Results demonstrated the males’ necessity in stabilizing the group’s interactions (Flack et al., 2005, 2006). Similarly, both macaque and chimpanzee males, once they have established high status, will inter-
vene in fights on the side of the loser (de Waal, 1978; Watanabe, 2001). Although there are many potential reasons for such behavior, it again indicates that these males recognized social inequalities in others’ interactions and were willing to act against their own short-term self-interest to rectify them. Of course, in the longer term, maintaining group stability always benefits a high-ranking male, and a physical intervention may additionally emphasize his power to others in his group. Nonetheless, there are short-term costs to entering into any fight, particularly in favor of an individual who is losing.
Recently, inequity has also been linked with play fighting in gorillas. Gorillas who gained the upper hand during a bout of play fighting worked to maintain it, possibly indicating that they were aware of the social dynamics and strove to maintain their competitive advantage (van Leeuwen et al., 2011). The authors emphasized the benefit of studying inequity in a natural social context and considered how responses to inequities may vary depending on whether the individual in question has the social advantage. Again, for practical experimental reasons, much work focuses on inequity of food rewards, but attention to inequity may have evolved in the social realm and only more recently adapted to physical outcomes, such as food. Future work will help to determine how responses to inequity vary depending upon the commodity in question.
INEQUITY AND RELATED BEHAVIORS
Inequity likely interacts with other behaviors, although, thus far, there have been few studies to explore these interactions.
Punishment
Recognizing inequity is only one of several behaviors that likely underpin cooperation. Punishment may also support cooperation. Cleaner fish, which cooperate with nonkin during the cleaning interaction, do not respond to inequity of rewards compared with a social partner (Raihani et al., 2012). However, the fish do punish one another for acting against the pair’s interests (Raihani et al., 2010). Thus, it may be that in species or situations in which finding a new partner would be difficult or costly, punishment to change the current partner’s behavior is used in lieu of finding a new partner (Raihani and McAuliffe, 2012).
Inequity and Contrast Effects
It seems likely that responses to inequity share cognitive mechanisms with behaviors such as loss aversion and contrast effects, both of which
involve reactions to unexpectedly lesser outcomes. These are rather difficult to untangle experimentally, given that inequity is essentially a contrast effect with another’s outcome as the referent (similarly, inequity is a loss with another’s outcome as the referent). However, we can compare the relative strengths of these motivations using well-designed experiments. For instance, whereas some species (e.g., chimpanzees and capuchin monkeys) were more sensitive to inequity with one’s partner than to contrast with previously offered outcomes, others (e.g., squirrel monkeys) showed the reverse. Mapping such preferences may help to determine the underlying influences on the expression of these behaviors, providing insight into the relative strength of social and nonsocial environmental stimuli on a species’ decision making.
FUTURE DIRECTIONS AND CONCLUSIONS
Although in the previous decade we have learned much about nonhuman primates’ responses to inequity, there remains much to learn. First and foremost, humans are primates, which makes nonhuman primates a natural group for comparison, yet other species also respond to inequity (Range et al., 2009; Horowitz, 2012; Raihani et al., 2012; Wascher and Bugnyar, 2013). More work is needed to identify which species respond to inequity, whether these responses are similar across species, and the nature of individual differences in response in these species. This will also provide a stronger test of the hypothesis that inequity responses correlate with cooperation.
Another important area of inquiry is distinguishing situations in which nonhuman species notice versus respond to inequity. All of the studies described above focus on behavioral responses to differing outcomes, but subjects may notice an inequitable situation that they do not respond to, for instance, because they are unable to inhibit their desire for the reward, because of social pressure, or for other reasons not yet known. One way to address this question is to explore changes in individuals’ affect across different conditions to identify responses that occur in the absence of an overt behavioral change. This approach has been successful in identifying recognition of inequity in young children (LoBue et al., 2009).
Inequity also occurs in a variety of contexts that are not yet reflected in the nonhuman literature because of the focus on distributional concerns in experimental tests. One major focus of human research that has received little attention in the nonhuman literature is procedural inequity (Skitka, 2012). Although preliminary investigations reveal no change in subjects’ behavior based on procedural variations (Brosnan et al., 2010b), nonhuman primates have expectations about others’ behaviors and it is
reasonable to assume that they are sensitive to procedural differences in some contexts.
All of the experimental work to date has involved an egocentric approach to fairness and justice, yet observational studies of natural behavior indicate that primates will intervene in the interactions of others, in some cases impartially. Although challenging, it is possible to study fairness in the laboratory removed from egocentric reactions. Experimental studies on reputation in chimpanzees indicates that these apes are able to form opinions about human experimenters from watching these experimenters interact with other chimpanzees (Subiaul et al., 2008). A fruitful line of inquiry would be to explore nonhuman primates’ judgments of the equity of others’ interactions. Such third-party assessments of others’ distributions would be the first research to experimentally examine justice in other species.
Finally, studies of justice and fairness will advance significantly when work from the laboratory is combined with what we know about animals’ natural behaviors in the wild (Janson and Brosna, 2013). In this way, we will better understand the social and ecological conditions under which these preferences manifest, which lends additional insight into their function. Additionally, combining these behavioral studies with what we know about cognition may yield insights into these behaviors, such as the overlap between inequity and delay of gratification (Price and Brosnan, 2012).
Humans are not alone in responding negatively to differential treatment as compared with a partner. This response is shared with other species and appears to be instrumental in successful cooperation. Understanding the evolution of this response provides insight into our own behavior. Although nonhuman primates do not show a sense of justice or fairness in the same way as humans do, gaining knowledge about the evolution of justice and fairness from comparative studies will improve our understanding of these judgments in humans as well as in other animals.
ACKNOWLEDGMENTS
I thank L. Salwiczek for helpful comments on an earlier draft of this manuscript. Funding was provided by National Science Foundation (NSF) Faculty Early Career Development Award SES 0847351 and NSF Grant SES 1123897.




















