
5
The Behavior of Dolphins and Tuna in the ETP
A great deal of effort has been expended to understand the impact of purse seining on dolphin and tuna stocks of the ETP. Features such as growth, reproduction, distribution, and trends in abundance of various tuna and dolphin stocks under exploitation have all been under continuing analysis for more than a decade. In addition, sporadic direct studies of dolphin and tuna behavior have been done during the course of purse-seining operations. The major impediment to doing comprehensive behavioral studies has been that such studies require work aboard seiners. While work on most other aspects of dolphin biology and mortality can go on while a seiner goes about its work, behavioral studies involve long-term dedication of a vessel to such study. Such an intensive long-term effort has not been undertaken.
The empirical experiments of fishermen (with input from NMFS) are largely responsible for the gear and methods we see today. They represent a subtle exploitation of the behavioral biology of tuna and dolphins. Such development has resulted, however, in an effective tuna capture method that continues to kill thousands of dolphins each year. For these reasons, the behavioral work of both scientists and fishermen needs to be extended.
The sole behavioral effort during actual seining operations was the Dedicated Vessel Program of 1978–1979 (see Fox and Lenarz, 1975; Stuntz, 1977; Bratten et al., 1979; Powers et al., 1979; DeBeer, 1980; Holbrook, 1980), and this program allowed only a relatively few experimental net sets to be made. Even though it was short, much of the work discussed here came from that pioneer program. In nearly every case, ideas could not be followed very far, and many tests were ended before their real promise could be assessed.
GENERAL DOLPHIN BIOLOGY AND ECOLOGY
Several species of dolphins are found in association with tuna ( Table 5-1). The spotted dolphin (Stenella attenuata) is by far the most important from the point of view of frequency of association with tuna and use by fishermen for catching tuna. Three stocks of this species are in the ETP ( Figure 5-1). The northern offshore and southern offshore stocks are considered separately; the coastal stock is subjected to very little fishing effort. The different boundaries on the figures correspond to the limits used in different assessments of abundance of the dolphin populations.
The frequent appearance of spinner dolphins (Stenella longirostris) in sets makes this species quite significant as well, although in almost all cases it appears in mixed herds with the spotted dolphin. Figure 5-2 shows the stock boundaries proposed for this species. Recent research by Perrin (1990) suggests that the northern and southern stocks of whitebelly spinner dolphins be merged into a single stock. In this case, every stock except the Costa Rican stock is to be considered separately. (See Perrin, 1990, for taxonomic analysis of subspecies of S. longirostris; he describes the Costa Rican stock as a separate subspecies.)
The common dolphin (Delphinus delphis) is another important species, although sets on this species are less frequent than on the previous two. There are three stocks—northern, central, and southern ( Figure 5-3).
A few other species are found in association with tuna but much less frequently. These include the striped dolphin (Stenella coeruleoalba), the roughtoothed dolphin (Steno bredanensis), the bottlenose dolphin (Tursiops truncatus), and Fraser's dolphin ( Lagenodelphis hosei).
TABLE 5-1 Species of Dolphin Found in Association with Tuna as Determined by Frequency of Sets on Different Species a
|
Dolphin |
Percent Sets in 1988 b |
|
Northern Offshore Spotted |
81.6 |
|
Eastern Spinner |
23.7 |
|
Northern Whitebelly Spinner |
8.6 |
|
Central Common |
4.7 |
|
Southern Offshore Spotted |
2.1 |
|
Northern Common |
2.0 |
|
Southern Common |
1.2 |
|
Southern Whitebelly Spinner |
0.8 |
|
a Data from IATTC. b The sum of the percentages exceeds 100 because many sets are made on mixtures of species or stocks; thus many are counted twice (e.g., almost all sets on eastern spinner dolphins also include spotted dolphins). |
|
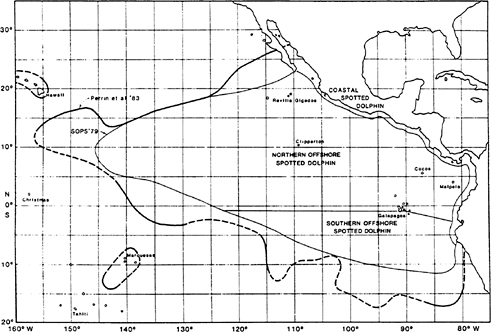
FIGURE 5-1 Known distribution of the spotted dolphin in the eastern Pacific, showing the 1979 species range and stock boundaries presented at the Status of Porpoise Stocks (SOPS) Workshop and the 1983 species range (Perrin et al., 1983). Source: Perrin et al., 1985.
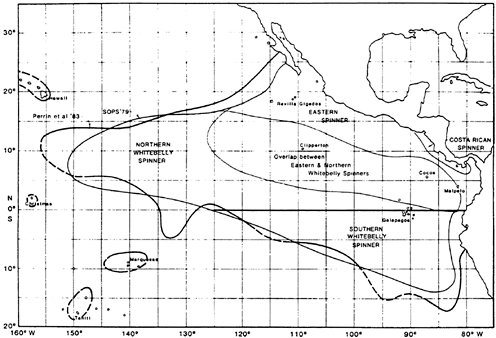
FIGURE 5-2 Known distribution of the spinner dolphin in the eastern Pacific, showing the SOPS 1979 species range and stock boundaries and the 1983 species range (Perrin et al., 1983). Source: Perrin et al., 1985.
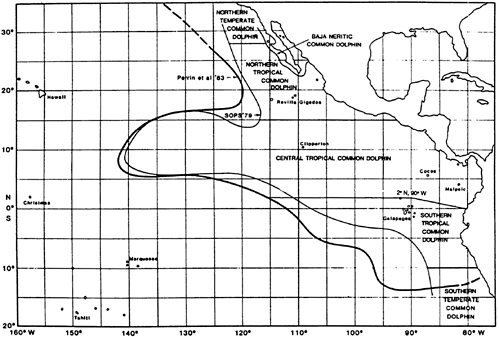
FIGURE 5-3 Known distribution of the common dolphin in the eastern Pacific, showing the SOPS 1979 species range and stock boundaries and the 1983 species range (Perrin et al., 1983).
Source: Perrin et al., 1985.
THE TUNA-DOLPHIN RELATIONSHIP
The bond linking tuna and spotted dolphins is a remarkably strong one. It may persist through much or all of the seining operation. This bond means that during seining, tuna and dolphins continue to associate so tightly that to catch dolphins is also to catch tuna.
It has been a subject of debate whether the tuna-dolphin relationship is one-sided or symmetrical (i.e., is the attraction mutual or is one species attracted to the other?). The point is worthy of attention not only because the nature of the relationship may predict the course of fish accumulation but also because it will affect choices made on whether to manipulate one partner or the other. If, for example, the dolphins are central in the association and the tuna are followers, an operation that attempts to release dolphins through the net perimeter is likely to result in tuna following.
Some modestly convincing evidence indicates that the dolphins, especially the spotted dolphin, are the central species in the association and not the tuna. That is, the attraction involved is of tuna for dolphin and is neither a mutual
-
-
bond nor a dolphin-to-tuna bond. This view is prevalent among fishermen and is supported by the following observations.
-
No evidence suggests that dolphins protect tuna, rather the association appears passive on the part of the dolphin. It seems reasonably clear that even though fishermen repeatedly remove tuna from dolphin herds, the two species continue to reaggregate daily and thus to sustain the fishery.
-
The seining operation itself strongly suggests that tuna follow dolphins. The seining operation works because it can corral the air-breathing dolphins by chasing and encircling them in a wake spiral. The dolphins must breathe frequently, especially under the high-exertion conditions of the chase, and hence they must stay near the surface where the seining operation is effective. The tuna are under no such constraint, and unless they adhere to the dolphin herd, it is unclear how seining catches them.
-
During bait-fishing days (jack-pole fishing), dolphins would stay near tuna vessels. Bait fish were tossed into the water (chumming) to hold fish near the boat where they could be hooked. If the dolphins remained near the vessel, tuna could be caught, but if the dolphin herd moved away, so did the tuna (H. Medina, El Cajon, Calif., personal commun., 1990).
-
During seining, if a dolphin herd becomes fragmented during the chase and part succeeds in escaping, the escaped dolphins often carry tuna with them. Conversely, if the maneuver successfully contains all of the dolphins, then the fish school tends to remain in the net (H. Medina, personal commun., 1990).
-
Tuna have been observed underwater in a purse seine following dolphins (Norris et al., 1978) but dolphins do not seem to follow tuna.
One possible cause for the tuna-dolphin bond is that the echolocation system of the dolphins allows them superior environmental surveillance related either to food finding or to protection or to both. Considering the moderate visibility in daytime surface waters of the ETP, dolphins should be able to detect food schools by echolocation at least three to five times farther away than a fish could detect them by eye. Since light is extinguished rapidly with increasing depth (McFarland and Loew, 1983), such a disparity should be greatly heightened during even rather shallow dives by the combined fish and dolphin herd.
Of course, food detection may be based on a sense other than vision. Many fish have very sensitive chemoreceptors (taste and smell) and can detect some compounds at extremely low concentrations (Hasler, 1966). Dolphins have highly developed organs of taste but do not have a sense of smell, so it is unclear if one species has an advantage in locating food with these means. It is likely, however, that the dolphin is superior to the tuna in terms of detailed environmental inspection beyond the reach of sight because of its echolocation.
-
FEEDING BEHAVIOR
Both spotted dolphin and yellowfin tuna feed predominantly on aggregations of prey that are thought to school in or above the thermocline, although each species eats a slightly different aggregation of species from this food source. Both are thought to feed primarily during daylight or twilight (Alverson, 1963; Perrin et al., 1973; Scott, 1991), although spotted dolphins may also feed at night to some extent (Leatherwood and Ljungblad, 1979; Scott and Wussow, 1983).
The spinner dolphins that often are seined with the spotted dolphin and tuna are a diving species that feeds primarily at night on small mesopelagic prey that typically concentrate in daytime below the thermocline. Some of the dolphin and tuna food species rise to the surface or close to the surface after dark (Fitch and Brownell, 1968; Perrin et al., 1973). Because prey that are not known to reach the surface are in the spinner's diet, the spinners are believed to dive to feed (Fitch and Brownell, 1968) in tuna areas, as they do elsewhere (Norris and Dohl, 1980a). Norris et al. (in press) suggest that the open-ocean spinner dolphin of the ETP aggregate with the spotted dolphin as part of a diurnally symmetrical pair of species that may be protective of the partners, one species resting while the other feeds.
The association of large tuna, but not small ones, with spotted dolphins is probably due to physical constraints. Small tuna, whose swimming costs at a given velocity are higher than those of larger dolphins, simply may not be able to sustain such a relationship. Body length is a major determinant of the energy expenditures of swimming (Lang, 1966), just as hull length is for ships. Large tuna have body masses and lengths similar to the dolphins they associate with. In addition, there are behavioral, ecological, and probably physiological requirements for the partners in this association, not all of which are understood.
BEHAVIOR OF DOLPHINS AND TUNA IN THE TUNA-SEINING OPERATION
The events in a tuna purse-seine set involve a subtle interplay between the maneuvers of the vessel and the sensory capabilities and behavior of the dolphins and the fish. The fishermen have evolved a means by which they have turned an empirical understanding of the behavior of the dolphins and tuna into a remarkably effective maneuver to catch fish. What happens in a set is worth exploring in some detail since these events also frame the causes of most dolphin injury or mortality. These events also might be used to find ways to release or maneuver dolphins in the net without losing tuna.
The entire ETP yellowfin-tuna seine fishery depends on the unwillingness of yellowfin tuna and the dolphin species most commonly associated with
them (spinner, spotted, and common dolphins) to spend much time below the shallow thermocline of this area of ocean during daylight hours (Au and Perryman, 1977; Au et al., 1979; Sharp and Dizon, 1978).
This association has been observed—and used by fishermen—in other oceans. Reports of association of yellowfin tuna with dolphins in other oceans include Cayre et al., 1988 (eastern Atlantic); Coan and Sakagawa, 1982 (eastern Atlantic); Levenez et al., 1980 (eastern Atlantic); Maigret, 1981, 1990 (eastern Atlantic); Mitchell, 1975 (eastern Atlantic); Pereira, 1985 (eastern Atlantic); Simmons, 1968 (eastern Atlantic); Stretta and Slepoukha, 1986 (eastern Atlantic); Stuntz, 1981 (central Pacific); Living Marine Resources, Inc., 1982 (Gulf of Mexico); Leatherwood and Reeves, 1989 (Indian Ocean); Montaudouin et al., 1990 (Indian Ocean); Potier and Marsac, 1984 (Indian Ocean); Northridge, 1984 (many areas); Caldwell and Caldwell, 1971 (western Atlantic); Dolar, 1990 (western Pacific); and Pacific Tuna Development Foundation, 1977 (western Pacific).
Some of these mention only observations of the association, and in one or two cases personal communications from third parties are mentioned; others include data on the sets made on dolphins.
In general terms, the frequency of the tuna-dolphin association seems to be much lower in the other ocean areas; for example, off the coast of West Africa the percentage of sets made on dolphins ranges from 0.4% in the Cape Lopez area to 4.7% off Senegal. There are no precise estimates of mortality for any other ocean area, and the proportion of trips with scientists or observers aboard is extremely low. The only mentions of mortality rates are in Levenez et al. (1980, 15 dolphins per set), Dolar (1990, 5 dolphins per set), and Mitchell (1975, several hundred dolphins in one set), but the data were not collected by independent observers; they came from surveys of fishermen and other sources.
In the ETP tuna grounds, a seine that is made deep enough can hang downward through the thermocline, and fish and dolphins will be reluctant to escape through the open bottom of the net. A possible exception is the so-called “untouchables,” an apparently regional collection of dolphin herds that may have learned to escape seines before pursing takes place. Some dolphin species, such as the bottlenose dolphin, routinely dive to safety through still open seines. The thermocline of the ETP is shallowest near the Central American coast and deepens to the west, where successful seining of dolphin-associated tuna becomes more difficult at about 80-meter depths (Sund et al., 1981).
Sighting Dolphin Herds
The dolphin herds under which tuna may aggregate are usually first located by ships' lookouts using high-powered binoculars. One of the cues is the
sighting of bird flocks circling near the horizon. Plunging or surface-feeding birds such as boobies and terns are often accompanied overhead by frigate birds, which, because their feathers are wettable, are restricted to stealing prey from other birds or to catching flying fish flushed from the water by the animals below. Fishermen have reported that the general size of a dolphin-tuna group below a bird flock can be predicted by the number of frigate birds flying over it (Norris and Dohl, 1980a). This general relationship between bird flocks and marine mammals appears to be widespread (Au and Pitman, 1988). The bird flocks can be detected also by sensitive radar.
The Chase
Some evidence (Au and Perryman, 1982) indicates that some dolphin herds sense and turn away from vessels as far away as seven nautical miles. The same evidence suggests that evasive maneuvers by the dolphins may quickly increase when a vessel turns toward such herds. Once a dolphin herd has been located, the seiner launches its speedboats, which race into position outside the fleeing animals and then move ahead and turn them while the seiner follows. The entire entourage changes the dolphin's swimming path into an inward bending arc. Both the speedboats and the churning vessel leave strong wakes and produce much underwater noise, which are thought to turn the dolphins (Norris et al., 1978). Shortly, the arc becomes a circle with the vessels now curving around in the second turn of the set. The average duration of the chase is one-half hour (M. Hall, personal commun., 1991).
The dolphins sometimes attempt to cut across ahead of the bows of the fast-moving seiner and to escape out through the relatively clear water ahead of the vessel. At the slightest hint that they might attempt this, the speedboats are dispatched ahead of the seiner to race in tight noisy circles in the clear water, beating it to a froth, in attempts to force the dolphins back into the wake spiral.
The Set
As the seiner spirals inward, a diameter is reached for which the circle's circumference corresponds to the length of the purse seine plus the towline. The net skiff is then released off the seiner' s stern, pulling the net with it. The timing of this release is made by the skipper on the basis of existing wind, wave, and current conditions. Typically, the intent is to close the circle of net at the completion of a full turn and have the seiner complete the turn on the down-wind side of the net.
If, after the pickup of the skiff-end of the net, there is a gap at the stern of the seiner, the seiner backs against the net to close the circle; once the net is fully closed the pursing operation begins. After the net is pursed and no fish
or dolphins can escape, net retrieval begins as the net is drawn up through the power block on the boom and stacked on deck.
Before the net circle is closed, dolphins seem reluctant to penetrate the wake even though it is unlikely to extend downward much more than 20 meters. A speculation is that dolphins' echolocation cannot easily penetrate the wake bubbles and that this sensory deficit may contribute to the mammals' spiral course. Free bubbles are exceptionally powerful reflectors of echolocation pulses. At resonance, the scattering and absorption cross-sections of a typical bubble at sea theoretically is of the order 103 times its geometrical cross-section (Clay and Medwin, 1972). Further, bubbles of the most effective scattering size for dolphin echolocation (about 60 micrometers in diameter; Glotov, 1962) may persist for long periods while smaller bubbles dissolve quickly and larger bubbles rise quickly to the surface.
Net Closed and Pursed
When the net circle is completed and pursed, the dolphins, far across the net from the now stationary seiner, appear as a slowly milling herd, located with its outer edge about 70 meters inside the far corkline and about 450 meters (or more with recent, larger nets) from the vessel (Norris et al., 1978). These netted herds move slowly, even though many individual dolphins within the herd are obviously in fairly rapid motion and chases between dolphins are frequent. Two distinct activity classes of dolphins are often present. These classes are called rafting animals and active animals. No specific species or age groups of dolphins can be associated with these two activity classes (Norris et al., 1978; Pryor and Kang, 1980). Rafting animals hang quietly and can be seen throughout netted herds. Rafts include passive columns of animals as deep as 20 meters below the surface that may from time to time rise to the surface (Pryor and Kang, 1980).
Rafters tend to be rather closely bunched together, though mostly out of actual contact with one another except for fin-tip contact between some pairs. A better understanding of rafting might contribute to understanding the problems and possibilities of release.
The herd of dolphins with many members in vigorous motion tends to remain at a single location in the net circle. The active animals tend to dive somewhere near the periphery of the herd, swim back under it, and resurface near the far edge, to turn back and dive again, back and forth, back and forth. This behavior defines what Norris (1991) has dubbed a “teacup formation,” in which passive dolphins are to some extent left above and within the excursions of these more active animals, and in a fluid way the active animals roughly define the boundaries of a “cup.”
Entire herds of dolphins, including rafters and more active animals, can be moved within the net circle by bringing strongly aversive stimuli near them. If,
for example, a speedboat, moving by means of its noisy engine, approaches the corkline near such a herd, the entire dolphin herd has been observed to move slowly away (Norris et al., 1978). Speedboats also are able to move the herd, whereas sometimes swimmers and rubber rafts deployed inside the net cannot. The trapped dolphins tolerate swimmers moving near them, without evident flight responses. Such behavior would be unthinkable in an unrestrained herd. Encircled dolphin herds probably can be maneuvered to specific locations in the net circle to allow release efforts.
Norris et al. (1978) observed that aggression is a notable feature of active non-rafting dolphins. Aggression was also typical of patterns described in another study by Norris et al. (1985). In undisturbed spinner dolphin herds studied elsewhere in the Pacific Ocean, aggressive patterns were observed much less frequently (Norris et al., 1985).
Both Norris et al. (1985) and Pryor and Kang (1980) reported the prevalence of male coalitions in trapped dolphin herds, which has also been reported for undisturbed Hawaiian spinner herds. These coalitions consist of small, closely coordinated groups of dolphins that move as clearly distinguishable subgroups through the trapped herds. Their sex is obvious for spinner dolphins but much more cryptic for spotted dolphins (Perrin, 1972).
A protective role for these coalitions has been suggested because they frequently interposed themselves between observers and the rest of the herd, even in undisturbed herds (Norris et al., 1985). Such coalitions are active in tuna nets before backdown but have not been reported to remain during backdown. Similar male coalitions of bottlenose dolphins have been observed to cut out and sequester sexually ready female bottlenose dolphins (Connor, 1987). Although bottlenose dolphins are seldom associated with tuna or involved in the ETP tuna fishery, this observation strengthens the probability that the male coalitions have a protective role as part of their purpose.
These features indicate an organization of dolphin herds that persists early in a set and that might be utilized to assist release. If given an opportunity to escape while this protective structure is still in place, dolphins may be able to help themselves escape. In the crowding of the backdown channel, however, such social arrangements may be crowded out of existence.










