
2—
HISTORY OF THE WILD POPULATION AND CAUSES OF ITS DECLINE
This chapter reviews what is known about the special ecological requirements of the 'Alala, for example, its original habitat and geographic range, food requirements, and direct and indirect factors that probably influence the number of birds. Data on the demographics of other island populations of corvids, though not perfectly analogous to the situation in Hawai'i, are also considered in an attempt to estimate the probability of extinction of the 'Alala.
Each endangered species presents its own array of questions that need to be answered before any action can be prescribed and taken. Although the questions can be general, the uniqueness of each species requires that some answers be specific. In the case of the 'Alala, useful and pertinent information is sparse, and it is probably too late to obtain certain types of information. Despite the fragmentary nature of the information, a species-specific judgment must be made now concerning both the environmental factors that have contributed to the 'Alala's decline and the recovery actions that are most likely to promote the re-establishment of a self-sustaining wild population.
Past and Present Distribution of the Wild Population
The ancestral origins of the 'Alala are unknown, but it was probably derived from colonization of a Corvus ancestor from the Australasian region. Two recently extinct species on the islands of O'ahu (C. impluviatus), and O'ahu and Maui (C. viriosus) have been described; they are easily distinguished from the 'Alala by bill shape and body size (Olson and James, 1982b; James and Olson, 1991). C. hawaiiensis has a short, moderately arched bill, C. impluviatus had a high, impressively arched bill, and C. viriosus had a long, relatively straight bill. The 'Alala has the smallest body of these species, C. impluviatus the largest. Whether the three species were derived from a single ancestral colonizing species or several has not yet been determined (James and Olson, 1991).
Banko and Banko (1980) compiled a detailed record of distribution and published and unpublished sightings of the 'Alala. The earliest specimen was taken during the visit of Captain Cook in 1778. Sightings and specimens obtained later by naturalists and collectors clearly reveal that the range of the 'Alala was relatively restricted on the island of Hawai'i even before its historical decline (Wilson and Evans, 1893; Perkins, 1903). Specimens collected between 1887
and 1902 were all taken on the island in a narrow V-shaped belt of dry woodlands and 'ohi'a forests and moist-'ohi'a koa forests at elevations of 500–1,800 m (Perkins, 1893; Banko and Banko, 1980). The belt extended from Hualalai on the northwestern side of the island to the southern and western sides of Kilauea Crater, perhaps to Kipuka Puaulu, which is now part of Hawai'i Volcanoes National Park (Henshaw, 1902) (Figure 2.1). There were a few sightings in that area in the 1960s and 1970s. The species might have occupied high-elevation forests along the Hamakua coast on Mauna Kea, but disappeared from these forests before ornithologists explored there. The restricted distribution of the 'Alala has never been fully explained and remains an enigma, but it suggests a species with rather narrow habitat requirements or preferences.
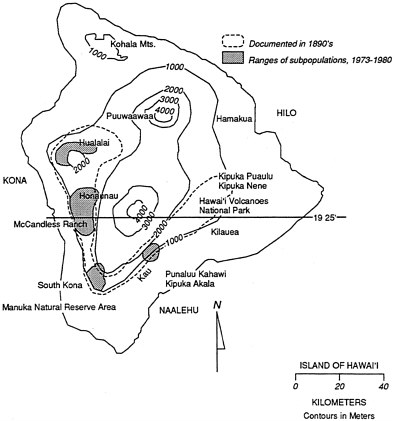
Figure 2.1 Map of the island of Hawai'i
The Geological Setting
The distribution of the 'Alala has been almost entirely confined to the slopes of two active shield volcanoes, the north and southwestern slopes of Hualalai and the southwestern slope of Mauna Loa. Some facts about the recent geological history of these volcanoes might aid in understanding the special ecology of past and present populations of the species. Radiocarbon dating of many lava flows shows that the surface of Mauna Loa, including the entire Kona coast, is being replaced at a rate of 40% per 1,000 years (Lockwood and Lipman, 1987) and the surface of the slightly older Hualalai at 25% per 1,000 years (Moore et al., 1987). Those high rates of replacement mean that only very small areas have forests that are more than 4,000 years old (Figure 2.2). As molten lava emerges from high-altitude rifts, it flows down toward the sea as a fiery river that destroys the vegetation in its path. Two predominant types of lava are a'a (rough) and pahoehoe (smooth). Detailed maps of historical and prehistoric flows show an intricate stripe-like pattern of parallel and partially overlapping flows of varied widths. Figure 2.2 documents the ages of surface flows on Mauna Loa. On Hualalai, the situation is essentially similar, but with somewhat older flows. In the high-rainfall areas, each flow appears to have been followed fairly rapidly by revegetation; a substantial forest with relatively large trees is attained in about 200 years, depending on the type of lava being colonized.
Several recent observers have noted that 'Alala appear to prefer the interface between intact forest with tall trees and areas that are open. Before the establishment of ranches, such habitat would have been characteristic of the contact zone between forested older flows and newer, open flows. Clearly, the concept of a stable ''ancient forest" habitat for the crow anywhere on this entire slope of the island is not valid. It is almost certain that former 'Alala populations continually adjusted their distribution in response to shifts in ecological conditions imposed by the periodicity of vulcanism and forest succession.
Extensive studies of the modes of natural colonization of new lava flows by plants have been made on a transect on the southeastern slope of Mauna Loa (Atkinson, 1970; Mueller-Dombois et al., 1981; Mueller-Dombois, 1987). These resulting data can serve as useful models for the understanding of the ecosystems associated with new lava flows on the Kona coast, i.e., the habitat of the 'Alala. In particular, one can estimate the characteristics of the bird's original habitat before the clearing of the forests for ranching and logging. Very few relevant areas on the Kona coast are still being colonized naturally.
When viewed in detail, each new bare lava flow is affected by unique sets of colonization events; these often result in differences in frequency of species composition from one new flow to the next. For example, the composition of major tree species is obviously similar to and derived from adjacent, older flows, but the vagaries of chance colonization, often from a distance, results in a complex mosaic of frequencies of tree species and their accompanying climbing and understory plants. Some of them are especially important as food plants for the 'Alala (see below).
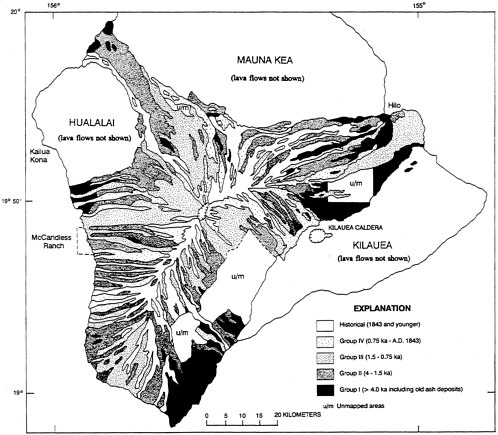
Figure 2.2 Age distribution map of Mauna Loa lava flows, showing in particular the natural parallel arrangement of the flows in present and former 'Alala habitat on the Kona coast. Most flows are more recent than 4,000 years ago. Greatly generalized from 1:24,000 mapping; adapted from Figure 18.2 of Lockwood and Lipman (1987). ka = 1,000 years before present.
As newer raw flows repeatedly cover partially vegetated older ones, the surface pattern includes various surviving pockets of older vegetation growing on an earlier flow at a slightly lower level. Such areas are called "kipukas"; some of these can be seen on large scale maps like Figure 2.2 (note dark areas), but many small-scale events are similar and generally much more significant on the local scene. Kipukas are obvious when seen totally isolated by new raw flows, but are less conspicuous as these younger flows become newly colonized. All this greatly increases the complexity of these ecosystems' mosaics. Many native species, furthermore, display exuberant genetic variation as they colonize, sometimes leading to new populations that can be recognized as genetically distinct (Mueller-Dombois et al., 1981; Carson et al., 1990).
A prominent feature of the original flora of these evolving ecosystems is the great scarcity both of mature soil and of endemic grasses, especially at the elevations where the 'Alala was originally found. It is clear that the 'Alala originally had no relationship to grasslands. The bare, rough, soil-less lavas and ashfalls were colonized naturally by a number of understory and climbing plant species that are adapted to germinate on these areas.
Ranching practice has been to remove the understory plants and trees that are close together and to plant exotic forage grasses. In this long-term process, begun two centuries ago, many weed grasses became established with the forage grasses desired by the ranchers. The process also encourages many other invasive, exotic weeds. Even though it is common practice to leave some tree overstory, the resulting pastures effectively discriminate against the maintenance of native herbs and shrubs. Cattle not only browse down the native plants and grasses, which lack spines or chemicals that naturally protect them from herbivores, but also continue to spread both forage and weed grasses and larger weeds effectively.
The spreading novel grassland forms tight mats in thin soil over the lava in which the native food plants of the 'Alala cannot germinate. Recovery and renewal of native understory can occur in an area only if cattle are excluded and a program of grass and weed removal is instituted.
Geographic Distribution of the 'Alala
Late in the nineteenth century, the 'Alala was reported to be "abundant locally" (Henshaw, 1902; Perkins, 1903; Munro, 1944; Banko and Banko, 1980) and "extremely numerous" (Henshaw 1902). "Hundreds" were seen in some ranch areas in the Kona District as late as the 1930s, but the observation by Perkins (1903) that the species was declining throughout its range turned out to be prophetic. Commercial logging and the continuing conversion of forest to agriculture and ranching were accompanied by steady declines in the number of crows. Many were shot as agricultural pests in the late 1800s and early 1900s, and this unnatural mortality undoubtedly contributed to the species' decline in some areas (Munro, 1944; Giffin et al., 1987). Unlike the American Crow (Corvus brachyrhynchos), which thrives
today in farmland, the 'Alala disappeared from highly disturbed areas—a further suggestion of relatively specialized habitat requirements.
In the Ka'u District on the southeastern section of the island, the number of 'Alala declined steadily throughout the 1900s (Munro, 1944; Baldwin, 1969; Banko and Banko, 1980). Baldwin (1969) did not encounter 'Alala during a 3-day field trip between Kipuka Akala and a point northeast of Punalu'u Kahawi, including Kipuka Nene, although some birds were still reported in this area. Three 'Alala were encountered in open-canopy 'ohi'a-koa forests with mixed native shrub understories during an extensive survey of the Ka'u Forest Reserve by USFWS in 1976 (Scott et al., 1986), and a single bird was recorded in the Hawai'i Volcanoes National Park in 1977 (Banko and Banko, 1980). The Ka'u District subpopulation is now believed to be extirpated, although to our knowledge no additional attempts have been made to locate 'Alala within the district.
The history of the 'Alala in the Kona District, on the western side of the island, has been well documented, especially since the early 1970s, and might afford insights into factors that are contributing to its critical status. Figure 2.3, compiled mainly from data supplied to the committee by the state of Hawai'i (J.G. Giffin, pers. comm., 1991), summarizes the changes. Between 1969 and 1976, W. E. and P. C. Banko made extensive population surveys of the Kona District for USFWS (Banko, 1974, 1976; Banko and Banko, 1980). The state of Hawai'i, USFWS, and others have continued to monitor the population since then (Giffin, 1983). Joint federal and state transect surveys were conducted in the Kona area from 1983 through 1987, except on the McCandless Ranch, where access was denied by the owner (Scott and Kepler, 1985; Scott et al., 1986; Giffin et al., 1987). In the Pu'uwa'awa'a Ranch area, on the north slope of Hualalai in north Kona, the 'Alala were once common in dry 'ohi'a-koa forest. In the 1940s, 'Alala occurred in the wet forests up to the summit of Hualalai (Baldwin, 1969; Berger 1981). But from the beginning of the counts made in the 1970s, it became clear that the 'Alala was declining precipitously in this area (Figure 2.3.a). The 'Alala population on Hualalai declined from 26 birds in 1974 to the single banded female that was seen in March 1991 adjacent to the Hualalai Ranch; the last nest was found in 1983 (Giffin et al., 1987; J. G. Giffin, pers. comm., 1991). Census data from the Honaunau Bishop Estate Lands in central Kona parallel those from Hualalai (Figure 2.3). The only exception to the decline appears to be on the privately owned McCandless Ranch, also in central Kona, where the only known wild population of 'Alala occurs. The owners of the ranch have reported that 10–25 birds have been present for the last decade. They permitted very little access to the property between 1980 and 1989. In 1989 and 1990, single groups of nine and four birds, respectively, were seen and videotaped on the McCandless Ranch by USFWS biologists. In January 1992, at least eight birds were seen by members of this committee on the McCandless Ranch. Surveys conducted in April 1992 have determined the current wild population on McCandless Ranch to be 11 birds (J. Engbring, pers. comm., 1992). The breeding range of the crow appears to have been further constricted since the mid-1970s (J. Engbring, pers. comm., 1992).
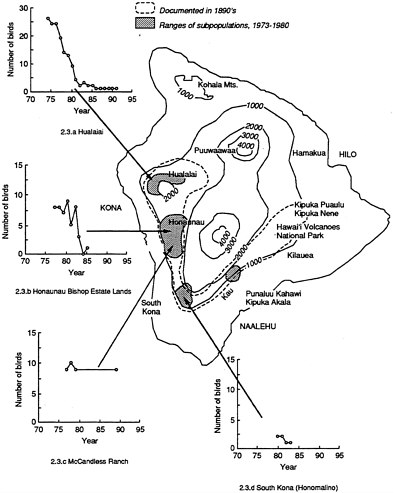
Figure 2.3 Distribution of 'Alala from survey data collected by J. Giffin (1991).
In summary, the 'Alala was declining but was still common in a narrow forest belt on the leeward southern and western side of the island of Hawai'i until about 1940. The Federal 'Alala Recovery Plan (Burr et al., 1982) gives three locations in the Kona District where at least one pair nested in the 1970s: Hualalai (north Kona), Honaunau (central Kona) and south Kona (see Figure 2.3). Those three and a location in the Ka'u Forest Reserve, also known to be in
the 'Alala's historical range, were designated as essential habitat to be protected and managed for the 'Alala. Since the Alala Recovery Plan was issued, the population has continued to decline. With the possible exception of a single female in north Kona, the small remaining wild population is now believed to be restricted to a single location in central Kona—the McCandless Ranch.
Assessment of Extrinsic Causes of the Decline in the Population
A decline in the numbers of any population is a function of low rates of reproduction, low rates of survival, or both. This section explores factors that are likely to be affecting those rates and discusses the particular ecological requirements of the species. As with many other endangered species, many of the factors associated with the decline of the 'Alala have been proposed as causes. Each of these factors could still be contributing to the decline, but it is also possible that the factors operating now are different in kind and magnitude from those which were most important in previous decades. Because many factors are likely to have acted in concert, it is not possible, on the basis of the scientific data now available, to determine the extent of their individual contributions.
Figure 2.4 is a graphic summary of environmental factors that are known to affect the numbers of 'Alala.1 The envirogram shows that activities of humans are likely to have negatively affected the survival and reproduction of this species in various ways. If data were available on the rates of the effects of each of the 10 direct and 9 indirect factors and all the important factors had been identified, we could say what is causing the decline of the 'Alala; however, that information is not available. The following discussion details what is known about the factors.
Habitat and Food
The first two directly acting factors are habitat and food: the 'Alala inhabits fairly closed (canopy cover more than 60%) to moderately open moist 'ohi'a-koa and wet or dry 'ohi'a forests with a diverse understory of fruit-bearing trees and shrubs. Recent records of 'Alala note nest sites in wet forests (Giffin, 1983; Giffin et al., 1987). In wet forests, tall, emergent koa trees serve as lookout posts and resting and displaying sites for the birds. All natural-history data and banding results indicate that 'Alala are permanently territorial and extremely site-faithful once they become breeding adults. Treetop sitting is undoubtedly related to territorial advertisement and defense, as in other corvids (Kilham, 1985a,b; McGowan and Woolfenden, 1989). Nests are usually built in the upper branches of 'ohi'as. In the current forest belt of the Kona District,
koa trees are restricted to a narrower and higher range than are 'ohi'as, usually above 1,500 m. Crows move short distances up the slopes of Hualalai and Mauna Loa into this region during the breeding season (Giffin, 1983). Seasonal movements to the lower elevations in the nonbreeding season appear to track the fruiting periods of the principal food plants (Giffin et al., 1987).
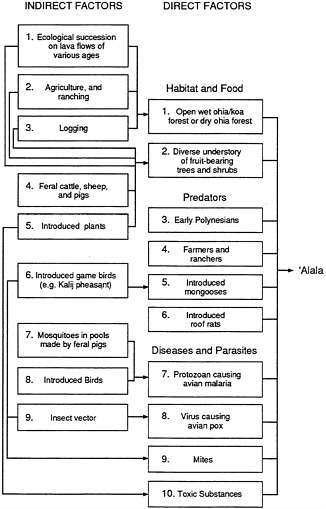
Figure 2.4 Factors influencing numbers of 'Alala
At least five indirect factors affect the suitability of both habitat and food sources for the 'Alala. The first is the impact of lava flows in the 'Alala's historical range, resulting in alterations to which the bird must continually adjust.
Other indirectly acting factors are habitat changes caused by agriculture, ranching, and logging. A prevailing hypothesis is that clearing of land by logging and, more important, by cattle and sheep grazing has so altered the available food supply that fecundity and survivorship have been severely affected (Banko and Banko, 1980; Giffin et al., 1987). It cannot be disputed that outright conversion of 'ohi'a-koa forest lands into open pasture has greatly reduced the total amount of forest habitat throughout the original range of 'Alala (Burr, 1984; Scott et al., 1986; Giffin et al., 1987). Widespread clear-cutting and grazing also fragmented the formerly continuous 'Alala population into numerous smaller subpopulations, each of which then became susceptible to additional perils (see Chapter 3 for a discussion of small populations). Thus, clearing of forests to produce grazing lands has clearly caused substantial reductions in the numbers of wild 'Alala (Banko and Banko, 1980; Giffin et al., 1987).
In the first half of the 1900s, substantial portions of the lower-elevation forests along the Kona and Ka'u coasts were developed into pastures, orchards, and settlements; by the 1940s, the 'Alala was essentially extirpated from these areas (Banko and Banko, 1980). Similarly, the disappearance of crows from most of their former range in central Kona was associated with heavy logging of valuable koa trees on the slopes of Mauna Loa from the mid-1920s to the late 1950s. Dry forests at low and middle elevations were significantly modified by the harvesting of sandalwood (Santalum spp.) in the 1800s, as well as by the later spread of exotic fountain grass (Pennisetum setaceum) and periodic wild fires. The last 'Alala disappeared from the dry forests on the north slope of Hualalai in 1964 (Tomich, 1971). Logging was often accompanied by the introduction of pigs, goats, sheep, and cattle, and grazing further modified the forest by preventing regeneration of koa trees and eliminating many native understory plants (Ralph and van Riper, 1985).
Much of the food of the 'Alala consists of fruits and invertebrates taken from the forest understory (Banko, 1976; Scott et al., 1986). Many of the understory plants and climbing vines are known to be important sources of fruit eaten by the 'Alala. Although 'Alala are omnivorous, fruits account for 33–66% of the adult diet (Sakai and Carpenter, 1990). Fruits of historical importance include 'ie'ie (Freycinetia arborea), 'olapa (Cheirodendron trigynum), 'oha-kepau (Clermontia spp.), oha (Cyanea spp.), mamaki (Pipturus albidus), pilo (Coprosma spp.), akala (Rubus hawaiiensis), and lama (Diospyros spp.). Seeds of manono (Gouldia terminalis) and ho'awa (Pittosporum hosmeri) are also used (Burr et al., 1982). The introduced banana poka (Passiflora mollissima) has become a major item in the diet. In addition, isopods, arachnids, and land snails are taken from understory trees and shrubs or gleaned from branches and clumps of vegetation in the upper canopy. The diet fed by 'Alala to their nestlings includes these items plus nestling songbirds and mice (Perkins, 1903; Munro, 1944; Sakai and Ralph, 1980; Sakai et al., 1986; Scott et al., 1986).
Another indirect factor that affects the habitat and food of the 'Alala is the common practice in Hawai'i of introducing feral ungulates into the forest. Both on private land and on land leased by the state, cattle, pigs, and sheep have occupied the forest belt for many years, often with considerable impact (Stone and Loope, 1987). Where the animals are overstocked, their foraging destroys native understory plants and deprives the 'Alala of many of its sources of fruit.
Since 1950, the state of Hawai'i has had a habitat-improvement program. Some areas that had been severely degraded by timber harvest and feral ungulates are recovering owing to the removal of ungulates, and that is the case in the Hualalai Reserve established in 1984. Cattle had been present in the wet forest of Hualalai for the previous 100 years. On the drier northern slopes in the Pu'uwa'awa'a Reserve, from which cattle and pigs have also been removed, the koa trees have begun to recover. Bird surveys in this area and in the Manuka Natural Area in south Kona, however, have continued to show declines in the numbers of 'Alala and other native birds (Giffin, 1983; Sakai et al., 1986; Giffin, 1990, 1991).
Ranching practices and the behavior of foraging cattle commonly result in reduction and even elimination of understory and subcanopy vegetation, and ground vegetation is replaced with introduced forage grasses and weeds. Even on ranches where mature trees are left in abundance, the resulting parklike environment often supports a greatly reduced diversity of plant species, in which natural regeneration of canopy trees is dramatically reduced (Baldwin and Fagerlund, 1943). Understory vegetation of the native forest included a rich diversity of fruit-producing species (e.g., Lobeliaceae and Rubiaceae). Many of those plants are eaten preferentially by grazing cattle and are uprooted by feral swine; when fully exposed and accessible to livestock and pigs, they become scarce and patchy. Even on the McCandless Ranch, where 'Alala are still found, fruiting species of the understory are widely scattered as individual plants, frequently growing out of fallen logs or tree crotches above easy reach of cattle (observations by this committee, January 1992). Under such conditions, these understory species have greatly reduced capacities to regenerate, recruit into dense stands, or recolonize areas from which they have been removed.
Biologists familiar with the plant life of the Kona slopes unanimously report a strong negative association between cattle and several of the most important food plants of the 'Alala. That is especially true for the 'ie'ie, which was shown in early twentieth century photographs to have covered huge areas of 'ohi'a forest in dense tangles. They were visited by 'Alala in great numbers, and crows were said to have nested directly among the vine tangles (Perkins, 1903). Evidence exists that several Hawaiian bird species, including 'Alala, were important pollinators of 'ie'ie (Cox, 1983); substantial loss of such important food sources could seriously alter time and energy budgets, and adversely affect reproduction and survival of the 'Alala.
As a direct result of logging and ranching, essentially no pristine example of the closed to moderately open moist 'ohi'a-koa and wet or dry 'ohi'a forest zone exists anywhere on the
Kona slopes. As of 1992, most of the remaining stands of intact humid forests are at the lower limits of 'Alala's modern range and consist of moist 'ohi'a forest growing on aa lava flows. These stands were used by 'Alala for late summer and winter foraging, but not as primary breeding habitat, which appears to be at an elevation of 1,000–1,600 m.
A dramatic correlation between loss of understory habitat and decline of an 'Alala population occurred in the late 1970s on Hualalai (Giffin, 1983; Carson, pers. comm. 1992). According to Giffin (1983; p.24), "one of the most important 'Alala nesting areas on Hualalai was partially cleared with bulldozers in the fall of 1976. Understory vegetation was removed and exotic pasture grasses were planted. Continuous grazing by livestock during the next 4 years further opened the forest floor. The number of active crow nests in that area decreased from four in 1977 to none in 1980."
The persistence of 'Alala on one particular ranch does not in itself mean that this property provides the only genuinely optimal habitat remaining within the historical range—only that the habitat there remains suitable and might be less degraded than other areas of former habitation. The McCandless Ranch lies directly at the center of the region where 'Alala historically was most common (middle elevations of central Kona). Because small populations are subject to extinction from chance events and because 'Alala are sedentary and unlikely to colonize new areas quickly, there could well be other suitable habitat areas within the historical distribution that are simply unoccupied.
Another complicating aspect is that 'Alala began to decline before 1900 in areas not dramatically altered by ranching, and throughout this century they have persisted in habitats where cattle ranching has been the dominant land-use activity. Numerous anecdotal reports and observations indicate that 'Alala in recent decades have preferred forest habitats that are broken by clearings and edges of habitats, of the kind typical on forested cattle ranches throughout middle and upper elevations of the Kona region (e.g., Berger, 1981). However, the overall record shows that past land-use practices have severely degraded the forest and in many areas removed aspects of the native forest that the bird might require, and the degradation is likely to have been a major factor in the overall decline of the 'Alala (Tomich, 1969, 1971; Banko, 1974; Burr et al., 1982; Giffin 1983; Sakai et al., 1986; Giffin et al., 1987). The fact that apparently suitable native forest is now unoccupied by 'Alala is difficult to explain (Banko, 1974) other than by suggesting that it occurs in fragments that might not be discovered by such a basically sedentary, traditional species or that the fragments are too small or too low in elevation to provide sustenance throughout the year.
Predators
Direct factors 3–6 in Figure 2.4 are predators that directly affect the numbers of 'Alala: early Polynesians, farmers and ranchers, introduced mongooses (Herpestes auropunctatus), and introduced roof (Polynesian) rats.
Polynesians used crow feathers in some of their ceremonies, but not to the extent that they used feathers of the brightly colored Honeycreepers (Drepanidinae spp.). European settlers are known to have hunted crows regularly for sport. When crows were common, they entered poultry yards, and many were shot by farmers as agricultural pests (Banko and Banko, 1980; Munro in Berger, 1981; Burr et al., 1982). Continuing, but unsubstantiated, reports of 'Alala shootings in recent years in Hualalai and other areas of north Kona despite legal protection (J.G. Giffin, pers. comm., 1991) point to the need for increased enforcement and public education. Considering the small size and relict distribution of the extant population, even a low mortality stemming from shooting could seriously hamper recovery efforts.
The Hawaiian Islands have no native mammals except bats, and there is no evidence that native hawks or owls have constituted a problem as predators of the 'Alala. But two introduced predatory mammals are abundant in the forest belt, and they take a heavy toll on the native fauna. Mongooses, introduced in 1883, are effective diurnal ground predators on young crows. Like other corvids, 'Alala fledglings leave the nest before they are strong fliers and spend some time on the ground or climbing the understory vegetation while still being fed by their parents. It is a period of high vulnerability to mongooses and presumably also feral cats (Tomich, 1969, 1971; Banko, 1974, 1976; Giffin, 1983). The McCandless Ranch, with some financial support from USFWS, has had a trapping program to control its mongoose population for the last few years (see Figure 2.5). An indirect factor that might contribute to predation on young 'Alala by mongooses is the presence in the forest of large numbers of the introduced Kalij Pheasants (Lophura leucomelana) and Turkeys (Meleagris gallipavo). Those birds are common on the McCandless Ranch (Engbring, 1990; committee observations, January 1992) and elsewhere within the 'Alala's former distribution (Lewin and Lewin, 1984); they and their eggs and young are important prey of the mongoose. Their abundance might help to support a large mongoose population, which in turn could increase the juvenile mortality of 'Alala.
The introduced roof rat is also a common predator in the forest belt (Tomich, 1969). This nocturnal arboreal predator can take eggs and nestlings. Partial brood loss from nests should probably be attributed to predation by rats (Tomich 1969, 1971). No evidence supports or refutes the idea that introduced predators are the major cause of the decline in the wild 'Alala population.
Diseases and Parasites
Factors 7–10 in Figure 2.4 are diseases, an ectoparasite, and potentially toxic substances. Two avian diseases—pox and malaria—introduced to Hawai'i after colonization by Europeans appear to have played an important role in the decline of many endemic birds (Warner, 1968; van Riper et al., 1986). Avian pox is caused by an arbovirus and is transmitted by direct contact with an infected bird, by secondary contact with a contaminated object, or mechanically by vectors, such as mosquitoes and biting flies (van Riper and van Riper, 1980; Cavill, 1982). Birds with pox usually develop lesions at the immediate site of infection in the mouth and upper
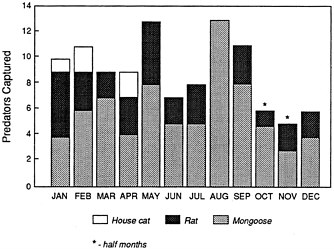
Figure 2.5 1991 Predator Trapping Results, McCandless Ranch. Information from P. Simmons (pers. comm. 1992).
respiratory tract or on the integument on the bill or legs or around the eye. Exposure to the disease is generally believed to confer immunity, but young birds seem particularly prone to severe, debilitating lesions. For example, in Galapagos Mockingbirds (Nesomimus p. parvulus), Vargas (1987) found a higher incidence of active lesions in nestlings and juveniles than in adults and higher post fledgling mortality in infected than noninfected birds. Nestling and fledgling 'Alala have been observed with pox-like lesions on a number of occasions (Banko and Banko, 1980; Giffin, 1983), and pox has been implicated in contributing to mortality in young 'Alala (Jenkins et al., 1989). Avian pox apparently was widespread in the native Hawaiian avifauna, including 'Alala, by the turn of the century (Perkins, 1903), and it persists today (Ralph and van Riper, 1986; Scott et al., 1986). Although the incidence rate in wild 'Alala is unknown, Jenkins et al. (1989) detected precipitating antibodies against fowlpox virus in blood sera collected from three adults (100% incidence), and two of five (40% incidence) nestlings. All three of the adult samples also reacted to the Hawaiian poxvirus isolate.
The blood protozoan that causes avian malaria (Plasmodium relictum capistranoae) is believed to have arrived in Hawai'i in the 1920s with introduced passerine birds (van Riper et al., 1985; Stone and Loope, 1987), and it has been identified in a variety of endemic Hawaiian birds, including the 'Alala (cf. Banko and Banko, 1980; Jenkins et al., 1989). As with avian pox, there is relatively little information on the rate of occurrence of avian malaria in 'Alala.
Jenkins et al. (1989) detected a heavy avian malaria infection in only one of 10 nestlings and in none of six adults. However, these authors noted that young birds are more susceptible to infection than adults, and that the debilitating effects of malaria would be expected to increase both direct and indirect mortality in nestlings and fledglings. The principal vector for this parasite is the introduced night-biting, ornithophilous mosquito, Culex quinquefasciatus, although another introduced species of mosquito, Aedes albopictus, can also act as a vector (Scott et al., 1986). The vulnerability of nestlings to malaria is probably enhanced by the fact that mosquito populations generally peak during the 'Alala's breeding period (Jenkins et al., 1989). Results of inoculation experiments conducted by van Riper et al. (1985) indicate that native Hawaiian birds are much more susceptible than introduced species to mortality from malaria, and Scott et al. (1986) have noted the negative relationship between the elevational distribution of mosquitoes and that of the surviving populations of native birds (see also Warner, 1968). Moreover, rooting by feral pigs, and stock ponds and troughs for cattle, provide sites for mosquito breeding, and hence the spread of malaria. The results lead us to conclude, with Ralph and van Riper (1985) and others, that avian diseases—particularly avian pox and malaria—have probably been a major factor in the decline and distribution of the 'Alala.
Banko (1974) found an 'Alala fledgling that was heavily infested with the introduced northern fowl mite (Ornithonyssus sylviarum). Although the significance of this parasite is unknown, it has been reported to cause mortality in young poultry (Banko, 1984). Heavy mite infestations could result from weakness or disease brought about by other factors. The extent to which introduced birds, such as the Kalij Pheasant, serve as reservoirs for disease and ectoparasites is poorly known, but certainly merits further investigation. The possibility that the seeds or fruits of introduced species of plants, such as the banana poka, that are now regularly exploited by the crow might contain potentially harmful substances also warrants further study.
Estimates of Population Viability and Time to Extinction
Approaches
The committee was charged with estimating the ''minimum viable population for survival" of the 'Alala. That cannot be done, in part because no set population size is sufficient to ensure survival within the changing, fragmented landscape of the Hawaiian forests. We can only estimate probabilities of extinction as a function of population size. The number of birds is too low for long-term survival to be possible without active management.
What follows are an evaluation of the species' current demography in the wild and an estimate of its time to extinction if there is only passive management.
Demographic Analyses
Given what we know about the demographics and approximate size of the wild population of the 'Alala, we built a deterministic model of the population dynamics of the 'Alala. Such a model, by definition, lacks the chance events that we know can doom small populations. Nonetheless, the exercise provides clues about the decline of the 'Alala in most areas and about its persistence on the McCandless Ranch.
Limited banding data at the McCandless Ranch (where the numbers appear not to have changed much in a decade) demonstrate that adults are long-lived, with annual survivorship approaching 90% (Banko and Banko, 1980). The species has a relatively low natural reproductive rate, with protracted periods of nestling development and fledgling dependence. As previously noted by Banko and Banko (1980), the 'Alala appears to have a smaller average clutch size than other members of Corvus, although current comparative information is primarily restricted to mainland species (Table 2.1). The incubation period of the 'Alala is similar to those of other members of Corvus, but the fledging time seems to be slightly longer than expected (Table 2.2). Pairs appear to occupy permanent ("traditional") territories of comparatively small size (about 1 km2), which also contain nonbreeding offspring from the breeding seasons of the preceding 1 or 2 years. The social roles of the nonbreeding offspring are unknown; however, evidence suggests that they are not involved in cooperative rearing of young, which occurs in a number of corvid species (Woolfenden, 1973; Kilham, 1984a,b,c; Woolfenden and Fitzpatrick, 1984). Nonbreeders might help a breeding pair to defend their territory against conspecifics. There is, however, little evidence of general reproductive failure.
Several workers (Banko, 1976; Burr et al., 1982; Giffin, 1983) have concluded that the rate of recruitment of young birds into the adult population is unusually low in the 'Alala. Although recruitment might have been higher under the pristine conditions in which the species evolved (which included no mammalian predators) than it has been in recent decades, the data in Table 2.3 indicate that the 'Alala's reproductive success is somewhat lower than that of temperate corvids. Comparative data for other tropical corvids, however, do not exist.
Existing information on age structure and age distribution of the 'Alala is in Box Table 1. It includes data from Banko and Banko (1980), who reported 72 known nesting attempts; data on 27 banded birds spanning a period from 1973 until 1980, when banding ceased, plus resightings of these birds; and the census data in Figure 2.3. The observations, although limited in scope, allow us to estimate demographic measures, given two precautions. The first is that, until April 1992, no sustematic observations were made for nearly a decade (during the 1980s), because the remaining birds were on the McCandless Ranch, where access was not permitted. The second is that the samples of banded birds are very small. Nonetheless, many of our conclusions are consistent with observations of 'Alala behavior and population biology during the 1970s and 1980s, both on and off the McCandless Ranch.
Table 2.1 Average clutch size and range of clutch sizes reported in various Corvus species
|
Species |
No. Clutches |
Mean |
Range |
Location |
Source |
|
Corvus hawaiiensis |
4 |
3.5 |
2–5 |
Hawai'i |
Banko and Banko, 1980 |
|
|
11 |
2.2 |
1–4 |
Hawai'i |
Temple and Jenkins, 1981 |
|
C. caurinus |
189 |
4.0 |
1–4 |
Canada |
Richardson et al., 1985 |
|
C. cryptoleucus |
|
|
3–8 |
N. America |
Bent, 1946 |
|
C. brachrhynchus |
|
4.4 |
2–6 |
N. America |
Emlen, 1942 |
|
C. frugilegus |
182 |
4.2 |
1–7 |
England |
Holyoak, 1967 |
|
|
151 |
4.3 |
2–7 |
England |
Lockie, 1955 |
|
C. monedula |
80 |
4.2 |
1–6 |
Switzerland |
Zimmermann, 1951 |
|
|
29 |
5.1 |
3–6 |
Finland |
Antikanen, 1978 |
|
|
233 |
4.4 |
1–6 |
England |
Lockie, 1955 |
|
|
431 |
4.3 |
2–7 |
England |
Holyoak, 1967 |
|
C. corone |
28 |
4.1 |
— |
Scotland |
Yom-Tov, 1974 |
|
|
49 |
4.1 |
2–6 |
Scotland |
Picozzi, 1975 |
|
|
168 |
3.9 |
1–7 |
England |
Holyoak, 1967 |
|
|
74 |
4.5 |
— |
W. Germany |
Wittenberg, 1968 |
|
|
39 |
4.6 |
3–6 |
Finland |
Antikanen, 1978 |
|
|
198 |
4.7 |
1–7 |
Finland |
Tenovuo, 1963 |
|
|
39 |
4.6 |
3–6 |
Norway |
Slagsvold et al., 1984 |
|
C. capensis |
7 |
3.9 |
3–4 |
S. Africa |
Skead, 1952 |
|
C. orru |
52 |
4.8 |
1–6 |
Australia |
Rowley, 1973 |
|
C. bennetti |
21 |
4.2 |
1–6 |
Australia |
Rowley, 1973 |
|
C. coronoides |
137 |
4.4 |
1–6 |
Australia |
Rowley, 1973 |
|
C. mellori |
570 |
4.2 |
1–6 |
Australia |
Rowley, 1973 |
|
C. tasmanicus |
11 |
4.2 |
3–6 |
Australia |
Rowley, 1973 |
|
C. corax |
45 |
6.0 |
3–7 |
N. America |
Stiehl, 1985 |
|
|
67 |
5.2 |
3–7 |
England |
Holyoak, 1967 |
|
|
245 |
4.5 |
— |
Wales |
Newton et al., 1983 |
Table 2.2 Incubation lengths and fledging ages reported in various Corvus species
|
Species |
Incubation Lengtha days |
Fledging Ageb days |
Source |
||
|
Corvus hawaiiensis |
19–20 |
39–43 |
Banko and Banko, 1980 |
||
|
C. kubaryi |
~ 21 |
~ 33 |
R.E. Beck (pers. comm.) |
||
|
C. caurinus |
18–19 |
30–35 |
Butler et al., 1984 |
||
|
C. brachyrhynchos |
16–21 |
26–35 |
Emlen, 1942; Bent, 1946; Chamberlain-Auger et al., 1990 |
||
|
C. monedula |
17–18 |
~ 18 |
Zimmermann, 1951 |
||
|
C. frugilegus |
16–18 |
32–39 |
Lockie, 1955; Richards, 1973; Goodwin, 1976 |
||
|
C. capensis |
18–19 |
~38 |
Skead, 1952; Goodwin, 1976 |
||
|
C. corone |
17–21 |
30–36 |
Holyoak, 1967; Wittenberg, 1968; Coombs, 1978 |
||
|
C. macrorhynchus |
17–19 |
|
Goodwin, 1976 |
||
|
C. orru |
19–20 |
38–48 |
Rowley, 1973 |
||
|
C. bennetti |
~17 |
29–31 |
Rowley, 1973 |
||
|
C. coronoides |
19–21 |
40–45 |
Rowley, 1973 |
||
|
C. mellori |
19–20 |
34–41 |
Rowley, 1973 |
||
|
C. albus |
18–19 |
~43 |
Lamm, 1958; Goodwin, 1976 |
||
|
C. ruficollis |
20–22 |
37–45 |
Goodwin, 1976 |
||
|
C. corax |
18–21 |
38–44 |
Gwinner, 1965; Stiehl, 1985 |
||
|
a Although some of reported variation in incubation length probably reflects attentiveness of individual females, large proportion of variation is probably due to method. Accurate determination of incubation in the field is complicated by female corvids' period before oviposition and beginning of incubation usually before clutch is complete, so that eggs hatch asynchronously (cf. Lockie, 1955; Wittenberg, 1968; Butler et al., 1984). Because most workers have used standard method of determining incubation length, time between laying and hatching of last egg (Nice, 1954), some degree of inaccuracy is probable (see Greenlaw and Miller, 1983). b Many corvids leave nest several days before fully capable of flight. Additionally, observer-caused disturbance is known to cause premature fledging (cf. Butler et al., 1984). |
|||||
Table 2.3 A comparison of reproductive success reported in various Corvus and other species
|
Species |
% Eggs Hatched |
% Nestlings Fledged |
% Nests Successful |
Fledglings per Nest |
Source |
|
Cyanocitta stelleri |
92 |
27 |
31 |
- |
Ligon, 1971 |
|
C. coerulescens |
54 |
56 |
53 |
1.5 |
Woolfenden, 1978 |
|
Psilorhinus morio |
- |
76 |
80 |
3.0 |
Lawton and Guinton, 1981 |
|
Pica pica |
80 |
- |
75 |
- |
Evendon, 1947 |
|
|
- |
39 |
- |
1.6a |
Husby, 1986 |
|
|
- |
- |
41–78a |
1.1–1.8a |
Dhindsa and Boag, 1990 |
|
|
- |
- |
61 |
2.5 |
Buitron, 1988 |
|
Corvus caurinus |
74 |
44 |
78 |
1.3 |
Butler et al., 1984 |
|
C. brachyrhynchus |
- |
33 |
- |
- |
Emlen, 1942 |
|
|
- |
- |
- |
2.0 (0.8–3.3) |
Chamberlain-Auger et al., 1990 |
|
C. monedula |
- |
73 |
- |
2.9b |
Lockie, 1955 |
|
C. frugilegus |
85 |
72 |
- |
2.0–3.7b |
Owen, 1959 |
|
|
- |
71 |
- |
2.7b |
Lockie, 1955 |
|
C. corone |
46 |
58 |
43 |
1.2a |
Yom-Tov, 1974 |
|
|
90 |
19 |
- |
- |
Tenovuo, 1963 |
|
C. coronoides |
72 |
48 |
54–86 |
1.53 (0.9–1.8) |
Rowley, 1973 |
|
C. mellori |
75 |
47 |
70 |
1.99 (0.2–2.0) |
Rowley, 1973 |
|
C. orru |
70 |
31 |
60 |
1.0c |
Rowley, 1973 |
|
C. bennetti |
72 |
5 |
14 |
0.1c |
Rowley, 1973 |
|
C. corax |
- |
70 |
- |
2.3 (2.2–2.5) |
Stiehl, 1985 |
|
C. hawaiiensis |
- |
- |
66 |
0.8 (0.5–1.3)d |
Banko and Banko, 1980 |
|
|
- |
- |
43 |
0.7 (0.5–0.8)e |
Temple and Jenkins, 1981 |
|
a Data from control nests only. b Unsuccessful nests excluded from calculation. c Data collected during year of severe drought. d Calculations based on 50 nests located and observed during 1973–1978. e Calculations based on 16 nests located and observed during 1979–1980 |
|||||
On the basis of data obtained through 1980 (Banko and Banko, 1980), the age at first breeding appears to be 2 years. Of four fledglings later found nesting, three were 2 years old, and the other was 3 years old. It is not known whether the 3-year-old breeder (Hualalai bird 012) was missed as a breeder at age 2. Young 'Alala accompany adults for up to a year (maybe more) after fledging, and delayed breeding is typical of such group-living corvids. Therefore, it is extremely unlikely that 1-year-old 'Alala breed.
By assuming 2 years to be the age at first breeding, we can represent the demography of the 'Alala as a three-stage model with several parameters (see Box Figure 1). In addition to the number of young produced per breeding adult, three survivorship parameters are estimated: adult survivorship (i.e., the proportion of the population of breeding adults surviving from year t to year t + 1, for t = 2), juvenile survivorship (proportion of recently fledged birds surviving to the end of their first year), and yearling survivorship (from 1 year old to 2 years old).
Average adult survivorship can be estimated in two ways. In the observation weighted method, birds are assumed to have died immediately after they were last seen. The number of year-to-following-year survivals is averaged across all the birds. Thus in Box Table 1, adult 025 survived 12 of a possible 12 year-to-following-year intervals. In contrast, bird 006 was not seen after reaching 2 years of age: as an adult, it survived zero year-to-following-year intervals of a possible one. Pooling these data for all birds establishes the minimum estimate of average survivorship; bird 006, for instance, might have survived several years after last being seen, but might not have been seen clearly enough to identify its color-bands.
The second method for estimating survival is a time-weighted method. Of five birds known to have been adults on the McCandless Ranch, two were alive after 9 years. If yearly survival is s, then 2-year survival is s2, 3-year survival s3, and so on. Hence, s9 = 2/5, and hence s = 0.90.
For estimating survivorship and reproduction of 'Alala, we separated the small pool of data on birds on the McCandless Ranch (where the population was approximately stable) from the pool of data on birds at Hualalai and Honaunau (where 'Alala were precipitously declining). The results are different from that of most previous authors regarding the proximate causes of 'Alala decline from 1974 through 1982: reproduction and recruitment were nearly normal among breeding pairs, but adult survival and nesting attempts were exceedingly low, except on the McCandless Ranch.
Fledgling production per nesting pair averaged 0.92 on McCandless Ranch and 0.87 elsewhere (Box Table 4). Juvenile survival was 50% on the McCandless Ranch and 43% elsewhere (Hualalai, Honoaunau and south Kona), and yearling survival was 90%+ everywhere (Box Table 2). For modeling purposes, we use 50% and 90% as juvenile (first-year) and yearling (second-year) survivorship, respectively. None of these estimates of reproduction and subadult survival is atypical of a healthy corvid population, especially given the small average
clutch size and extended nestling period of 'Alala. Yearling survival (between 1 and 2 years) is often the highest of any year, because the birds have avoided the potentially fatal mistakes of inexperience, yet do not have the responsibilities of parenthood. Those numbers also match observations, both on and off the McCandless Ranch, that show that the birds are breeding successfully and that dependent young often accompany adults throughout the year. However, an extremely important observation is that fledgling production drops to 0.66 away from the McCandless Ranch when all pairs that failed to nest (or whose failed nests went unobserved) are added to the sample (Box Tables 3 and 4). Even off the McCandless Ranch, however, there was no "general reproductive failure" of the kind postulated by many previous authors (Banko and Banko, 1980; Burr et al., 1982; Burr, 1984; Jenkins et al., 1989). Some 'Alala were breeding, often successfully, even during the period when the population was crashing.
The sharpest contrast between the McCandless Ranch and elsewhere appears to be in adult survival (Box Table 2). On the McCandless Ranch, annual survival was between 80% and 90%, which is typical of corvids. At Hualalai and Honaunau, the minimum estimate of adult survival is 66%. A time-weighted estimate is more difficult, because birds were banded over a 6-year period. However, only one of 10 'Alala (Hualalai bird 025) is known to have survived more than 2 years as an adult! Seven 'Alala were alive as young adults in 1980. Had they survived, all seven could have been recorded during a thorough census of these regions in 1986. Only bird 025 was alive after these 6 years; the time-weighted estimate of annual survival is about 72%.
Except on the McCandless Ranch, an average of one-third of all remaining adult 'Alala died or disappeared annually between 1975 and 1986. This conclusion is entirely consistent with the observed rate of decline in 'Alala numbers from census counts at Hualalai and Honaunau during that period (Figure 2.3). This adult survivorship is extremely low for a corvid.
Florida Scrub Jays (Aphelocoma c. coerulescens) share many of the social and demographic characteristics of 'Alala. They are sedentary, omnivorous corvids living in permanently defended territories, and they exhibit delayed reproduction (Woolfenden and Fitzpatrick, 1984, 1990). Offspring remain with the parents for one or more years, creating a "standing crop" of nonbreeding individuals that constantly interacts with the population of breeders and survive at rates different from those of the breeders (Fitzpatrick and Woolfenden, 1986). Annual survivorship of breeding jays averages about 80% and has dropped below 70% only once in 23 years, during a catastrophic epidemic (Woolfenden and Fitzpatrick, 1991). Adult survival between 80% and 90% is reported among Santa Cruz Island Scrub Jays (Aphelocoma coerulescens insularis) (Atwood et al., 1990) and Mexican Jays (Aphelocoma ultramarina) (Brown and Brown, 1990), both of which also exhibit delayed breeding. Recently, adult survival of American Crows (Corvus branchyrhynchos) was found to exceed 90% near Los Angeles, where first-year birds do not appear to breed (C. Caffrey, pers. comm., 1992). No corvid in the world is known to have adult mortality anywhere near as high as that exhibited by 'Alala during the 1970s and early 1980s away from the McCandless Ranch.
|
Box Table 1: Banding records and resightings  Notes: 1. Band numbers are the last three digits of the eight-digit U. S. Fish and Wildlife Service band. 2. Key to symbols: * bird born in year; A bird known to be adult in year; B bird known to have bred in year; - bird known to be alive in year; # bird known to be dead in year. Column of # for 1986 at Honaunau and Hualalai reflects counts in these areas that showed only one bird alive (band number 025). Data from Banko and Banko, 1980; Giffin, 1991. |
|
Box Figure 1. On the basis of banding records, it appears reasonable to group the 'Alala into three age groups: juveniles (0–1 year old), yearlings (1–2 years old), and breeding adults (=2 years old). According to census data, it seems necessary to separate the populations on the McCandless Ranch, which have remained constant, from those elsewhere, which have declined. Nesting information (from Banko and Banko, 1980) plus banding information (supplied by Giffin, 1991) allow estimation of the relevant demographic parameters:
To predict the likely deterministic dynamics of these populations, let Xt, Yt, and Zt be the numbers of juveniles, yearlings, and adults, respectively. The equations describing the McCandless situation above are:
For the equations at Hualalai and Honaunau, comparable coefficients can be substituted. These equations can be investigated analytically, but for simplicity we show the results of two simulations below. As can be seen in Box Figure 2, barring unforeseen demographic accidents or chance environmental disturbances, the McCandless population is predicted to show a very slow increase in population size; the populations elsewhere are predicted to decline rapidly (although not as rapidly as actually observed.) The data and their interpretation follow in Box Table 2. 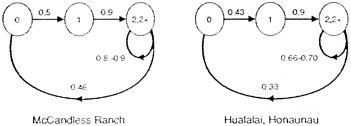 |
|
Box Table 2: Analysis of banding records and survival Juveniles are ≤1 year old, yearlings 1–2 years old, and adults =2 years old. Juvenile Survival: McCandless Ranch Juvenile survivorship: 3 of 6 survived the first year = 0.50 Juvenile Survival: Honaunau and Hualalai Juvenile survivorship: 6 of 14 survived the first year = 0.43 Yearling Survival: all sites Yearling survivorship: 6 of 6 survived the year at Honaunau and Hualalai, and 3 of 6 survived the year at McCandless, resulting in a survivorship of 1.00 at both places. As explained in the text, such a value is not unexpected. However, given that 100% survival is unlikely, we have suggested that it might be lower. A survivorship value of 0.9 is used in Box Figure 1.
Adult survivorship: 22/27 = 0.81. As indicated in the text, this is a minimum value, although, given the rapid disappearance of birds from these sites, it cannot be far below the actual value. Using the time-weighted method (see text), we estimated the adult survivorship as 0.9.
Adult survivorship: 19/29 = 0.66. Again, this is a minimum value, and the time-weighted method yields a value of 0.7. |
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
Analysis of nesting data Nesting data are drawn from Banko and Banko (1980), who calculated a fledgling rate of about 0.8 bird per pair per year. Three important, related ideas emerge from their raw data: 1. The value of 0.8 is the average value that combines all nests found, including renesting and nests that were found very late in the nesting cycle. Nests that failed before being encountered were not included in any way. (There is a statistical procedure for estimating true nest success from such data with the observed daily probabilities of failure. We did not do this calculation, because we felt that it would not alter the results by very much.) Renests should not be counted separately in calculating the observed production of fledglings per year. Box Table 4 shows that from 1970 to 1978 in all areas, including McCandless Ranch, 58 fledglings were produced by 67 pairs that attempted to nest. This yields a value of 0.87 fledgling per pair (0.43 per bird). 2. Of more concern is that numerous records exist during the 1970s for which no nesting was documented, despite considerable searching efforts. It is vital to correct our estimates to incorporate these unsuccessful pairs, because those which did produce fledglings in unfound nests were incorporated into the overall summary (as ''unobserved nests") in Banko and Banko (1980). The pairs that never nested or whose unsuccessful nests were never found are not counted anywhere. We catalogued these pairs as closely as possible with the documentation in Appendix I of Banko and Banko (1980), aided by the narrative on pages 10–25 (see Box Table 4). Our initial estimate of 0.87 fledgling per pair is therefore almost certainly too high. Virtually all the nonreproductive pairs were encountered at sites where the 'Alala has been known to breed. In some cases, the observers missed a year, and nests were located at the same sites they were in both the previous and subsequent years. Many represented the final year of documented territorial occupancy before disappearance. We were conservative in counting only the birds and places where permanent occupancy was strongly suspected. Had these pairs actually produced a fledgling in those years, they would have been logged as reproductive and listed in Banko and Banko's (1980) Table 1. As indicated in Box Table 4, the corrected fledgling production is 58 fledgling in 83 pair-years, or productivity of 0.70 fledgling per pair. That value should be considered the overall average productivity of the 'Alala islandwide. It is on the low side for the family of Corvidae (see Table 2.4), although data on other species are few. 3. Perhaps most important is that, on the McCandless Ranch, all pairs attempted to breed each year. From 1972 through 1978 on the McCandless Ranch, there were 11 fledglings during 12 pair-years—an average productivity of 0.92 per pair. If those fledglings are subtracted from the total islandwide production, average production for the areas off the ranch was 47 fledglings in 71 pair-years, or 0.66 fledgling per pair. In sum, we estimate the average fledgling production per individual adult per year to be 0.66/2 = 0.33 in Hualalai and Honaunau and 0.92/2 = 0.46 on the McCandless Ranch. Those are the values found in Box Figure 1. |
|
Box Table 3: Nonreproductive pairs on territories in Kona District, 1971–1978
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
Box Table 4: Fledgling production on territories in Kona District, 1970–1978 All areas, including McCandless Ranch
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
It is possible that reduced reproductive success, and especially the high frequency of failure of pairs to attempt nesting at Hualalai and at Honaunau, can be attributed to high adult mortality. Especially high mortality among established breeders has the effect of scrambling the few remaining territory holders each year, forcing re-pairing among unfamiliar and inexperienced birds (Woolfenden and Fitzpatrick, 1984, 1991). In a stable population of sedentary corvids, mateless territory holders are quickly discovered by dispersing prebreeders or other mateless breeders and re-pairing is rapid. In fragmented populations, however, solo birds, especially orphaned nonbreeders, might leave the territory in search of other occupied territories and conspecifics. Such a scenario among 'Alala is supported by the frequent observations of wandering, extra-limital birds during the late 1970s (Banko and Banko, 1980).
It is not clear why adult survivorship on the McCandless Ranch has remained high or why it was lower elsewhere. It should be noted that these estimates of adult survival are strongly influenced by the one or two longest-lived individuals at each location (Box Table 2). Such sensitivity calls attention to the uncertainty inherent in estimating this life-history parameter, and it also suggests the vulnerability of these populations to demographic accidents or chance events. Possibly, by chance alone, a few birds with enough "know-how" (or immunity from disease) to escape the most common causes of death elsewhere are on the ranch. One such bird persisted on Hualalai (025) for at least 12 years, while an average of one-third of her compatriates disappeared each year. Such a possibility, of course, is a worst-case scenario: unless their numbers are augmented soon, the death of such special, experienced birds would be accompanied by the final extinction of 'Alala in the wild.
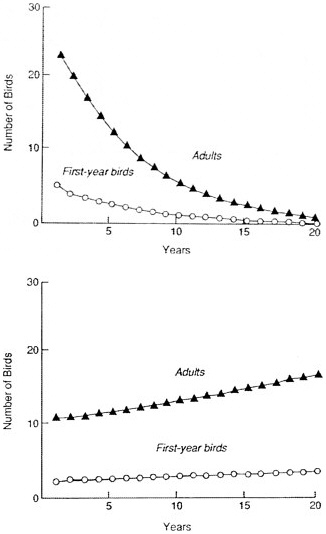
Population projections based on our estimates of reproduction and first-year, second-year, and adult survival replicate the observed decline (effective extirpation) of non-McCandless 'Alala (Box Figure 2). Our analysis suggests that the wild population of the 'Alala on the ranch could be stable or even increasing very slightly. The McCandless Ranch could still be a "net exporter" of 'Alala, but any dispersing bird would have to colonize areas where high adult mortality is known to have occurred. The analysis also allows us to predict that, even if it is at equilibrium or exporting a few 'Alala, the McCandless Ranch population will remain at low numbers for decades (Box Figure 2) and cannot exceed a few territories (Figure 2.6), because the social system and ecological requirements of 'Alala prevent it. Given the low numbers, demographic accidents alone could easily extirpate all remaining 'Alala in just a few years, and the risk of chance extinction will remain high for decades.
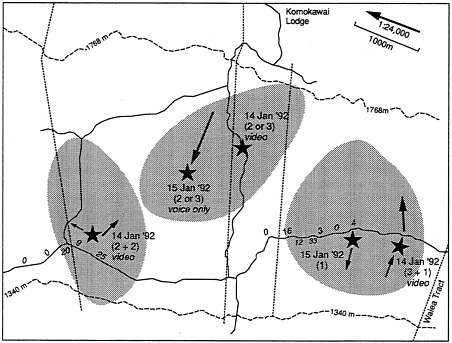
Figure 2.6 Map of the portion of McCandless Ranch where 'Alala are regularly recorded, according to ranch personnel. Thin solid lines are jeep trails; thin dotted lines are fences (these and contour lines traced from U.S. Geological Survey quadrangle 'Kaunene'). Small numbers along jeep trails indicate numbers of 'Alala encounters recorded by McCandless Ranch trapper throughout 1991 (N = 128 encountered by sight or sound). Trapper noted 1 (N = 82), 2 (N = 36), 3 (N = 10), or 4 (N = 3) 'Alala in each encounter—never more than 4 at one site—and committee had a maximum of 10 total 'Alala records on 14–15 January 1992. Thick solid lines encircle three hypothetical 'Alala territories consistent with all observations by committee and ranch personnel. No evidence of more than three territories on McCandless Ranch during the 1970s or 1980s exists. It is possible that only two territories exist and that records suggesting the middle territory (including McCandless personnel records at sandalwood exclosure downslope from Komokawai Lodge) represent birds from the other two territories.
Times to Extinction of Corvids on Islands
There are two broad approaches to estimating time to extinction. The first is used when a population is declining continuously and involves a simple extrapolation to the point of
extinction. For instance, extrapolation of 1974–1979 data on the population of 'Alala on Hualalai would have predicted the population's extinction by 1982 or 1983 (see Figure 2.3.a), and indeed 1983 was the last year in which the species bred in that area. Such an approach is not applicable to the small population on the McCandless Ranch, where, in contrast with its behavior everywhere else, the species has not obviously declined in the last decade.
The second approach to predicting time to extinction asks what determines the time to extinction of such small populations that might appear to be stable; it is the approach developed in this section. Two kinds of evidence are examined: from other very small populations of corvid species that have been counted for long periods and from an examination of the known features of the 'Alala's demography (analyzed above).
First, however, we review some of the background on predicting times to extinction. Theoretical studies recognize two kinds of extinction processes that concern small populations. The first, demographic accidents, stem from the vagaries of birth and death. For example, a population consisting of only a few breeding pairs has a high risk of demographic accidents: all the young born in one generation might be of the same sex, or all the adults might die in one generation for independent reasons. Demographic risks quickly diminish as the population increases. In nature, an additional risk comes from events that are external to the population. Unpredictable environmental disturbances can cause many animals to die—a forest fire, a hot summer, a cold winter, an unusual influx of predators, die-off of prey, a hurricane, and so on. Experience with small animal and plant populations shows that small populations, even if they are not declining, are extremely vulnerable to extinction caused by such chance events. The resulting fluctuations in population size impose a considerable risk of extinction, especially for populations that have high year-to-year variations in numbers. Consideration of the magnitude of size fluctuations and the additional risk that they impose can be an important issue in deciding on a management strategy, as they frequently represent the most important factor in determining extinction probabilities (Goodman, 1987).
In addition to the theoretical models, empirical methods can be used to predict how long small populations are likely to last before they become extinct. In the present case, data are available on very small populations of corvid species elsewhere in the world. In addition, other kinds of information might be available on the species of interest:
-
Population data. How many, where, and how these change over time.
-
Distributional data. Whether the species is only in habitats larger than some specified size.
-
Life history data. Age- or stage-specific fecundity and survival.
Even if a great deal is known about a species, we might not know the right things. What is known—and particularly what is not known—dictates the analytical methods that can be used. For example, all that is known about some warblers in the eastern United States is that they are
typically absent from woodlots of a particular size—that allows prediction of population if the patterns of forest fragmentation are known. For other species, information about life-history characteristics might permit age-dependent or stage-dependent modeling. These modeling approaches have some severe drawbacks: To be compelling, the predictions need both estimates of parameters and estimates of their variability. For example, for a long-lived species like the 'Alala, stage-dependent models often find adult longevity to be the critical parameter in determining time to extinction. The models merely predict that the population will last until the adults die in some unexpected, episodic way—e.g., as a result of a bad storm, a cold winter, a major forest fire, a drought, or shooting. Without knowledge of the nature and frequency of such deaths, predictions of that type are of limited use.
The second kind of evidence we examine involves estimates of times to extinction for the 'Alala derived from observed times to extinction of very small populations of other species of corvids. There is an extensive theoretical literature on times to extinction, but few data. Entire populations must be counted repeatedly for long periods, and analyses require a large set of such counts. The best estimates of how long small populations of corvids survive come from yearly counts made on islands off the coasts of Great Britain and Ireland. Analysis of those data leads to the conclusion that the median time to extinction for a maximum density of three breeding pairs is 8 years. (See Appendix A for a detailed summary of the derivation of estimates of time to extinction for those small island populations.)
The effective population size of the 'Alala on the McCandless Ranch is likely to be three or fewer breeding pairs. There is the estimate of "3 young produced in the wild in 1991" provided by Peter Simmons, manager of the McCandless Ranch. A maximum of three pairs is also consistent with the observations of this committee in January 1992, and 3 nests were sighted in April 1992 (J. Engring, pers. comm, 1992). A comparison with the island data noted in Appendix A suggests that as long as the 'Alala population on the McCandless Ranch remains at its current size (and does not increase), it has a 50% chance of extinction within a decade (and each subsequent decade).
Suppose that the 'Alala on the McCandless Ranch could be considered a "healthy" small population, i.e., the population is not suffering an inexorable decline and has numbers that remain approximately constant from year to year. Assume that habitat loss and predation are no longer factors, because of the protection afforded the species on the McCandless Ranch; that there is no decline attributable to disease; and so on. In short, the population has now stabilized. The British island populations, too, are "healthy" in this sense. Long-term population data from the British islands show that all but one of the species of crows are increasing; the exception is the Chough, which suffered local declines on Great Britain (now reversed), but tended to be unchanged on the small islands. Despite that favorable news, even ''healthy" populations become extinct because of chance events. So the British island data are comparable with the best-case scenario for the 'Alala.
Although comparisons with other tropical island endemic species would be valuable, there are few precise data for corvids. Craig (1991) has called attention to the persistence over many decades of small populations (fewer than 20 pairs) of island birds in New Zealand, and Jones and Owadally (1988) summarize examples on Mauritius. On the basis of current knowledge, however, the British crows seem to be the best available models for the 'Alala for estimating the likelihood of extinction resulting from demographic accidents. The likelihood of chance extinction is known to depend generally on the dispersal abilities of the species, how long individuals live, how many young the species produces per year, the magnitude of year-to-year changes in the population, the social behavior of the species, and most important, the first-observed size of the population. The 'Alala population on the McCandless Ranch is isolated, whereas the island populations noted above are not isolated. Indeed, one reason why some of the British island populations last so long might be that the populations received immigrants from the mainland. The immigrants might also rescue a population genetically—providing an influx of genes that helps to overcome the dangers of inbreeding. (A more thorough discussion of inbreeding is found in Chapter 3.) That difference in isolation makes the estimates of how long the populations last very optimistic—truly isolated populations might not last as long.
Corvids, in general, live a long time (see records provided by Clapp et al., 1983), and these British island populations have some of the lowest year-to-year variations in numbers ever observed in any species anywhere (Pimm, 1991). Moreover, tropical birds tend to live a long time, to reproduce slowly, and (on the basis of very few data) supposedly to vary less from year to year than temperate species. Long life and low variability reduce the chance of extinction; slow reproduction increases it. In short, there is nothing obviously special about the basic dynamics of the 'Alala versus other corvids. Therefore, we think that it is unlikely that the island data suggest shorter times to extinction than will be observed in the 'Alala.
Social factors might be very important. Some species of corvids nest colonially. Those species are highly prone to extinction, presumably because a small number of birds cannot survive: the Jackdaw seems to be an example. Were the 'Alala to be colonial or loosely colonial in its nesting habits, the predicted times to extinction would have to be revised sharply downward. (Little or nothing is known about the degree of coloniality of the 'Alala.)
Simply, the 'Alala population has a short time to extinction, because of its size and the inevitable demographic accidents that affect it and other tiny populations. Any additional unexpected environmental disturbance would shorten its time to extinction considerably.
Regardless of the particular causes of the decline of the 'Alala, its management, given its present critical status, should involve all ways to increase its rates of reproduction and survival at all stages of its life history.
The best-case scenario is that the small wild population on the McCandless Ranch is not declining and that three to five pairs of birds are breeding each year. Under such circumstances,
normal populations of corvid species—those not undergoing inexorable decline—often encounter local extinction on a scale of 1–2 decades. There is no evidence that a comparable global time to extinction is unrealistic for the 'Alala. Moreover, various factors—such as an inexorable decline because of disease, predation, or a dysfunctional social structure for the species—could all cause the expected times to extinction to be even shorter.
The demography of the population is unusual. Everywhere (including the McCandless Ranch), breeding success, fledgling survival, and yearling survival are not dramatically atypical of corvid species; nor, on the ranch, is adult survivorship. Off the McCandless Ranch, however, perhaps one-third of all the adults died each year, until the populations became extinct in those areas. That represents a catastrophic annual mortality for such a typically long-lived corvid species.
Estimates of times to extinction suggest that in the absence of any intervention, the McCandless Ranch population is likely to become extinct within a few decades. Of course, serious environmental disturbance or disease could extirpate a population even much larger than this one.
Population projections based on these admittedly limited demographic parameters suggest that, although the McCandless Ranch population appears stable and may even export a few dispersers, the population will remain at levels at which demographic accidents alone will continue to be a highly probable cause of extinction for the next several decades. For this reason, we believe it unlikely that the 'Alala can survive in the wild without intensive management.

































