
13
Endocrinological Responses to Dietary Salt Restriction During Heat Acclimation
Ralph P. Francesconi,1 Lawrence E. Armstrong, Natalie M. Leva, Robert J. Moore, Patricia C. Szlyk, William T. Matthew, William C. Curtis, Jr., Roger W. Hubbard, and Eldon W. Askew
INTRODUCTION
There are many, albeit sometimes inconsistent, reports related to the salt (NaCl) requirements of people working in hot environments. Dill (1938) recommended that 10 to 15 g per day of NaCl would be adequate to maintain electrolyte equilibrium despite the increased salt loss of men working in the heat. Later, Taylor et al. (1944) and Consolazio (1966) suggested an optimal intake of 13 to 17 g per day and 15 to 20 g per day, respectively. At about the same time, other investigators argued that, in heat-acclimatized men, salt deficits could be avoided by consuming only 5 g per day (Ladell, 1957) or 6 g per day of NaCl (Conn, 1949). Of course, the latter view is supported by the considerable populations that flourish in extremely hot global environments despite dietary NaCl intakes of from 2 to 6 g per day (Ladell, 1957; Orr and Gilks, 1931). A recent review from this laboratory (Hubbard et al., 1986) has pointed out that some of these early studies were uncontrolled with respect to heat acclimation, exercise level, and the period of dietary stabilization.
It has been well established that the acquisition of heat acclimation or natural acclimatization reduces significantly the amount of salt lost in the sweat (Allan and Wilson, 1971; Kirby and Convertino, 1986) and urine (Bonner et al., 1976; Costill et al., 1975; Finberg and Berlyne, 1977; Francesconi
et al., 1977). However, it is unknown whether these adaptive responses are sufficient to acquire and sustain acclimation during unaccustomed restriction of dietary salt intake. For example, there have been no comprehensive studies on the minimal daily consumption of salt necessary to acquire and sustain heat acclimation in individuals who ordinarily ingest 8 to 15 g per day of NaCl. This question is particularly important to military planners. When troops are rapidly deployed from garrison to field conditions, their normal salt intake in many cases drops precipitously due to the altered salt content of the issued field rations and the generally reduced total ration consumption, especially during the first several days of the deployment. If the mobilization destination is a desert or jungle environment, the problem is compounded by enhanced salt losses in sweat, especially during the early stages of heat acclimation.
It has been extensively documented, however, that reduced consumption of salt in the human diet results in hormonal adaptations designed to reduce urinary and sweat losses of sodium (Na+). Initially, reductions in Na+ content of the glomerular filtrate are perceived by cells of the distal tubules, thus stimulating the biosynthesis of renin (Rowell, 1986). Elevations in plasma renin activity (PRA) are followed rapidly by an increased biosynthesis and release of aldosterone (ALD), which promotes reabsorption of Na+ by the distal tubules, with obligatory retention and return of water to the extracellular space. Thus, PRA and ALD, in conjunction with argininevasopressin (antidiuretic hormone, AVP), are the humoral factors most instrumental in the regulation of fluids and electrolytes, especially under conditions of reduced availability of either.
The responses of these hormones have been extensively studied during passive heat exposure (Adlerkreutz et al., 1977; Kosunen et al., 1976), during exercise in the heat (Francesconi et al., 1983, 1985), and during periods when these stressors have been combined with restricted or supplemented sodium intake (Brandenberger et al., 1986; Davies et al., 1981). Armstrong et al. (1987) reported that during heat acclimation subjects who consumed a diet providing 5.7 g per day of NaCl had higher heart rates and rectal temperatures, as well as attenuated increments in plasma volume, than when the same group repeated the acclimation while ingesting 23 g per day of NaCl. Endocrinological responses were not described.
The current study offered a unique opportunity to assess and quantitate the endocrinological responses during, and perhaps integral to, the acquisition of heat acclimation and prolonged periods of work in a desert environment (Costill et al., 1976). Further, consumption of a low-salt diet during the acclimation period was expected to amplify these hormonal adaptations (Follenius et al., 1979) and provide important information on their role in the individual's response to recurrent and prolonged exercise in the heat. A
prior dietary stabilization period in which all test subjects consumed the same daily moderate level of Na+ provided for a relatively homogeneous range of initial levels of ALD, PRA, and AVP, thus stabilizing the usual lability of these variables in young adult men. Finally, the opportunity to evaluate hormonal responses across a 10-day acclimation period permitted the addition of confirmational data to earlier findings on the effects of acclimation on the responses of these hormones to exercise in the heat (Francesconi et al., 1983, 1984).
METHODS
Subjects
Seventeen young adult males (mean age 19.8 ± 0.6 years) participated in this study after providing their written voluntary consent. Average height was 179.4 ± 1.6 cm, average weight was 78.5 ± 2.4 kg, and body surface area was 1.97 ± 0.03 m2. Subjects were briefed orally on the procedures, risks, and benefits of the study, as well as on their right to withdraw at any time for any reason without penalty. Prior to their participation, all subjects were medically screened and examined and found to be in good health; the mean maximal oxygen consumption for these subjects was 46.5 ± 1.1 ml per minute per kg.
Design
The study was divided into two time intervals: (a) a 7-day dietary stabilization period under dormitory conditions and a temperature of 21°C and (b) a 10-day heat acclimation period, each day of which simulated an ''average'' 8-hour workday in a desert environment. During the complete 17-day interval, test volunteers were dedicated to the study and supervised 24 hours per day by test investigators who had total dietary control. During the 7-day dietary stabilization period, all volunteers consumed approximately 4000 kcal per day, which was adequate to sustain initial weights; the NaCl content of the diet was constant at 8 g per day for all subjects during this control interval. On day 8 of the study (day 1 of heat acclimation and work in the heat), subjects were randomly divided into two dietary groups, one of which continued to consume the moderate NaCl diet (8 g per day) and served as the control, while the second was placed on a low-salt diet (4 g per day) for the remainder of the test. Appropriate caloric consumption was sustained during the 10-day heat acclimation period for both groups as evidenced by minimal reductions in body weight over the experimental interval with no effects of diet on body weight.
Heat Acclimation and Work in the Heat
To ensure that both the duration of heat exposure and the amount of work done in the heat were adequate to elicit effects of the low-salt diet, subjects remained in the hot environment for approximately 8.5 hours per day. They entered a large environmental chamber (41°C dry bulb, 21 percent relative humidity, 1.1 to 1.2 meters per second wind speed) at approximately 7:30 a.m. on each of the 10 heat acclimation days and remained standing for at least 20 minutes to stabilize body fluid compartments prior to blood sampling (Hagan et al., 1978). At approximately 8:00 a.m. on each day of acclimation (days 8 to 17), subjects began exercise (treadmill, 5 percent grade, 5.6 km per hour) in this hot environment. They then walked for 30 minutes and rested for 30 minutes each hour for the next 8 hours with water available (temperature of water, 10° to 15°C). Fluid consumption and body weight were monitored at 30-minute intervals, and subjects were instructed to maintain euhydration levels by consuming the full complement (based on body weight measures) that was allowed during each 30-minute interval. Lunch was presented at approximately 12:30 p.m. (fifth rest period), and subjects were allowed to exit the chamber shortly after the eighth walk upon completing other physiological and behavioral measures (approximately 4:00 p.m.). If a person was unable to complete all of the work cycles on a particular day, he remained in the chamber and participated in all other aspects of the protocol (for example, diet, drinking, other tests). Thus, the continuity of the heat exposure component of the acclimation process was uninterrupted. After completing the daily chamber exposure, subjects returned to their dormitory setting until the regimen was repeated the next morning.
Blood Sampling
During the dietary stabilization period, on days 1, 4, and 7 a 6-ml sample of blood was removed by venipuncture from a superficial arm vein by a trained phlebotomist using aseptic techniques. This sample was removed at approximately 7:45 a.m. after subjects had been standing for 20 minutes; room temperature was 21° ± 1°C during the entire stabilization period. During the heat-work period (days 8 to 17) three samples were taken on each of experimental days 8, 11, 15, and 17, which corresponded to days 1, 4, 8, and 10 of heat acclimation. Because of the requirement for repeated blood sampling on these days, a catheter was aseptically inserted into each subject's superficial arm vein. The first blood sample of the day (T1) was taken after subjects had stood in the heat for 20 minutes, prior to exercise, at approximately 7:45 a.m. to correspond with the time of blood sampling during the dietary stabilization period. The second blood sample
(T2) was removed immediately after the fourth work bout (approximately 11:30 a.m.). The final blood sample (T3) was withdrawn after the final walk (approximately 3:30 p.m.). Collecting tubes were immediately placed into ice and transported to the laboratory for centrifugation (4°C, 10,000 rpm); EDTA plasma or serum was removed and stored (-20°C) for subsequent analysis.
Radioimmunoassays
Aldosterone was quantitated in serum using commercially available kits purchased from the Diagnostic Products Corporation (Los Angeles, California) and following techniques described in their technical bulletin (Aldosterone, No Extraction, Coat-A-CountR). This technique provides an approximate detection limit of 16 picograms (pg) per ml and is extremely specific for aldosterone; the range for this hormone is usually 4 to 31 nanograms (ng) per deciliter in salt-replete, standing adults (Aldosterone, No Extraction, Coat-A-CountR). Plasma renin activity was estimated by the quantitation of angiotensin I in EDTA plasma. Commercially available test kits (RIANEN Angiotensin I [125I] RIA Kit) were purchased from DuPont NEN Products (Boston, Massachusetts), and the assay was performed according to techniques outlined in their technical manual (RIANEN Assay System, Angiotensin I, Instruction Manual). When endogenous converting enzyme and angiotensinases of human plasma are appropriately inhibited, then angiotensin I formation quantitatively reflects PRA. Control values in adult men ordinarily range from 1 to 4.1 ng angiotensin I formed per ml per hour (Young, 1987).
Arginine-vasopressin was quantitated in EDTA plasma according to the techniques outlined by LaRose et al. (1985). One ml of EDTA plasma was treated with 10 µl per ml of 50 percent trifluoroacetic acid to acidify the sample to a pH of 4.0 to 4.5. Rabbit antibody to arg8-vasopressin was purchased from the Calbiochem Corporation (San Diego, California), and 125I-AVP was purchased from the DuPont NEN Corporation. Prepared standards were purchased from the Incstar Corporation (Stillwater, Minnesota). The range of circulating AVP in healthy adult males has been reported to be non-detectable to 4.7 pg per ml (Incstar, Vasopressin 125I RIA Kit).
Statistical Analysis
Repeated measures analysis of variance was performed using statistical package BMDP4V (BMDP Statistical Software, Los Angeles, California). Tukey's mean critical difference test was applied post hoc to determine significant differences of appropriate mean values. The null hypothesis was rejected at p < .05.
RESULTS
Due to the chronicity of the heat acclimation regimen and the need to complete eight work-rest cycles on a given day, all subjects were unable to complete the 80 treadmill walks (224 km). The main reasons for not completing all trials included foot blisters, inner thigh skin chafing, heat rash, and leg muscle pain. However, there was no significant difference in the proportion of the maximal possible walks completed between the two groups (control = 82.7 percent, low-salt = 75.4 percent). Further, as noted previously, if subjects could do only a portion of the total walks on a given day, they remained in the chamber and maintained the same rigorous nutritional, hydrational, and psychological testing requirements of the study as their walking counterparts. For these reasons the endocrinological data of subjects who did not walk the full complement of trials on all days were included in the mean values reported as well as the statistical analyses.
Figure 13-1 illustrates the effects of the low-salt diet and recurrent exercise in the heat on plasma levels of aldosterone. During the dietary stabilization period (days 1, 4, and 7) there were no significant differences (p > .05) noted between the control and low-salt groups on any of the days; in fact, the two groups displayed remarkably consistent between-group values in this sometimes labile variable. The slight elevation in levels of both groups on day 4 may have been in response to the dietary stabilization intake of NaCl (8 g), which probably represented a decrement in salt intake for most of these young adult men in comparison to their normal garrison consumption (approximately 11 to 15 g per day; Szeto et al., 1987). This increment nearly achieved statistical significance (for example, on day 1, low-salt, mean = 18.3 ng per dl; on day 4, low-salt, mean = 33.2 ng per dl, minimal critical difference of the means necessary for significance = 15.1 ng per dl, p > .05).
On the first day of both heat acclimation and dietary manipulation (eighth experimental day) plasma ALD levels at T1, T2, and T3 were not significantly different between groups (p > .05). However, the effects of exercise in the hot environment are noted in the elevated plasma ALD levels in both groups at T2, which achieved statistical significance by T3 in both the control and the low-salt groups (p < .05). By day 11, the marked effects of the low-salt diet on circulating ALD were manifested in significant (p < .01) increments at all sampling times when compared to the control levels. In the low-salt group, the increment observed between T1 and T2 narrowly failed to achieve significance (difference of means = 45.73, minimal difference for significance = 46.46 ng per dl).
A strikingly similar pattern emerged on day 15. Thus, at each of the sampling times, plasma ALD in the low-salt group was significantly elevated (p < .01) when compared to the respective mean of the control
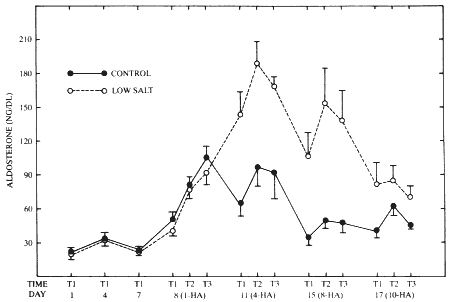
FIGURE 13-1 The effects of consuming a low-salt diet (4 g per day) or a moderate-salt diet (8 g per day) and of work in the heat (41°C; 21 percent relative humidity, treadmill, 5 percent grade, 5.6 km per hour, 30 minutes per hour, 8 hours per day) on plasma levels of aldosterone. During the dietary stabilization period (days 1 to 7), blood samples were taken at approximately 7:45 a.m. on days 1, 4, and 7; during the heat acclimation (HA) period (days 8 to 17), the first blood sample (T1) was removed after each subject remained upright in the heat chamber for at least 20 minutes, also at about 7:45 a.m. The second (T2) and third (T3) samples were removed after the fourth and eighth walks, respectively, on days 8, 11, 15, and 17. Mean values ± standard errors of the mean are depicted for all values.
group. Likewise, the apparent increment in circulating ALD in the low-salt group between T1 and T2 once again barely failed to achieve statistical significance (difference of means = 46.13, minimal difference for significance = 46.46 ng per dl). However, by day 17 (corresponding to day 10 of both dietary and heat-work manipulations), the effects of the low-salt diet were minimized, and there were no significant effects of either NaCl consumption or exercise in the heat on circulating levels of ALD. There were no between-diet effects on circulating Na+ concentrations, and these levels remained within the range of normal (135.6 to 140.8 mEq per liter) throughout the experimental period. As anticipated, urinary Na+ in the low-salt group fell precipitously (to less than 10 mEq per liter) during the 10 days of low-salt-exercise in the heat.
The responses of ALD to the low-salt-heat-work regimen were nearly mirrored in the effects of these parameters on PRA (Figure 13-2). In examining first the PRA of both groups during the dietary stabilization period, there were no significant differences in plasma levels between groups on day 1. In progressing from day 1 to day 4, a trend occurred toward elevations in PRA in both groups, but neither achieved statistical significance. However, in the low-salt group, the sharp decrement between day 4 and day 7 resulted in a significant difference (p < .05) in mean levels. On the first heat acclimation day, although there were no significant intergroup effects, exercise in the heat elicited a significant elevation (p < .01) in PRA in the low-salt group (T1 versus T3); other trends toward increased activity on day 8 failed to achieve statistical significance. On day 11 (fourth day of heat acclimation), there was a significant elevation (p < .05) of PRA in the low-salt group at T1. However, because of the slight decrements in activity in
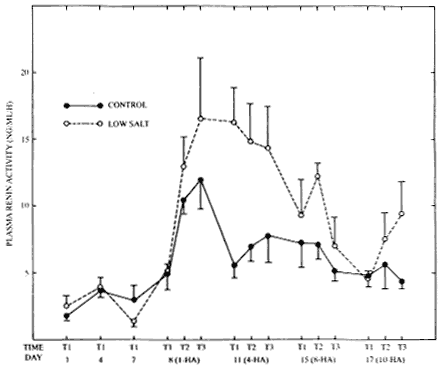
FIGURE 13-2 The effects of consuming a low-salt or moderate-salt diet and of recurrent exercise in the heat on levels of plasma renin activity. All conditions, times, and parameters are as noted in Figure 13-1. HA = days of heat acclimation period and T1, T2, and T3 are the first, second, and third blood samples, respectively.
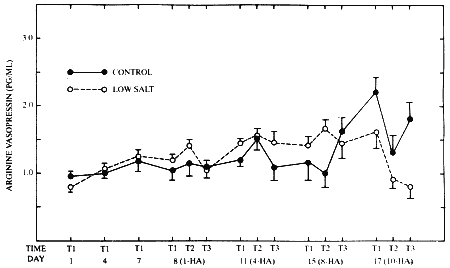
FIGURE 13-3 The effects of consuming a low-salt or moderate-salt diet and of recurrent exercise in the heat on circulating levels of arginine-vasopressin. All conditions are as noted in Figure 13-1. HA = days of heat acclimation period and T1, T2, and T3 are the first, second, and third blood samples, respectively.
the low-salt group and the concomitant minor increments in PRA in the controls, no further significant differences were noted in any of the other time intervals. Moreover, as the chronicity of heat acclimation increased (days 15 and 17), all effects of the low-salt diet as well as exercise in the heat were negated, and no further intergroup or between-time differences were observed.
During the dietary stabilization period, there were no significant intergroup or across-time differences in levels of AVP (Figure 13-3). Further, even after dietary manipulation and consecutive days of exercise in the heat, the data indicated that throughout the period of heat acclimation there were no significant effects on AVP of either the dietary manipulation or the recurrent exercise in the hot environment.
DISCUSSION
Because the endocrinological variables under consideration in the current experiments are significantly affected by dietary salt consumption (McDougall, 1987), hydration state (Convertino et al., 1981), exercise and training (Geyssant et al., 1981), and thermal exposure (Kosunen et al., 1976), it was considered important that all test subjects undergo an adequate stabi-
lization period prior to salt restriction and recurrent exercise in the heat. Thus, during the first 7 days of the experiment all subjects remained under the 24-hour control of test investigators with obligations only to the study (as was the case throughout the experiment). During this interval, subjects consumed the same test diets—which delivered 8 g of NaCl daily for each volunteer—and adhered to a structured and consistent schedule of activity that included completion of questionnaires (for example, environmental symptoms, thermal comfort), simulated target acquisition and firing, a maximal oxygen uptake test on day 4, and at least 4 hours per day of light recreational activity that included reading, videos, and games. All of this activity took place after blood sampling on all days.
During the stabilization period, circulating levels of all three hormones were generally consistent between groups, with some minor variation over time. All values were well within the normal range for young adult males, except for ALD levels on day 4, which were approximately 4 ng per dl above the upper limit of the normal range. This increment was probably due to the reduced NaCl content of the stabilization diet (8 g per day) as compared to the normal salt content of military dining hall rations, which is usually 11 to 15 g per day (Szeto et al., 1987). By day 7, mean ALD levels had returned to within the normal range.
Francesconi et al. (1983, 1985) and others (Finberg and Berlyne, 1977; Finberg et al., 1974) have reported—and current results confirm—that the acquisition of acclimation attenuates the response to exercise in the heat of both PRA and ALD. In the current experiments, the effects of acclimation were manifested in mean daily ALD levels of 79 and 84.9 ng per dl on days 8 and 11 (days 1 and 4 of heat acclimation), respectively, which were reduced by days 15 and 17 (days 8 and 10 of heat acclimation) to 44.3 and 51.3 ng per dl, respectively, in the control group. Even in the low-salt group, the mean plasma ALD concentration on day 11 was a markedly elevated 167.3 ng per dl, which was reduced to 133.5 and 78.7 ng per dl on days 15 and 17, respectively. By day 17 there were no significant differences in plasma ALD either between the control and low-salt groups at any of the individual sampling times or between the T1, T2, and T3 values for either group. These results suggest that the chronicity of both the heat acclimation regimen and the low-salt diet engender temporally related endocrinological accommodations to these experimental manipulations. Thus, the attenuated hormonal responses over time may be related to the following: increased plasma volume of heat acclimation (Bonner et al., 1976), generally reduced adrenocorticotrophic activity with decreased physiological strain (that is, a reduced stress response to the experimental conditions) (Francesconi et al., 1984), decreased sensitivity to adrenocorticotrophic hormone (McDougall, 1987), increased renal sensitivity to ALD-induced sodium reabsorption mechanisms (Smiles and Robinson, 1971), and a general improvement of the Na+
balance due to improved Na+ conservation at the level of the kidney and sweat gland (Allan and Wilson, 1971).
Similar arguments and rationale can be made for the pattern of PRA levels observed in the current experiments. For example, in the control group, mean PRA peaked on day I of heat acclimation (9.04 ng per ml per hour) followed by a declining trend (6.8, 6.6, and 5.3 ng per ml per hour) on days 4, 8, and 10, respectively, of heat acclimation. Moreover, in the low-salt group, by using the rationale developed above for ALD, maximal effects of the low-salt diet and exercise in the heat would be expected on day 4 of heat acclimation (15.2 ng per ml per hour) followed by moderation on day 8 (9.6 ng per ml per hour) with further reductions on day 10 (7.3 ng per ml per hour). The endocrinological adaptations that occur early in the acclimation process are necessary to maintain electrolyte balance and ultimately to expand extracellular fluid volume and may be closely related to the acquisition of full acclimation. Moreover, when the physiological strain of exercise in the heat has been reduced after full acclimation and expansion of plasma volume have been achieved, these hormonal responses are obviated.
In the current experiments, subjects on the low-salt diet gradually achieved the same state of acclimation as the control group over the 10-day acclimation period as suggested by significant reductions in heat-and exercise-induced heart rate, rectal temperature, urinary Na+, and perceived exertion. Further, neither these variables nor the distance walked during the 10-day acclimation period differed significantly between the control and low-salt groups. The moderation of the endocrinological responses during the latter phase of the acclimation interval probably reflects, most importantly, the attendant plasma volume expansion and achievement of electrolyte balance. However, successful acclimation to recurrent and prolonged exercise in the heat on 4 g per day of NaCl was accomplished under the specific conditions of this study. It is unknown whether successful adaptation to this low-salt diet could have been accomplished if the exercise or environmental conditions were more intense.
It is generally agreed that AVP is most responsive to dehydration and increments in plasma osmolality (Von Ameln et al., 1985). Accordingly, Greenleaf et al. (1983) reported that when water intake was increased during an acclimation period from 450 ml per hour on day 1 to approximately 1000 ml per hour on days 5 to 8, there also occurred a significant decrement in plasma AVP. In the current experiments, subjects were weighed every 30 minutes during each of the 8-hour workdays, were encouraged to drink, and were provided fresh, cool water at 30-minute intervals. Thus, dehydration was not a significant observation in either group of subjects, and AVP levels remained generally consistent throughout both the stabilization and the heat acclimation intervals. In fact, the range of AVP for both groups over all
sampling periods (0.80 to 2.21 pg per ml, mean values) is within the range of normal for young adult men with plasma osmolality less than 290 milliosmoles per kg.
CONCLUSIONS AND RECOMMENDATIONS
As shown from these studies, young adult men consuming a daily diet containing only 4 g of NaCl can work consecutive days in a desert environment and achieve full acclimation to this specific heat-work scenario. The endocrinological adaptations occurring especially during the first several days of the dietary-heat-work regimen may be important to the physiological adaptations and electrolyte equilibria needed for achieving acclimation. These hormonal responses moderated during the latter portions of the experimental regimen, which indicates a dietary and acclimation steady-state characterized by greatly reduced physiological strain to the test volunteers. These results imply that healthy young individuals can acclimate quite rapidly to work in desert environments despite relatively restricted salt intake.
ACKNOWLEDGMENTS
The authors gratefully acknowledge the technical, technological, and logistical support of many U.S. Army Research Institute of Environmental Medicine and U.S. Army Natick Research, Development, and Engineering Center personnel. A special thanks to the test volunteers who participated in this study. Without their cooperation, it could not have been executed.
REFERENCES
Adlerkreutz, H., K. Kosunen, K. Kuoppasalmi, A. Pakarinen, and S. Karonen 1977 Plasma hormones during exposure to intense heat. Proc. Cong. Int. Med. 13:346–355.
Allan, J.R., and C.G. Wilson 1971 Influence of acclimatization on sweat sodium concentration. J. Appl. Physiol. 30:708–712.
Armstrong, L.E., D.L. Costill, and W.J. Fink 1987 Changes in body water and electrolytes during heat acclimation: Effects of dietary sodium. Aviat. Space Environ. Med. 58:143–148.
Bonner, R.M., M.H. Harrison, C.J. Hall, and R.J. Edwards 1976 Effect of heat acclimatization on intravascular responses to acute heat stress in man. J. Appl. Physiol. 41:708–713.
Brandenberger, G., V. Candas, M. Follenius, J. Libert, and J. Kahn 1986 Vascular fluid shifts and endocrine responses to exercise in the heat: Effects of rehydration. Eur. J. Appl. Physiol. 55:123–129.
Conn, J.W. 1949 Acclimatization to heat. Adv. Int. Med. 3:337.
Consolazio, D.F. 1966 Nutrient requirements of troops in extreme environments. Army Res. Dev. Mag. Nov:24–27.
Convertino, V.A., L.C. Keil, E.M. Bernauer, and J.E. Greenleaf 1981 Plasma volume, osmolality, vasopressin, and renin activity during graded exercise in man. J. Appl. Physiol. 50:123–128.
Costill, D.L., R. Cote, E. Miller, T. Miller, and S. Wynder 1975 Water and electrolyte replacement during repeated days of work in the heat. Aviat. Space Environ. Med. 46:795–800.
Costill, D.L., G. Branam, W. Fink, and R. Nelson 1976 Exercise induced sodium conservation: Changes in plasma renin and aldosterone. Med. Sci. Sports 8:209–213.
Davies, J., M. Harrison, L. Cochrane, R. Edwards, and T. Gibson 1981 Effect of saline loading during heat acclimatization on adrenocortical hormones. J. Appl. Physiol. 50:605–612.
Dill, D.B. 1938 Life, Heat, and Altitude. Cambridge, Mass.: Harvard University Press.
Finberg, J.P.M., and G.M. Berlyne 1977 Modification of renin and aldosterone response to heat by acclimatization in man. J. Appl. Physiol. 42; 554–558.
Finberg, J.P.M., M. Katz, H. Gazit, and G.M. Berlyne 1974 Plasma renin activity after acute heat exposure in nonacclimatized and naturally acclimatized man. J. Appl. Physiol. 36:519–523.
Follenius, M., G. Brandenberger, B. Reinhardt, and M. Simeoni 1979 Plasma aldosterone, renin activity, and cortisol responses to heat exposure in sodium depleted and repleted subjects. Eur. J. Appl. Physiol. 41:41–50.
Francesconi, R., J.T. Maher, G. Bynum, and J. Mason 1977 Recurrent heat exposure: Effects on plasma and urinary sodium and potassium in resting and exercising men. Aviat. Space Environ. Med. 48:399–404.
Francesconi, R.P., M.N. Sawka, and K.B. Pandolf 1983 Hypohydration and heat acclimation: Plasma renin and aldosterone during exercise. J. Appl. Physiol. 55:1790–1794.
1984 Hypohydration and acclimation: Effects on hormone responses to exercise/heat stress. Aviat. Space Environ. Med. 55:365–369.
Francesconi, R.P., M. Sawka, K. Pandolf, R. Hubbard, A. Young, and S. Muza 1985 Plasma hormonal responses at graded hypohydration levels during exercise/heat stress. J. Appl. Physiol. 59:1855–1860.
Geyssant, A., G. Geelen, C. Denis, A.M. Allevard, M. Vincent, E. Jarsaillon, C. A. Bizollon, J.R. Lacour, and C. Gharib 1981 Plasma vasopressin, renin activity, and aldosterone: Effect of exercise and training. Eur. J. Appl. Physiol. 46:21–30.
Greenleaf, J.E., P.J. Brock, L.C. Keil, and J.T. Morse 1983 Drinking and water balance during exercise and heat acclimation. J. Appl. Physiol. 54:414–419.
Hagan, R.D., F.J. Diaz, and S.M. Horvath 1978 Plasma volume changes with movement to supine and standing positions. J. Appl. Physiol. 45:414—418.
Hubbard, R.W., L.E. Armstrong, P.K. Evans, and J.P. DeLuca 1986 Long-term water and salt deficits: A military perspective. Pp. 29–48 in Predicting Deficits in Military Performance Due to Inadequate Nutrition. Washington, D.C.: National Academy Press.
Kirby, C.R., and V.A. Convertino 1986 Plasma aldosterone and sweat sodium concentrations after exercise and heat acclimation. J. Appl. Physiol. 61:967–970.
Kosunen, K. J., A.J. Pakarinen, K. Kuoppasalmi, and H. Adlerkreutz 1976 Plasma renin activity, angiotensin II, and aldosterone during intense heat stress. J. Appl. Physiol. 41:323–327.
Ladell, W.S.S. 1957 Disorders due to heat. Trans. R. Soc. Trop. Med. Hyg. 51:189–216.
LaRose, P., H. Ong, and P. DuSouich 1985 Simple and rapid radioimmunoassay for the routine determination of vasopressin in plasma. Clin. Biochem. 18:357–361.
McDougall, J.G. 1987 The physiology of aldosterone secretion. News Physiol. Sci. 2:126–128.
Orr, J.B., and J.L. Gilks 1931 Studies of nutrition: The physique and health of two African tribes. Medical Research Council London, Special Reports, Series 155:82, His Majesty's Stationery Office, London.
Rowell, L.B. 1986 Human Circulation: Regulation During Physical Stress. New York: Oxford University Press.
Smiles, K.A., and S. Robinson 1971 Sodium ion conservation during acclimatization of men to work in the heat. J. Appl. Physiol. 31:63–69.
Szeto, E.G., D.E. Carlson, T.B. Dugan, and J.C. Buchbinder 1987 A comparison of nutrient intakes between a Ft. Riley contractor-operated and a Ft. Lewis military-operated garrison dining facility. U.S. Army Research Institute of Environmental Medicine Tech Rep. T2-88:1–65, Natick, Mass.: U.S. Army Research Institute of Environmental Medicine.
Taylor, H.L., A. Henschel, O. Mickelson, and A. Keys 1944 The effect of sodium chloride intake on the work performance of man during exposure to dry heat and experimental heat exhaustion. Am. J. Physiol. 140:439–451.
Von Ameln, H., M. Laniado, L. Rocker, and K.A. Kirsch 1985 Effects of dehydration on the vasopressin response to immersion. J. Appl. Physiol. 58:114–120.
Young, D.S. 1987 Implementation of SI units for clinical laboratory data. Ann. Int. Med. 106:114–129.
DISCUSSION
PARTICIPANT: What was driving the aldosterone and plasma renin activity responses? What was the relationship between those hormones and plasma volume, plasma osmolality, plasma sodium concentrations?
DR. FRANCESCONI: Well, the circulating sodium concentrations were actually not too different between groups, as Dr. Armstrong indicated. In addition to circulating sodium levels, circulating potassium levels, circulating protein levels, and osmolality were also not really significantly different.
Aldosterone biosynthesis is ordinarily stimulated by reduced dietary sodium, heat exposure, exercise, and usually by increases in plasma renin activity although we have seen conditions where the two have been separated.
So I can only conclude that increased sodium and fluid reabsorption in the face of reduced dietary salt intake was the driving factor for those endocrinological responses.
Increased plasma renin activity was probably due to diminished splanchnic blood flow, as Rowell (Rowell, 1986) has shown; decreased plasma volume or reduced renal blood flow ordinarily elicits a very immediate response in terms of renal tubular biosynthesis and release of plasma renin. These endocrinological responses thus stimulate very efficient electrolytes and, thus, fluids.
PARTICIPANT: I was going to ask somewhat of a similar question. I was trying to figure out the mechanisms, first, when you kept everything constant, the aldosterone and renin went up. I was fascinated that they went back down.
DR. FRANCESCONI: They did and that could be a result of just the achievement of new steady states after the full acquisition of acclimation and expansion of plasma volume. It could he an increased sensitivity of both the sweat glands and the kidney cells to the activity of the PRA and the aldosterone.
Nothing that we saw would indicate that any of the subjects were becoming either hyponatremic or hypokalemic in these studies.
PARTICIPANT: As acclimation progressed, the intensity of these endocrine responses was moderated. Is that just due to increased sensitivity to PRA and ALDO.
DR. FRANCESCONI: That is just one of the mechanisms proposed.
PARTICIPANT: But, the mechanisms remain very vague.
DR. FRANCESCONI: As an aside, I have had rats on low sodium diets for up to 70 or 80 days with no real significant effects on circulating sodium levels and almost nonexistent urinary or salivary sodium.
PARTICIPANT: I am not so sure that is a change in sensitivity because in one of those slides that you showed where you looked at preacclimatization and postacclimatization, if you looked at the two lines there was a difference between the two, but those two lines looked like they were parallel. They appeared to be rising at the same rate for PRA and ALDO.
It looked as if the sensitivity was the same but just the initial values were different.
DR. FRANCESCONI: Actually, on day 1 of heat acclimation the responses
were quite similar, but, of course, on that day the volunteers in both groups were coming off the same stabilization diet and level of salt intake. By day 4 of heat acclimation, the T1 values for PRA and ALD were remarkably different due to the low-salt diet.
And clearly, if acclimation increases plasma volume, which it has been reported to do up to 18 percent, there should indeed be a reduction in both the T1 levels and the responses as acclimation progresses.
In fact, at one time we infused hyperoncotic albumin to test volunteers to increase plasma volume in a matter of hours versus the six or eight days that Dr. Armstrong showed were required here in this study.
And in that condition also we had repression of baseline levels of those hormones, not great, but statistically significant.
PARTICIPANT: Did you measure blood pressure during that period of time? Because that could change glomerular filtration rate and explain some of these differences.
DR. FRANCESCONI: There were actually some decreases in blood pressure during heat acclimation. In fact, I think Lt. Colonel William Curtis has much data on that which he is about to present at a different meeting. There were some decrements in blood pressure.
PARTICIPANT: I am trying to remember, from the military initiatives study, what was the sodium intake of soldiers in garrison? What were measured in some of those studies?
PARTICIPANT: At least 4500 mg of sodium per day.
PARTICIPANT: So that would be roughly 9 grams of salt, or 11 or 12. So maybe a little more.
Would you speculate that if you were to take soldiers coming right off the food that they would be consuming normally when they are on base and were dropped into Saudi Arabia and eating only MREs getting 4 grams of salt, that you would see the same kinds of results that you saw there?
DR. FRANCESCONI: I think I probably would. If they had responsive and well-functioning endocrinological systems, young, healthy adult males and if they were drinking well, yes.
DR. NESHEIM: Any other questions?
PARTICIPANT: Just point of clarification. Our current MRE is much higher in salt than the 4 grams.
DR. FRANCESCONI: Yes. That is true.
PARTICIPANT: Do you expect any differences in that level for women?
DR. FRANCESCONI: No, I don't. The hypohydration study which I have referenced actually had 12 women in that group of 24 test subjects and I saw no differences in hormonal responses and I don't think Dr. Sawka saw any differences in physiological responses in that study.
PARTICIPANT: My question is, do you anticipate that—this is a very small population you are looking at. There are some changes that are occurring here. Do you expect that there might be some people in a group of young men like this that might not respond the same way.?
DR. FRANCESCONI: Clearly, we can only speak for the volunteers and the conditions in this study, at least at this time. However, the consistency of the responses indicated that such responses should generally occur, especially considering the numbers of papers in the literature describing the effects of heat and exercise on circulating levels of these hormones.
PARTICIPANT: It could represent a little caution in interpreting that everybody would respond that way.
PARTICIPANT: That is what I am worried about. If you look at the number of people who have heat stroke, it is a small number but they do occur.
PARTICIPANT: I just wanted to ask—Larry [ed. Dr Armstrong] mentioned that some people couldn't make it through this rigorous heat/exercise protocol for orthopedic reasons. How bad was the drop-out rate and maybe these people don't have the endocrinological response.
DR. FRANCESCONI: I didn't see any correlation. As Dr. Armstrong mentioned, the drop-outs were primarily for reasons that you would expect on this kind of a march—foot blisters primarily, ankle pain, intra-thigh chafing, especially for subjects that were a little heavier, all those kinds of things. In some instances the volunteers would miss one or two of the 30-minute marches and then rejoin the group. In other instances they may have missed a day that was not a blood-sampling day. However, they did remain in the heat all day on those days.
DR. JOHNSON: One note that might address your question: Even though people may have stopped walking on the treadmill, they stayed in the wind tunnel. They didn't leave the hot environment. They were still there.


















