
3
Physiological Responses to Exercise in the Heat
Michael N. Sawka,1 C. Bruce Wenger, Andrew J. Young, and Kent B. Pandolf
INTRODUCTION
Humans often exercise strenuously in hot environments for reasons of recreation, vocation, and survival. The magnitude of physiological strain imposed by exercise-environmental stress depends on the individual's metabolic rate and capacity for heat exchange with the environment. Muscular exercise increases metabolism by 5 to 15 times the resting rate to provide energy for skeletal muscle contraction. Depending on the type of exercise, 70 to 100 percent of the metabolism is released as heat and needs to be dissipated in order to maintain body heat balance. The effectiveness of the thermoregulatory system in defending body temperature is influenced by the individual's acclimatization state (Wenger, 1988), aerobic fitness (Armstrong and Pandolf, 1988), and hydration level (Sawka and Pandolf, 1990). Aerobically fit persons who are heat acclimatized and fully hydrated have less body heat storage and perform optimally during exercise-heat stress. To regulate body temperature, heat gain and loss are controlled by the autonomic nervous system's alteration of (a) heat flow from the core to the skin via the blood and (b) sweating. Thermoreceptors in the skin and body core provide input into the hypothalamic thermoregulatory center where this information is processed, via a proportional control system, with a resultant
signal for heat loss by the thermoregulatory effector responses of sweating and alterations in skin blood flow (Sawka and Wenger, 1988).
This chapter reviews human temperature regulation and normal physiological responses to exercise-heat stress. In general, muscular exercise and heat stress interact synergistically and may push physiological systems to their limits in simultaneously supporting the competing metabolic and thermoregulatory demands.
CORE TEMPERATURE RESPONSES TO EXERCISE
During muscular exercise, core temperature initially increases rapidly and subsequently increases at a reduced rate until heat loss equals heat production, and essentially steady-state values are achieved. At the initiation of exercise, the metabolic rate increases immediately; however, the thermoregulatory effector responses for heat dissipation respond more slowly. The thermoregulatory effector responses, which enable sensible (radiative and convective) and insensible (evaporative) heat loss to occur, increase in proportion to the rise in core temperature. Eventually, these heat loss mechanisms increase sufficiently to balance metabolic heat production, allowing achievement of a steady-state core temperature.
During muscular exercise, the magnitude of core temperature elevation is largely independent of the environmental condition and is proportional to the metabolic rate (Gonzalez et al., 1978; Nielsen, 1938, 1970). This concept was first presented by Nielsen (1938) who had three subjects perform exercise at several intensities (up to approximately 3.0 liters oxygen per minute) in a broad temperature range (5° to 36°C with low humidity). Figure 3-1 presents the heat exchange data for one subject during an hour of cycle exercise at a power output of 147 watts and at a metabolic rate of approximately 650 watts. The difference between metabolic rate and total heat loss represents the energy used for mechanical work and heat storage. The total heat loss and, therefore, the heat storage and elevation of core temperature were constant for each environment. The relative contributions of sensible and insensible heat exchange to total heat loss, however, varied with environmental conditions. In the 10°C environment, the large skin-to-ambient temperature gradient facilitated sensible heat exchange, which accounted for about 70 percent of the total heat loss. As ambient temperature increased, this gradient for sensible heat exchange diminished, and there was a greater reliance upon insensible heat exchange. When the ambient temperature was equal to skin temperature, insensible heat exchange accounted for almost all the heat loss. In addition, when the ambient temperature exceeded the skin temperature, there was a sensible heat gain to the body.
Nielsen's finding that the magnitude of core temperature elevation is
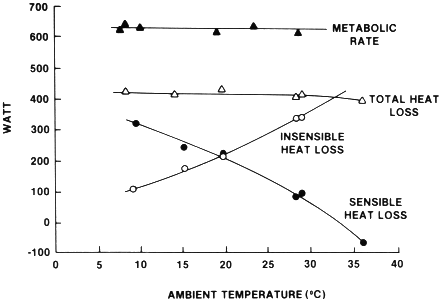
FIGURE 3-1 Heat exchange data averaged over 1 hour for one subject performing constant intensity exercise in a variety of ambient temperatures. The difference between metabolic rate and total heat loss is the sum of mechanical power (147 watts) and mean rate of heat storage. SOURCE: Sawka and Wenget (1988), used with permission. Redrawn from Nielsen (1938).
independent of environmental conditions is inconsistent with the personal experience of most athletes. For example, a runner will experience greater hyperthermia if he or she competes in a 35°C environment (Robinson, 1963). Lind (1963) showed that the magnitude of core temperature elevation during exercise is independent of the environment only within a certain range of conditions or a ''prescriptive zone.'' Figure 3-2 presents a subject's steady-state core temperature responses during exercise performed at three metabolic intensities in a broad range of environmental conditions. The environmental conditions are represented by the "old" effective temperature, which is an index that combines the effects of dry-bulb temperature, humidity, and air motion. Note that during exercise the greater the metabolic rate, the lower the upper limit of the prescriptive zone. In addition, Lind found that even within the prescriptive zone there was a small but significant positive relationship between the steady-state core temperature and the "old" effective temperature. It seems fair to conclude that throughout a wide range of environmental conditions, the magnitude of core temperature elevation during exercise is largely, but not entirely, independent of
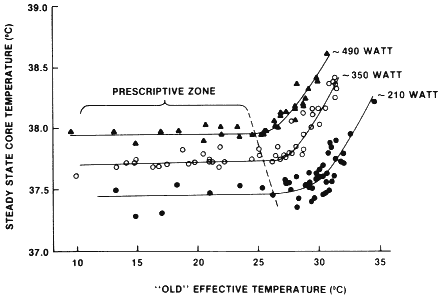
FIGURE 3–2
Relationship of steady-state core temperature responses during exercise at three metabolic rates to the environmental conditions. SOURCE: Sawka and Wenger (1988), used with permission. Redrawn from Lind (1963).
the environment. During exercise with a substantial metabolic requirement, the prescriptive zone might be exceeded, and there is a further elevation of steady-state core temperature.
As stated, within the prescriptive zone, the magnitude of core temperature elevation during exercise is proportional to the metabolic rate (Nielsen, 1938; Saltin and Hermansen, 1966; Stolwijk et al., 1968). Although the relationship between metabolic rate and core temperature is strong for a given individual, it does not always hold well for comparisons between different individuals. Åstrand (1960) first reported that the use of relative intensity (percentage of maximal oxygen uptake), rather than actual metabolic rate (absolute intensity), removes most of the intersubject variability for the core temperature elevation during exercise.
METABOLISM
Metabolic Rate
The effects of acute heat stress on a person's ability to achieve maximal aerobic metabolic rates during exercise have been thoroughly studied. Most
investigators find that maximal oxygen uptake is reduced in hot compared to temperate environments (Klausen et al., 1967; Rowell et al., 1969; Saltin et al., 1972; Sen Gupta et al., 1977), but some investigators report no differences (Rowell et al., 1965; Williams et al., 1962). For example, in one study (Sawka et al., 1985) maximal oxygen uptake ![]() was 0.25 liter per minute lower in a 49°C, as compared to a 20°C, environment (see Figure 3–3). Clearly, heat stress reduces
was 0.25 liter per minute lower in a 49°C, as compared to a 20°C, environment (see Figure 3–3). Clearly, heat stress reduces ![]() relative to that achieved in a temperate environment. In addition, the state of heat acclimatization did not alter the approximate 0.25 liter per minute decrement in
relative to that achieved in a temperate environment. In addition, the state of heat acclimatization did not alter the approximate 0.25 liter per minute decrement in ![]() . The question remains, What physiological mechanism(s) is/are responsible for this reduction in
. The question remains, What physiological mechanism(s) is/are responsible for this reduction in ![]() ? It can be theorized that thermal stress might result in a displacement of blood to the cutaneous vasculature, which could (a) reduce the portion of cardiac output perfusing the contracting musculature or (b) result in a decreased effective central blood volume and thus reduce venous return and cardiac output. As skin blood flow can reach 7 liters per minute
? It can be theorized that thermal stress might result in a displacement of blood to the cutaneous vasculature, which could (a) reduce the portion of cardiac output perfusing the contracting musculature or (b) result in a decreased effective central blood volume and thus reduce venous return and cardiac output. As skin blood flow can reach 7 liters per minute
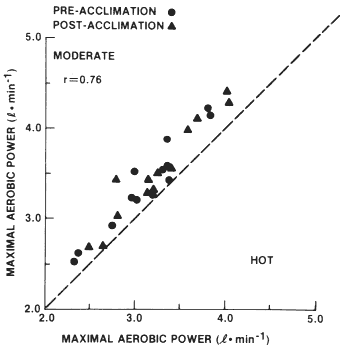
FIGURE 3-3 Maximal aerobic power values (liters per minute) for the pre-and postheat acclimatization tests in a moderate (21°C, 30 percent relative humidity) and a hot (49°C, 20 percent relative humidity) environment, r = Pearson product-moment correlation coefficient. SOURCE: Sawka et al. (1985), used with permission.
TABLE 3–1 Papers Reporting the Effect of Heat on Metabolic Rate During Exercise
|
Study |
Exercise Mode |
Metabolic Intensity |
Test Environment |
Change in Metabolic Rate Rate |
Comment |
|
Brouha et al., 1960 |
Cycle |
460 watts |
25 vs 32°C |
NC* |
— |
|
|
|
|
25 vs 37°C |
|
|
|
Consolazio et al., 1961 |
Treadmill |
400 watts |
26 vs 40°C |
|
Acclimated |
|
|
|
|
26 vs 41°C |
|
|
|
|
Cycle |
230 watts |
26 vs 40°C |
|
|
|
|
|
|
26 vs 41°C |
|
|
|
Consolazio et al., 1963 |
Cycle |
205 watts |
21 vs 29°C |
NC |
— |
|
|
|
545 watts |
|
NC |
— |
|
|
|
205 watts |
21 vs 38°C |
|
— |
|
|
|
545 watts |
|
|
— |
|
Dimri et al., 1980 |
Cycle |
10–90% |
27 vs 37°C |
|
Acclimated |
|
|
|
|
27 vs 40°C |
|
|
|
Fink et al., 1975 |
Cycle |
70–85% |
9 vs 41°C |
|
Unacclimated |
|
|
|
|
|
|
|
|
Peterson and Vejby-Christensen, 1973 |
Cycle |
87–347 watts |
24 vs 32°C |
|
Unacclimated |
|
|
|
|
|
|
|
|
Williams et al., 1962 |
Cycle |
— |
20 vs 36°C |
|
Acclimated |
|
|
|
|
|
|
|
|
Young et al., 1985 |
Cycle |
793 watts |
21 vs 49°C |
|
Unacclimated and Acclimated |
|
|
|
|
|
|
|
|
* NC = No change. † LA = Plasma lactate. |
|||||
(Rowell, 1986) during maximal vasodilation, the contracting musculature could receive less perfusion at a given cardiac output level. Rowell et al. (1966) reported that during high-intensity exercise in the heat, cardiac output can be reduced by 1.2 liters per minute below control levels. A reduction in maximal cardiac output by 1.2 liters per minute could account for a 0.25-liter-per-minute decrement in ![]() with heat exposure, because each liter of blood could deliver about 0.2 liter of oxygen (1.34 ml oxygen per g hemoglobin × l5 g hemoglobin per 100 ml of blood).
with heat exposure, because each liter of blood could deliver about 0.2 liter of oxygen (1.34 ml oxygen per g hemoglobin × l5 g hemoglobin per 100 ml of blood).
Acute heat stress increases resting metabolic rate (Consolazio et al., 1961, 1963; Dimri et al., 1980), but the effect of heat stress on an individual's metabolic rate for performing a given submaximal exercise task is not so clear (see Table 3-1). Such an effect would influence the calculation of the heat balance and might have implications for the nutritional requirements of individuals exposed to hot environments. Many investigators report that to perform a given submaximal exercise task, the metabolic rate is greater in a hot than temperate environment (Consolazio et al., 1961, 1963; Dimri et al., 1980; Fink et al., 1975). Some investigators, however, report lower metabolic rates in the heat (Brouha et al., 1960; Petersen and Vejby-Christensen, 1973; Williams et al., 1962; Young et al., 1985). Heat acclimation state does not account for whether individuals demonstrate an increased or decreased metabolic rate during submaximal exercise in the heat. However, other mechanisms can explain this discrepancy. Most investigators have only calculated the aerobic metabolic rate during submaximal exercise, ignoring the contribution of anaerobic metabolism to total metabolic rate.
Dimri et al. (1980) had six subjects exercise at three intensities in each of three environments. Figure 3-4 presents their subjects' total metabolic rate (bottom) and the percentage of this metabolic rate that was contributed by aerobic and anaerobic metabolic pathways. The anaerobic metabolism was calculated by measuring the postexercise oxygen uptake that was in excess of resting baseline levels. Although there are limitations to this methodology, the study provides useful information. Note that to perform exercise at a given power output, the total metabolic rate increased with the elevated ambient temperature. More importantly, the percentage of the total metabolic rate contributed by anaerobic metabolism also increased with the ambient temperature. The increase in anaerobic metabolic rate exceeded the increase of total metabolic rate during exercise at the elevated ambient temperatures. Therefore, if only the aerobic metabolic rate had been quantified, Dimri et al. (1980) would probably have reported a decreased metabolic rate in the heat for performing exercise at a given power output. Investigations that report a lower metabolic rate during exercise in the heat also report increased plasma or muscle lactate levels (Petersen and Vejby-Christensen, 1973; Williams et al., 1962; Young et al., 1985) or an increased respiratory exchange ratio (Brouha et al., 1960), which also suggests an
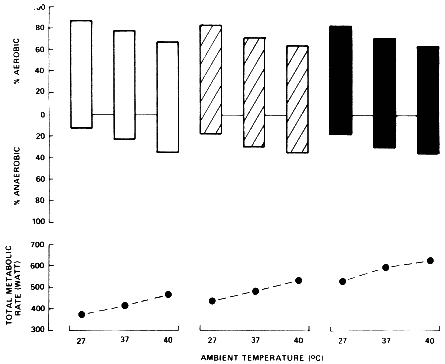
FIGURE 3-4 The total metabolic rate and percentage contribution of aerobic and anaerobic metabolism during exercise at different ambient temperatures. SOURCE: Sawka and Wenger (1988), used with permission. Data from Dimri (1980).
increased anaerobic metabolism. Likewise, other investigators report that plasma lactate levels are greater during submaximal exercise in a hot as compared to a comfortable environment (Dill et al., 1930/1931; Dimri et al., 1980; Fink et al., 1975; Nadel 1983; Robinson et al., 1941).
Interestingly, the oxygen uptake response to submaximal exercise does appear to be affected by heat acclimatization (Sawka et al., 1983). Most reports indicate that oxygen uptake and aerobic metabolic rate during submaximal exercise are reduced by heat acclimatization, although a significant effect is not always observed (see Table 3-2). Large effects (14 to 17 percent reductions) have been reported for stair-stepping (Senay and Kok, 1977; Shvartz et al., 1977; Strydom et al., 1966), but some of the reduction in ![]() during stair-stepping can be attributed to increased skill and improved efficiency acquired during the acclimatization program (Sawka et al., 1983). In other studies, although the acclimatization-induced reductions were statistically
during stair-stepping can be attributed to increased skill and improved efficiency acquired during the acclimatization program (Sawka et al., 1983). In other studies, although the acclimatization-induced reductions were statistically
TABLE 3-2 Papers Reporting the Effect Heat Acclimatization Has on Metabolic Rate During Exercise
|
Study |
n |
Acclimatization (days) |
Exercise Mode |
Test Environment |
Change in Metabolic Rate (ΔM) |
%Δ |
|
Cleland et al., 1969 |
3 |
7 |
Treadmill |
Hot |
NC* |
— |
|
Eichna et al., 1950 |
3 |
10 |
Treadmill |
Hot |
|
4 |
|
|
|
|
|
Cool |
|
2 |
|
Gisolfi, 1973 |
6 |
8 |
Treadmill |
Hot |
|
12 |
|
Jooste and Strydom, 1979 |
4 |
7 |
Treadmill |
Hot |
|
? |
|
|
|
|
|
Cool |
|
? |
|
Piwonka and Robinson, 1967 |
3 |
4 |
Treadmill |
Hot |
NC |
— |
|
Robinson et al., 1941 |
5 |
10–23 |
Treadmill |
Hot |
|
8 |
|
Rowell et al., 1967 |
6 |
11–12 |
Treadmill |
Hot |
NC |
— |
|
Sawka et al., 1983 |
42 |
6–10 |
Treadmill |
Hot |
|
3 |
|
|
|
|
|
Cool |
|
5 |
|
Senay and Kok, 1977 |
5 |
8 |
Step |
Hot |
|
14 |
|
|
|
|
|
Cool |
|
7 |
|
Shvartz et al., 1977 |
7 |
8 |
Step |
Hot |
|
16 |
|
|
|
|
|
Cool |
|
14 |
|
|
7 |
8 |
Step |
Hot |
|
14 |
|
|
|
|
|
Cool |
|
14 |
|
|
7 |
8 |
Step |
Hot |
|
10 |
|
|
|
|
|
Cool |
|
9 |
|
Strydom et al., 1966 |
5 |
12 |
Step |
Hot |
|
17 |
|
Weinman et al., 1967 |
5 |
8 |
Treadmill |
Hot |
NC |
— |
|
|
5 |
8 |
Treadmill |
Hot |
NC |
— |
|
Wyndham et al., 1976 |
4 |
10 |
Cycle |
Hot |
NC |
— |
|
* No change. SOURCE: Modified from Sawka et al. (1983). |
||||||
significant, the magnitude of the effects was reported to be smaller for treadmill and cycle-ergometer exercise.
Skeletal Muscle Metabolism
Several investigations examined the effects of environmental heat stress on skeletal muscle metabolism during exercise. Fink et al. (1975) had six subjects perform 45 minutes of cycle exercise (70 to 85 percent of ![]() ) in a cold (9°C) and a hot (41°C) environment. They found greater plasma lactate levels and increased muscle glycogen utilization during exercise in the heat. Also, muscle triglyceride utilization was reduced during exercise in the heat as compared to the cold. In addition, serum glucose concentration increased, and serum triglyceride concentration decreased during exercise in the heat, compared to the opposite responses during exercise in the cold. During exercise in the heat, the increased muscle glycogen utilization was attributed to an increased anaerobic glycolysis resulting from local muscle hypoxia, caused by a reduced muscle blood flow. Because these investigators (Fink et al., 1975) did not perform control experiments in a temperate environment, it is not known if the differences reported are due partially to the effects of the cold exposure.
) in a cold (9°C) and a hot (41°C) environment. They found greater plasma lactate levels and increased muscle glycogen utilization during exercise in the heat. Also, muscle triglyceride utilization was reduced during exercise in the heat as compared to the cold. In addition, serum glucose concentration increased, and serum triglyceride concentration decreased during exercise in the heat, compared to the opposite responses during exercise in the cold. During exercise in the heat, the increased muscle glycogen utilization was attributed to an increased anaerobic glycolysis resulting from local muscle hypoxia, caused by a reduced muscle blood flow. Because these investigators (Fink et al., 1975) did not perform control experiments in a temperate environment, it is not known if the differences reported are due partially to the effects of the cold exposure.
Young et al. (1985) had 13 subjects perform 30 minutes of cycle exercise (70 percent of ![]() ) in a temperate (20°C) and a hot (49°C) environment. They found skeletal muscle and plasma lactate concentrations were greater during exercise in the heat. There was no difference in muscle glycogen utilization between the two experimental conditions. Young et al. (1985) speculated that during exercise in the heat, an alternative glycolytic substrate might have been utilized, such as blood glucose. Rowell et al. (1968) demonstrated a dramatic increase in hepatic glucose release into the blood during exercise in a hot compared to a temperate environment. Such an increased release of hepatic glucose could account for the elevated serum glucose concentration reported in the hot environment by Fink et al. (1975).
) in a temperate (20°C) and a hot (49°C) environment. They found skeletal muscle and plasma lactate concentrations were greater during exercise in the heat. There was no difference in muscle glycogen utilization between the two experimental conditions. Young et al. (1985) speculated that during exercise in the heat, an alternative glycolytic substrate might have been utilized, such as blood glucose. Rowell et al. (1968) demonstrated a dramatic increase in hepatic glucose release into the blood during exercise in a hot compared to a temperate environment. Such an increased release of hepatic glucose could account for the elevated serum glucose concentration reported in the hot environment by Fink et al. (1975).
Data from Dimri et al. (1980) and Young et al. (1985) support the concept of increased anaerobic metabolism during submaximal exercise in the heat. Much of the other support for this concept is based on the findings that, during submaximal exercise, the plasma lactate accumulation is greater in a hot than in a comfortable environment. However, any inference about metabolic effects within the skeletal muscle from changes in plasma lactate is open to debate. Plasma lactate concentration reflects the balance between muscular production, efflux into the blood, and removal from the blood. Rowell et al. (1968) have shown that during exercise in the heat the splanchnic vasoconstriction reduced hepatic removal of plasma lactate. Therefore, the greater blood lactate accumulation during submaximal exercise in the heat
can be attributed, at least in part, to a redistribution of blood flow away from the splanchnic tissues.
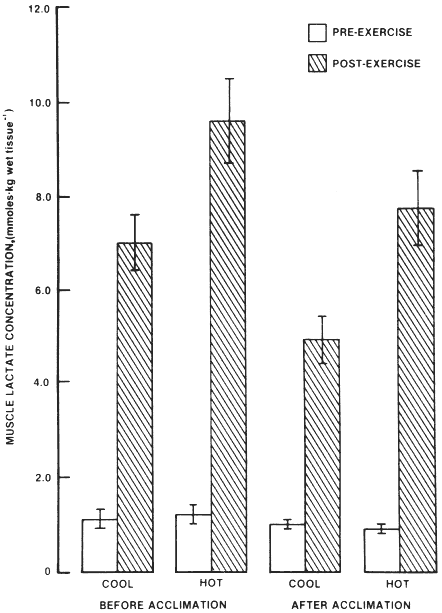
Lactate accumulation in blood and muscle during submaximal exercise is generally found to be reduced following heat acclimatization (Young, 1990). Figure 3-5 shows that heat acclimatization resulted in lower postexercise muscle lactate concentrations. Muscle lactate concentrations were still higher in the heat than in the cool, and changes in blood lactate concentrations followed exactly the same patterns (Young et al., 1985). King et al. (1985) and Kirwan et al. (1987) observed that heat acclimatization reduced muscle glycogen utilization during exercise in the heat by 40 to 50 percent compared to before acclimatization. Young et al. (1985) also observed a statistically significant glycogen sparing effect due to heat acclimatization, but the reduction in glycogen utilization was small and apparent only during exercise in the cool conditions. Glycogen utilization during exercise in the heat was negligibly affected. The mechanism(s) for the reduction in lactate accumulation during exercise associated with heat acclimatization remains unidentified.
Evaporative Heat Loss
Figure 3-1 illustrates that when ambient temperature increases, there is a greater dependence on insensible (evaporative) heat loss to defend core temperature during exercise. In contrast to most animals, respiratory evaporative cooling is small in humans when compared to total skin evaporative cooling. The use of skin provides the advantage of having a greater surface area available for evaporation. The eccrine glands secrete sweat on the skin surface, which is cooled when the sweat evaporates. The rate of evaporation depends on the wetted area, air movement, and the water vapor pressure gradient between the skin and the surrounding air; the wider the gradient, the greater the rate of evaporation.
For a given person, sweating rate is highly variable and depends on environmental conditions (ambient temperature, dew point temperature, radiant load, and air velocity); clothing (insulation and moisture permeability); and physical activity level (Shapiro et al., 1982). Adolph et al. (1947) reported that for 91 men studied during diverse military activities in the desert, the average sweating rate was 4.1 liters every 24 hours, but values ranged from 1 to 11 liters every 24 hours. The water requirements of soldiers on the modern battlefield may be even greater. The threat of chemical warfare may require military personnel to wear nuclear-biological-chemical (NBC) protective clothing, which prevents noxious agents from reaching the skin. Characterized by low moisture permeability and high insulating properties, NBC clothing prevents the normal dissipation of body heat. As a
result, both core and skin temperatures can rise excessively and result in high levels of sweat output, which cannot evaporate within the garments. For example, during light-to moderate-intensity (about 150 to 400 watts) exercise in hot environments, soldiers wearing NBC clothing routinely have sweating rates of 1 to 2 liters per hour (Muza et al., 1988; Pimental et al., 1987).
For athletes, the highest sweating rates occur during prolonged highintensity exercise in the heat. Figure 3-6 (Sawka and Pandolf, 1990) provides an approximation of hourly sweating rates and, therefore, water requirements for runners based on metabolic rate data from several laboratories. The sweating rates were predicted by the equation developed by Shapiro et al. (1982). The amount of body fluid lost as sweat can vary greatly, and sweating rates of 1 liter per hour are very common. The highest sweating
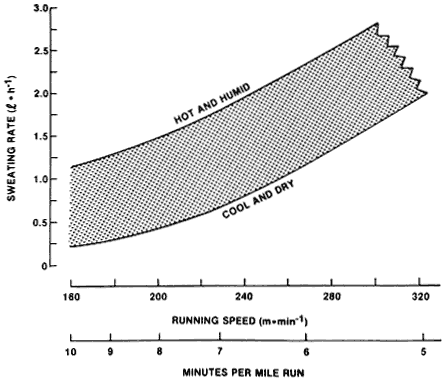
FIGURE 3-6 An approximation of the hourly sweating rates (liters per hour) for runners. Running speed is indicated in meters per minute. SOURCE: Sawka and Pandolf (1990), used with permission.
rate reported in the literature is 3.7 liters per hour, measured for Alberto Salazar during the 1984 Olympic Marathon (Armstrong et al., 1986).
If sweat loss is not fully replaced, the individual's total body water will be decreased (dehydration). Because sweat is more dilute than plasma, dehydration from sweat loss results in an increased plasma tonicity and decreased blood volume, both of which will act to reduce sweat output and skin blood flow (Sawka and Pandolf, 1990). As a result, the body's ability to dissipate heat will be decreased, and dehydration will result in a greater rise in core temperature during exercise-heat stress. In addition, the combination of an elevated core temperature and a reduced blood volume will increase the circulatory strain.
SKIN BLOOD FLOW AND CIRCULATORY RESPONSES
Blood flow from the deep body tissues to the skin transfers heat by convection. When core and skin temperatures are low enough that sweating does not occur, raising skin blood flow brings skin temperature nearer to blood temperature, and lowering skin blood flow brings skin temperature nearer to ambient temperature. This phenomenon allows the body to control sensible (convective and radiative) heat loss by varying skin blood flow and thus skin temperature. In conditions in which sweating occurs, the tendency of skin blood flow to warm the skin is approximately balanced by the tendency of sweating to cool the skin. Therefore, there is usually little change in skin temperature and sensible heat exchange after sweating has begun, and skin blood flow serves primarily to deliver to the skin the heat that is being removed by sweat evaporation. Skin blood flow and sweating thus work in tandem to dissipate heat under such conditions.
During exercise-heat stress, thermoregulatory skin blood flow, although not precisely known, may be as high as 7 liters per minute (Rowell, 1986). The higher skin blood flow will generally, but not always, result in a higher cardiac output, and one might expect the increased work of the heart in pumping this blood to be the major source of cardiovascular strain associated with heat stress. The work of the heart in providing the skin blood flow necessary for thermoregulation in the heat imposes a substantial cardiac strain on patients with severe cardiac disease (Burch and DePasquale, 1962). In healthy subjects, however, the cardiovascular strain associated with stress results mostly from reduced cardiac filling and stroke volume (Figure 3-7), which necessitate a higher heart rate to maintain cardiac output (Nadel et al., 1979; Sawka and Wenger, 1988). This change occurs because the venous bed of the skin is large and compliant and dilates reflexively during heat stress. Therefore, as skin blood flow increases, the blood vessels of the skin become engorged and blood pools in the skin, thus reducing central blood volume and cardiac filling.
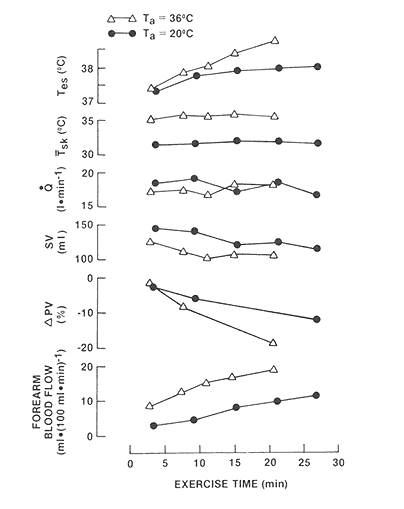
FIGURE 3-7 Thermal and circulatory responses of one subject during cycle exercise at 70 percent ![]() in ambient temperatures (Ta) of 20° and 36°C, showing (from top) esophageal (Tes) and mean skin (Tsk) temperatures, cardiac output (Q), stroke volume (SV), percentage change in plasma volume (PV), and forearm blood flow. SOURCE: Sawka and Wenger (1988), used with permission. Redrawn from Nadel et al. (1979).
in ambient temperatures (Ta) of 20° and 36°C, showing (from top) esophageal (Tes) and mean skin (Tsk) temperatures, cardiac output (Q), stroke volume (SV), percentage change in plasma volume (PV), and forearm blood flow. SOURCE: Sawka and Wenger (1988), used with permission. Redrawn from Nadel et al. (1979).
Several reflex adjustments compensate for peripheral pooling of blood and decreases in blood volume to help maintain cardiac filling, cardiac output, and arterial pressure during exercise-heat stress. Splanchnic and renal blood flows are reduced during exercise in proportion to relative exercise intensity (that is, as a percentage of  (Rowell, 1986). These blood flows also undergo a graded and progressive reduction in subjects who are heated while resting; and in the splanchnic bed, at least, the vasoconstrictor effects of temperature and of exercise appear to be additive, so that at any exercise intensity, the reduction in splanchnic blood flow is greater at a higher skin temperature (Rowell, 1986). Reduction of renal and splanchnic blood flow allows a corresponding diversion of cardiac output to skin and exercising muscle. A substantial volume of blood can thus be mobilized from these beds to help maintain cardiac filling during exercise and heat stress.
(Rowell, 1986). These blood flows also undergo a graded and progressive reduction in subjects who are heated while resting; and in the splanchnic bed, at least, the vasoconstrictor effects of temperature and of exercise appear to be additive, so that at any exercise intensity, the reduction in splanchnic blood flow is greater at a higher skin temperature (Rowell, 1986). Reduction of renal and splanchnic blood flow allows a corresponding diversion of cardiac output to skin and exercising muscle. A substantial volume of blood can thus be mobilized from these beds to help maintain cardiac filling during exercise and heat stress.
During exercise in the heat, the primary cardiovascular challenge is simultaneously to provide sufficient blood flow to exercising skeletal muscle to support metabolism and to provide sufficient blood flow to the skin to dissipate heat. In hot environments, the core-to-skin temperature gradient is less than in cool environments, so that skin blood flow must be relatively high to achieve sufficient heat transfer to maintain thermal balance (Rowell, 1986; Sawka and Wenger, 1988). This high skin blood flow causes pooling of blood in the compliant skin veins, especially below heart level. In addition, as discussed, sweat secretion can result in a net loss of body water, and thereby a reduction in blood volume (Sawka and Pandolf, 1990). Heat stress can reduce cardiac filling through pooling of blood in the skin and through reduced blood volume. Compensatory responses include reductions in splanchnic and renal blood flow; increased cardiac contractility, which helps to defend stroke volume in the face of impaired cardiac filling; and increased heart rate to compensate for decreased stroke volume. If these compensatory responses are insufficient, skin and muscle blood flow will be impaired, possibly leading to dangerous hyperthermia and reduced exercise performance.
SUMMARY
-
Acclimatization state, aerobic fitness and hydration level are important factors influencing a person's ability to dissipate body heat to the environment.
-
The higher the ambient temperature, the greater the dependence on evaporative heat loss to maintain body heat balance.
-
During exercise, the elevation in core temperature is dependent on the metabolic rate, when the environment has sufficient capacity for heat exchange.
-
Heat stress reduces a person's ability to achieve maximal metabolic rates during exercise.
-
Heat stress increases the total metabolic rate and anaerobic participation during submaximal exercise, and these increases are somewhat abated by heat acclimatization.
-
Exercise-heat stress reduces hepatic blood flow and increases hepatic glucose release.
-
Individuals routinely have sweating rates of 1 liter per hour when working in hot environments.
-
Dehydration from sweat loss increases plasma tonicity and decreases blood volume, both of which reduce heat loss and result in elevated core temperature levels during exercise-heat stress.
-
During exercise-heat stress, competing metabolic and thermoregulatory demands for blood flow make it difficult to maintain an adequate cardiac output.
REFERENCES
Adolph, E.F., and associates 1947 Physiology of Man in the Desert. New York: Interscience Publishers.
Armstrong, L.E., and K.B. Pandolf 1988 Physical training, cardiorespiratory physical fitness and exercise-heat tolerance. Pp. 199–226 in Human Performance Physiology and Environmental Medicine at Terrestrial Extremes, K.B. Pandolf, M.N. Sawka, and R.R. Gonzalez , eds. Indianapolis, Ind.: Benchmark Press.
Armstrong, L.E., R.W. Hubbard, B.H. Jones, and J.T. Daniels 1986 Preparing Alberto Salazar for the heat of the 1984 Olympic Marathon. Physician Sportsmed. 14(3):73–81.
Åstrand, I. 1960 Aerobic work capacity in men and women. Acta Physiol. Scand. 49(suppl. 169):64–73.
Brouha, L., P.E. Smith, Jr., R. De Lanne, and M.E. Maxfield 1960 Physiological reactions of men and women during muscular activity and recovery in various environments. J. Appl. Physiol. 16:133–140.
Burch, G.E., and N.P. DePasquale 1962 Hot Climates, Man and His Heart. Springfield, Ill.: Charles C. Thomas.
Cleland, T.S., S.M. Horvath, and M. Phillips 1969 Acclimatization of women to heat after training. Int. Z. Angew. Physiol. 27:15–24.
Consolazio, C.F., R. Shapiro, J.E. Masterson, and P.S.L. McKinzie 1961 Energy requirements of men in extreme heat. J. Nutr. 73:126–134.
Consolazio, C.F., L.O. Matoush, R.A. Nelson, J.A. Torres, and G.J. Isaac 1963 Environmental temperature and energy expenditures. J. Appl. Physiol. 18:65–68.
Dill, D.B., H.T. Edwards, P.S. Bauer, and E.J. Levenson 1930/1931 Physical performance in relation to external temperature. Arbeitsphysiologie 3:508–518.
Dimri, G.P., M.S. Malhotra, J. Sen Gupta, T.S. Kumar, and B.S. Aora 1980 Alterations in aerobic-anaerobic proportions of metabolism during work in heat. Eur. J. Appl. Physiol. 45:43–50.
Eichna, L.W., C.R. Park, N. Nelson, S.M. Horvath, and E.D. Palmes 1950 Thermal regulation during acclimatization in a hot, dry (desert type) environment. Am. J. Physiol. 163:585–597.
Fink, W.J., D.L. Costill, and W.J. Van Handel 1975 Leg muscle metabolism during exercise in the heat and cold. Eur. J. Appl. Physiol. 34:183–190.
Gisolfi, C.V. 1973 Work-heat tolerance derived from interval training. J. Appl. Physiol. 35:349–354.
Gonzalez, R.R., L.G. Berglund, and A.P. Gagge 1978 Indices of thermoregulatory strain for moderate exercise in the heat. J. Appl. Physiol. 44:889–899.
Jooste, P.L., and N.B. Strydom 1979 Improved mechanical efficiency derived from heat acclimation. S. Afr. J. Res. Sport Phys. Ed. Rec. 2:45–53.
King, D.S., D.L. Costill, W.J. Fink, M. Hargreaves, and R. A. Fielding 1985 Muscle metabolism during exercise in the heat in unacclimatized and acclimatized humans. J. Appl. Physiol. 59:1350–1354.
Kirwan, J.P., D.L. Costill, H. Kuipers, M.J. Burrell, W.J. Fink, J.E. Kovaleski, and R.A. Fielding 1987 Substrate utilization in leg muscle of men after heat acclimation J. Appl. Physiol. 63:31–35.
Klausen, K., D.B. Dill, E.E. Phillips, and D. McGregor 1967 Metabolic reactions to work in the desert. J. Appl. Physiol. 22:292–296.
Lind, A.R. 1963 A physiological criterion for setting thermal environmental limits for everyday work. J. Appl. Physiol. 18:51–56.
Muza, S.R., N.A. Pimental, H.M. Cosimini, and M.N. Sawka 1988 Portable ambient air microclimate cooling simulated desert and tropic conditions. Aviat. Space Environ. Med. 59:553–558.
Nadel, E.R. 1983 Effects of temperature on muscle metabolism. Pp. 134–143 in Biochemistry of Exercise, H.G. Knuttgen, J.A. Vogel, and J. Poortmans, eds. Champaign, Ill.: Human Kinetics Publishers.
Nadel, E.R., E. Cafarelli, M.F. Roberts, and C.B. Wenger 1979 Circulatory regulation during exercise in different ambient temperatures. J. Appl. Physiol. 46:430–437.
Nielsen, M. 1938 Die Regulation der Körpertemperatur bei Muskelarbeit. Skand. Arch. Physiol. 79:193–230.
1970 Heat production and body temperature during rest and work. Pp. 205–214 in Physiological and Behavioral Temperature Regulation, J.D. Hardy, A.P. Gagge, and J.A.J. Stolwijk, eds. Springfield, Ill.: Charles C. Thomas.
Petersen, E.S., and H. Vejby-Christensen 1973 Effect of body temperature on steady-state ventilation and metabolism in exercise. Acta Physiol. Scand. 89:342–351.
Pimental, N.A., H.M. Cosimini, M.N. Sawka, and C.B. Wenger 1987 Effectiveness of an air-cooled vest using selected air temperature and humidity combinations. Aviat. Space Environ. Med. 58:119–124.
Piwonka, R.W., and S. Robinson 1967 Acclimation of highly trained men to work in severe heat. J. Appl. Physiol. 22:9–12.
Robinson, S. 1963 Temperature regulation in exercise. Pediatrics 32:691–702.
Robinson, S., D.B. Dill, J.W. Wilson, and M. Nielsen 1941 Adaptations of white men and Negroes to prolonged work in humid heat. Am. J. Trop. Med. 21:261–287.
Rowell, L.B. 1986 Human Circulation: Regulation During Physical Strain. New York: Oxford University Press.
Rowell, L.B., J.R. Blackmon, R.H. Martin, J.A. Mazzarella, and R.A. Bruce 1965 Hepatic clearance of indocyanine green in man under thermal and exercise stresses. J. Appl. Physiol. 20:384–394.
Rowell, L.B., H.J. Marx, R.A. Bruce, R.D. Conn, and F. Kusumi 1966 Reductions in cardiac output, central blood volume and stroke volume with thermal stress in normal men during exercise. J. Clin. Invest. 45:1801–1816.
Rowell, L.B., K.K. Kraning II, J.W. Kennedy, and T.O. Evans 1967 Central circulatory responses to work before and after acclimatization. J. Appl. Physiol. 22:509–518.
Rowell, L.B., G.L. Brengelmann, J.B. Blackmon, R.D. Twiss, and F. Kusumi 1968 Splanchnic blood flow and metabolism in heat-stressed man. J. Appl. Physiol. 24:475–484.
Rowell, L.B., G.L. Brengelmann, J.A. Murray, K.K. Kraning, and F. Kusumi 1969 Human metabolic responses to hyperthermia during mild to maximal exercise. J. Appl. Physiol. 26:395–402.
Saltin, B., and L. Hermansen 1966 Esophageal, rectal and muscle temperature during exercise. J. Appl. Physiol. 21:1757–1762.
Saltin, B., A.P. Gagge, U. Bergh, and J.A.J. Stolwijk 1972 Body temperatures and sweating during exhaustive exercise. J. Appl. Physiol. 32:635–643.
Sawka, M.N., and K.B. Pandolf 1990 Effects of body water loss on exercise performance and physiological functions. Pp. 1–38 in Perspectives in Exercise Science and Sports Medicine. Vol. 3, Fluid Homeostasis During Exercise, C.V. Gisolfi and D.R. Lamb, eds. Indianapolis, Ind.: Benchmark Press.
Sawka, M.N., and C.B. Wenger 1988 Physiological responses to acute exercise-heat stress. Pp. 1–38 in Human Performance Physiology and Environmental Medicine at Terrestrial Extremes, K.B. Pandolf, M.N. Sawka, and R.R. Gonzalez, eds. Indianapolis, Ind.: Benchmark Press.
Sawka, M.N., K.B. Pandolf, B.A. Avellini, and Y. Shapiro 1983 Does heat acclimation lower the rate of metabolism elicited by muscular exercise? Aviat. Space Environ. Med. 54:27–31.
Sawka, M.N., A.J. Young, B.S. Cadarette, L. Levine, and K.B. Pandolf 1985 Influence of heat stress and acclimation on maximal aerobic power. Eur. J. Appl. Physiol. 53:294–298.
Sen Gupta, J., P. Midri, and M.S. Malhotra 1977 Metabolic responses of Indians during sub-maximal and maximal work in dry and humid heat. Ergonomics 20:33–40.
Senay, L.C., and R. Kok 1977 Effects of training and heat acclimatization on blood plasma contents of exercising men. J. Appl. Physiol. 43:591–599.
Shapiro, Y., K.B. Pandolf, and R.F. Goldman 1982 Predicting sweat loss response to exercise, environment and clothing. Eur. J. Appl. Physiol. 48:83–96.
Shvartz, E., Y. Shapiro, A. Magazanik, A. Meroz, H. Birnfeld, A. Mechtinger, and S. Shibolet 1977 Heat acclimation, physical fitness and responses to exercise in temperate and hot environments. J. Appl. Physiol. 43:678–683.
Stolwijk, J.A.J., B. Saltin, and A.P. Gagge 1968 Physiological factors associated with sweating during exercise. Aerospace Med. 39:1101–1105.
Strydom, N.B., C.H. Wyndham, C.G. Williams, J.F. Morrison, G.A.G. Bredell, A.J.S. Benade, and M. Von Rahden 1966 Acclimatization to humid heat and the role of physical conditioning. J. Appl. Physiol. 21:636–642.
Weinman, K.P., Z. Slabochova, E.M. Bernauer, T. Morimoto and F. Sargent II 1967 Reactions of men and women to repeated exposure to humid heat. J. Appl. Physiol. 22:533–538.
Wenger, C.B. 1988 Human heat acclimatization. Pp. 153–197 in Human Performance Physiology and Environmental Medicine at Terrestrial Extremes, K. B. Pandolf, M.N. Sawka, and R.R. Gonzalez, eds. Indianapolis, Ind.: Benchmark Press.
Williams, C.G., G.A.G. Bredell, C.H. Wyndham, N.B. Strydom, J.F. Morrison, J. Peter, P.W. Fleming, and J.S. Ward 1962 Circulatory and metabolic reactions to work in heat. J. Appl. Physiol. 17:625–638.
Wyndham, C.H., G.G. Rogers, L.C. Senay, and D. Mitchell 1976 Acclimatization in a hot, humid environment: Cardiovascular adjustments. J. Appl. Physiol. 40:779–785.
Young, A.J. 1990 Energy substrate utilization during exercise in extreme environments. Pp. 65–117 in Exercise and Sport Sciences Reviews, K.B. Pandolf and J.O. Holloszy, eds. Baltimore, Md.: Williams and Wilkins.
Young, A.J., M.N. Sawka, L. Levine, B.S. Cadarette, and K.B. Pandolf 1985 Skeletal muscle metabolism during exercise is influenced by heat acclimation. J. Appl. Physiol. 59:1929–1935.




















