
1
Genetic Vulnerability and Crop Diversity
Over the ages the tendency of crop improvement efforts has been to select varieties with traits that give the highest return, largely by concentrating on genetic strains that combine the most desirable traits. The resulting homogeneity and uniformity can offer substantial advantages in both the quantity and quality of crop harvested, but this same genetic homogeneity can also reflect greater susceptibility or pathogens. Thus it appears the more that agricultural selection disturbs the natural balance in favor of variety uniformity over large areas, the more vulnerable such varieties are to losses from epidemics. The increased risks presented by genetic selection and the increased cultivation of only a few selected cultivars are easily perceived. Chapters 1 and 2 of this reports focus on crop vulnerability, because it is a broadly recognized problem. The issue of genetic vulnerability, however, is only one of several important problems affecting the management of global genetic resources.
Significant efforts have been made by national and international institutions to collect and preserve crop genetic resources as an insurance policy against future disasters. The recognition, however, that the products of a few major breeding programs are now planted over entire continents under increasingly intensive conditions raises new concerns that are global in scope. These concerns prompted the committee to reassess where the world agriculture community stands today with respect to the risks of genetic vulnerability and progress in diversifying crop gene pools.
The concerns became apparent in 1972 with the report Genetic Vulnerability of Major Crops (National Research Council, 1972). It was prepared by scientists who, alerted by the 1970 epidemic of southern corn leaf blight (Helminthosporium maydis) the United States, became concerned about the potential for similar outbreaks in other major crops. They sought to warn the agricultural community about trends in modern plant breeding programs that may have contributed to the crisis and recommended that the genetic foundations of major crops be diversified to reduce the risk of future outbreaks. Nearly 2 decades later, the issue of genetic vulnerability is still fresh, and debate continues on the risk that it poses.
This chapter addresses genetic vulnerability, first by defining it. The chapter discusses when vulnerability becomes a cause for concern and how the risk it presents can be measured. It examines the trends that contribute to genetic vulnerability and the strategies for reducing it. It evaluates the impact of modern plant breeding programs on genetic vulnerability, and the changes that have occurred in recent years that have increased or decreased genetic vulnerability. Finally, it assesses what remains to be done to minimize vulnerability.
WHAT IS GENETIC VULNERABILITY?
The term genetic vulnerability has been widely used to invoke fears of disastrous epidemics and threats to global food security, often without clarifying what is meant by the term. This chapter uses genetic vulnerability to indicate the condition that results when a crop is uniformly susceptible to a pest, pathogen, or environment hazard as a result of its genetic constitution, thereby creating a potential for disaster.
Critical Factors and Assumptions
Two important factors interact to increase the potential for crop failure: (1) the degree of uniformity for the trait controlling susceptibility to the hazardous agent or environmental stress, and (2) the extent of culture (often monoculture) of the susceptible variety. The greater the uniformity for a susceptible trait and the more extensive the area of cultivation, the greater the risk of disaster. In the case of pest or pathogen attack, two additional factors enhance the risks: (1) a highly dispersible disease or insect agent, and (2) favorable environmental conditions for the multiplication of the agent.
Susceptibility to diseases, insects, and environmental stresses is a natural phenomenon characteristic of all plants, no matter how primitive
or highly bred. Natural epidemics, drought, or other adverse conditions may affect any landraces or cultivars grown by farmers. The extent and impact of the disaster on the food supply and regional economy may, however, be lessened on agricultural systems that incorporate a variety of crop and landraces.
There is cause for concern when extensively planted cultivars of major crops are derived from limited gene pools and, hence, are uniform for a high percentage of traits with narrow based resistances to common pathogens or other agents. These concerns have prompted surveys of plant breeders on their perceptions of the gravity of the problem and a reevaluation of trends in international varietal development and distribution.
Although some breeders and scientist are encouraged by the wider availability of crop gene pools into which exotic plant genes have been introduced, others are worried that genetic uniformity may be increasing on a global scale because of the widespread adoption of modern varieties with similar genetic backgrounds across continents where large numbers and mixtures of landraces were formerly grown. Farmers in developing countries who intensify their cultivation practices and adopt improved varieties may increase their chances of losses from epidemics.
One of the rationales for crop germplasm conservation is that plant genes have utilitarian value. When they are effectively used they can decrease crop susceptibility to natural predators, pathogens, and stresses. Thus, genetic vulnerability can be characterized in terms of genetic resources use. Genetic vulnerability results from the improper or inadequate deployment of these genetic resources, in conjunction with biotic and abiotic stresses. It may also be more inherent in some agricultural systems than in others.
The Role of Uniformity
Genetic uniformity is desirable for many agronomic traits of concern to farmers, processors and consumers and does not, by itself, make a crop vulnerable. In other words, uniformity and susceptibility need not be synonymous. Crops can be vulnerable even when they are genetically quite dissimilar if they have in common a trait (often governed by a single gene) that renders them susceptible to a pathogen, pest, or environmental stress. The chestnut blight (Cryphonectria parasitica) epidemic is a classic case in which genetically heterogeneous species was virtually destroyed by a fungal pathogen that is highly damaging to the American and several other species of chestnut trees (Anagnostakis, 1982). However, such cases are unusual.
More commonly, genetically similar crop varieties that may have in common the same critical gene for resistance to important diseases or insects are grown over large areas in successive seasons. This dependence on a single source of resistance is, arguably, the crux of the problem. Breeders can strengthen plant resistance against epidemics by broadening the diversity of resistance genes and "pyramiding" multiple genes from different sources and genes controlling other mechanisms of resistance. One key to the uniformity issue is understanding the genetic basis of the plant-pathogen, plant-pest, or plant-stress interaction and identifying preventive measures that breeders can take to build in greater resistance. Another is to reduce contiguous areas of land planted to the same varieties.
The Role of Monoculture
Intensive and continuous cultivation of uniform crop varieties enhances opportunities for pathogen or pest evolution and the natural selection of new strains able to attack their hosts successfully. In a monoculture of a single variety or genetically uniform group of
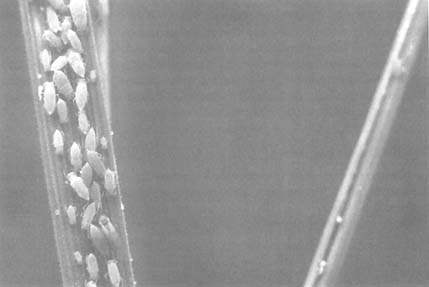
As Russian wheat aphids feed, the wheat leaves tend to roll around them, making them difficult to kill with conventional pesticides. Credit: U.S. Department of Agriculture, Agricultural Research Service.
varieties, the pest must overcome only one genotype, as opposed to numerous genotypes in mixed cultures. Dense stands can facilitate the spread of the attacking population. The effect is particularly severe in tropical environments, where natural reproductive cycles of the insect or disease often are not curtailed by changing climatic conditions. Failure to rotate crops, or sequential planting of two crops that share a common genotype and the same pathogens or insect pests, produces similar results. Nevertheless, the requirements of mechanization and consumer demands follow-cost, high-quality food make it difficult to alter these patterns.
Extensive monocultures clearly pose a risk, yet the spread of closely related, high-yielding varieties (HYVs) continues, despite the risk of vulnerability. Farmers are generally not concerned about vulnerability until production is affected. They depend on plant breeders to supply replacement varieties when existing ones fail. Replacement varieties are often readily available in industrialized countries, but may not be in many developing countries because breeding or seed production programs may be limited or nonexistent.
The Role of Environment
Environmental conditions play both direct and indirect roles in triggering genetic and physiologic plant responses. Weather conditions may promote crop susceptibility. Warm conditions tend to favor the development of fungal and bacterial pathogens, whereas climates that are only periodically moist may enhance damage from insect-vectored viral diseases and seedborne toxins (for example, aflatoxins or toxins of Fusarium kernel rot). Also, the weather greatly influences off-season survival of the attacking population and the amount of off-season inoculum available to initiate a seasonal epidemic. In these situations, networks of collaborating scientists monitoring the effect of environmental conditions on the development and spread of pest problems provide the best prediction of possible vulnerability.
Vulnerability to abiotic stresses may also arise from genetic uniformity for susceptibility. Plants may vary considerably in their tolerance of or susceptibility to abiotic stresses. Drought or excessive salt levels can cause complete crop failure. Cold or hot weather during flowering may induce pollen sterility, and high temperatures may cause flower drop and poor seed set. Although it is sometimes possible to incorporate resistance into crop varieties, the genetic control of stress resistance traits is usually complex. Experimental testing, and thus varietal development, are often difficult.
Measuring Genetic Vulnerability
Three components must be considered when assessing genetic vulnerability. First, the area devoted to major cultivars must be monitored by analyzing volume and distribution of seed sales (which is closely held information in the private sector) through actual farm surveys. Such information was more readily available in the past than it is at present, because many national agricultural agencies (including the U.S. Department of Agriculture [USDA]) have ceased gathering such data in recent years.
Second, trends in pest and pathogen evolution must be monitored to identify those that pose the greatest threats to common cultivars under specified conditions. Given the global patterns of varietal distribution, this should be done on a worldwide basis. For rice, maize, wheat, beans, and the several other commodities, information could be developed through the international agricultural research centers (IARCs). Hundreds of collaborating scientists around the world grow and evaluate breeding lines developed by the IARCs. They collect data on agronomic factors and on susceptibility to pests and pathogens.
Similar public networks (although much more limited in scope) operate for a few crops in the United States through the USDA and state agricultural experiment station collaborators and in Europe. These networks can be excellent sources of information on developing pest or pathogen problems. However, information is not readily available for minor crops and commodities traditionally developed in the private sector (for example, hybrid corn, coffee, bananas, and other plantation crops), although major multinational seed companies often find it worthwhile to develop their own networks. Networking can be a very cost-effective means both of disseminating germplasm and monitoring evolving pest problems.
Third, the extent of genetic uniformity among major cultivars must be determined. In comparison with determination of acreage devoted to major cultivars and pest and pathogen evolution, data on the extent of genetic uniformity among major cultivars are scarce and difficult to assemble. The number of traits involved is enormous, detection methods are laborious, and cultivar pedigrees (when available) usually do not reveal the ecogeographic origins of important resistance genes. Biotechnological methods (see Chapter 7) could be used to develop gene probes and to construct genetic maps, circumventing some of these problems, but the costs are high and the techniques have not yet been broadly applied.
One of the few thorough studies of genetic uniformity was done on rice by scientists based at the International Rice Research Institute
(IRRI). Hargrove et al. (1985) analyzed the diffusion of rice genetic materials among 27 breeding programs over a 20-year period and found a disturbing similarity in the genealogies of improved rice varieties across Asia. Some narrowly focused studies have been done on the geographic distribution of specific traits, such as genes for fertility restoration in hybrid rice based on cytoplasmic male sterile germplasm (Li and Zhu, 1989). Nevertheless, there are few such studies, and key pedigree information is not available for varieties created by the private sector.
The product of these analyses is not a quantitative measurement but a pattern formed by correlation of the trends in the amount of land planted to certain crops, pest-pathogen development, and extent of uniformity for major resistance genes. Congruent with the definition used here for genetic vulnerability, the greatest risk results when widely planted cultivars are exposed to increasingly severe pathogen or pest problems for which they lack broadly based resistance.
Measurement of the trends discussed above, as well as others, indicates the degree to which crops are at risk from genetic vulnerability, a risk that is aggravated by intensive cultivation practices. The following is an examination in greater depth of biological inter-actions that lead to vulnerability and the impact of breeding programs design to manipulate those interactions.
PLANT, PATHOGEN, PEST, AND ENVIRONMENTAL RELATIONSHIPS
The genetic theories of mutation, selection, population biology, and host-pest genetic interaction apply equally to both pathogens (bacteria, viruses, and fungi) and invertebrate pests (aphids, ants, weevils, other arthropods, and nematodes), both of which are used throughout this section as examples. When plant, pest, or pathogen responses to environmental stresses are genetically controlled, the same theories apply.
Evolutionary Nature of the Relationship
A crop variety's responses to a pathogen— whether it is resistance or susceptibility— is a result of its evolutionary and genetic relationship with that organism. This accounts for the phenomenon known as the breakdown of resistance, in which a variety previously resistant becomes susceptible to new forms or races of the pathogen population. In fact, the crop's genetic resistance is unchanged and is still effective against the original pest or pathogen population. It is the genetics of the pathogen that has been altered through natural selection so that it
can attack the crop variety. The variety is then considered susceptible, although no change in its genetics has occurred. The task of the plant breeder becomes to incorporate into the now-susceptible variety additional resistance alleles that are effective against the new pathogen population.
Seldom, if ever, is total genetic protection from disease possible. Resistant varieties are those with significantly decreased disease or pest susceptibility that does not have a negative impact on the growth and biomass production of the plant, and therefore the yield. This decreased susceptibility, however, lasts only until the pathogen or pest population gains new genetic ability to overcome the resistance.
Simmonds (1991) recognizes four broad types of genetically controlled resistance (Table 1-1): major gene or vertical resistance, which is pathotype specific; polygenic or horizontal resistance, which is
TABLE 1-1 The Four Main Kinds of Resistance
|
Kind of Resistance |
Specificity |
Geneticsa |
Durability |
|
Pathotype-specific or vertical, VR/SR |
Very high |
Oligogenes |
Mobile pathogens, durability usually bad |
|
Pathotype-nonspecific major gene resistance, NR |
Nil |
Oligogenes |
Immobile pathogens, durability may be good |
|
General or horizontal resistance, HR/GR |
Nil/low |
Polygenes |
High |
|
Interaction or mixture resistance, IR/MR |
Some |
Heterogeneous oligob |
Probably good |
|
a Oligogenes are single genes that produce a pronounced phenotypic (expressed character) effect, as opposed to polygenes, which have individually small effects. b Some authors (Simmonds, 1979) are inclined to attribute some weight to heterogeneity for polygenic systems, but this matter appears to be undecided. SOURCE: Adapted from Simmonds, N.W. 1988. Synthesis: The strategy of rust resistance breeding. Pp. 119-136 in Breeding Strategies for Resistance to the Rusts of Wheat, N.W. Simmonds and S. Rajaram, eds. Mexico, D.F.: Centro International de Mejoramiento de Maíz y Trigo. Reprinted with permission, ©1988 by Centro Internacional de Mejoramiento de Maíz y Trigo. |
|||
pathotype nonspecific; major gene resistance, which is pathotype nonspecific; and mixture or interaction resistance.
Types of Relationships
Under the genetically controlled resistance patterns described above, the susceptibilities of crop plants to pests or pathogens can develop in many different ways. These host-pathogen relationships fall into three general groups: those characterized by (1) changes in the pathogen or its effect, (2) movement of varieties or pathogens to new biomes, or (3) changes in agronomic practices or pest control strategies. In the first group, major pathogens may evolve, minor pathogens may become more serious, or the environment may enhance the severity of the pathogen's attack. In the second group, a crop may prove to be susceptible to newly encountered pathogens when grown in a new location, or exotic pests introduced into the crop's home region may be successful in attacking it. In the third group, new agronomic traits or practices may render the crop susceptible to new pathogens, or the crop may be vulnerable to previously discounted pests when control strategies or priorities change. Most these responses are intensified by genetic uniformity, ecologic uniformity, of agronomic conditions, or both. Examples illustrating the complexity of interactions that plant breeders must deal with when seeking pest and pathogen resistance in the crops are discussed below.
Evolution of Major Pathogens
Plants with resistance conferred by a single gene or with resistance to a specific pathogen race are said to have vertical resistance. This means they can be damaged by a different virulent race. There are major concerns about varieties that contain vertical resistance genes effective for dealing only with the prevailing races in the breeders' plots. The critical question is: How likely is the new race to develop or to be introduced? Put differently, how soon will there be severe losses because of breaching of the resistance by the rest or pathogen species? The answer depends on how frequently different races arise in the existing pathogen population, how well adapted they are for survival in the off season, and the size and the dispersion of the pest or pathogen population.
The record of single major genes in providing lasting or durable resistance is mixed. The R genes for potato late blight (Phytophthora infestans) resistance and the Sr genes for stem rust resistance in wheat are two classic examples. In both cases, at least some resistant varieties
collapsed because of the selection and spread of new pathogen races able to attack varieties carrying the resistant genes. In wheat, certain combinations of Sr genes in new varieties led to longer-term resistance to stem rust (Puccinia graminis) . In the United States, no widespread epidemic of wheat stem rust has occurred since the late 1950s. However, outbreaks have appeared in other regions.
The biology of the pest and its interaction with the host plant greatly influence the epidemic potential of any new race or biotype that may be selected (Simmonds, 1979). For example, a soilborne fungus that is not also seedborne may develop a new race in a particular field and remain highly localized for decades. In general, vertical resistance genes are ephemeral and when introduced in crops, are soon rendered ineffective by new races of pest and pathogens. This is specially true for cases of pathogens that are windborne (for example, the powdery mildews) or of pests that are highly mobile or that have high reproductive rates (such as the brown plant hopper, (Nilaparvata lugens) .
It is important to consider the evolutionary relationship of the resistance genes and the pest or pathogen. If the two have coevolved in the primary center of diversity, it is highly likely that the pest population contains genes effective against each plant gene conferring resistance (Ashri, 1971; Browning,1972). In developing new varieties, plant breeders must consider whether resistance is sufficiently durable to be economically useful. Several guidelines emerge from past experience. For pests with a high epidemiologic potential that have coevolved with the host plants, major genes for resistance are ephemeral in their utility, if used singly. In some coevolved systems, pyramiding of many genes may work in augmenting the durability of the resistance. Genes for vertical resistance can be used for cases in which experience has shown that selection of new races is slow, the spread of new races is slow, or pathogen survival from season to season is low.
Minor Pathogen Enhancement
In addition to the major pests and pathogens that a breeder addresses in a breading program, there are many others that have only a minor impact on crop performance. It is assumed that selection for high yield will ensure sufficient resistance against yield reduction by these minor pests.
However, minor pathogens can become important if there is a lack of challenge in the breeders' plots and a difference in environmental conditions between the plots and farmers' fields. The inoculum for
the minor pathogen may be so low in the plots that increased susceptibility will not be detected. Changes in the agronomic environment could result in an increased pathogen population. Thus, without any shift in the genetics of the pathogen, a minor pest may become a major problem. A good example is the recent development of gray leafspot disease (Cercospora zeae-maydis) of maize into a disease of potential importance in the United States.
Environmental Enhancement of Pathogen Effects
The environment affects disease levels by altering, increasing, or decreasing pest development. Thus, levels of resistance adequate in one environment may be much too low in another.
If the environment is not conducive to pathogen or pest increase. It is difficult to select for resistance. In general, selection in an arid environment not favorable to airborne fungal or bacterial diseases often results in plant genotypes inadequately protected genetically against many fungal and bacterial pathogens when the crop is grown in wetter climates. For viral diseases with insect vectors and for many insect pests, climates that are only periodically moist may favor heavier crop damage.

In 1985, rains broke the worst drought in a decade in the Sahel, but they also caused an outbreak of the Senegalese grasshopper. An observer (right) from the Food and Agriculture Organization examines the remains of a farmer's crop of millet. Credit: Food and Agriculture Organization of the United Nations.
Varieties Vulnerable in New Regions
Not all pest are widely distributed. At one time the major continents contained separately evolving host-pathogen systems. The equilibria of these system have been breached by the movement of crop species and pests into new regions (Buddenhagen, 1977).
When crops are moved between continents or regions, they may encounter pathogen or pest species for the first time, with unpredictable results. Examples of new encounters resulting from the introduction of the crops into the North America are fire blight (Erwinia amylovora) of apples and pears, Pierce's disease of grapes (Xylella fastidiosa ) , and dwarf bunt of wheat (Tilletia controversa) . Examples of such encounters in the tropics are African cassava mosaic virus, cacao swollen shoot virus, maize streak virus, and Moko disease ( Pseudomonas solanacearum) of banana (Buddenhagen,1977).
Lack of pests or pathogens can be serious impediment to an effective breeding program. The potential impact of the organisms on the crop cannot be assessed when new varieties are develop in the absence of a pathogen or pest.
Varieties Vulnerable to Exotic Pathogens
Many of the great epidemics of the past have been caused by the movement of pest into new areas where a crop species has developed in the absence of the pathogen. The late blight (Phytophthora infestans) epidemic of potatoes in Ireland in the 1840s is a well-known example. Dutch elm disease (Ceratocystis ulmi), white pine blister rust (Cronartium ribicola), and chestnut blight (Cryphonectria parasitica) in North America are three more recent examples. Maize, barley, and wheat have all succumbed to rust epidemics (caused by Puccinia spp.) where they have previously escaped on continents distant to their origin—maize in Africa in 1950s, barley in South America in the 1970s, and wheat in Australia in the 1970s (Simmonds and Rajaram,1988).
Some of the great tropical plantation cultures have developed specifically because the crop escaped potentially devastating coevolved pathogens by being moved to distant continents. Coffee in Latin America escaped coffee rust (Hemileia vastatrix) from Asia and Africa until very recently. Bananas in Latin America escaped Sigatoka (Mycosphaerella musicola) until 1932 and the more virulent black Sigatoka (M. fijiensis ) until the 1970s. Rubber in both Africa and Southeast Asia still escapes South American leaf blight (Dothidella ulei) of its Amazonian homeland (Buddenhagen,1977).
Agronomically Related Vulnerability
New cultivars may be more vulnerable to pests because of either new agronomic traits or practices that are changed to use the new cultivars to their fullest yield potential. This is specially true when breeders target whole continents, where there much variability in agronomic technology.
The semidwarfing genes of both wheat and rice have affected the epidemiology of fungal pathogens by reducing plant height and interleaf distance, consequently reducing air circulation and increasing the humidity around the plant that can contribute to enhanced pathogen densities and rate of development. This has been studied intensively in Septoria leaf blight of wheat (Scott et al., 1982, 1985). In another crop, such as beans, changes in plant form for easier mechanical harvesting also reduces the potential of infection by pathogen. An upright plant shape reduces contact with the soil, promotes improved air circulation and lower humidity, and helps reduce infection and spread of diseases.
Breeders often seek to develop varieties with shorter life cycles. Fewer days to maturity may mean fewer pests or pathogen generations during crop growth, or less chance of exposure to weather conditions favorable to the pathogen. However, in tropical latitudes, shorter life cycles nay lead to raising of two successive crops (double cropping) with increased exposure to pests.
A more subtle, highly unpredictable case is when an agronomic gene actually modifies the intrinsic level of pest susceptibility or resistance in the crop. This may due to multiple characteristics affected by a single gene (pleiotropy), or an agronomic gene may be on the same chromosome and near a gene for susceptibility (linked).
The corn blight epidemic of 1969–1970 was the result of susceptibility conferred by a pleiotropic gene. A cytoplasmic (mithochondrial) gene was used by breeders to produce maize plants that did not have viable pollen. This simplified the production of commercial F1 hybrids. However, it was later discovered that this gene, producing what is called Texas (or T) cytoplasmic male sterility, had an additional effect of increasing susceptibility to a race of the fungus Bipolaris maydis. When climatic conditions were right, the fungus reproduced explosively on millions of acres and over many varieties of maize.
Change in Vulnerability to Discounted Pests
All new varieties are, to some degree, susceptible to various pathogens and pests, but it is assumed that this will be relatively insignificant.
For some crops, chemical treatment or other practices are used as controls to protect them. In other cases, occurrence of the pest or pathogen is infrequent, sporadic, or absent; or the level of susceptibility is so low that little damage will result. For many horticultural, orchard, or ornamental crops, product quality is so important that resistance or susceptibility to pests and pathogens is often largely ignored in breeding, on the assumption that these high-priced crops will be protected by chemicals.
BREEDING STRATEGIES AND THEIR IMPACT ON GENETIC DIVERSITY
Modern plant breeding is essentially an evolutionary process, characterized by a more rapid rate of change than that which occur in the slow natural selection of wild species or the landraces of primitive agriculture. Two major differences exist between the processes of early domestication of crop plants and current breeding practices. First, early development of the crop frequently occurred in the center of diversity for coevolving pests and pathogens. Second, during the early evolution of the crop, hybridization often occurred with related wild species. Today, many crops are grown outside their centers of origin and are distant from related wild species or coevolved pathogens. Few breeding programs use wild species or primitive cultivars in the breeding process; instead, they rely on the proven elite breeding lines (Marshall, 1989; Wilkes, 1989 a).
Genetic Diversity in Primitive and Modern Varieties
In most primitive farming systems, genetic diversity within and among the crops was the norm. Genetic heterogeneity, however, did not provide absolute assurance against epidemic pest attacks nor against environmentally caused yield losses. Rather, the rate of loss varied within the population because of both genetic heterogeneity and crop admixtures. Information on the genetics of landraces in primitive agriculture is fragmentary (Janzen, 1973). Little is known about gene flow and interpopulation versus intrapopulation variation, especially when wild relatives are included (Altieri et al., 1987; Landizinsky, 1989).
Modern varieties, in addition to their high-yield capacity, possess resistances or tolerances to many diseases and insect pests and to normally expected environment stresses. It is often alleged that new hybrids and varieties are weak and prone to disease, and that they must be given plentiful supplies of water, fertilizer, and pesticides to well (Fowler and Mooney, 1990; Mooney, 1979, 1983).
Most modern varieties, however, show superior yield under the intensive cultural conditions of modern agriculture that increase competition for water, nutrients, and sunlight and favor the development of pest populations. Under these conditions many older varieties exhibit sterility, poor root growth, lodging, and susceptibility to pests and diseases (Duvick, 1987).
Many genetic traits give varieties their higher-yield potential. For example, modern maize hybrids show improved root strength, resistance to fungal species that cause stalk-rots, resistance to heat and drought, ability to withstand poor nitrogen nutrition, resistance to the European corn borer (Ostrinia nubilalis) , and ability to grow when densely planted. Because these genetic improvements are typically present with the new resistance genes, the net result is a so-called pyramiding of resistance and tolerance. Except in extreme environments, new hybrids yield more whether grown under good or bad conditions (Anderson et al., 1988).
Germplasm Varietal Development and Vulnerability
The breeding system of a crop species—whether self- or cross-pollinated—can have a great effect on its genetic diversity. Those that are cross-pollinated, such as many perennials, are highly heterozygous, and self-pollinated crops are more generally homozygous. However there are many exceptions. Even though maize has a long history of cultivation as a cross-pollinated, heterozygous crop modern maize hybrids are genetically highly uniform. Although every plant is usually highly heterozygous because it results from the cross of two homozygous inbred lines, it is genetically identical to all other plants of the hybrid.
Most perennial crop species are obligate outcrossers and exhibit a wide array of genetic variability in subsistence agriculture. But modern perennial cultivars intended for large-scale production such as sugarcane, banana, apple, and citrus are often selected as individual plants and are then multiplied clonally to produce many replicates of one heterozygous individual. Such clone are genetically identical to one another as the single-cross maize hybrids are, they differ only in the way the selected genotype is multiplied.
Self pollinating annuals, such as wheats, are typically grown as genetically uniform cultivars tracing back to one nearly homozygous selected plants. However, blends of several uniform, but genetically distinct, cultivars are sometimes purposely grown together, thus producing genetically heterogeneous populations. These can impede the
spread of disease and prevent an epidemic provided that effective resistance genes are present in some of the cultivars.
Performance superiority in crop species results from the accumulation of favorable alleles. The use of unadapted germplasm in crosses introduces unfavorable genes and increases the likelihood of breaking favorable linkages that may be difficult to recover. Therefore, breeders tend to rely on crosses between adapted lines in which many favorable alleles and linkage groups may be present in both parents. They are reluctant to use unadapted exotic germplasm for crossing purposes (particularly if they continue to make acceptable progress without its use) and therefore do not make heavy use of landraces and wild and weedy relatives (Duvick, 1984a; Peeters and Galwey, 1988). This is especially true for crops for which there have been long-standing breeding programs, such as the cereals. This strategy increases the odds for steady performance improvement, but it also tends to enhance genetic uniformity.
If a new crop-threatening hazard (for example, a disease or an insect) appears for which genetic resistance is unavailable in the breeder's own nurseries, the breeder is forced to look elsewhere for the needed genes. Improved germplasm in the major crop species can be found in national, regional, and international screening nurseries and as a result of trials conducted by international agricultural research centers, the USDA Agricultural Research Service, and a few land-grant university experiment stations. Advanced lines and varieties undergoing evaluation in such trials are heavily used by many breeders as parents in new crosses. Performance data on unfamiliar material, when available, are more useful to breeders than are germplasm bank descriptor data, because they provide information on productivity and adaptation that is particularly sought by breeders in choosing parental materials.
Exotic germplasm is most often used in the pursuit of long-range goals. It reaches large breeding programs via national, regional, and international evaluation networks and by personal contacts. These same programs generally maintain large breeders' collections of germplasm that are heavily used for hybridization with exotic materials (Centro International de Mejoramiento de Maíz y Trigo, 1985a, 1987a).
The greatest reliance on germplasm bank collections is made by breeders of plant species that have little or no history of improvement. Generally, these are pulses (beans or lentils), some root crops, vegetables, and some industrial crops. For these crops, the breeder who has initiated an improvement program often has no alternative but to use exotic collected material for local evaluation and selection.
The most frequently voiced criticism from breeders who use germplasm banks is the lack of reliable and complete descriptor information and passport data. This forces breeders to screen large numbers of accessions more or less blindly. They would prefer to identify, for initial screening, a smaller number of accessions best suited to the production environment for which they are breeding, but performance data are rarely available. Finally, the diversity of accessions in a collection may be more indicative of the history of collecting efforts in selected geographic regions than of the breadth of genetic diversity of the species (Peeters and Williams, 1984).
Genes identified in primitive materials usually need to be introgressed or backcrossed repeatedly to improved genotypes before breeders will use them. This process is known as germplasm enhancement, or prebreeding. However, unless there is need for an all-out search for a particular trait, breeders may be reluctant to use the older, more primitive accessions preserved in banks. Rather, they would prefer the enhanced materials, which may not be in germplasm banks (see also Chapter 6).
Gains in Productivity
Progress in plant breeding of the major grain crops can, in part, be measured as increase in productivity or yield per unit area (Buddenhagen, 1985). Changes in farming practices, such as increased use of nitrogen fertilizer and better weed and pest control, have also been responsible for major gains in productivity in the major U.S. crops (maize, wheat, sorghum, and soybeans). The most important input, however, has been that of improved varieties and hybrids from breeding programs (Duvick, 1987).
Repeated studies for each of four major crops have shown that about 50 percent of yield gains in the United States over the past 30 to 50 years were due to varietal improvement (Fehr, 1984). The rate of yield gains as a result of genetic improvement has been estimated at about 1 percent per year for each crops and has been greater for wheat (2 percent) and maize (3 percent). Parallel trends are documented in other developed countries, for example in the United Kingdom, where yield gains for wheat progressed at the rate of 3 percent annually during the period from 1949 to 1984 (Silvey, 1986).
Vulnerability and Varietal Replacement Over Time
Varieties are adopted, become popular, eventually lose their popularity and are discontinued for many reasons. Yield is an important consideration
of variety adoption by farmers. The prevailing markets also affect popularity, the rate at which a variety may be adopted, and the rate at which it eventually will be abandoned.
The area devoted to major U.S. crop varieties in 1969 is shown in Table 1-2. Varietal dominance for major grains ranged from six lines used in hybrids grown on at least 71 percent of the land in corn to nine varieties on 50 percent of the land in wheat. Such data raised serious concerns about greater vulnerability due to narrowing of the genetic base (Duvick, 1984a). Unfortunately, USDA no longer monitors the amount of land devoted to certain crops or the varieties of crops that are planted. Thus, it is no longer possible to assess vulnerability in this way.
A question remains if crops are more or less vulnerable today than in 1970. In 1981, Duvick (1984a) examined data from the seed
TABLE 1-2 Area and Farm Value of Major Crops in the United States and Extent to which Small Numbers of Varieties Dominate Crop Area, 1969
|
Crop |
Area in Hectares (106) |
Value in Dollars(106) |
Number of |
Area Planted to Major Varieties (percent) |
|
|
Total Varieties |
Major Varieties |
||||
|
Bean, dry |
0.6 |
143 |
25 |
2 |
60 |
|
Bean, snap |
0.1 |
99 |
70 |
3 |
76 |
|
Cotton |
45.3 |
1,200 |
50 |
3 |
53 |
|
Maizea |
26.8 |
5,200 |
197b |
6 |
71 |
|
Millet |
0.8 |
— |
— |
3 |
100 |
|
Peanut |
0.6 |
312 |
15 |
9 |
95 |
|
Peas |
0.2 |
80 |
50 |
2 |
96 |
|
Potato |
0.6 |
616 |
82 |
4 |
72 |
|
Rice |
0.7 |
449 |
14 |
4 |
65 |
|
Sorghum |
6.8 |
795 |
— |
|
— |
|
Soybean |
17.2 |
2,500 |
62 |
6 |
56 |
|
Sugar beet |
0.6 |
367 |
16 |
2 |
42 |
|
Sweet potato |
0.04 |
63 |
48 |
1 |
69 |
|
Wheat |
18.0 |
1,800 |
269 |
9 |
50 |
|
NOTE: Dashes indicate that the information was not available. a Maize includes seeds, forage, and silage. b Released public inbreds only. SOURCE: National Research Council. 1972. Genetic Vulnerability of Major Crops. Washington, D.C: National Academy Of Sciences. |
|||||
TABLE 1-3 Perception of Vulnerability Among Breeders
industry to estimate the concentrations of the six most widely planted cultivars of cotton, soybean, and wheat and the correspondingly important lines of maize (Table 1-3). Duvick argued that, from 1969, diversity increased and that the lead varieties were replaced in most cases. However, Smith (1988a) argued that increased numbers of released varieties do not necessarily reflect an increase in genetic diversity because many varieties share common ancestries.
The average time for wide-scale use of a successful variety from the time of its introduction is about 7 years (Duvick, 1984a). Farmers often continue to grow a variety as long as it continues to perform well for them, and they sometimes persist for years after experts have warned against its use. This happened in India, where the wheat variety Sonalika was removed from the recommended list of wheat because of known susceptibility to leaf rust (Puccinia recondita) . However, it continues to be planted over vast areas because it can be harvested earlier than many varieties, has desirable grain color, and is suitable for multiple croppings (Dalrymple, 1986a).
The Seed Savers Exchange has documented varietal losses in the North American vegetable seed industry on the basis of U.S. and Canadian mail-order seed catalogs. In 1984, a total of 4,963 nonhybrid varieties were available from 230 companies, but 54 percent of these were offered by one source only. When a second inventory was completed in 1987, there was a net loss of 15 companies, the majority of which were smaller companies that had been rich sources of unique
varieties (Whealy, 1988). The last such comprehensive inventory of commercially available varieties by the USDA was in 1901 and 1902.
In general, most annual field crop varieties change considerably over time. A very different picture emerges for log-lived tree crops such as apples, for which turnover is measured in decades. In these crops there is less of a tendency for genetic change and more of a dependence on chemical and cultural practices to overcome pest and pathogen problems. For both tree and annual crops, recent trends away from heavy chemical control combined with less energy-intensive cultural practices require intensive breeding efforts to maintain crop quality and productivity.
Alternative Strategies to Enhance Diversity and Increase Stability
Modern agricultural practices often use large inputs of fertilizers, herbicides, fungicides, and insecticides, in combination with monoculture, to optimize yields. However, it appears that additional applications of inputs have leveled off (and may be dropping) relative to yields (Duvick, 1987).
In recent years, there has been a growing interest in agricultural systems that mimic natural ecosystems. The stability of early farming systems was due not only to the genetic diversity and heterogeneity of their landraces but also to the spatial separation of farms and the temporal separation of crops that reduced vulnerability to pests and promoted more efficient use of water, nitrogen, and light (Gould, 1983). Alternative agricultural strategies use a mixture of management and technological options that try to take advantage of natural cycles and biological interactions (National Research Council, 1989a). These options include crop rotations to reduce biotic stresses and enhance soil fertility, modified tillage practices for soil conservation and weed control, integrated pest management practices for insect and disease control, and the use of cultivars with enhanced genetic resistance to various stresses (National Research Council, 1989a).
Most IARC programs include some multiple-cropping or farming systems research, such as cropping practices with beans for erosion control in Africa (Centro Internacional de Agricultura Tropical, 1989) or rotation systems for vegetables in Asia (Asian Vegetable Research and Development Center, 1988). A range of alternative rice production practices developed at IRRI is tested at over 180 sites through the Asian Farming Systems Network (International Rice Research Institute, 1985).
Alternative genetic strategies include modern multiline mixtures and relay plantings of distinctly different genotypes that enhance
agroecosystem stability (Wolfe, 1985, 1988). In a multiline variety, individual lines differ in specific resistance genes. The lines are often segregates from common parents but are not isogenic; varietal mixtures may include lines with different genetic backgrounds and resistance genes. Varietal mixtures appear to be the best option to supply heterogeneity for disease resistance (Browning, 1988; Simmonds and Rajaram, 1988; Wolfe, 1988). Their disease-dampening effect is due to a combination of disease restriction factors, a dilution of inoculum that reduces the success rate of sporulation, and slowing of the spread of disease and postponing the development of a broadly effective pathogen (Marshall and Pryor, 1978). In addition, varietal mixtures make no new demands on plant breeding programs because currently available varieties are adequate, in most cases, for mixture development.
Some reports suggest that a field made up of heterogeneous cultivars provides yields at least as good as those provided by the separate components, whether they are multilines (Borlaug, 1958; Browning and Frey, 1969) or varietal mixtures (Brim and Schutz, 1968; Wolfe, 1985). Such mixtures are usually more stable than their component genotypes with respect to environmental interactions (Schutz and Brim, 1971; Simmonds, 1962). However, multiline or varietal mixtures may present problems for the processing industry, especially when there are industrial restrictions, as there are for wheat.
VULNERABILITY AND CROP DIVERSITY SINCE 1970
The 1970 southern corn leaf blight epidemic focused public awareness on the risks of genetic vulnerability in crops. Trends toward genetic uniformity in crop breeding programs and extensive monoculture were identified by the National Research Council (1972) as major factors contributing to the disaster, but there were no comprehensive data with which to measure changes in crop vulnerability over time. In that report, the data on crop production and the area devoted to major crop varieties served as a benchmark for estimating changes in crop vulnerability from varietal turnover and altered breeding strategies 2 decades later.
Informed measures of vulnerability involve a correlation of estimates of genetic uniformity among major varieties and their land areas with trends in pest and pathogen evolution. An increase in the number of available varieties is not a reliable indicator of increasing genetic diversity, because very similar cultivars may be marketed under different names or may share many genes in common with many other varieties. Modern methods (isoenzyme electrophoresis, high-performance liquid chromatography, DNA restriction fragment
length polymorphism analysis, and gene mapping) provide more precise means for measuring genetic similarity between two cultivars, but they cannot be used to predict susceptibility to unknown pests or pathogens. The following are assessments of changes in diversity of dry beans, wheat, maize, rice, and several minor crops.
Dry Beans
Dry beans (Phaseolus vulgaris L.) Offer a unique example of a crop for which susceptibility to an epidemic was accurately predicted. The National Research Council (1972:225) report recognized that "for a considerable part of the edible dry bean acreage in the United States, annual production rests upon a dangerously small germplasm base." At that time, two market classes of P.vulgaris, the Michigan navy bean and the pinto bean, accounted for 60 percent of 600,000 ha planted in the United States in 1969 and 1970.
Within 5 years of the report, one of the authors had refined a predictive model for dry bean genetic vulnerability, which suggested that pinto beans faced the highest risk of an epidemic of any dry bean commodity class (Adams, 1977). Adams calculated a genetic distance index based on 36 chemical and agronomic traits of each cultivar to assess the homogeneity of beans grown in each production region. By combining these estimates of genetic similarity with the acreage covered by each cultivar, Adams calculated a homogeneity index for each bean-producing state. The assumption was that if the homogeneity of cultivars in a region was great, less time would be required for a pathogen to adapt to them as hosts and the infection rate would be higher.
Within five growing seasons, the pinto bean fields in Colorado and Wyoming — the two states at greatest risk—suffered an epidemic of rust (Uromyces appendiculatus) that caused damage of $15 million to $20 million in 1982 alone (Stavely, 1983). Pinto bean yield losses ranged from 25 to 50 percent in this region in 1981 and 1982 (Venette and Jones, 1982), causing the widespread destruction clearly recognized as a potential in dryland regions in the 1972 report. Yet, the majority of pinto bean farmers continued to grow the rust-susceptible cultivars up until the time of the epidemic. Today, susceptible varieties continue to occupy 40 percent of the land planted to pinto beans in Colorado, southwestern Nebraska, and northwestern Kansas, despite another, lighter rust epidemic in this area in 1987 (Stavely, 1988). Nevertheless, the 1981-1982 epidemic prompted a significant continuing trend to develop rust-resistant cultivars.
The severity of rust led to a multi-institutional commitment to
resistance breeding that has produced dramatic results. Since 1983, when USDA enabled one of its pathologists to concentrate on bean rust, 70 pathogenic races of U. appendiculatus have been identified from field samples, and 1,118 USDA plant introduction bean accessions have been evaluated for rust susceptibility (Stavely, 1988). Two breeding lines of pinto beans have been released that are homozygous for dominant genes for resistance to 33 races of rust and homozygous for a second independent gene for resistance to many of the same races.
There are now both privately and publicly bred pinto bean cultivars in production with resistance to some of the prevalent rust races in the High Plains of the United States. The area planted to pinto beans is shared by several different cultivars and is far more heterogeneous than it was in 1970. Breeders and pathologists point to the exchange of germplasm with the Centro Internacional de Agricultura Tropical (CIAT, International Center of Tropical Agriculture) as one of the major factors that allowed them to diversify the gene pool of pinto beans. Latin American germplasm screened by CIAT has proved particularly valuable, and USDA has sent samples of its rust-resistant accessions to CIAT and other breeding programs overseas as well.
Wheat
Wheat (Triticum spp.) illustrates two conflicting trends in genetic diversity. In the United States, there has been a significant increase in the number of cultivars and the range of exotic germplasm from which they are derived (Cox et al., 1988a; Dalrymple, 1988). At the same time, farmers in developing countries are using fewer landraces and cultivars because of the success of high-yielding cultivars. Their replacement of landraces in wheat's centers of origin continues to concern genetic conservationists (Centro Internacional de Mejoramiento de Maíz y Trigo, 1989).
More than 20 years ago, a report of the National Research Council (1972:135) warned that a "highly vulnerable situation exists in the United States, where a relatively small number of varieties dominates the wheat acreage." Duvick's (1984a) survey of plant breeders suggested that the concentration of U.S. land planted to the six leading cultivars of wheat was about as great in 1980 as it was in 1970. A survey covering a longer time span, however, indicated a steady increase in the number of wheat varieties grown each year since 1949; the increase was from 263 varieties in 1969 to 429 varieties in 1984 (Dalrymple, 1988). In addition, the amount of land covered by the leading wheat variety and the other top 10 varieties has declined
dramatically since 1969, as more privately bred varieties have come to share the market. Nevertheless, caution must be used in assessing the extent of new genetic diversity, because many varieties may be closely related.
Cox et al. (1986) analyzed the common parentage and genetic similarities of wheat cultivars grown in the United States within the previous 10 years. The pedigrees of all 400 U.S. winter wheat varieties may be traced to 74 ancestors, with the bulk of the germplasm still derived from a small number of pre-1919 varieties. At the same time, the use of exotic germplasm by wheat breeders is increasing. Genes from exotic landraces or other species occurred in a quarter of the soft red winter wheats and 7 percent of the hard red winter wheats grown on at least 16,000 ha in 1984. More than 60 percent of the hard red winter wheat breeding lines in the U.S. Southern Regional Performance Nursery now contain chromosome segments from other species. Of 17 wheat breeders contacted in a recent survey, 14 were making crosses with other species, accounting for about 2 to 4 percent of their crosses, advanced lines, and releases (Cox et al., 1988a).
Exotic germplasm is being used to reduce genetic susceptibility of wheat cultivars to powdery mildew (Erysiphe graminis), dwarf bunt (Tilletia controversa), stripe rust (Puccinia striiformis), Hessian fly (Mayetiola destructor), stem rust (P. graminis), leaf rust (P. graminis), Septoria leaf blotch (Septoria nodorum), and greenbug (Schizaphis graminum) (Cox et al., 1988a). The resistance will sometimes last longer in cultivars in which two or more different genes for it are bred into the same cultivar (Gould, 1986). This strategy, termed pyramiding, is particularly promising for cultivars grown in the presence of rapidly evolving Hessian fly biotypes (Gallun and Khush, 1980; Kiyosawa, 1982).
Trends in developing countries provide a dramatically different picture. There, a high percentage of wheat lands are planted to a small number of high-yielding, semidwarf varieties (Figures 1-1 and 1-2). In Bangladesh, for example, HYVs covered about 96 percent of the wheat area in 1984 with 67 percent of the wheat land planted to the variety Sonalika (Dalrymple, 1986a).
In India, about 76 percent of the wheat area was planted to HYVs in 1983. Sonalika, a variety developed in India and released in 1967, alone occupied 30 percent of India's wheat land (Dalrymple, 1986a). India has suffered several severe wheat epidemics since 1969. Shoot fly (Atherigona spp.) killed 50 to 100 percent of all tillers on semidwarf and triple-dwarf HYVs in 1974, and Karnal but (Tilletia indica) affected 67 percent of the Sonalika crop and eight other HYVs in 1975.
The wheat varieties in Mexico appear to be somewhat more diverse
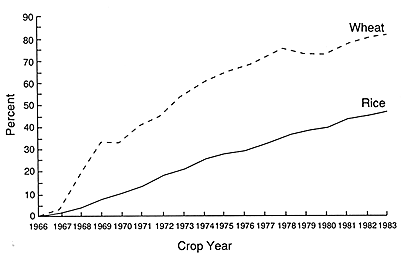
FIGURE 1-1 The estimated proportion of areas planted with high-yielding varieties of wheat and rice in southern and southeastern Asian nations increased between 1965–1966 and 1982–1983. Source: Dalrymple, D. G. 1986. Development and Spread of High-Yielding Rice Varieties in Developing Countries. Washington, D.C.: U.S. Agency for International Development.
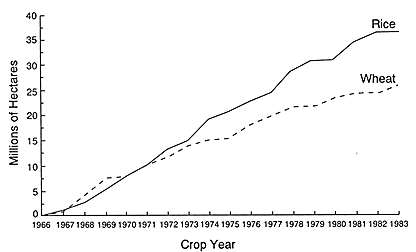
FIGURE 1-2 The estimated area planted to high-yielding varieties of wheat and rice in southern and southeastern Asian nations increased steadily between 1965–1966 and 1982–1983. Source: Dalrymple, D. G. 1986. Development and Spread of High-Yielding Rice Varieties in Developing Countries. Washington, D.C.: U.S. Agency for International Development.
than those of other developing countries. In 1983, HYVs occupied 95 percent or more of Mexican wheat fields, with about 32 percent of the area planted to three varieties. From 1976 to 1981, the breakdown of leaf rust resistance in HYVs such as Nacozari caused a rapid turnover in widely planted varieties (Centro Internacional de Mejoramiento de Maíz y Trigo, 1985b).
The past 2 decades have seen positive trends toward the greater use of exotic germplasm in U.S. wheat breeding programs, increased numbers of available varieties, and decreased dominance by a few leading varieties. Similar trends have not been seen in developing countries. Rather there is a worrisome dominance by a small number of HYVs. Multiplication and distribution of a greater number and diversity of varieties are restricted in developing countries by the typically low capacities of national seed agencies and limited private sector involvement.
Maize
The National Research Council (1972) suggested that too little genetic diversity was available in U.S. maize varieties and recommended the restoration of genetic diversity through the development of new and unrelated types of maize resistant to southern corn leaf blight ( Biopolaris maydis) and other hazards. In 1981, questionnaires were sent to public and private plant breeders in the United States on this an related topics (Duvick, 1984a). The dominance of the six most popular inbred lines of maize decreased from 71 percent of the land planted to hybrids in 1969 to 43 percent in 1980. At the same time, only 5 percent of the maize breeders believed that genetic vulnerability was a major problem for maize, 32 percent felt it was of some concern, and 64 percent doubted that it was a serious concern (Table 1-3). Duvick (1984a) concluded that breeders felt they could produce new resistant cultivars quickly enough to keep farmers from suffering dramatic yield or economic losses.
The major inbred lines used in maize breeding had undergone considerable turnover since 1970, and novel sources of resistance continued to be discovered in elite lines. The message was that the U.S. maize industry had already built in four strategies for reducing the magnitude of its vulnerability: diversity through time (varietal turnover), diversity through anticipation (early-warning alerts), diversity in reserves (numerous advanced lines), and rapid deployment of genetic diversity (the ability of the U.S. seed industry to bring in materials from anywhere in the world).
Other surveys have interpreted changes in the maize industry
differently. Zuber (1975) concluded that the U.S. maize germplasm base was nearly static from 1970 to 1975. Only after 1975 did large changes in the frequency of use of major inbred lines begin, but two germplasm sources (Iowa Stiff Stalk Synthetic and Lancaster Sure Crop) continued to dominate (Zuber and Darrah, 1981). Since 1980, the use of privately available inbred lines as opposed to publicly developed lines of maize has increased dramatically (Darrah and Zuber, 1986).
Many of the maize hybrids sold under different names, however, continue to depend on a few closely related inbred lines. Thus, the apparent changes through time may not be as dramatic as some studies suggest. Comparing isozyme data for 138 commercial hybrids used in 1986 with the set of hybrids used in 1981, Smith (1988a) concluded that genetic diversity in U.S. hybrid maize grown in the Corn Belt has remained at an identical level quantitatively. However, the time period for comparison was short in relation to the rate of varietal turnover. Nevertheless, most surveys have shown that there is little immediate prospect for a large-scale increase in genetic diversity of hybrid maize (Cox et al., 1988a; Goodman, 1985; Zuber and Darrah, 1981).
Maize breeders claimed that their programs had broader germplasm bases in 1981 than they did in 1970 (Duvick, 1984a). By 1986, however, very few genes from exotic germplasm and none from related species had been bred into advanced lines or cultivars. Overall, exotic germplasm is rarely used in hybrids grown in the United States. Less than 1 percent of U.S. hybrid maize has exotic germplasm traceable to sources other than North American landraces and lines (Cox et al., 1988a). Lack of adequate facilities to evaluate materials that are sensitive to day length hamper the use of tropical germplasm in the United States (National Research Council, 1991a). Some evaluation of 400 tropical accessions with promising traits for temperate breeding programs has been done in the southern states of Florida and Texas (Castillo-Gonzalez and Goodman, 1989) and breeding materials adapted to the United States are being developed from elite tropical sources (Holley and Goodman, 1988).
Numerous tolerance and resistance traits can be obtained from exotic as well as adapted maize varieties. Exotic germplasm is a source of resistance to maize bushy stunt mycoplasma, rust (Puccinia sp.), leaf blight, maize streak virus, maize chlorotic dwarf virus, maize chlorotic mottle virus, rootworms (Crambus caliginosellus) , stalk borders (Papaipema nebris), and earworms (Heliothis zea) (Cox et al., 1988a; Nault and Findley, 1981; Prescott-Allen and Prescott-Allen, 1986). Exotic landraces and other species are the only known sources of resistance
to certain viruses (Nault and Findley, 1981). Many maize breeders report that they can also find resistance in already adapted elite lines (Duvick, 1984a). They may be picking up resistance factors from the earlier introgression of exotic genes, however. Continuation of introgression of exotic germplasm into adapted materials can be valuable for future breeding maize programs and most likely will fall to public sector and international programs like Centro Internacional de Mejoramiento de Maíz y Trigo (CIMMYT, International Maize and Wheat Improvement Center).
The international maize outlook has changed considerably over the past 20 to 30 years. During the period 1961–1965 to 1983–1985, world maize production increased by 84 percent, representing an annual growth rate of 4 percent (2.8 percent in developing countries) (Centro Internacional de Mejoramiento de Maíz y Trigo, 1987b). By 1985, improved genotypes were grown on 40 million ha or about one-half of the total maize area in developing countries. Thirty million hectares are planted with hybrids, with the People's Republic of China, Brazil, and Argentina accounting for 80 percent of that area (Centro Internacional de Mejoramiento de Maíz y Trigo, 1987b). Ten million ha are planted with improved open-pollinated varieties. In the remaining half of the total maize area in developing countries, however, adoption rates of improved varieties are low. This may be due to seed production systems that restrict the diffusion of improved varieties (Timothy et al., 1988) or because farmers prefer landraces.
The rate of variety turnover to hybrid maize in developing countries can be illustrated by using Kenya as an example. Maize is the major staple in Kenya. Until the 1950s, nearly all varieties grown in Kenya were local landraces traceable to Tuxpeno-Hickory King landraces brought from North America in the nineteenth century (Timothy et al., 1988). In the late 1950s, two exotic varieties from Latin America, Ecuador 573 and Costa Rica 76, were introduced as breeding stocks to hybridize with local landraces. By the mid-1960s, the first inbred lines and hybrids were released in Kenya and were grown on half of the large-scale farms there. The impact of the hybrids' high yields was so great (30 percent higher than earlier synthetic hybrids) that Kenya's hybrid maize area increased from 0 to 600,000 ha between 1963 and 1981. Both large- and small-scale farmers have become dependent on hybrids, so that 61 percent of Kenya's total maize area was devoted to hybrids in 1985, and another 5 percent to improved open-pollinated varieties (Timothy et al., 1988).
CIMMYT has been one of the primary suppliers of germplasm to Kenyan national programs, even though most of the CIMMYT populations are generally susceptible to common rust (Puccinia sp.) and
turcicum blight (Helminthosporium turcicum), both of which are problems in Kenya (Timothy et al., 1988). Streak virus tolerance has been transferred to inbred lines from a maize composite developed by the International Institute of Tropical Agriculture (IITA). Despite these introductions of exotic germplasm, the germplasm, base in Kenya remains limited because of the policies of the Kenyan Seed Company. From 1963 to 1981, the Kenyan Seed Company and its government-controlled predecessors released only five open-pollinated varieties, four varietal crosses, three double-cross hybrids, two three-way-cross hybrids, and two top-cross hybrids; some of these used the same CIMMYT parents (Timothy et al., 1988). It is therefore not surprising that a severe outbreak of leaf blight (Alternaria triticina) began in 1978 and became widespread by 1979 (Singh et al., 1979). It affected two HYVs and several of the remaining local landraces as well. A major concern is that the Kenyan and other nationalized seed industries in developing countries may be modeling their breeding programs after the IARCs, but are not yet able to respond rapidly to resistance breakdown in released cultivars (Lipton and Longhurst, 1989).
Maize hybrids developed in both the private and public sectors have spread to developing countries where they were previously absent and, for the present, have brought novel germplasm to those countries. Dominance by just a few hybrids or a few sources of germplasm may now be of concern in some of those countries. However, 49 percent of the total maize-growing area in developing countries remains planted to locally adapted landraces and the progeny of purchased, open-pollinated seed (Centro Internacional de Mejoramiento de Maíz y Trigo, 1987b).
Those countries with a high proportion of area sown to the new HYVs face a dilemma. They probably cannot return to cultivating the indigenous maize varieties without reducing yields, yet they cannot continue to plant the same high-yielding varieties indefinitely because new pest races likely will appear, causing disastrous epidemics. The vulnerability is particularly acute in tropical and subtropical areas that lack a cold season (and often little dry season). Lack of support for public plant breeding efforts in many developing countries makes it unlikely that they will be able to mobilize new varieties in sufficient time to prevent disaster.
Rice
Genetic uniformity is a greater concern in rice than it is in wheat because of this crop's limited diversity for three major traits: the single locus for semidwarfism (sd - 1); a common cytoplasmic ancestry
for many HYVs; and a widely used, single source of cytoplasmic male sterility in Chinese hybrid rices (Chang, 1984a; Chang et al., 1985).
Cytoplasmic uniformity, a concern in maize since 1970, is now a concern in rice and other major crops (Hargrove et al., 1980). Resistance or susceptibility to disease is influenced by cytoplasmic inheritance not only in maize and rice but also in strawberries, potatoes, wheat, grain sorghum, cotton, and grapes(Hargrove et al., 1980). In rice, 38 percent of the female parents used worldwide in rice breeding in 1983 and 1984 traced maternally to one Chinese-Indonesian variety named Cina, which is commonly used at IRRI and elsewhere. The use of semidwarf parents in breeding began to increase in 1965. Although the use of locally developed semidwarf lines as females has dominated various national breeding programs since then, nearly all of them can be traced back to sd-1 semidwarf ancestors. Hargr et al. (1985:9) concluded that "the genealogies of improved rice varieties across Asia appear disturbingly similar." Although no weakness in Cina cytoplasm or sd-1 semidwarfs has been reported, in strong recommendations were made to diversify the genetic base of future rice varieties.
The area planted to rice HYVs (more than 73 million ha in 1982 and 1983) exceeds those planted to wheat and maize HYVs in developing countries (Dalrymple, 1986b). It is well known that a very small number of varieties accounts for a very large proportion of the area planted to rice HYVs. In Malaysia, for example, three varieties accounted for about 40 percent of all Malaysian rice plantings in 1981 and 1982. In Indonesia, two varieties accounted for 54 percent of the total rice plantings in 1983 and 1984. In the Philippines, it was estimated that two IRRI varieties occupied about 90 percent of the entire rice-growing area during the 1984 dry season (Dalrymple, 1986b).
The widespread planting of a few closely related varieties with intensive cultivation practices and multiple cropping has led to a breakdown in varietal resistance and to outbreaks of tungro virus, grassy stunt virus, and the brown planthopper (Nilaparvata lugens) in parts of Bangladesh, southern India, Indonesia, Malaysia, the Philippines, Thailand, and southern Vietnam, although none of these was as destructive as the southern corn leaf blight was in the United States (Chang, 1988). In contrast, no serious epidemics have been reported from parts of Thailand, where traditional varieties are commonly grown in rotation with HYVs (Chang, 1984a). Multiple and continuous cropping of the same HYV and staggered planting dates across vast production areas have contributed to the breakdown of monogenic (or vertical) resistance because of changing biotypes of
the brown planthopper in the Philippines, Indonesia, Sri Lanka, and Vietnam. Sequentially released new varieties (for example, IR26 with the gene bph-1, IR36 and IR42 with the gene bph-2, and IR56 and IR60 with the gene bph-3) with related genetic backgrounds were each developed by adding a gene to deal with an emerging insect biotype that had rendered resistance in earlier releases ineffective. Although these varieties provided some relief, new pest biotypes soon evolved (Heinrichs et al., 1985). The new insect populations also transmitted virus more effectively, as in the case of the tungro virus transmitted by the green leafhopper (Nephotettix spp.) (Dahal et al., 1990). Through the misuse of host resistance, rice farmers have suffered severe losses from the resulting "boom-and-bust" cycles of oligogenic resistance (Chang, 1984a).
During the 1970s brown planthopper and the associated grassy stunt virus inflicted heavy losses to the rice crop over a broad region (Dyck and Thomas, 1979). Following a decade of planting, the virus resistance gene (Gsv) obtained from Oryza nivara was rendered ineffective when a second biotype of the virus emerged. Similarly, when semidwarfs were widely grown, the predominant stem borers shifted from striped borers (Chillo zacconius) to yellow borers (Scirpophaga incertulas ) (Chang, 1988).
Indiscriminate use of insecticides has also been linked to a resurgence of the brown planthopper in some areas (Chelliah and Heinrichs, 1980). By contrast, Indonesia has made notable progress controlling pest populations by varietal rotation, pest monitoring, community wide pest control, and synchronization of planting dates (Manna et al., 1985).
With more intensive cultivation of fewer varieties, rice diseases and insects continue to grow in number, intensity, and geographic distribution (Chang, 1984a, 1988). Examples include bacterial blight (Xanthomonas campestris), which reduced seed yield by 50 percent, largely on an HYV under irrigation in Niger in 1983 (Reckhaus and Adamou, 1986), and rice yellow mottle virus, which spread from irrigated paddy rice in Niger, Burkina Faso, and Mali and has infected HYVs from IRRI in Côte d'Ivoire and neighboring countries (Awoderu et al.,1987).
The Republic of Korea initially reported spectacular increases in grain yield after using the semidwarf gene in Tong-il and other varieties. In 1978, about 76 percent of the rice area was planted to HYVs. However, a cool season and widespread blast epidemic in 1979 affected the HYVs more severely than they did the traditional varieties and led to heavy losses (Rural Development Administration, 1985). The area of HYVs dropped to 20 percent in 1988, and national rice production slumped.
Hybrid rice in the People's Republic of China represents another potentially vulnerable situation. The F1 hybrids grown in the major rice production areas share a common cytoplasmic male sterility source and the sd-1 locus. The area planted with hybrids has rapidly expanded from 5 million ha in 1979 to 15 million ha in 1990. So far the Wild Abortive cytoplasm remain the most stable source of male sterility (Yuan and Lin, 1989).
Rice faces special pressures from narrowly based diversity for major traits, continuous use of the same variety or close relatives in multiple cropping, dangerously intensive cultivation practices, reliance on monogenic resistance to important insect pests, and warm and humid environments that sustain the multiplication of pests and rapid shifts within insect populations. The genetic base has also narrowed in temperate regions, such as in Japan (Kimura et al., 1986) and the United State (Dilday, 1990). The inclusion of wheat and rice in continuous cropping systems (Bangladesh, the People's Republic of China, and India) poses new risks of developing new pathogens common to the two hosts (Chang, 1988). The custom of growing traditional varieties in rotation with HYVs in different crop seasons, as is practiced in Thailand, can reduce potential crop losses.
Minor Crops
In 1988, the Agricultural Research Service (ARS) of USDA requested that the Crop Advisory Committees (CACs) assess the status of genetic resources utilization, evaluation, conservation, and vulnerability for their particular crops. The USDA has CACs for all major crops and most minor crops of importance to U.S. agriculture (National Research Council, 1991a).
For apples, two cultivars account for more than 50 percent of the entire U.S. crop. The CAC for apples concluded "commercially cultivated apples are so lacking in genetic diversity that they could easily become victim to a catastrophe" (Apple Crop Advisory Committee, 1987:16). Walnuts are represented by a few cultivars, all of which come from a small gene pool of progenitors. Three diseases and two pests do economic damage to the walnut crop.
Prunus cultivars in the United States— peach, plum, cherry, almond, and apricot— have a limited genetic base. Most commercial peach cultivars can be traced back to fewer than a dozen parents, with the variety Chinese Cling represented in the majority of them. Commercial nectarine cultivars can be traced back to four parents. The entire tart cherry industry is based on a single heirloom cultivar, Montmorency. The sweet cherry industry uses the Bing cultivar on

A crop of sunflowers is grown for the production of sunflower oil. Credit: Food and Agriculture Organization of the United Nations.
75 percent of its acreage in the northwestern United States. Apricots have a narrow gene base, whereas plums have the broadest of any of the Prunus crops in the United States. The Prunus CAC considered all of these stone fruit tree crops in the United States to be susceptible to insect and disease pests. Because the ecologic risks associated with certain chemical pesticides limit their use in pest and disease control, vulnerability will likely increase.
The Vitis CAC said that grape is a prime example of genetic vulnerability because of uniformity. It cites example of "the narrow genetic base upon which most of the world's commercial grape production rests" (Grape Commodity Advisory Committee, 1987:10). It suggested that a majority of vineyards in the United States contained grapevines susceptible to a wide range of pests. Pests and diseases are being introduced into formerly "clean environments" (Grape Commodity Advisory Committee, 1987:11). Also, new abiotic stresses from herbicides, ozone, and sulfur dioxide contamination are affecting grapes.
Sugar beets also suffer from a narrow genetic base derived from very few parents. They are vulnerable to four diseases and two major pests. Between 1974 and 1976, the sugar beet crop in the central and western parts of the United States suffered from a powdery mildew epidemic (Erysiphe betae) . Another root crop, the sweet potato, is considered to be in a critical state. One cultivar makes up 75 percent
of sweet potato production in the United States, and 79 percent of all cultivars are derived from just three parents.
Sunflower hybrids grown in the United States also have a very narrow genetic base, with most being derived from a inbred lines produced by USDA. Only one source of cytoplasmic male sterility is used throughout the entire world. Although genes for resistance to some major insect and disease stresses can be found in the remarkably complete assemblage of wild North American sunflower (Heiser, 1975) now 1975) now maintained by USDA, the wild sunflower program has been severely curtailed because of a lack of funds.
Although the CAC reports identify other crops for which the genetic bases have been broadened (cowpeas, peanuts, tomatoes, clover), on the whole, genetic diversity in the minor crops appears to be seriously restricted and to lag behind gains that have been made for major crops. The level of resources it has taken to build in early warnings of potential epidemics and alternate protection strategies for major crops is proportionately reduced or absent for minor crops. Yet, the costs in loss of employment, closure of farms, or disappearance of industries and businesses resulting from an epidemic would be severe, both economically and socially.
Genetic Vulnerability Today
For major crops in developed countries, varietal turnover and the number of varieties planted have increased since 1972, indicating that the overall level of genetic vulnerability may have decreased. The genetic basis of elite germplasm, however, was found to be shallow because of extensively shared ancestry and limited use of exotic germplasm. Continued efforts to broaden the genetic diversity of primary breeding pools of major crops would provide enhanced and more stable resistance to biotic and abiotic stresses.
In the United States most minor crops have extremely narrow genetic bases, and progress in diversifying them lags far behind that in major crops. The relatively small industries associated with many of these crops do not have the resources to incorporate rapidly needed genes for resistance into new cultivars. The fruit crops are particularly vulnerable to epidemics in the near future, because many of the chemicals that have buffered them from disaster over the past 3 decades are now being withdrawn from the market. Small farmers and hand laborers may be adversely affected by the lack of genetic protection bred into these crops.
In the developing world, high-yielding hybrids and modern varieties of the major crops (especially rice and wheat) have come into
dominance within the past 15 to 25 years. The immediate effect of these introductions is to make available novel genes for resistance to widespread diseases to which landraces may have been susceptible. Simultaneously, rapid losses of native diversity are occurring. As a few releases from the same breeding programs come to dominate a developing country's production, increased vulnerability will almost certainly emerge. It is doubtful whether breeding programs in developing countries can react as rapidly to future epidemics as U.S. maize breeders did between 1970 and 1972.
For the past 20 years, leadership in wheat and rice breeding for developing countries has come from CIMMYT and IRRI, respectively. There are indications that these institutes will devote smaller proportions of their effort to variety development in the future. The national breeding programs in developing countries are not nearly as well funded nor as effective as are IARC breeding programs. There is some question, therefore, whether sufficient breeding efforts worldwide will be devoted to developing and using new wheat and rice varieties at frequent intervals for the developing world. This is of particular significance if the IARCs move away from breeding in favor of other activities, such as biotechnology or development of farming systems for marginal environments before national breeding capabilities are more uniformly strengthened.
At least 5 to 10 years of effort are needed to put strong national breeding programs into place or to improve ineffective ones. Those countries with undersized or ineffective programs that are now depending on the IARCs for new HYVs must begin as soon as possible to build their national breeding programs to the size and efficiency that will serve their countries properly. The pooling of several small programs can, in theory, provide a sufficient base for developing and using improved varieties at frequent intervals. Unfortunately, the histories of planned, government-led, intercountry cooperation show few successes. Remarkable skills in management and diplomacy and stable resources are needed to make such cooperation productive. However, commodity-based networks such as the Programa Regional Cooperativo de Papa (PRECODEPA, Cooperative Regional Potato Program) for potatoes and the Latin American Maize Project for maize promise to be effective in disseminating germplasm and promoting crop improvement.
RECOMMENDATIONS
Assessment of genetic vulnerability involves the synthesis of four components: the area devoted to major cultivars, trends in pest and
pathogen evolution, the extent of genetic uniformity for resistance or susceptibility to biotic and abiotic stresses, and the ability for development of replacement cultivars. Such data are essential to monitoring the potential for severe crop losses. Reductions in public sector funding for agricultural research and service agencies have seriously diminished capabilities for monitoring and responding to crop vulnerability in the United States and other nations.
The potential for crop vulnerability must be nationally and globally monitored.
Information on potential threats should be exchanged among cooperating institutions (for example, the Food and Agriculture Organization of the United Nation, IARCs, USDA, and other national agencies). Responsibility for gathering data lies in national authorities that oversee varietal development and use, and in international and multinational organizations that distribute seed widely.
Genetic uniformity per se does not cause crop susceptibility or epidemics; indeed, uniformity for many traits is demanded by farmers, processors, and consumers. When susceptibility to diseases, pests, or environmental stresses exists, then uniformity for that trait (regardless of whether the variety is otherwise homogeneous or heterogeneous) enhances the risk of crop damage.
A wide range of genetic and agronomic strategies should be employed to minimize crop uniformity and consequent susceptibility.
Plant breeders and agronomists should balance the need for uniformity in agronomic traits with the need for diversity in resistance to biotic and abiotic stresses. Extension specialists should work with processor and consumer groups to win acceptance of greater diversity in crop varieties.
Complex interrelationships among plant hosts, pathogens, pests, vectors, and the environment are evolutionary in nature and determine the crop response— whether it will be resistant or susceptible. Pyramiding of resistance genes provides more durable resistance; single-gene resistance is often more easily overcome by evolving pest populations. Varietal mixtures and varietal deployment can be used to supplement genetic manipulation of resistance. Additionally, the genotypes of successive releases must vary significantly to provide genetic diversity over time.
Agricultural systems that mimic natural ecosystems through use of, for example, multiple lines, varietal mixtures, relay cropping, and integrated pest management, are significantly more stable than monocultures of a few genetically close varieties and, in many cases, provide yields at least as good as those of monoculture systems.
Greater emphasis is needed on agronomic management strategies (for example, multiple cropping, crop rotation, tillage practices, crop mixtures, varietal mixtures, multiple lines, integrated pest management, pest trap crops) through extension services and private-sector marketing campaigns as preventative measures to pest and pathogen attack.






































