
2
Mechanisms of Action of Metabolic Modifiers
During the past 10 years extraordinary progress has been made in understanding the mechanisms of action of the metabolic modifiers under discussion. This chapter discusses the biology, structure, mechanisms of action, and treatment effects of somatotropin (ST), β-adrenergic agonists, and anabolic steroids.
SOMATOTROPIN
It is important to appreciate the benefits that have been realized from research in animal agriculture. With respect to ST, the expansion of our knowledge base has enabled scientists to conceptualize and develop strategies to (1) modify carcass composition, (2) improve feed efficiency, and (3) enhance growth rate and milk yield. This exciting new era in animal agriculture offers unprecedented potential and, in large part, depends on our understanding of the basic biological processes that regulate growth and lactation. It is also important to emphasize that further progress in our understanding of these processes should lead to additional increases in both production performance and productive efficiency.
Downs (1930) and Bierring and Nielsen (1932) were the first to show that an alkaline extract of the anterior pituitary gland reduced carcass fat in rats. This was verified by Lee and Schaffer (1934) who reported that pair-fed rats injected with a crude alkaline extract of bovine pituitaries not only gained more weight, but also contained proportionally more muscle and less fat. Their paper introduced the concept that the "growth hormone" of the anterior pituitary gland could affect the quantity of fat in animals. It was not until 1945, however, that growth hormone was isolated from the anterior pituitary (Li et al., 1945). This allowed Li et al. (1948) to conduct the first experiment to show that crude preparations of ST would reduce carcass fat in rats. Rats were treated 6 days/week for 437 days with a graded injection regimen increasing from 0.4 mg/day to 2.0 mg/day. Carcass fat was reduced by 47 percent.
Results of early studies showing that preparations of ST could decrease carcass fat of rats prompted a number of studies to evaluate the effects of pituitary preparations of porcine ST (pST) on growth and carcass composition of pigs (Giles, 1942; Turman and Andrews, 1955; Henricson and Ullberg, 1960). These studies were inconclusive, probably because the ST preparations were not pure. In 1972, Machlin established that treating pigs with pituitary-derived pST significantly improved weight gain and feed efficiency (Machlin, 1972).
The impact of ST on lactation was also recognized during this same interval. Early studies indicated that administration of anterior pituitary extract affected lactation in laboratory animals (Stricker and Grueter, 1928) and increased milk yield of lactating goats (Asdell, 1932). In 1937, Asimov and Krouze conducted the first substantial study involving more than 500 dairy cows and demonstrated that injections of crude pituitary extract increased milk yield (Asimov and Krouze, 1937). During the 1940s, further research established that the component in the crude pituitary extracts that increased milk yield and stimulated growth was the same molecule, ST (Li et al., 1945; Young, 1947). The first study of the effects of long-term administration of bovine ST (bST) on lactation was conducted by Brumby and Hancock (1955). Milk production was increased approximately 50 percent when twin cows received daily injections of bST for 12 weeks. Similar results were obtained in a subsequent study by Machlin (1973) using a more highly purified source of bST.
Prior to the 1980s, studies with ST were limited in domestic livestock. There was little interest in commercial application for two reasons. First, there was no means for producing large quantities of ST, thus, the supply was limited to that extracted, with varying purity, from the pituitary glands of slaughtered animals. Second, the mechanism of action for ST was thought to involve an acute stimulation in the use of body fat reserves. Accordingly, scientists believed it would be effective only in fat dairy cows with a low milk
yield or in growing animals at the end of the finishing period when fat stores are extensive. Advances in biotechnology made it possible to produce large quantities of ST by recombinant DNA technology. This made it feasible to conduct more extensive investigations which demonstrated that the previously proposed mechanisms were wrong. More recent studies have conclusively demonstrated that chronic administration of recombinant ST results in unprecedented increases in milk production and productive efficiency of dairy cows. It also dramatically improves growth rate, carcass composition, and productive efficiency of pigs.
Structure of Somatotropin
Somatotropin is a protein synthesized in and secreted from the anterior pituitary gland. Secretion of ST is regulated by two peptides that act to either stimulate [growth hormone-releasing factor (GRF)] or inhibit (somatostatin) release of ST from the pituitary gland. Somatotropins from domestic livestock contain 191 amino acids and share a high degree of sequence identity (see Figure 2-1). There are 18 positions that differ between the amino acid sequences pST and bST, but only 2 residues differ between ovine ST and bST. Sequence differences between chicken ST (cST) and ST from other species (Figure 2-1) are much greater (e.g., cST has 77 percent sequence

FIGURE 2-1
Comparison of amino acid sequences for somatotropin from different species. pST, porcine somatotropin; bST, bovine somatotropin; oST, ovine somatotropin; cST, chicken somatotropin; hST, human somatotropin; A, Ala; C, Cys; D, Asp; E, Glu; F, Phe; G, Gly; H, His; I, Ile; K, Lys; L, Leu; M, Met; N, Asn; P, Pro; Q, Gln; R, Arg; S, Ser; T, Thr; Y, Tyr; V, Val; W, Trp. Amino acid sequences were from the following: cST (Souza et al., 1984); pST (Seeburg et al., 1983); bST, hST, and oST (Miller and Eberhardt, 1983; Nicoll et al., 1986).
identity to bST). Variants of ST are also produced by the pituitary gland. For example, in the bovine there are four major variants (Bauman and Vernon, 1993).
It has been known for almost 40 years that pituitary preparations of ST from farm animals and other nonprimates are not biologically active in humans (Bennet et al., 1950; Froesch et al., 1957; Raben, 1959; Juskevich and Guyer, 1990). Subsequent research established that this is because there are considerable differences between the amino acid sequences of human ST (hST) and bST or pST (Wallis, 1975, 1989). Because of these differences, bST and pST are unable to effectively bind to the ST receptor on human cells and initiate a biological response (Carr and Friesen, 1976; Lesniak et al., 1977; Moore et al., 1985; Hocquette et al., 1989).
Effects of Somatotropin
GROWTH PERFORMANCE
Numerous studies have been conducted to investigate the effects of different doses of pST on growth performance of pigs. Furthermore, we have a reasonable understanding of the effects pST has on amino acid requirements. Unfortunately, there is less information available on the effects of ST on growth performance of beef cattle, sheep, and chickens, which limits discussion of the effects of ST on nutrient requirements in these species.
Pigs The effects of pST on growth performance have been widely documented. Responses to different doses of pST are summarized in Table 2-1. Magnitude of response in these studies has varied primarily because of experimental design differences including initial body weight, length of study, breed, sex, dose of pST, and diet composition. Despite these differences, however, it is evident that maximally effective doses of pST can increase average daily gain (ADG) by as much as 10 to 20 percent, improve feed efficiency 15 to 30 percent, decrease adipose tissue mass and lipid accretion rates approximately 70 percent, and concurrently increase protein deposition 50 percent. These changes in performance are associated with a decrease in feed intake of approximately 10 to 15 percent.
Some studies have examined the relationship of pST dose to production response (Boyd et al., 1988; Evock et al., 1988; McLaren et al., 1990). These studies have established that the relationship varies considerably among the various parameters that define growth performance (Figure 2-2). For instance, ADG and rates of protein and ash accretion are maximally stimulated at a daily pST dose of about 100 µ/kg BW. In contrast, lipid accretion rate and feed: gain ratio decrease in a more linear manner across a range of pST doses up to 200 µg/kg BW (see Figure 2-2). The difference in the shape of the dose-response curves not only implies that pST affects growth and metabolism of these tissues by mechanisms
TABLE 2-1 Effects of Porcine Somatotropin (pST) on Pig Response
|
pST Dose (µg/kg/day) |
Average Daily Gain (%) |
Feed: Gain (%) |
Carcass Fata (%) |
Carcass Proteinb (%) |
Reference |
|
22 |
10 |
-4 |
NR |
6 |
Chung et al., 1985 |
|
30 |
10 |
-19 |
-18 |
36 |
Etherton et al., 1986 |
|
70 |
14 |
-17 |
-25 |
19 |
Etherton et al., 1987b |
|
100 |
16 |
-24 |
-32 |
37 |
Campbell et al., 1988 |
|
140 |
19 |
-25 |
-68 |
28 |
Evock et al., 1988 |
|
100 |
16 |
-32 |
-51 |
62 |
Campbell et al., 1989a |
|
100 |
36 |
-28 |
-63 |
46 |
Campbell et al., 1989b |
|
70 |
18 |
-22 |
— |
— |
Bryan et al., 1989 |
|
NOTE: NR, no response;—, not evaluated. a Response represents a composite of percent of carcass fat, kilograms of adipose tissue, or lipid accretion rates. b Response represents a composite of percent of carcass protein, kilograms of muscle, or protein accretion rates. |
|||||
that differ but also has important implications from a nutritional perspective. The marked change in compositional growth and growth rate indicate that changes in the dietary nutrient-calorie relationship must be made to support rates of protein deposition observed for a specific dose of pST. As is detailed in Chapter 5, this becomes particularly important in view of the decrease in feed intake of pST-treated pigs.
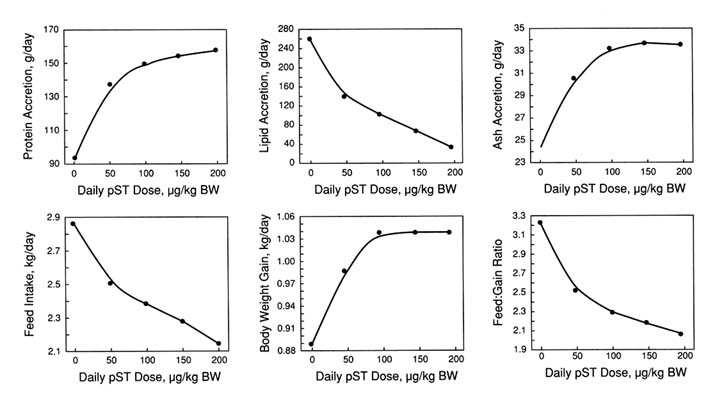
FIGURE 2-2
The dose-response relationship between pST and different parameters of pig growth performance. BW, body weight. The data were summarized from studies reported by Krick et al. (1992).
Somatotropin must be administered by injection because it is not orally active. In studies reported to date, pST has been injected daily, an approach with limited feasibility for treating a large number of pigs. Thus, for the product to be commercially accepted, it is likely that prolonged-release formulations will need to be developed. A few recent abstracts have described the effects of sustained-release
formulations of pST (Knight et al., 1988, 1989; Klindt et al., 1992), and this is an area of active investigation.
Ruminants Studies examining the effect of ST on growth performance of sheep and cattle have been less extensive than those with pigs. Enright (1989) compiled the results of studies in which cattle and sheep were treated with ST. In general, it appears that the ST dose/growth performance response relationships are similar to those observed with pigs but the magnitude of the responses, in particular the effects on rates of protein and lipid accretion, are substantially less. Possible explanations for these differences are discussed in Chapter 4.
Chickens The effects of cST on chicken growth performance are equivocal (see Table 2-2 and Chapter 6) and differ from those observed for other domestic animals. In some studies, administration of cST increases adipose tissue accretion (Leung et al., 1986; Cogburn et al., 1989a), a response that is the opposite of results for growing pigs and cattle. In addition, some studies have shown that cST decreases protein deposition. The most encouraging results are from Vasilatos-Younken and colleagues (1988) in which episodic administration of cST to older birds markedly improved growth performance whereas continuous infusion had no effect. These authors suggested that the pattern of cST administration is important to attain a stimulatory effect.
TABLE 2-2 Effects of Chicken Somatotropin (cST) on Chicken Response
|
cST Dose (µg) |
Average Daily Gain (%) |
Feed: Gain (%) |
Carcass Fata (%) |
Carcass Proteinb (%) |
Reference |
|
50 |
7 |
NRc |
8 |
NR |
Leung et al., 1986 |
|
500 per kg, 3 × per day |
4 |
4 |
5 |
NR |
Burke et al., 1987 |
|
100 per day |
NR |
— |
— |
— |
Bowen et al., 1987 |
|
150 per kg |
21 |
-34c |
-33 |
NR |
Vasilatos-Younken et al., 1988 |
|
200 per kg per day |
NR |
NR |
17 |
-3 |
Cogburn et al., 1989a |
|
NOTE: NR, no response; — not tested. a A response represents a composite of percent of carcass fat, grams adipose tissue, or lipid accretion rates. b A response represents a composite of percent of carcass fat, grams muscle, or protein accretion rates. c Data were presented as body weight gain:feed consumed in this study. |
|||||
TABLE 2-3 Increase in Milk Yield (kg milk/day above controls) in Response to Bovine Somatotropin (bST)
|
|
bST (mg/day) |
||||
|
Controls |
5 |
10-15 |
20-27 |
31-50 |
Reference |
|
24.9 |
+3.6 |
+3.8 |
+5.0 |
+5.7 |
Chalupa and Galligan, 1989a |
|
26.0 |
+2.8 |
+4.1 |
+5.3 |
+6.2 |
Chilliard, 1988b |
|
a Summarized from 7 studies. b Summarized from 20 studies. |
|||||
LACTATION
The effects of bST on milk yield have been documented and discussed in several hundred studies (see reviews by Peel and Bauman, 1987; Chilliard, 1988; Chalupa and Galligan, 1989; Bauman, 1992). Administration of bST following peak milk production (approximately 60 days postpartum) results in a substantial increase in milk yield, and a marked improvement in the persistency of lactation is generally observed. Results from 27 lactation studies are summarized in Table 2-3. Milk yield increases in a dose-dependent manner and the composition of milk is unaltered. Increases of 4 to 6 kg/day have been most frequently observed in long-term studies. In addition, responses have been observed for all dairy breeds examined and in animals of different parity and genetic potential (see Chapter 3).
SUMMARY OF EFFECTS OF SOMATOTROPIN ON GROWTH AND LACTATION
Studies show that ST markedly increases growth performance in pigs and ruminants and enhances milk production in dairy cows. Overall, this results in an impressive improvement in productive efficiency. The effects of ST on chicken growth performance are equivocal at the present time. It should be emphasized that ST is not magic and the greatest
factor affecting the magnitude of response is quality of management (Bauman et al., 1989b). One management factor of particular importance is the nutritional program. Indeed, there are several studies with growing pigs and lactating cows in which the diet was inadequate in amount and/or nutrient balance, and these investigations observed a negligible response to ST.
Mechanisms of Somatotropin Action
In order to cause such dramatic biological effects on growth performance and lactation, it is evident that ST orchestrates many diverse physiological processes to enable more nutrients to be used for lean tissue accretion (during growth) or milk synthesis (during lactation). Research conducted during the past 10 years has indeed established the diversity of physiological effects of ST (see Table 2-4).
ST affects numerous target tissues in ways that are highly coordinated to effect marked changes in nutrient partitioning among these tissues. The biological effects of ST can be broadly classified as either somatogenic or metabolic. The somatogenic effects are those in which ST stimulates cell proliferation. These effects are thought to be mediated by insulin-like growth factor I (IGF-I) (Rechler, 1988). IGF-I is a potent mitogen that shares some sequence similarities with insulin. In contrast, many of the metabolic effects are a direct action of ST. These involve a variety of tissues, and affect the metabolism of all nutrient classes—carbohydrate, lipid, protein, and minerals (see Table 2-4). As a result the coordinated changes in tissue metabolism alter nutrient partitioning and thus play a key role in increasing growth performance or milk yield.
The principal effect of ST is on partitioning of absorbed nutrients. In lactating cows or growing cattle treated with bST, digestibilities of dry matter, carbon, nitrogen, and energy are not altered (Bauman et al., 1989a; Boyd and Bauman, 1989; Chalupa and Galligan, 1989). The energy expenditure for maintenance or the partial efficiency of milk synthesis is not altered in dairy cows treated with bST (Tyrrell et al., 1988; Sechen et al., 1989a; Kirchgessner et al., 1991a). Likewise, studies with growing pigs and cattle have shown that the energetic efficiency of specific processes is not altered. However, maintenance costs are increased by pST administration in pigs, which is consistent with the fact that pST-treated animals have a greater proportion of lean tissue at a given body weight. Verstegen et al. (1989) reported that the maintenance requirement (MEm) increased about 10 percent; Campbell et al. (1988) found that maintenance requirement increased about 17 percent.
The biological effects of ST are initiated by binding to the ST receptor on the target cell. The ST receptor has been cloned for humans (Leung et al., 1987), rabbits (Leung et al., 1987), rats (Baumbach et al., 1989; Mathews et al., 1989), mice (W. C. Smith et al., 1989), cattle (Hauser et al., 1990),
TABLE 2-4 Effects of Somatotropin on Animal Tissue and Systems during Growth or Lactation
|
Tissue/System |
Physiological Effect |
|
Skeletal muscle (growth) |
Increased protein accretion; Increased protein synthesis |
|
Mammary tissue (lactation) |
Increased synthesis of milk with normal composition; Increased uptake of nutrients used for milk synthesis; Increased activity per secretory cell; Increased maintenance of secretory cells; Increased blood flow consistent with change in milk synthesis |
|
Adipose tissue |
Decreased glucose uptake and glucose oxidation; Decreased lipid synthesis, if in positive energy balance; Increased basal lipolysis, if in negative energy balance; Decreased insulin stimulation of glucose metabolism and lipid synthesis; Increased catecholamine stimulated lipolysis; Increased ability of insulin to inhibit lipolysis |
|
Liver |
Increased glucose output; Decreased ability of insulin to inhibit gluconeogenesis |
|
Intestine |
Increased absorption of calcium and phosphorus required for milk (lactation) or bone (growth); Increased ability of vitamin D (1,25-dihydroxycholecalciferol) to stimulate calcium binding protein; Increased calcium binding protein |
|
Various systems |
Increased insulin-like growth factor (IGF)-I and IGF-binding protein (IGFBP)-3; Decreased IGFBP-2; Decreased amino acid oxidation and blood urea nitrogen; Decreased glucose clearance; Decreased glucose oxidation; Decreased response to insulin tolerance test; Increased free fatty acid oxidation, if in negative energy balance; Increased cardiac output consistent with increases in milk output (lactation) |
|
NOTE: References are presented in reviews by Peel and Bauman (1987), Etherton (1989a, b), and Bauman et al. (1989b). |
|
sheep (Adams et al., 1990), and pigs (Cioffi et al., 1990). Based on these studies, the transmembrane ST receptor is a protein of 634 to 638 amino acids with an estimated molecular weight of approximately 70 kDa. Because of similarities in amino acid sequence, the ST receptor is a member of an expanding superfamily of receptor molecules called the cytokine, hematopoietin, or growth hormone/prolactin receptor superfamily (De Meyts, 1992). The extracellular domain consists of about 250 amino acids, the transmembrane domain is comprised of about 25 amino acids, and the intracellular domain has approximately 350 amino acids. A major advance in our understanding of ST
action was made when it was discovered that hST binds to two ST receptor extracellular domains (Ultsch et al., 1991; De Vos et al., 1992). The fact that hST mutants that cannot induce dimerization of ST receptors are biologically inactive (De Vos et al., 1992) suggests that this is an important step in initiating the ST signal transduction pathway.
The remainder of this section discusses the mechanisms by which ST exerts its biological effects. The objective here is to provide an overview of the mechanisms; more specific aspects of ST action in domestic animals are discussed in greater detail in numerous reviews (Etherton and Walton, 1986; Peel and Bauman, 1987; Bauman et al., 1989b; Boyd and Bauman, 1989; Etherton 1989a, b; Vernon and Flint, 1989; Bauman and Vernon, 1993).
LIPID METABOLISM
Changes in lipid metabolism play an integral role in the responses observed in ST-treated animals. The precipitous decrease in the rate of lipid accretion of meat animals is the most graphic illustration of the profound changes that occur in adipocyte metabolism (Figure 2-2). These metabolic changes are important because they (1) establish the rate of lipid accretion and, therefore, the extent to which ST affects carcass composition in a growing animal; (2) play a key role in redirecting nutrients (e.g., glucose) normally destined to be deposited as lipid to other tissues, thereby supporting the nutrient needs for the increased lean tissue accretion during growth or milk synthesis during lactation; and (3) result in improvements in productive efficiency because of the reduction in the proportion of nutrients used for synthesis of body fat.
Accretion of lipid in adipose tissue is a function of the relative rates of lipid synthesis (lipogenesis) and lipid mobilization. Biological effects of ST on lipid metabolism are not acute (as was previously thought) but rather are chronic, and the extent to which they involve lipogenesis versus lipolysis is a function of energy balance. When animals are in positive energy balance ST causes a reduction in rates of lipid synthesis, whereas effects on rates of lipolysis are minimal (Etherton and Walton, 1986; Walton and Etherton, 1986; Sechen et al., 1989a; Dunshea et al., 1992b). This represents the typical situation for growing animals treated with ST but is also observed for bST treatment of lactating cows that are in substantial positive energy balance. In contrast, when animals are in negative energy balance, rates of lipogenesis are already low and ST treatment affects adipose tissue by increasing rates of lipid mobilization (Machlin, 1972; Eisemann et al., 1986a; Bauman et al., 1988). This situation typically occurs in early lactation in dairy cows during the first weeks of treatment (prior to the increase in voluntary intake), but it can also be observed in growing animals when caloric intake is restricted.
In growing pigs, ST affects lipid synthesis in two ways: (1) there is a striking decrease in both glucose uptake and use of glucose for lipid synthesis by adipocytes; (2) the stimulatory effects of insulin on glucose uptake and metabolism are attenuated (Etherton and Smith, 1991; Etherton and Louveau, 1992). Studies conducted with pig adipose tissue cultured in the presence of pST (Walton et al., 1986) or obtained from pST-treated pigs (Walton and Etherton, 1987; Walton et al., 1987; Magri et al., 1990) and in vivo kinetic studies (Dunshea et al., 1992c) have shown that pST markedly decreases glucose uptake, lipogenic enzyme activities, and lipogenic rate. Studies with ovine and bovine adipose tissue cultured with bST (in which acetate incorporation has been studied) have also shown that bST decreases lipogenesis (Vernon, 1982; Etherton et al., 1987a). Thus, the decrease in nutrient utilization for lipid synthesis in adipose tissue enables nutrients to be redirected to other tissues to support the increases in lean tissue growth or milk synthesis.
One of the important regulatory effects of ST is to alter the response of adipose tissue to homeostatic signals. In growing pigs, pST decreases the stimulatory effects of insulin on glucose uptake and utilization by adipose tissue (Walton et al., 1987; Magri et al., 1990) and decreases whole-body response when insulin tolerance tests are conducted (Gopinath and Etherton, 1989b; Wray-Cahen et al., 1990, 1991). This decrease in insulin sensitivity is not associated with any change in insulin binding or insulin receptor tyrosine kinase activity in pig adipocytes (Magri et al., 1990). ST treatment also causes increased lipolytic responses to epinephrine in growing pigs (Wray-Cahen et al., 1991; Novakofski et al., 1988), steers (Peters, 1986; Boisclair et al., 1989b), and lactating cows (McCutcheon and Bauman, 1986; Sechen et al., 1989b, 1990). Other studies have shown that the antilipolytic effects of insulin are increased by bST and that glucose uptake response to insulin in lactating dairy cows is decreased (Sechen et al., 1989b, 1990).
As previously mentioned, changes in lipid metabolism of the bST-treated cow differ somewhat from changes observed for ST treatment of growing animals, and the differences are a function of energy balance. When bST treatment causes cows to be in negative energy balance, lipid mobilization is increased as illustrated by decreases in body fat, chronic elevations of nonesterified fatty acids (NEFA), and increases in milk fat percent and the proportion of long-chain fatty acids found in milk (Bitman et al., 1984; Eppard et al., 1985; Brown et al., 1989). In this situation, the irreversible loss rate (ILR) of NEFA is increased in a manner that is proportional to the extent the cow is in negative energy balance (Bauman et al., 1988). The increase in ILR of NEFA facilitates the decrease in glucose oxidation observed in dairy cows as will be discussed later.
When bST-treated cows are in positive energy balance, adipose tissue lipogenesis is decreased; however, there is no change in NEFA ILR, milk fat percent, or milk fatty acid composition (Eppard et al., 1985; Sechen et al., 1989a). During
chronic bST treatment, feed intake increases; this is associated with a gradual adjustment in lipogenesis that enables the cow to replenish body reserves over the lactation cycle.
It is important to understand that the biological effects of ST are chronic rather than acute. The effects of ST are not observed in short-term (2 hour) incubations with adipose tissue but only become apparent after 24 hours (Walton and Etherton, 1986, 1987; Walton et al., 1986). This suggests that ST acts to inhibit nutrient utilization in adipose tissue by changing the mass of glucose transporter proteins and/or key lipogenic enzymes either by transcriptional or post-transcriptional regulation. Recent evidence provides support for this hypothesis. Mildner and Clarke (1991) have shown that pST decreases fatty acid synthase mRNA levels by 75 percent in pig adipose tissue. Furthermore, when pigs are treated with pST for 7 days, there is a 20 to 40 percent decrease in glucose transporter (GLUT4) mRNA levels in adipose tissue and an associated 40 percent decrease in GLUT4 protein (Etherton and Louveau, 1992). Other studies also have shown that ST decreases acetyl CoA carboxylase activity in cultured sheep adipose tissue (Vernon et al., 1991) and that in vivo ST treatment of lactating cows (Lanna et al., 1992) and pigs (Harris et al., 1990; Liu et al., 1991) reduces enzyme activity. The fact that ST reduces acetyl CoA carboxylase, fatty acid synthase, and GLUT4 mRNA abundance in adipose suggests that ST affects lipid metabolism by altering transcription of these key metabolic genes. Because insulin sensitivity of adipose tissue is reduced and these genes are insulin regulated, it appears that ST acts, in part, by impeding the insulin signal pathway(s), which results in a diminution in transcription of insulin-regulated genes. This insulin antagonistic effect of ST, however, does not appear to universally affect all insulin-regulated genes, since some of the effects (i.e., antilipolytic) of insulin are not reduced by ST (Sechen et al., 1989a, 1990).
During the past 10 years there has been a remarkable increase in our understanding of the physiological effects that ST exerts on adipose tissue of domestic animals. Despite this, little is known about the ST intracellular signal pathway(s) that cause these alterations in lipid metabolism. Furthermore, it remains unclear how the metabolic changes that occur in adipose tissue in response to ST are coordinated with those that take place in the liver, muscle, mammary tissue, and other tissues to effect the remarkable increases in production observed.
CARBOHYDRATE METABOLISM
ST has numerous effects on carbohydrate metabolism (see Table 2-4). This is of particular importance in the dairy cow, in which glucose originates from gluconeogenesis and typically 60 to 85 percent of the glucose turnover is used for milk synthesis. Treatment of cows with bST increases glucose ILR and reduces whole-body glucose oxidation (Bauman et al., 1988). The increase in glucose ILR is the result of an increase in hepatic gluconeogenesis (Pocius and Herbein, 1986; Cohick et al., 1989; Knapp et al., 1992) and one of the tissues that reduces its use of glucose is the hind limb muscle (McDowell et al., 1987). Adaptations in glucose production and oxidation in bST-treated cows are quantitatively equal to the extra glucose required for the increased milk synthesis (Bauman et al., 1988). In pigs (in the postabsorptive state) treated with pST, there is also an increase in hepatic output of glucose (Gopinath and Etherton, 1989b). In both cattle and pigs, liver responses to insulin are decreased (Boisclair et al., 1989a; Gopinath and Etherton, 1989b). In bST-treated lactating cows this is particularly important because the liver is the predominant source of glucose for milk synthesis, and insulin acts to inhibit hepatic production of glucose. Thus, the reduction in hepatic response to insulin allows the liver to sustain the increased rate of gluconeogenesis that is critical to support the increase in milk synthesis.
When pigs are treated chronically with pST, plasma glucose and insulin concentrations are elevated (Gopinath and Etherton, 1989a; Dunshea et al., 1992a). The increase in plasma glucose is most likely related to a reduction in glucose uptake, particularly by adipose tissue. Because a significant quantity of glucose is metabolized in adipose tissue of pigs, a decrease in glucose utilization by adipose tissue redirects a considerable quantity of glucose to other tissues. For example, it has been shown that approximately 40 percent of whole-body glucose uptake measured in the basal state and 25 to 30 percent measured in the insulin-stimulated state are used by adipose tissue of barrows (Dunshea et al., 1992c). In contrast, glucose utilization by adipose tissue of pST-treated pigs amounts to only about 7 percent of whole-body glucose turnover (Dunshea et al., 1992c).
PROTIEN METABOLISM
Less is known about the effects of ST on protein metabolism than about either lipid or carbohydrate metabolism. It is clear the ST treatment increases muscle protein deposition in growing animals and milk protein secretion in lactating cows. No studies have examined the effects of ST on the kinetics of protein metabolism during ST treatment in dairy cows. In growing pigs and cattle, one of the most characteristic responses to ST treatment is a dose-dependent decrease in blood urea nitrogen concentration. This suggests that whole-body oxidation of amino acids and the concomitant hepatic conversion of ammonia to urea are reduced. These adaptations in amino acid metabolism are consistent with an increased use of amino acids for protein accretion.
The kinetics of amino acid metabolism have been examined in growing cattle treated with bST. Eisemann et al. (1986a,b) have reported that ST treatment of beef heifers fed slightly more than maintenance amount increased nitrogen
retention and whole-body protein synthesis without affecting protein degradation. Subsequently, Eisemann et al. (1989b) reexamined this in rapidly growing steers and found that bST treatment increased L-[14C]-leucine use for protein synthesis and that whole-body rates of leucine oxidation decreased. Furthermore, they noted that the additional nitrogen retained was deposited with an incremental efficiency of approximately 50 percent. Other studies with growing lambs and cattle have demonstrated that the fractional rate of protein synthesis is increased in skeletal muscle with bST treatment (Pell and Bates, 1987; Eisemann et al., 1989a). A recent study with growing pigs indicates that pST treatment increased rates of whole-body protein turnover; however, the absolute increment in protein synthesis rate was greater than that for breakdown, leading to the increased net nitrogen retention (Tomas et al., 1992).
One of the critical events that occurs during postnatal muscle growth is an increase in muscle DNA content. This event is important because postnatal accretion of DNA is a key factor in regulating muscle growth (Allen, 1988). This increase is caused by proliferation of satellite cells that reside between the sarcolemma and basement membrane of myofibers. These cells have the ability to fuse with the myofiber and thereby contribute their nucleus to the cell. Thus, during postnatal muscle growth the increase in muscle DNA is coordinated with the increase in muscle protein. As discussed above, ST increases the rate of protein accretion markedly. The effects of ST appear to be not only caused by changes in protein metabolism but also by changes in the rate of satellite cell proliferation. One of the growth factors that has been shown to stimulate proliferation of satellite cells is IGF-I (Allen, 1988). This observation is significant because the mitogenic effects of ST are mediated indirectly by IGF-I (Florini, 1987).
MAMMARY GLAND METABOLISM
Administration of bST to lactating dairy cows results in major adaptations in mammary tissue metabolism. The change that occurs in the shape of the lactation curve with long-term bST administration indicates that the number of secretory cells in the glands and/or the synthetic capacity of each mammary epithelial cell must increase (Bauman et al., 1985). Recent studies with lactating goats have demonstrated that bST treatment prevented the normal decline in mammary cell number and increased the activity of key enzymes involved in milk synthesis (22 weeks; 27 percent increase in milk yield) (Knight et al., 1990). Baldwin (1990) demonstrated that bST-treated cows had increased RNA per mammary gland, indicating an increase in protein synthetic capacity. In addition, scientists have reported significant increases or trends for increases in activities of several enzymes from bST-treated cows and goats (Baldwin, 1990; Knight et al., 1990, 1992).
A major paradox is that bST does not appear to directly mediate its effects on the mammary gland. bST does not have any direct effect on milk synthesis in bovine mammary tissue in vitro (Gertler et al., 1983) and bovine mammary cells appear to lack bST receptors (Akers, 1985; Collier et al., 1989). However, this is not completely resolved; some recent studies have reported mRNA for somatotropin receptors in mammary tissue from several species including cows (Glimm et al., 1990; Jammes et al., 1991). There has been some suggestion that IGF-I may mediate the galactopoietic effects of bST because bovine mammary epithelial cells do have receptors for IGF-I (Collier et al., 1989). Prosser et al. (1989) reported that infusion of IGF-I (1.1 nMol/min) for 6 hours into the pudic artery of lactating goats increased milk secretion by 30 percent. Recently, Davis et al. (1989) conducted a study in which IGF-I was infused (43 nMol/hour via jugular catheter) into lactating goats on days 4 to 6 of a 10-day experimental period. Although bST administration increased milk production, there was no increase in milk yield of the group infused with IGF-I, even though blood concentrations of IGF-I were comparable between the two groups. An additional complication is that IGFs in physiological fluids are bound to soluble, high-affinity IGF-binding proteins (IGFBP). Although we do not have a clear understanding of how the IGF complex is able to mediate mammary function, it is apparent that changes in circulating concentrations of IGF-I and some of the IGFBP are closely tracking the biological events and magnitude of milk responses that occur with bST treatment (see review by Bauman and Vernon, 1993).
Consistent with the increases in milk yield, bST treatment has been shown to increase cardiac output and mammary blood flow (Mepham et al., 1984; Davis et al., 1988; Fullerton et al., 1989). Based on current concepts of mammary biology, it is probable that the alteration in blood flow is a consequence of changes in mammary tissue metabolism rather than the cause of these changes.
Summary of Effects of Somatotropins
Somatotropins alter an array of physiological processes in domestic animals treated chronically with the hormone. These effects are precisely coordinated to alter the flow of nutrients among the tissues of the body. ST alters many metabolic pathways in numerous tissues and changes tissue responses to other endocrine signals. Overall, these alterations in metabolism and cell proliferation lead to the production responses observed in meat and dairy animals. It is likely that as we increase our understanding of how ST functions, we will be able to develop ways to further potentiate the stimulatory effects of ST or identify alternative strategies that increase not only growth performance and milk yield but, more important, the efficiency of production.
β-ADRENERGIC AGONISTS
Synthetic compounds called β-adrenergic agonists exhibit profound effects on growth and metabolism of skeletal muscle and adipose tissue. They share some similarity in structure and function with the naturally occurring catecholamines. Three major catecholamines (dopamine, norepinephrine, and epinephrine) are found in mammals. They circulate in the blood plasma, can act at sites removed from their origin (a relationship that is used to define a hormone), and regulate a wide range of physiological responses in many tissues. Epinephrine in particular, but also norepinephrine, are major regulators of metabolism. Examples of the various physiological actions of catecholamines include the following: regulation of the speed and force of heart contractility, motility and secretory responses of various portions of the gastrointestinal tract, bronchodilation, salivary gland and pancreatic insulin secretion, blood vessel constriction and dilation, uterine contraction, and spleen capsule contraction.
The three endogenous catecholamines are related in structure, biosynthetic sequence, and function. General information regarding catecholamine structure, biosynthesis, metabolism, and adrenergic control of physiological and metabolic function has been reviewed (Martin, 1985; Norman and Litwack, 1987; Timmerman, 1987; Mersmann, 1989b; Weiner and Molinoff, 1989; Hoffman and Lefkowitz, 1990). The chemical structures of dopamine, norepinephrine, and epinephrine are presented in Figure 2-3. Epinephrine is the primary hormone secreted by the adrenal medulla.
External stimuli cause the adrenal medulla to release epinephrine and rapidly elevate the peripheral concentration. Stimulation can cause the release of some norepinephrine from the adrenal medulla as well. The relative plasma concentration of these two naturally occurring catecholamines varies among different species. Norepinephrine is usually present at two-to-five times the concentration of epinephrine under resting conditions, whereas dopamine is present at similar or lower concentrations than epinephrine in most mammalian species (Buhler et al., 1978).
All three catecholamines, but particularly norepinephrine and epinephrine, precipitate an extremely large spectrum of physiological functions either by stimulating central nervous system synaptic activity or direct innervation of an organ by the sympathetic nervous system or by acting as plasma hormones. Response to catecholamines requires the presence of a receptor that will bind the particular neurotransmitter or hormone whose concentration has been increased and then couple the receptor binding to an effective intracellular response system. Because most organs of the mammalian body possess receptors for catecholamines, these substances have a major role in regulating many metabolic processes. For example, catecholamines are instrumental in stimulation of glycogen phosphorylase and inhibition of glycogen synthase to stimulate the production
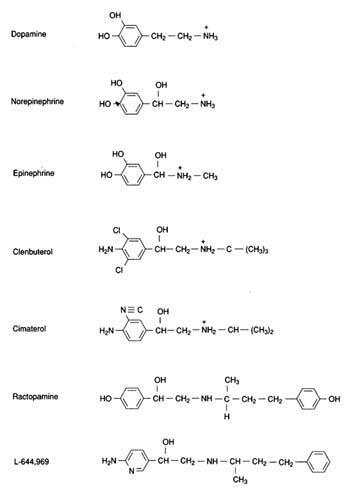
FIGURE 2-3
Chemical structures of the endogenous catecholamines dopamine, norepinephrine, and epinephrine and of select synthetic β-adrenergic agonists.
of glucose from glycogen stores. Catecholamines also stimulate lipolysis to cause the release of free fatty acids from adipose tissue triacylglycerol stores. Thus, catecholamines have a role in control of plasma concentrations of two primary oxidative substrates—glucose and free fatty acids.
Adrenergic Receptors
The many and sometimes antipodal functions regulated by the naturally occurring catecholamines lead to the concept that different receptors must exist in different organs. For example, norepinephrine stimulates mammalian heart contractility at a lower concentration than epinephrine, whereas epinephrine is more potent for stimulation of spleen capsule contraction. Observations such as this led to the concept of distinct α- and α-adrenergic receptors that control various physiological and metabolic functions. This type
of classification was aided by the synthesis of a large number of analogs of norepinephrine. As the number of analogs increased, compounds became available that would preferentially stimulate (or inhibit) a particular function. With an increased spectrum of norepinephrine analogs and continued investigation of the adrenergic control of additional biological functions, it became appropriate to divide the β-adrenergic receptor class into β-1 and β-2 subclasses. Eventually, the β-adrenergic receptor class was divided into β-1 and β-2 subclasses. These classification schemes for adrenergic receptors are attempts to codify biological responses so that the complex functions and plethora of chemical structures can be integrated into a rational pattern.
There have been attempts to establish additional subclasses of receptors based on pharmacological properties and distribution of both α- and α-receptors (Martin, 1985; Norman and Litwack, 1987; Timmerman, 1987; Mersmann, 1989a; Weiner and Molinoff, 1989; Hoffman and Lefkowitz, 1990). Protein purification techniques (Lefkowitz and Caron, 1988) and molecular biology techniques using nucleotide sequences of cDNAs have definitively established the existence of distinct α-1- (Cotecchia et al., 1988), α-2-(Kobilka et al., 1987b), α-1- (Machida et al., 1990), and α-2-(Kobilka et al., 1987a) adrenergic receptors. Molecular characterization of the human α-3-receptor has also been reported (Emorine et al., 1989). It is too early to know the extent of homology between receptor types purported to be α-1- (or α-2- and α-3-) adrenergic receptors when examined in the same tissue in a number of species or in a variety of tissues within a single species.
It is important to note that earlier attempts to classify adrenergic receptors, agonists, and antagonists was confounded by arbitrary selection of ligands, choice of variable biochemical or physiological events, and use of discrete ''all-or-none" response criteria. These physiological data are complemented by ligand binding studies in some but not all cases. In addition, ligand binding can be affected by subtle differences among the same cell type across species or in different cell types within a given species. Receptor classification becomes extremely confounded by use of a variety of species, a multiplicity of analogs, several diverse cell types, and numerous experimental approaches (McGonigle et al., 1986; Neve et al., 1986; Timmerman, 1987; Mersmann, 1989b; Lafontan et al., 1990). Although most of the synthetic adrenergic agonists found to exhibit repartitioning effects on growth of mammalian species have been characterized as β-agonists; whether their effects are directly mediated through the β-receptors is equivocal.
Distribution of adrenergic receptor types is of equal importance in determining the nature or magnitude of a response. It has become apparent that many or even most mammalian organs, tissues, or cell types do not have a pure population of α- or α-adrenergic receptors; rather a mixture of subtypes of these receptors is usually present, albeit at different levels (Minneman et al., 1979). For example, heart contractility usually is considered to be stimulated by α-1-adrenergic receptors, although mammalian heart muscle appears to have both α-1-and α-2-adrenergic receptors. The proportion may vary from essentially 100 percent α-1-adrenergic receptors in the guinea pig ventricle (Hedberg et al., 1980) to 35 percent α-2-adrenergic receptors in the human ventricle (Heitz et al., 1983).
Skeletal muscle has β-adrenergic receptors as evidenced by the stimulation of glycogenolysis and the production of lactate by epinephrine, norepinephrine, and the analog isoproterenol both in vitro and in vivo. The antagonist propranolol inhibits this response. More direct measurement of β-adrenergic receptors by ligand binding techniques also indicates the presence of β-receptors, with the β-2 subtype predominating over the β-1 subtype (Liggett et al., 1988). Expression of β-3-receptor mRNA has been demonstrated in rat skeletal muscle (soleus), as well as adipose, liver, and ileum, but was not observed in brain, skin, heart, and lung (Emorine et al., 1989). Two β-adrenergic agonists that dramatically enhance skeletal muscle deposition, clenbuterol and cimaterol, are purportedly specific for the β-2-receptor subtype (O'Donnell, 1976; Kim and Sainz, 1990), whereas ractopamine is primarily a β-1-agonist (Anderson et al., 1991).
Mammalian adipose tissue cells contain β-adrenergic receptors as indicated by stimulation of lipid breakdown (lipolysis) by epinephrine, norepinephrine, and isoproterenol as well as a number of other norepinephrine analogs both in vitro and in vivo. These effects can be antagonized by propranolol or other β-adrenergic antagonists. Lipogenesis, both fatty acid and triacylglycerol biosynthesis in the adipocyte, is inhibited by β-adrenergic agonists and such effects can be antagonized by β-adrenergic antagonists (Fain and Garcia-Sainz, 1983; Buttery and Dawson, 1987; Timmerman, 1987; Mersmann, 1989a; Yang and McElligott, 1989). Establishment of the β-adrenergic receptor subtypes on the mammalian adipocyte has not been particularly successful. Some studies have indicated the receptor is of the β-1-adrenergic subtype, others indicate a mixture of β-1- and β-2-adrenergic receptors, and yet others indicate that a totally different receptor, the β-3-adrenergic receptor, is also present (Emorine et al., 1989; Lafontan et al., 1990). How much of the diversity in description of the adipose tissue adrenergic receptor subtypes is the result of studies in different species, use of different agonists and antagonists, or use of different methodologies, including ligand binding compared to measurement of cellular function, is not yet understood.
Effects of β-Adrenergic Agonists on Growth and Carcass Composition
Since the early 1980s, several synthetic analogs of epinephrine and norepinephrine have been investigated for their
ability to promote skeletal muscle growth and reduce the fat content of animal carcasses. These orally active materials are usually referred to as β-agonists. Structures of some of the compounds that have been studied and for which effects have been reported in the literature are shown in Figure 2-3. These substances bind predominantly to β-receptors found in the membranes of cells. There are relatively few substances that bind almost exclusively to one type of receptor. Several of the agonists currently being evaluated for use as metabolic modifiers in the livestock industry show a marked specificity toward the β-2-receptor; stimulation of the β-1-receptor results in tachycardia. Transient increases in heart rate and systemic blood flow are observed with dietary administration of cimaterol in lambs (Beermann, 1987) and clenbuterol in cattle (Eisemann et al., 1988).
The β-adrenergic agonists reported in the literature as potential metabolic modifiers are all orally active, unlike ST and most of the anabolic steroids. Their main effects on the carcass are to increase skeletal muscle and reduce adipose tissue mass, with little or no effect on bone. This is sometimes accompanied by an increase in growth rate or feed efficiency. It is not surprising, then, that responses in live-weight gain and feed efficiency are related to the dose rate, with indications that efficacy is reduced at extremely high doses (Ricks et al., 1984; Hanrahan et al., 1986; Reeds et al., 1986). Effects on the overall body weight are of course markedly influenced by the relative changes in fat and muscle, which are in turn influenced by the dose. Mass of visceral tissues and most organs is not increased; in some cases liver mass is decreased. Therefore, percent of live weight present in the carcass is usually increased. All farm animal species tested (including poultry, ruminants, and pigs) show similar but variable effects (see Table 2-5).
In mammals the magnitude of the response generally appears to be greater in ruminants than in single-stomached animals, although a functioning rumen does not appear to be required in calves (Williams et al., 1987) or lambs (Williams et al., 1989). The 20 to 40 percent increases in skeletal muscle mass commonly observed in growing lambs and cattle are rarely observed in swine (see reviews by Hanrahan et al., 1987; Beermann, 1989, 1993; Anderson et al., 1991; Moloney et al., 1990). Likewise, the 20 to 40 percent reductions in adipose tissue mass observed in lambs and cattle are approximately half as large in pigs.
The magnitude of the influence of these compounds on the adipose tissue content of the carcass appears to be related to the tendency of the control animals to lay down fat (i.e., the magnitude of carcass or empty-body lipid accretion rate). Responses are less significant in preweaning and young rapidly growing animals, in which lipid accretion rates are low. Likewise, the enhancement of skeletal muscle growth is also less in these younger animals. Jones et al. (1985) and Moser et al. (1986) studied the impact of cimaterol dose on pigs treated from approximately 60 to 105
TABLE 2-5 Representative Responses in Farm Animal Species to Dietary Administration of β-Adrenergic Agonists
kg BW. Effects on average daily gain and feed efficiency were small, but dressing percentage was increased and carcasses contained up to 10 percent less fat and 10 percent more skeletal muscle in proportion to cimaterol dose. More recent studies with another compound, ractopamine, have demonstrated 5 to 20 percent improvements in growth performance and dose-dependent improvements in carcass composition, including 8 to 20 percent more muscle mass (Adeola et al., 1990; Watkins et al., 1990; Bark et al., 1992) and 4 to 37 percent less adipose tissue (Watkins et al., 1990; Bark et al., 1992). Although cimaterol was effective in finishing pigs, it had no effect on growth performance or carcass composition in younger pigs fed similar doses from 10 to 60 kg BW (Mersmann et al., 1987); nor were there any effects on the several indices of lipid metabolism studied. Similar differential responses between young and older animals have been observed in ruminants. The dose-response effects of cimaterol (Quirke et al., 1988) and L-644,969 (Moloney et al., 1990) on growth performance and carcass composition in finishing cattle exceed the magnitude of response seen in veal calves (Williams et al., 1987). Effects on skeletal muscle growth of 10-day-old lambs (15 kg BW) fed milk replacer and cimaterol for 21 days (Williams et al., 1989) was minimal (10 to 15 percent), approximately half the reduction in lipid accretion rate as that observed with similar treatment in older ruminating lambs from the same genetic pool (O'Connor et al., 1991).
The lack of response in very young ruminants and nonruminants may be related to fewer receptors, lower binding affinity, or more rapid development of refractoriness to these compounds. Kim and Sainz (1990) have shown that the number of β-receptors in rat plantaris muscles decreased 28 to 42 percent after 3 days of dietary cimaterol administration, which preceded the attenuation of the muscle hypertrophy response over a 14-day treatment period.
There have been relatively few specific breed or genotype comparison studies reported for effects of β-agonists in farm livestock. Either little evidence of important genotype-by-treatment interactions or none at all has been reported
for lambs (Hanrahan et al., 1987) and pigs (Yen et al., 1990a,b; 1991). However, significant differences in responses among genotypes were observed in some comparisons (Warris et al., 1990; Gu et al., 1991a,b; Bark et al., 1992). Cimaterol and ractopamine increased skeletal muscle growth in both obese and lean genotypes of swine (Yen et al., 1990a,b; 1991), but anabolic responses to ractopamine were greater in genotypes that exhibited superior growth performance and carcass muscle and protein accretion rates than the inferior genotype to which they were compared (Bark et al., 1992; Gu et al., 1991b). Ractopamine-treated pigs of the inferior genotype still exhibited 12 kg less skeletal muscle mass than control pigs from the superior genotype when comparisons were made at similar body weights (Bark et al., 1992), which indicates that genotypic differences are not eliminated by β-agonist administration.
Responses to β-agonists in poultry tend to be, on a percentage basis, less significant than those seen in the mammals (see Table 2-5). This difference is probably caused by fundamental differences between adipose tissue metabolism and the type of β-receptor found in the muscle. There is, however, some evidence for the response in chickens to be related to the sex of the animal. Treatment of Hubbard × Hubbard broilers with cimaterol reduced carcass fat on the order of 10 percent in the female birds but only approximately 5 percent in the males (Dalrymple and Ingle, 1987). This sexual dimorphic effect may be caused by the tendency of female broilers to deposit more fat than their male counterparts. Cimaterol fed at 1 mg/kg to broiler chickens increased leg muscle weight more than breast muscle weight, and effects were greater after 56 days of treatment than after 38 days (Morgan et al., 1989). Cimaterol has also been shown to improve growth performance and body composition of ducks in a dose-dependent manner (W. F. Dean, Duck Research Laboratory, Eastport, Long Island, N.Y., personal communication, 1987). Ractopamine causes a dose-dependent improvement in growth performance, dressing percentage, and muscle content of turkeys when administered at the end of the feeding period (12 to 20 weeks) (Wellenreiter and Tonkinson, 1990a,b).
Mechanisms of β-Agonist Action
Although it is convenient to discuss en masse the β-agonists that seem to induce enhancement of skeletal muscle deposition and reduce carcass fat content, it should be remembered that marked differences are present in their structures, pharmacokinetics, and metabolism. Therefore, some differences in their mode of action and efficacy are to be expected.
The pharmacokinetic properties of β-adrenergic agonists will influence rates of absorption into the circulation, magnitude and temporal pattern of elevated concentrations in blood or plasma, and even selectivity for specific β-receptors (Timmerman, 1987). Variation among the compounds in how they are metabolized and eliminated from the circulation gives rise to estimated biological half-lives in rats ranging from 2 hours for fenoterol (Rominger and Pollmann, 1972) to 24 hours for clenbuterol (Kopitar and Zimmer, 1976). Species and mode of administration also contribute to variation among half-life estimates of the β-agonists. Published estimates of half-lives of β-agonists in farm animals are scarce. Affinity chromatography and high-performance liquid chromatography were used to describe the biphasic decline of plasma cimaterol concentration following a bolus intravenous injection in steers and yielded half-life estimates of 2.5 minutes for the distribution phase and 54 minutes for the elimination phase (Byrem et al., 1993). Biphasic elimination of clenbuterol in urine was demonstrated in veal calves fed 5 µg/kg BW twice daily for 3 weeks (Meyer and Rinke, 1991). Estimated half-life was 10 hours for the rapid phase and 2.5 days for the second phase. Half-life of clenbuterol in plasma could only be calculated for the fast phase and was estimated to be 18 hours. These estimates as well as those for other synthetic β-agonists developed for therapeutic use are much larger than for epinephrine and indicate that transfer to the peripheral compartment is very rapid. They also provide evidence that direct metabolic effects of cimaterol on specific tissues may be studied by close arterial infusion of cimaterol into specific vascular beds.
No formal proof exists for a common or shared set of specific actions of these compounds on skeletal muscle growth or lipid metabolism among all species in which they have been evaluated. The similar changes observed in protein and lipid deposition in growing animals suggest involvement of common effects. However, differences exist among the results of studies on mode of action. Review of the literature indicates that both quantitative and qualitative differences exist in the lipid metabolism response of different species to the same compound and of the same species to different compounds (Mersmann, 1989b). Similarly, attempts to block responses in muscle and adipose with receptor-specific compounds have given mixed results (Reeds et al., 1988; Choo et al., 1989). Therefore, caution must be exercised when drawing generalizations about mode of action of these compounds.
MODE OF ACTION IN SKELETAL MUSCLE
Treatment with β-agonists causes muscle hypertrophy rather than hyperplasia (Maltin et al., 1986; Beermann et al., 1987; Kim et al., 1987), but the response is not equal or not seen in all muscles (Beermann et al., 1986a; Bohorov et al., 1987; Dawson et al., 1988; Morgan et al., 1989). Responses in muscles containing a predominance of one fiber type (e.g., rat soleus) have been both greater than (Maltin et al., 1986) and equal to (Reeds et al., 1986; Thiel et al., 1987) responses
observed in mixed-fiber type muscles. It is apparent that type-II fibers (i.e., fast-contracting, mixed glycolytic-oxidative) account for the greater portion of hypertrophy when compared with the type-I fibers (slow contracting oxidative) (reviewed by Yang and McElligott, 1989). There is evidence that long-term treatment can cause an increase in the proportions of type-II fibers (Beermann et al., 1987; Zeman et al., 1988), although this has not always been observed (Kim et al., 1987).
Increases in muscle protein deposition (growth) can either be a result of changes in the rate of protein synthesis or in the rate of degradation or both. Several studies suggest that β-agonists reduce the rate of muscle protein degradation in sheep (Bohorov et al., 1987), rats (Reeds et al., 1986), cattle (Dawson et al., 1988), and broilers (Morgan et al., 1989). There are also data which suggest that muscle protein synthesis may be enhanced in rats (Emery et al., 1984), lambs (Claeys et al., 1989), cattle (S. B. Smith et al., 1989), and swine (Bergen et al., 1989; Helferich et al., 1990). Chronic feeding of clenbuterol increased the rate of β-amino nitrogen uptake by 44 percent in the hindquarters of steers (Eisemann et al., 1988). This was caused by chronic elevation of blood flow with no difference in arterio-venous concentration. Plasma urea nitrogen concentrations were reduced 20 percent with chronic cimaterol treatment. These data are consistent with an increase in protein synthesis and deposition and reduced amino acid oxidation with chronic administration of clenbuterol. Although oxygen utilization by the hindquarters was also increased, glucose uptake was not, indicating greater reliance on lipid oxidation to support the expected increase in energy required for protein synthesis and deposition.
Attempts to use isolated muscle preparations have also yielded equivocal results. For example, using two muscle cell lines, L6 and G8-1, Harper et al. (1990) obtained an increase in protein synthesis of about 12 percent following treatment with cimaterol, with the half-maximal effect occurring at a concentration consistent with the binding of cimaterol to the β-receptor on the cells. The effect was blocked by the antagonist propranolol. As mentioned earlier, Kim and Sainz (1990) demonstrated a temporal reduction of the number of β-receptors in rat muscle with cimaterol treatment, which preceded a diminished response in muscle hypertrophy during a 14-day treatment period. These data are taken as indicative of the involvement of the β-receptor. The presence of the β-receptor on L6 myoblasts (Pittman and Molinoff, 1983) and in muscle suggests that the agonists do have a direct effect on the muscle, especially in light of the finding that treatment of animals with propranolol (a β-antagonist) can block the myogenic action of β-agonists (MacLennan and Edwards, 1989).
Young et al. (1990) observed a 25 percent increase in the quantity of myofibrillar protein and a 30 percent increase in the quantity of myosin heavy chain in primary muscle cell cultures of broiler chicks with 10-7 M cimaterol. At higher levels of cimaterol the myosin heavy chain synthesis rate was increased 10 to 12 percent and protein degradation rate was decreased 10 to 15 percent. Clenbuterol (10-7) increased fusion rate and protein synthesis rate in neonatal rat myoblast cultures but failed to exhibit similar effects in rat satellite cell cultures or cultures of L6 myoblasts and myotubes and had no effect on neonatal fibroblast cultures (McMillan et al., 1992). McElligott et al. (1989) also observed no effect on protein metabolism of L6 cells treated with the agonist zinterol. It appears that origin and/or presence of the full complement of regulatory factor genes (present in animal-derived cells) may be important in responsiveness of myogenic cells to the synthetic β-agonists.
Taking an overview of the literature, it would appear that the rate of muscle protein synthesis is increased and the protein degradation rate may be reduced in animals fed these synthetic β-agonists. Temporal patterns of change are present that make it difficult to ascertain which might be the major route by which β-agonists increase the rate of muscle protein deposition. Measurement of calcium-dependent proteinase, calpastatin, and cathepsin activities in skeletal muscle of β-agonist-treated sheep (Higgins et al., 1988; Wang and Beermann, 1988; Beermann et al., 1989; Kretchmar et al., 1990), cattle (Wheeler and Koohmaraie, 1992), rabbits (Forsberg et al., 1989), and broiler chickens (Morgan et al., 1989) indicate that activities of calpastatin are increased and/or the microcalpain protease activity is reduced with cimaterol, L-644,969, and L-665,871 adrenergic agonist administration in vivo. The protein-sparing effects of β-agonist administration have been demonstrated in response to restricted energy intake and starvation. Cimaterol converted a daily loss of 2.3 g carcass protein to a daily gain of 4.1 g carcass protein in lambs maintained at zero energy balance (Kim et al., 1989). Starvation-induced skeletal muscle atrophy was significantly reduced when clenbuterol was given to rats (Choo et al., 1990). Indications that clenbuterol-induced reduction in basal nitrogen loss could be achieved in sheep (Hovell et al., 1989) have subsequently been shown in further studies to be transient, and nitrogen loss was equal in control and treated sheep after a 4-day treatment period (Inkster et al., 1989).
Indirect Effects of β-Adrenergic Agonists
Despite evidence for direct, receptor-mediated influences on skeletal muscle, it is possible that some of the changes in muscle protein metabolism in vivo are brought about by an indirect mechanism, that is, as a result of the changes in the circulating concentrations of some endogenous hormones (see review by Buttery and Dawson, 1987). Elevation of insulin concentrations have been observed in sheep (Beermann et al., 1986b; O'Connor et al., 1988) and cattle (M. Vestergaard, National Institute of Animal Science,
Tjele, Denmark, personal communication, 1990) with acute administration of cimaterol, but chronic treatment reduces insulin concentrations 25 to 50 percent (Beermann et al., 1987; O'Connor et al., 1988). Circulating levels of ST are not elevated with acute or chronic exposure of growing lambs to cimaterol (O'Connor et al., 1988), and no difference was observed in plasma concentrations of prolactin, cortisol, or thyroid hormones at 6 or 12 weeks. Similar results have been reported for growing steers fed cimaterol (Chikhou et al., 1991).
Cimaterol evokes similar stimulation of skeletal muscle growth and reduction in lipid accretion in animals surgically manipulated to remove the source of somatotrophic and metabolic hormone secretion. Cimaterol administration causes marked muscle hypertrophy in hypophysectomized rats (Thiel et al., 1987) and thyroidectomized rats (Forsberg and Wehr, 1990), and muscle hypertrophy is stimulated in severely diabetic rats and diabetic rats given a daily fixed dose of insulin (McElligott et al., 1987). These data suggest that ST, the thyroid hormones, and insulin, all important metabolic hormones required for normal muscle growth, are not involved to any great extent in the mediation of β-agonist-induced skeletal muscle hypertrophy. These results and the lack of significant changes in circulating metabolic hormone concentrations in response to β-agonist administration suggest that the β-agonist's mode of action involves direct, receptor-mediated stimulation of skeletal muscle growth.
MODE OF ACTION IN ADIPOSE TISSUE
It is generally accepted that β-agonists act directly on adipose tissue via the β-receptor to stimulate lipolysis. This is supported by the consistent observation of elevated plasma free fatty acids in treated animals (Beermann et al., 1987; Eisemann et al., 1988). Results from in vitro studies have not yielded such clear-cut results. For example, clenbuterol has been shown to stimulate lipolysis in adipose tissue from rats (Duquette and Muir, 1985) and chickens (Campbell and Scanes, 1985) but not pigs (Rule et al., 1987) or cattle (Miller et al., 1988; Dawson et al., 1989). However, isoproterenol, cimaterol, and ractopamine all stimulate lipolysis in pig adipose tissue in vitro (Liu et al., 1989; Peterla and Scanes, 1990). Several reports have shown that β-agonists can also affect the in vitro rate of lipogenesis (Mersmann, 1989a,b; Mills and Liu, 1990; Peterla and Scanes, 1990). In the absence of any effect on lipolysis, some reports have concluded that the reduction in lipogenesis is a very important component in the mechanism whereby total body fat is reduced (Miller et al., 1988). It is becoming clear that the incubation conditions used for these in vitro incubations is critical (Liu et al., 1989; Mersmann, 1989a,b; Mills and Liu, 1990). Liu and Mills (1990) have subsequently shown that clenbuterol and ractopamine reduce insulin binding to porcine adipocytes presumably through reduced insulin receptor number, thereby antagonizing insulin action on porcine adipocytes.
A major deterrent to conclusively identifying the mode of action is that it is difficult to measure rates of lipogenesis in vivo. It is also very likely that potencies differ among β-agonists, especially in terms of their relative effects on lipolysis and lipogenesis as well as the response between different species.
The ability of many β-agonists to induce a decrease in adipose tissue and at the same time an increase in skeletal muscle is a very useful attribute for animal production. However, there are agonists developed for other purposes that reduce adipose tissue without increasing lean mass, for example BRL 35135 (Arch and Ainsworth, 1983; Arch et al., 1984). It therefore seems that the increase in lean mass seen with many β-2-selective agonists is not simply a consequence of the reduced amount of energy stored in adipose tissue.
Interaction between Treatment and Dietary Intake
There have been relatively few reports in which the interaction between response to β-agonists and dietary protein and/or energy intake was investigated. Dry-matter intake is commonly reduced on initial exposure to the β-agonists but most often returns to normal within a short time and remains unchanged thereafter. The repartitioning effects of β-agonists are reported to occur in both adequate and restricted feeding conditions in lambs (Hovell et al., 1989; Kim et al., 1989) and pigs (Bracher-Jakob and Blum, 1990; Bracher-Jakob et al., 1990; Dunshea et al., 1991). However, significant increases in growth rate tend to occur only in well-fed animals. Even these effects can be lost if the animals become refractory to the compound or if the dose rate is increased (see, e.g., the data assembled by Williams, 1987; Beermann, 1993).
Providing adequate supplies of amino acids and energy is prerequisite to optimizing rate and efficiency of protein use for growth in normal management systems and may be particularly important when protein deposition rates are enhanced by β-agonist administration. Increased skeletal muscle protein deposition will increase the requirement for individual amino acids unless there is an increase in the efficiency with which dietary protein (amino acids) is used for growth. Inadequate protein intake constrains the magnitude of improvement in growth performance, nitrogen balance, or the degree to which protein accretion rate or skeletal muscle growth is enhanced by ractopamine in pigs (Anderson et al., 1987; Adeola et al., 1990; Dunshea et al., 1991; Mitchell et al., 1991). Anderson et al. (1987) observed that nitrogen retention in swine fed ractopamine is enhanced with an 18 percent crude protein diet, but nitrogen retention was reduced with a 12 percent protein diet.
Adeola et al. (1990) observed that ractopamine depressed ADG and gain: feed 9 and 10 percent, respectively, when the diet contained only 13 percent crude protein. However, ractopamine increased ADG and gain: feed 10 to 12 percent and 12 to 25 percent, respectively, when the diet contained 17 percent crude protein. These studies do not resolve the question of whether the efficiency of protein utilization is changed or whether protein requirement is increased.
If protein intake limits the rate of protein accretion, either the amount or the profile of amino acids supplied in the diet may account for the restriction of protein deposition. Therefore, in nonruminants protein intake titration experiments must be conducted using diets in which the amino acid profile is matched to the amino acid profile of deposited protein. These types of data are limited. Dunshea et al. (1991) demonstrated in gilts fed diets ranging from 8.3 to 23 percent crude protein concentration that carcass protein accretion rates were not increased by ractopamine at crude protein concentrations below 14 percent, and that additional crude protein was required (16.8 versus 14 percent) to accommodate the greater protein accretion rates achieved with 20 mg ractopamine/kg diet. No evidence was observed for ractopamine increasing the efficiency of protein use for growth. In ruminants, enhanced availability of amino acids does enhance skeletal muscle growth. Substitution of fishmeal for an equal amount of soy protein enhanced skeletal muscle mass in ram lambs by 15 to 19 percent, and effects were additive with cimaterol (Beermann et al., 1986a). Proximal hind limb muscles in lambs fed fishmeal and cimaterol were 40 to 45 percent larger than those in lambs that received no fishmeal or cimaterol.
An increase in skeletal muscle mass and in the basal metabolic rate of treated animals (see, e.g., MacRae et al., 1988; Kim et al., 1989) may increase the maintenance requirement of animals fed β-agonists. These increases would be offset only minimally by the reduction in fat deposition, but the shift of protein synthesis and accretion away from tissues with higher turnover rates (small intestine and liver) and toward skeletal muscle may help minimize changes in maintenance requirements. Although it is possible to speculate on the magnitude of any changes in nutrient response following treatment with β-agonists, it is doubtful that data are available to do this with any precision. In extrapolating from the data available for ST-treated animals, caution must be exercised because although both ST and β-agonists have similar effects on fat and skeletal muscle deposition, they have different effects on the relative growth of other tissues, especially the liver, kidneys, and other visceral organs. Different mechanisms of action could also dictate different effects or influences on nutrient requirements. This may result in differences in the response to changes in nutrient availability or to dietary manipulations.
Summary of Effects of β-Adrenergic Agonists on Growth and Composition
Chronic dietary administration of select β-adrenergic agonists markedly influences protein and lipid metabolism in farm animals, leading to marked increase in skeletal muscle protein accretion rate and, in most cases, significant reduction in lipid deposition rates. Significant improvement in carcass composition results without effect on growth of bone and with little effect on mass of visceral tissues and other organs. Improvements in growth performance appear to be greatest within the first few weeks of administration and diminish to a varying extent with continued administration. Ruminants appear to be more responsive than swine, and poultry respond least. Significant influences of diet and genotype on magnitude of response have been observed, but the nature of these interactions varies across species.
ANABOLIC STEROIDS
Naturally occurring and synthetic estrogens and androgens have been safely used to improve efficiency of growth and carcass composition of meat animals for more than 40 years. Historically, the first commercial use of an estrogen was in poultry, but this lasted only a short time. Anabolic steroids are not approved for use in growing swine in the United States; however, both estrogens and androgens are extensively used in growing cattle produced for beef. Several anabolic steroid implants are currently approved for use in beef cattle in the United States. Only one compound, zeranol, is approved for use in lambs. These approved steroid implants include the naturally occurring hormone, estradiol, the hormone progesterone in combination with estradiol or estradiol benzoate, the fungal metabolite with estrogenic properties, zeranol, the synthetic progestin, melengestrol acetate (MGA), testosterone propionate in combination with estradiol benzoate, and a synthetic testosterone analog, trenbolone acetate (TBA). Structures of estradiol, progesterone, zeranol, testosterone, TBA, and MGA are shown in Figure 2-4.
Classification of the anabolic agents previously or currently in use is based on their chemical structures and associated actions. A review of the biosynthesis and metabolism of the naturally occurring estrogens and androgens has recently been published (Hancock et al., 1991). Descriptions, approval dates, and recommended doses of the commercial products are found in papers by Schanbacher (1984), Muir (1985), and Hancock et al. (1991). Efficacy of these anabolic steroid implants is summarized in several reviews (Galbraith and Topps, 1981; Schanbacher, 1984; Muir, 1985; Roche and Quirke, 1986; Beermann, 1989; Hancock et al., 1991).
The literature on growth-performance responses to anabolic steroids indicates great variability, ranging from no
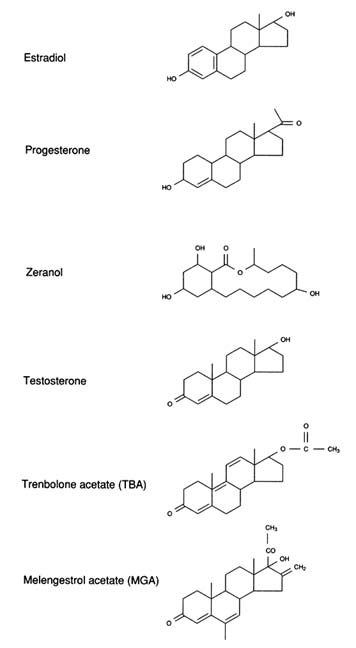
FIGURE 2-4
Chemical structures of the endogenous steroids estradiol, progesterone, and testosterone and of synthetic anabolic steroids.
response in feedlot bulls (Richards et al., 1986) to a 69.9 percent increase in average daily gain in heifers treated with TBA (Bouffault and Willemart, 1983). Gender determines which anabolic steroid will be administered. The estrogenic compounds are generally more effective in steers. The response in females is more variable and less consistent, but the androgenic steroids are superior. Use of a combination of anabolics generally produces an additive response compared to use of either estrogenic or androgenic implant alone. Response in bulls is generally less than that of steers, and implanted steers often achieve the growth performance observed in nonimplanted bulls (Fisher et al., 1986).
Growth Performance Response to Anabolic Steroids
Rate of live-weight gain is increased 10 to 20 percent on average by anabolic steroids (Bradley et al., 1957; Sharp and Dyer, 1971; Griffiths, 1982; O'Lamhna and Roche, 1984; Gill et al., 1987; Keane and Drennan, 1987; Perry et al., 1991), but responses approaching 50 percent have been observed in lambs (Sulieman et al., 1988), steers in calorimetry chambers (Lobley et al., 1985), and steers compensating for earlier growth restriction (Keane and Drennan, 1987). It appears that young animals may respond better to steroid implants than older animals (Mader et al., 1985; Whittington, 1986; Simms et al., 1988), and this may be particularly true in bulls (Richards et al., 1986). In some studies it was observed that the rate of mass increase was higher during the early period following implantation and then decreased, which may reflect the declining circulating concentration of the anabolics after the first few weeks (Schanbacher, 1984; Lobley et al., 1985; MacVinish and Galbraith, 1988; Hayden et al., 1992).
Results from TBA-estradiol combinations appear to be superior to either implant alone. Dose-response efficacy trials involving 1,296 steers conducted for FDA approval of TBa-estradiol combinations indicate that the ADG plateaued at 118 mg TBA combined with 24 mg estradiol, but the feed efficiency plateaued at 139 mg TBA combined with 28 mg estradiol (Bartle et al., 1992). ADG was increased 18 percent and the feed: gain ratio was reduced 9.5 percent, both of which exceeded the response to 30 mg estradiol alone. Implants of 140 mg TBA alone did not improve growth performance in this and other studies (Apple et al., 1991; Hayden et al., 1992). The authors further indicated that a TBA-estradiol ratio of 5:1 is optimum for feedlot steers fed a high-grain diet. Feed conversion efficiency is usually improved with anabolic steroids, but the magnitude of the response is variable. Improvements of 5 to 14 percent have been reported (Rumsey, 1978; Greathouse et al., 1983; Mathison and Stobbs, 1983; Steen, 1985; Thonney, 1987). TBA-estradiol combinations have been shown to decrease feed: gain ratios by 10 to 13 percent (Trenkle, 1987; Eversole et al., 1989; Perry et al., 1991; Bartle et al., 1992). The degree of response to anabolic steroids is clearly influenced by dry-matter intake changes, which are also variable, and by the degree of change in composition of gain. The majority of studies in which large increases in gain were observed also showed 5 to 10 percent increases in feed intake (Thonney, 1987; Perry et al., 1991; Bartle et al., 1992) and proportional increases in lean mass in cattle (Keane and Drennan, 1987) and lambs (Sulieman et al., 1986, 1988). However, no significant changes in feed intake were observed in several studies (Griffiths,
1982; Vanderwert et al., 1985; Calkins et al., 1986; Fisher et al., 1986).
Composition of Gain
Few studies investigating the effects of anabolic steroids on growth in ruminants in the United States include direct measurement of carcass or empty-body composition, a necessity for understanding the mode of action and defining nutrient requirements. Total lean carcass (muscle) increased 9.5 and 10.4 percent in steers implanted twice with 300 mg TBA/36 mg resorcylic acid lactone over the live-weight range of 250 to 400 kg (Griffiths, 1982). Separable fat was reduced 2 percentage points (P < 0.05) and efficiency of gain was greater in implanted cattle fed at the higher of two levels of energy intake. Dressing percentage was higher in implanted cattle, which implies that neither organ weights nor gut fill were increased with treatment. Nitrogen balance was increased 31 and 70 percent, respectively, in two subsets of animals used in metabolism trials in this study. Nitrogen intake was not different between implanted and nonimplanted cattle, and urinary nitrogen excretion was reduced 25 and 29 percent, respectively.
Nitrogen balance of Holstein bull calves indicate that TBA-estradiol combinations increase cumulative nitrogen retention 47 percent compared with 28 percent increase in controls treated with estradiol alone (VanderWal et al., 1975). When administered separately, zeranol, progesterone, testosterone, and TBA also did not lead to a significant increase in nitrogen retention. Galbraith (1980) observed a 106 percent increase in daily nitrogen retention in beef heifers treated with a 300-mg implant of TBA for 60 days.
Long-term administration of TBA and resorcylic acid lactone to heifers and steers fed to 491, 612, or 731 days of age exhibited greater absolute and relative amounts of carcass lean and lesser absolute and relative amounts of carcass fat than nonimplanted cattle (Keane and Drennan, 1987). Implanted cattle had 23.6 kg heavier side weights, 23.1 kg more lean, 3.2 kg more bone, and 2.6 kg less fat than nonimplanted cattle. Sex-by-implant interactions were not significant. Cattle implanted the longest exhibited a greater (P < 0.05) increase in carcass lean than cattle implanted for shorter periods. The increase in carcass weight was accounted for entirely by the increase in carcass lean observed in implanted heifers and steers, and the decrease in fat accretion was offset by increased bone growth. Dressing percentage was increased in the implanted cattle.
Choice of the experimental end point for cattle growth trials can influence the outcome and interpretation of results. Compositional differences are influenced by live weight, as dictated by normal allometric growth patterns in farm animal species. However, industry grading and pricing systems may dictate what end point is most appropriate in a commercial production system. Such is the case for fed cattle for which the degree of marbling in the longissimus muscle measured at the twelfth rib determines quality grade. Few studies on the effects of anabolic steroids in fed cattle have been conducted with degree of marbling as the end point. Growth performance and composition of gain responses to TBA-estradiol implants were compared in three breeds of steers representing different frame sizes—Holstein, Angus, and Simmental-crossbred—using rib dissection and the comparative slaughter technique at a common degree of marbling end point (Perry et al., 1991). Daily empty-body protein gain was increased 25 to 27 percent in all three breed groups with little effect on carcass composition, carcass quality grade, or retail cut distribution. However, live weight required to reach the common degree of marbling end point was increased 25 to 45 kg with the TBA-estradiol implant. Fat gain was increased by 19 percent on average in these cattle. These results suggest that anabolic steroids stimulate growth without dramatic effects on composition of gain and increase the weight at which a common carcass intramuscular fat concentration is achieved.
Studies using combined implants in growing and finishing lambs confirm the results observed in cattle. Lambs fed diets containing 16 percent crude protein ad libitum and implanted with TBA and estradiol-17β exhibited significant increases in intake (14 percent), live-weight gain (16 percent), and carcass lean mass (8 percent), as well as significant reductions in subcutaneous and total carcass fat (Sulieman et al., 1986). A subsequent study with similar lambs at two initial weights (24 and 37 kg) demonstrated even more substantial growth performance and carcass composition effects that were equivalent in both weight groups (Sulieman et al., 1988).
The consistent improvement in rate of protein deposition observed in growing ruminants indicates that anabolic steroid implants exert their primary influence through altering protein metabolism, with lesser effects on lipid metabolism. They appear to increase feed intake in most cases.
Mechanism of Anabolic Steroid Action
Few data are available that describe the effects of anabolic steroids on protein metabolism, but even fewer data exist for assessment of direct effects of anabolic steroids on lipid metabolism in growing ruminants. Protein metabolism studies suggest that both fractional protein synthesis rate and protein degradation rate might be reduced by TBA, with the degradation rate reduced to a greater extent, resulting in an overall increased protein accretion rate in rats (Vernon and Buttery, 1976, 1978a) and lambs (Vernon and Buttery, 1978b; Sinnett-Smith et al., 1983). Combined TBA-estradiol (140 and 20 mg, respectively) implant treatment increased daily live-weight gain 50 to 60 percent at similar feed intakes and increased daily nitrogen retention 100 and 146 percent in Hereford-Holstein steers during the
first 7 weeks of treatment in a study by Lobley et al. (1985). Estimates of whole-body protein synthesis rate (based on metabolic body size) were similar throughout the 10-week experiment, while amino acid oxidation was lower in treated steers at 2 and 5 weeks. Urinary 3-methyl histidine excretion was slightly less and total energy retention was unaffected in treated steers, indicating that reduction of protein degradation rate may account for most of the improvement in daily gain and nitrogen retention. Heat production was not increased in steers treated with the TBA-estradiol combination.
A recent experiment indicates that one possible mechanism responsible for TBA's ability to stimulate skeletal muscle hypertrophy may be through enhanced proliferation and differentiation of satellite cells as the result of increased sensitivity to IGF-I and fibroblast growth factor (Thompson et al., 1989).
Few data are available regarding effects of anabolic steroid implants on lipid metabolism in growing ruminants. Prior et al. (1983) demonstrated that lipogenic enzyme activity and fatty acid synthesis in vitro were elevated in subcutaneous adipose tissue from bulls implanted with estradiol. This may account for the increase in fat content of carcasses reported in some studies. St. John et al. (1987) found that TBA implants had no effect on lipogenesis in intact heifers and only tended to reduce lipogenic enzyme activities in ovariectomized heifers treated with TBA.
Assessment of indirect effects is difficult because few studies have reported determinations of circulating hormone or metabolite concentrations in animals treated with anabolic steroid implants. Treatment of lambs (MacVinish and Galbraith, 1988) or cattle (Henricks et al., 1988) with TBA and estradiol caused reduced plasma urea nitrogen concentrations, which would be expected with decreased rates of amino acid oxidation; but a causative effect has not been demonstrated. Galbraith (1980) observed no significant changes in blood hormone or metabolite levels in heifers treated with 300 mg TBA for 60 days. The review by Buttery and Sinnett-Smith (1984) notes the lack of any consistent change in ST, prolactin, insulin, or other metabolic hormones in a total of 15 studies summarized. Recent data obtained from a study in which TBA alone, estradiol alone, and TBA plus estradiol treatments were compared in yearling steers (Hayden et al., 1992) supports this conclusion.
Summary of Effects of Anabolic Steroid on Growth and Composition
The consistent net effect of anabolic steroid implant use in growing ruminants appears to be that of increasing the live weight at which carcass or empty-body fat content or concentration equals that of nonimplanted animals, thus increasing their potential mature size. Increased growth rate is usually accompanied by an increase in feed intake. Data suggest that protein metabolism is altered toward increased rates of protein synthesis and reduced rates of amino acid oxidation and/or lesser rates of protein degradation. There may be little direct effect on lipid metabolism and associated fat deposition. Indeed, as is noted by the lack of information in the literature, little is known about direct effects of anabolic steroids and combinations of same on lipid metabolism in growing ruminants.


















