
APPENDIX E
Evidence for Mixing Based on Parasites
Walters (1980)1 examined the prevalence of two species of parasites from three locales (western Atlantic Ocean, Mediterranean Sea, and the Bay of Biscay). Based on the prevalence of the parasites, he argued that annual mixing of the two stocks was about 15%. He concluded that some school tuna in the Bay of Biscay "may" have been spawned in the Gulf of Mexico and recommended further study to support the postulate, however, to date, this has not been carried out.
Of all the techniques used to argue the case for mixing, this is by far the simplest. It requires a sample of 20 to 30 fish, ages 0 to 2, from each locale. The sample can consist of only the heads of the fish, and they can be preserved in 10% formalin.
Little is known about the life cycle of these parasites or how they are acquired. The life cycle is direct: eggs are shed into the water, and immatures are free-living. Immatures in the water column usually attach to the skin or gill cavity of the host in response to unknown chemical cues, and they migrate to the preferred site (the nasal cavity in the case of Nasicola sp.; the gill chamber in the case of Elytrophora sp.). Generally speaking, adult parasites of this type do not pass from one host to another. There is no information on the longevity of these parasites.
The analyses by others of Walters' data are not critical reviews. Murphy (1990) confuses prevalence and intensity and simply restates Walters' postulate
that there is mixing of about 15% in both directions. Brunenmeister's (1980) analysis predates access to all of the Walters' data and so is incomplete. Bossert (1993) does not consider Walters' data. It is important then to restate Walters' view of his own data. He uses the terms "reasonable to postulate," "may have spawned," "a plausible explanation,'' "further study is indicated," "it appears that Nasicola sp. could be used," and "Elytrophora sp. appears to be acquired." Walters offers a possible explanation, but it is not more than that and always should be couched in those terms.
NASICOLA SP.
Most giant bluefin tuna in the western Atlantic Ocean are infested with Nasicola sp. (93% [Walters, 1980], 81% [Phipps, 1980]). None have been sampled from the eastern Atlantic Ocean.
The most definitive result in Walter's study is that mean parasite size increases with host size. Data of Phipps (1980) and Wheeler and Beverley-Burton (1987) confirm and extend this relation (Figure E-1). Walters suggested that the parasite may be very long lived because parasite size increases with host size and the host is long lived (20+ years). An alternative explanation, not uncommon in the parasitological literature, is that the parasites grow larger because they have more room to do so and that the giant fish repeatedly acquire the parasite each time they pass through water harboring immatures.
Table E-1 and Figure E-2A provide a summary of the data for Nasicola sp. The data of Phipps (i.e., worm size [1980]) suggest that the giant bluefin tuna used by Walters were probably small giant fish. Neither Walters nor Wheeler and Beverley-Burton (1987) give the host size of giant fish.
Walters used the data to suggest the following. The decrease in prevalence in the western Atlantic tuna (i.e., 0.86 at age 0 to 0.31 at age 3) was taken to indicate that the fish were being diluted by uninfested fish from the east; there is mixing. The presence of infested fish in the Bay of Biscay was taken to indicate that the uninfested fish from the Mediterranean Sea were being diluted by infested fish from the west; there is mixing.
The Nasicola klawei holotype (i.e., the specimens used to define the parasite species) was obtained from tuna other than bluefin (Yamaguti, 1963). Wheeler and Beverley-Burton (1987) proposed a new species (N. hogansi) for specimens obtained from western Atlantic bluefin tuna (Prince Edward Island, Canada). Their description is identical to that for a much larger sample described by Phipps (1980) also from western Atlantic giant bluefin tuna. It is likely the taxonomy is confused because workers have not sampled parasites from hosts of the same size. It is likely that the species from bluefin tuna described by Walters is not N. klawei but rather N. hogansi.
The high prevalence at age 4 was ignored by Walters; presumably it was taken to be anomalous. The slope of the line describing the decrease in prevalence
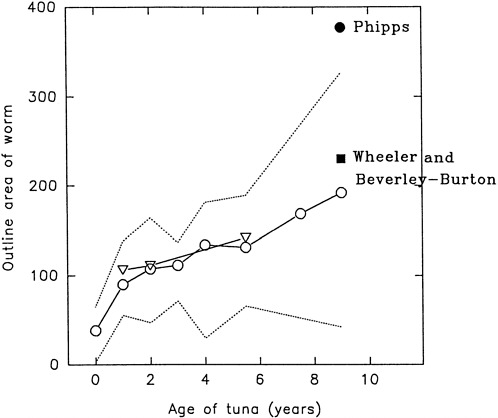
FIGURE E-1 The increase in size of Nasicola sp. with host size in Atlantic bluefin tuna. Age 9 on the x-axis actually refers to all fish 9 years or older (i.e., giant fish). Open circles are mean values for western bluefin tuna, and dashed lines indicate the range; open triangles are mean values for eastern bluefin tuna (Walters, 1980). Filled square (Wheeler and Beverley-Burton, 1987; tended line in Prince Edward Island) and filled circle (Phipps, 1980; fork length of 280 cm, traps in Nova Scotia) are mean values for western bluefin tuna.
from age 0 to age 8 is not significantly different from zero, thus weakening the dilution argument. The number of fish sampled from the east is very small. It is likely that the marked increase in prevalence between ages 8 and 9+ is related to a marked change in migration patterns. That is, at age 9 the giant fish return to warm water to spawn, but when they return to warm water they also acquire the parasite. An alternative and equally valid explanation of all of the data is that there are two regions where the parasite can be acquired, one in the west (in the region of the Florida Straits) and one in the east (near the Bay of Biscay). The prevalence data are some measure of the number of tuna passing through these regions at some time of the year, and the probability of a tuna
TABLE E-1 Prevalence of Nasicola sp. parasites in bluefin tuna related to age of the host and to locale. The fractions are the number of hosts infested divided by the number of hosts examined.
|
Age (years) |
Western Atlantic Ocean |
Mediterranean Sea |
Bay of Biscay |
|
0 |
19/22 = 0.86 |
0/9 = 0 |
— |
|
1 |
19/25 = 0.76 |
0/2 = 0 |
2/14 = 0.14 |
|
2 |
38/62 = 0.61 |
— |
2/24 = 0.08 |
|
3 |
8/26 = 0.31 |
0/1 = 0 |
— |
|
4 |
19/24 = 0.79 |
0/3 = 0 |
0/3 = 0 |
|
5 & 6 |
3/17 = 0.18 |
0/1 = 0 |
1/12 = 0.08 |
|
7 & 8 |
9/25 = 0.36 |
— |
— |
|
9+ |
53/57 = 0.93 |
— |
— |
|
9+ Phipps |
39/48 = 0.81 |
— |
— |
passing through the waters where they can acquire the parasite changes with their age.
ELYTHROPHORA SP.
Figure E-2B and Table E-2 provide a summary of the data for the copepod.
Walters used these data to suggest that the western fish ages 2 to 5 infested with this parasite represent fish that moved from the east to the west.
The taxonomy on this parasite is also confused, but a revision has not been published (Phipps, 1980).
About 20% of young western tuna were infested with the parasite, and fish that were parasitized had about five parasites each. Walters argued that this is a nonrandom distribution (i.e., of 100 fish one would expect each to have one parasite rather than 20 fish each to have five parasites). He suggested that the infested and noninfested fish had different histories and that the infested fish in the west had their origin in the east. It is equally likely that the two groups had different histories but that the infested fish passed through waters harboring the parasite and others avoided these waters. It is possible that the waters harboring the parasite are in the west and not in the east as argued by Walters.
Except for age 0, sample sizes from the Mediterranean Sea are too small to allow one to draw any inferences.
Age 4 fish from the western Atlantic Ocean are not treated as an anomaly, whereas in the Nasicola sp. analysis they were. As with Nasicola sp., it is likely that the marked increase in prevalence in age 9+ is related to a marked change in migration patterns. That is, at age 9 the giant fish return to warm water to spawn but in doing so also pass through waters that allow them to acquire the parasite.
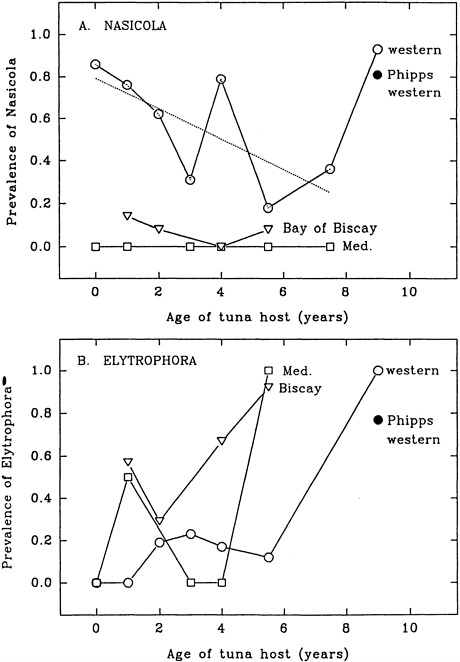
FIGURE E-2 Prevalence of parasites as a function of host age and locale of capture (A, Nasicola sp.; B, Elytrophora sp.). Age 9 on the x-axis actually refers to all fish 9 years or older (i.e., giant fish). Open circles are mean values for western tuna, and the dashed line is a linear regression for ages 0 to 8; open triangles are mean values for eastern bluefin tuna from the Bay of Biscay; open squares are mean values for eastern bluefin tuna from the Mediterranean Sea (Walters, 1980). Filled circles (Phipps, 1980; fork length of 280 cm, traps in Nova Scotia) are mean values for western bluefin tuna.
TABLE E-2 Prevalence of Elytrophora sp. parasites in bluefin tuna related to age of the host and to the locale of host capture. The fractions are the number of hosts infested divided by the number of hosts examined.
|
Age (years) |
Western Atlantic Ocean |
Mediterranean Sea |
Bay of Biscay |
|
0 |
0/22 = 0 |
0/9 = 0 |
— |
|
1 |
0/25 = 0 |
1/2 = 0.5 |
8/14 = 0.57 |
|
2 |
12/62 = 0.19 |
— |
7/24 = 0.29 |
|
3 |
6/26 = 0.23 |
0/1 = 0 |
— |
|
4 |
4/24 = 0.17 |
0/3 = 0 |
2/3 = 0.67 |
|
5 & 6 |
2/17 = 0.12 |
1/1 = 1.0 |
11/12 = 0.92 |
|
7 & 8 |
— |
— |
— |
|
9+ |
22/22 = 1.0 |
— |
— |
|
9+ Phipps |
53/69 = 0.77 |
— |
— |
An alternative and equally valid explanation of all of the data is that there are two regions where the parasite can be acquired, one in the west (in the region of the Florida Straits) and one in the east (near the Bay of Biscay). The prevalence data are some measure of the number of tuna passing through these regions at some time of the year, and the probability of a tuna passing through the area where they can acquire the parasite changes with their age.
HOSTS WITH BOTH PARASITES
Walters reported that only three of 129 western hosts ages 2 to 6 were infested with both parasites. Table D-3 is a summary of his data. Walters incorrectly reported the sum of those infested with Nasicola sp. as 52; the sum is 68.
Walters argued that if the parasites were randomly distributed in a homogeneous
TABLE E-3 Prevalence of Nasicola sp., Elytrophora sp., and both parasites in the same host for western Atlantic bluefin tuna ages 2 to 6.
|
Age (years) |
Nasicola sp. |
Elytrophora sp. |
Both parasites |
|
2 |
38/62 |
12/62 |
Not reported |
|
3 |
8/26 |
6/26 |
Not reported |
|
4 |
19/24 |
4/24 |
Not reported |
|
5 & 6 |
3/17 |
2/17 |
Not reported |
|
Sum |
68/129 = 0.53 |
24/129 = 0.19 |
3/129 = 0.02 |
host stock, one would expect 68 × 24/129 = 13 fish to harbor both parasites, whereas he observed only three. Walters draws the inference that the host stock is not homogeneous. It is equally likely that the parasites simply are not distributed randomly.
SUMMARY
One satisfactory explanation for the data is that there is mixing of young tuna: about 15% of tuna moving from east to west and a similar number moving in the other direction.
An equally valid explanation is that western bluefin tuna acquire both parasites in the relatively warm waters in the west, that eastern bluefin tuna can acquire both parasites in the relatively warm waters of the east, and that the water temperature that permits acquisition is different for the two parasites. The Walters study does not provide evidence to support mixing.
REFERENCES
Bossert,2 1993. Review of the ICCAT bluefin tuna working group report, draft of 10/2/93, and supporting documents.
Brunenmeister, S. 1980. A summary and discussion of technical information pertaining to the geographical discreteness of Atlantic bluefin tuna resources. ICCAT Coll. Vol. Sci. Pap. IX:506-527 (SCRS/79/95).
Murphy, G.I. 1990. A review of the evidence of stock structure in Atlantic bluefin tuna with an alternate stock hypothesis. Draft.
Phipps, M. 1980. Preliminary studies of population structure and mortality of bluefin tuna (Thunnus thynnus) in Nova Scotia, Canada. MSc thesis, University of Guelph.
Walters, V. 1980. Ectoparasites of eastern and western Atlantic bluefin tuna. ICCAT Coll. Vol. Sci. Pap. IX(2): 491-498 (SCRS/79/79).
Wheeler, T.A., and M. Beverley-Burton. 1987. Nasicola hogansi n.sp. (Monogenea: Capsalidae) from bluefin tuna, Thunnus thynnus (Osteichthyes: Scombridae), in the northwest Atlantic. Can. J. Zool. 65:1947-1950.
Yamaguti, S. 1963. Monogenetic Trematodes of Hawaiian Fishes. University of Hawaii Press, Honolulu. 287pp.








