
4
Proterozoic and Early Cambrian Protists: Evidence for Accelerating Evolutionary Tempo
Andrew H. Knoll
In the 50 years since G. G. Simpson published Tempo and Mode in Evolution , paleontological documentation of evolutionary history has improved substantially. Not only has the quality of stratigraphic and systematic data increased for animal, plant, and protistan taxa found in Phanerozoic * rocks; recent decades have witnessed a tremendous increase in the documented length of the fossil record. Speculation about a long pre-Cambrian history of life has been replaced by a palpable record of evolution that begins some 3000 Ma before the Cambrian explosion. In this paper, I examine the early fossil record of eukaryotic organisms, asking whether or not this longer record is amenable to the types of investigation used to estimate tempo in Phanerozoic evolution. Even though analysis is limited by incomplete sampling, patchy radiometric calibration, and taxonomic uncertainty, a robust pattern of increasing diversity and accelerating evolutionary tempo is evident.
Andrew H. Knoll is professor and chairman of the Department of Organismic and Evolutionary Biology at Harvard University, Cambridge, Massachusetts.
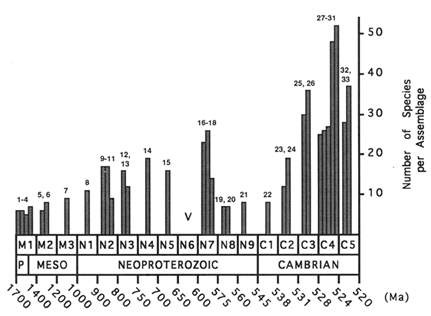
Figure 1 Species richness of selected protistan microfossil assemblages in 17 stratigraphic intervals running from the latest Paleoproterozoic era (P), through the Mesoproterozoic (Meso), Neoproterozoic, and Early Cambrian. Numbers 1–33 identifying assemblages refer to Table 1. V marks the Varanger ice age. Species richnesses are based on my taxonomic evaluation and do not necessarily reflect published tabulations. With a single exception (assemblage 31), all assemblages have been examined first-hand, resulting in uniform systematic treatment. Stratigraphic and systematic data for all figures and tables are available from the author.
The Nature and Limitations of the Record
Stratigraphic and Geochronometric Framework. The time interval considered here is 1700–520 Ma; that is, the latest Paleoproterozoic Eon to the end of the Early Cambrian Period (Figure 1 and Table 1). U-Pb dates on accessory minerals in volcanic rocks of known relationship to fossiliferous strata are limited for this interval—but then, such data are also limited for younger Paleozoic fossils on which much greater paleobiological demands are placed. Quantitative analysis of the Paleozoic fossil record is possible because a well-defined chronostratigraphic, or relative, time-scale has been calibrated by radiometric data in a few key sections.
The Proterozoic-Cambrian time scale is developing along the same
TABLE 1 Stratigraphic intervals used in analyses of tempo and representative acritarch assemblages
|
Interval (age in millions of years [Ma]) |
|||
|
Formation |
|
Location |
Reference |
|
Late Paleoproterozoic- and Mesoproterozoic |
|||
|
M1 |
(1700–1400 Ma) |
|
|
|
Satka [1] |
|
Urals, Russia |
Jank. 82 |
|
Bakal [2] |
|
Urals, Russia |
Jank. 82 |
|
Ust'-Il'ya [3] |
|
Siberia |
Veis. 92 |
|
McMinn [4] |
|
Australia |
Peat. 78 |
|
M2 |
(1400–1200 Ma) |
|
|
|
Omachtin [5] |
|
Siberia |
Vesi. 89 |
|
Zigazino-Kamarovsk [6] |
|
Urals, Russia |
Jank. 82 |
|
M3 |
(1200–1000 Ma) |
|
|
|
Baicaoping [7] |
|
China |
Yan. 92 |
|
Neoproterozoic |
|
|
|
|
N1 |
(1000–900 Ma) |
|
|
|
Lakhanda [8] |
|
Siberia |
Germ. 90 |
|
N2 |
(900–800 Ma) |
|
|
|
Miroyedikha [9] |
|
Siberia |
Germ. 90 |
|
Kwagunt [10] |
|
Arizona, USA |
Vida. 85 |
|
Dakkovarre [11] |
|
Norway |
Vida. 81 |
|
N3 |
(800–750 Ma) |
|
|
|
Andersby [12] |
|
Norway |
Vida. 81 |
|
Middle Visingsö [13] |
|
Sweden |
Vida. 76 |
|
N4 |
(750–700 Ma) |
|
|
|
Svanbergfjellet [14] |
|
Svalbard |
Butt. 94 |
|
N5 |
(700–650 Ma) |
|
|
|
Upper Visingsö [15] |
|
Sweden |
Vida. 76 |
|
N6 |
(650–600 Ma)* |
|
|
|
N7 |
(600–575 Ma; Volhyn) |
|
|
|
Pertatataka [16] |
|
Australia |
Zang. 92 |
|
Doushantuo [17] |
|
China |
Yin. 87 |
|
Kursovsky [18] |
|
Siberia |
Mocz. 93 |
|
N8 |
(575–560 Ma; Redkino) |
|
|
|
Redkino [19] |
|
Baltic |
Volk. 90 |
|
Mogilev/Nagoryany [20] |
|
Ukraine |
Asee. 83 |
|
N9 |
(560–545 Ma; Kotlin) |
|
|
|
Kotlin [21] |
|
Baltic |
Volk. 90 |
|
Early Cambrian |
|
|
|
|
C1 |
(545–538 Ma; Rovno) |
|
|
|
Rovno [22] |
|
Baltic |
Volk. 83 |
|
C2 |
(538–531 Ma; Lontova) |
|
|
|
Lontova [23] |
|
Baltic |
Volk. 83 |
|
Mazowsze [24] |
|
Poland |
Mocz. 91 |
|
C3 |
(531–528 Ma; Talsy) |
|
|
|
Talsy [25] |
|
Baltic |
Volk. 83 |
|
Lower Radzyn*/Kaplanosy [26] |
|
Poland |
Mocz. 91 |
|
Interval (age in Ma) |
|||
|
Formation |
Location |
Reference |
|
|
Early Cambrian (continued) |
|||
|
C4 |
(528–524 Ma; Vergale) |
|
|
|
Middle Radzyn*/Kaplanosy [27] |
|
Poland |
Mocz. 91 |
|
Qianzhisi [28] |
|
China |
Zang. 90 |
|
Tokammane [29] |
|
Svalbard |
Knol. 87 |
|
Vergale [30] |
|
Baltic |
Volk. 83 |
|
Buen [31] |
|
Greenland |
Vida. 93 |
|
C5 |
(524–520 Ma; Rausve) |
|
|
|
Upper Radzyn*/Kaplanosy [32] |
|
Poland |
Mocz. 91 |
|
Rausve [33] |
|
Estonia |
Volk. 83 |
|
Assembly numbers in brackets refer to Figure 1. * Interval includes Varanger ice age. |
|||
path (Harland et al., 1990; Semikhatov, 1991; Knoll and Walter, 1992; Compston et al., 1992; Bowring et al., 1993). A biostratigraphic framework based on stromatolites, microfossils, and (in younger rocks) both the body and trace fossils of animals can be used to divide this nearly 1200-Ma expanse into recognizable intervals of various lengths. Complementing this is an increasingly well-supported chemostratigraphic framework based on the distinctive pattern of secular variation in the isotopic compositions of C and Sr in carbonate rocks (Kaufman and Knoll, in press). These data define the chronostratigraphic scale now being calibrated. Within the period under consideration, younger intervals are shorter than older ones, both because strong Neoproterozoic isotopic variation has no parallel in the Mesoproterozoic record and, more importantly, because of the finer biostratigraphic resolution in younger successions.
For the purposes of this analysis, I have divided the period from 1700 to 520 Ma into 17 intervals as shown in Table 1 and Figures 1–3. Table 1 and Figure 1 also show my placement of representative microfossil assemblages into these intervals. Others might estimate the ages of interval boundaries differently, and one or two assemblages might be moved to bins adjacent to those chosen here. However, no assemblage placement or estimate of interval duration is so egregiously uncertain as to affect the analysis in a substantial way. That is, relative to the strength and time scale of the pattern observed, uncertainties of time are acceptably small.
The Paleontological Data Base: Taxonomy. For the estimation of evolutionary tempo, I will restrict consideration to the organic-walled micro-
fossils known as acritarchs (Figure 4). Structural features leave little doubt that all or nearly all were eukaryotic. Most were the vegetative and reproductive walls of unicellular protists, although the reproductive cysts of multicellular algae and even egg cases of early animals may be included.
The total number of clades that contributed to the observed record is unknown, but probably small. Some of the Early Cambrian microfossils included here are clearly the phycomata of green algal flagellates (Tappan, 1980). (The phycoma is a nonmotile vegetative stage of the flagellates' life cycle characterized by a wall that contains the degrada-
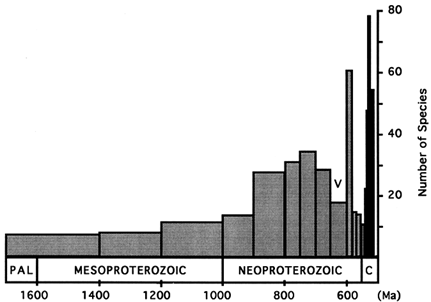
Figure 3 Histogram showing total species richness of protistan microfossils for the 17 intervals recognized in this study (Tables 1 and 2). The time scale along the abscissa is linear, underscoring the long initial interval of low diversity and the rapidity of later radiations. V marks the Varanger ice age. PAL, Paleoproterozoic; C; Cambrian.
tion-resistant polymer, sporopollenin.) Others, including most Neoproterozoic taxa, may also represent green algae (Tappan, 1980; Vidal and Knoll, 1983; Knoll et al., 1991), but phylogenetic relationships have not been established unequivocally.
The pre-Ediacaran record of seaweeds is too patchy for meaningful evaluation of evolutionary tempo, but these fossils do provide a paleobiological context for the interpretation of microfossil assemblages. In particular, fossils of multicellular algae relate the latest Mesoproterozoic and early Neoproterozoic diversification of acritarchs to the biological differentiation of ''higher" protists inferred from molecular phylogenies (Sogin et al., 1989; Knoll, 1992a).
If the view of eukaryotic evolution provided by molecular phylogenies is reliable, many of the protistan phyla that differentiated during the Proterozoic are not represented in the fossil record. Therefore, care must be exercised in ascribing generality to the preserved record. The prob-
lem is well understood by invertebrate paleontologists who enjoy excellent preservation for only a few of the more than 30 phyla of invertebrate animals. Estimates of evolutionary tempo and the timing of diversification and extinction events are broadly similar across phyla for Phanerozoic invertebrates and protists with good fossil records. Therefore, the limited clade diversity of Proterozoic and Early Cambrian acritarchs may reflect a broader pattern of early eukaryotic evolution.
A second type of biological uncertainty concerns the interpretation of paleospecies. In studies of acritarchs, analyses are necessarily done at the species level, because biologically meaningful higher taxa have not been defined. The uncertain phylogenetic relationships of most forms exacerbate the common paleontological problem of relating paleospecies to biological species. While the paleontological use of the term species is convenient and accurate in the sense of "most inclusive diagnosable units," what we can really measure is diversity of morphology. Estimates of tempo are, therefore, to be viewed as rates of morphological diversification and turnover within a preservable subset of early eukaryotes.
The Paleontological Data Base: Sampling Quality. Several hundred Proterozoic and Lower Cambrian formations are known to contain protistan microfossils (Schopf and Klein, 1992); however, many assemblages are indifferently preserved, poorly described, and/or accompanied by inadequate stratigraphic and paleoenvironmental information. Assessment of sample quality is better based on those assemblages that are well preserved, meticulously monographed, and well buttressed by stratigraphic and sedimentological data (Figure 1).
Older assemblages are both less numerous and less diverse than those of younger intervals. One might, therefore, suppose that low observed diversity is a product of poor sampling or poor preservation; however, sampling adequacy is not simply a function of assemblages per interval. It is also dependent on quality of preservation, facies and/or paleogeographic heterogeneity, and rates of taxonomic turnover. If fossils are well preserved, cosmopolitan, and slowly evolving, a limited number of samples may be sufficient to characterize the paleobiology of an interval.
This appears to be the case for the Mesoproterozoic acritarch record. The quality of fossil preservation in Mesoproterozoic mudstones (e.g., Peat et al., 1978) and silicified carbonates (e.g., Sergeev et al. , in press) matches the best seen in Neoproterozoic rocks, but the acanthomorphic (process or spine bearing) and other ornamented acritarchs seen in younger rocks of comparable environmental setting are not seen in these or any other rocks older than ca. 1100 Ma. In contrast, even metamorphosed Neoproterozoic rocks may contain ornamented acritarchs
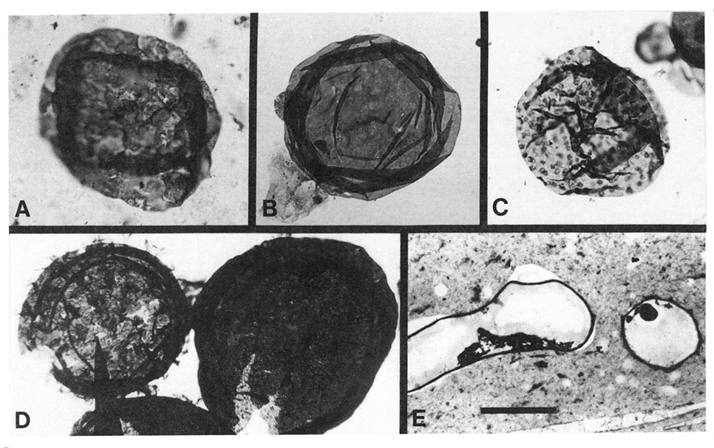
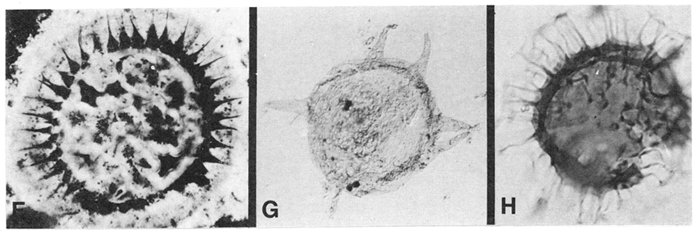
Figure 4 Representative Proterozoic and Early Cambrian acritarchs. A–C and the specimen on the left in D are spheromorphic; the specimen on the right in D and E–H are acanthomorphic. (A) Le iosphaeridia sp., Mesoproterozoic [M1] McMinn Formation, Australia. (B) Leiosphaeridia crassa, Neoproterozoic [N2] Miroyedikha Formation, Siberia. (C) Kildinosphaera verrucata, Miroyedikha Formation. (D) Shuiyousphaeridium macroreticulata (Left ) and Dictyosphaera incrassata (Right), late Mesoproterozoic [M3] Baicaoping Formation, China. (E) Trachyhystrichosphaera aimika, Neoproterozoic [N4] Draken Formation, Svalbard. (F) Tanarium densum, Neoproterozoic [N7] Doushantuo Formation, China. (G) Tanarium conoideum, Neoproterozoic [N7] Kursovsky Formation, Siberia. (H) Skiagia ciliosa , Early Cambrian [C4] Tokammane Formation, Svalbard. (Bar in E: A–C and H, 25 µm; D and G, 60 µm; E, 300 µm; G, 100 µm)
(Knoll, 1992b). Observations such as these suggest that differential preservation is not a principal determinant of observed diversity and turnover patterns.
In general, Proterozoic and Cambrian acritarch species have wide (and known) paleoenvironmental distributions and show little evidence of provincialism. This minimizes the likelihood that observed patterns are influenced strongly by differential sampling of facies among time intervals.
Perhaps the best indicators of sample quality are the degree of similarity among assemblages within an interval and the incremental taxonomic richness accompanying each new assemblage reported. The total number of assemblages known for the interval from 1700 to 1000 Ma is low, but the taxonomic similarity among samples is high. Insofar as knowledge of the age and environmental setting of an assemblage permits the prediction of taxonomic composition, the Proterozoic and Early Cambrian acritarch record appears to be sufficiently well sampled to permit the broad estimation of evolutionary tempo.
Despite my confidence that the existing record is governed more by evolution than by sampling, this paper should be read as a dispatch from the trenches and not as a definitive history. The events most likely to have escaped notice to date are short bursts of diversification and extinction of the type recorded in interval N7. The time intervals most likely to yield new assemblages that will modify the conclusions drawn here are those between 750 and 600 Ma ago, just prior to and including the Varanger ice age.
Early Eukaryotic Fossils: A Narrative Record
Acritarchs occur in rocks as old as 1900–1700 Ma (Zhang, 1986). The fossils are morphologically simple, but sedimentological distributions, size frequency distributions, and inferred excystment structures ally these remains to unequivocally eukaryotic microfossils that extend upward from this interval to the present. Independent evidence for the occurrence of late Paleoproterozoic to early Mesoproterozoic eukaryotes comes from significant sterane concentrations in bitumens (Summons and Walter, 1992) and problematic macrofossils (Walter et al., 1990; Han and Runnegar, 1992). There is no reason to assume that these remains document the evolutionary first appearance of the Eucarya nor even any necessary reason to insist that they mark the emergence of clades capable of synthesizing preservable walls or cysts. What can be concluded is that eukaryotic organisms were significant parts of marine ecosystems in the late Paleoproterozoic Era and that the fossil record of earlier periods is poor.
Acritarchs are widespread and abundant in late Paleoproterozoic and Mesoproterozoic rocks, and in all known assemblages morphological diversity is limited to a few taxa of smooth-walled spheromorphs (leiosphaerids; Figure 4A) differentiated on the basis of size, spheroids bearing a single filament-like emergence, and/or somewhat lumpy or irregular vesicles (Keller and Jankauskas, 1982; Jankauskas, 1989).
Morphological diversification began in the late Mesoproterozoic Eon with the appearance of striated spheromorphic vesicles (Valeria lophostriata ) and the first acanthomorphic acritarchs (Jankauskas, 1989; Knoll, in press). Chinese rocks poorly dated at ca. 1100 Ma contain the oldest known large (>100 µm) acanthomorphs (Yan and Zhu, 1992; Figure 4D)—a type of microfossil characteristically found in Neoproterozoic assemblages (Semikhatov, 1991; Knoll, in press). The 900- to 1000-Maold Lakhanda biota of Siberia (German, 1990) contains a moderate diversity of both acanthomorphs and lightly ornamented spheromorphs (Figure 4C). Latest Mesoproterozoic to early Neoproterozoic acritarch diversification is complemented by the first appearance of multicellular eukaryotes that can be placed in extant phyla. Red algae that display clear cellular differentiation are locally abundant in silicified peritidal carbonates of the Hunting Formation, arctic Canada (Butterfield et al., 1990); probable chromophytic algae are beautifully preserved in Lakhanda mudstones (German, 1990); and several green algal taxa occur in the younger Svanbergfjellet Formation of Svalbard (Butterfield et al., in press).
Acritarchs increase in both total and assemblage diversity in 900-to 800-Ma-old rocks (Figures 1–3), due largely to the differentiation of ornamented forms. Vase-shaped protistan tests also proliferate at this time. Most taxa that originated during the early-to-middle Neoproterozoic Era disappeared by the time of the great Varanger ice age (?650–590 Ma ago), but post-Varanger successions on three continents contain diverse assemblages of new and highly ornamented acritarchs (Yin, 1987; Knoll, 1992a; Zang and Walter, 1992; Moczydlowska et al., 1993; Figure 4F and G). This postglacial diversification is all the more remarkable because it is so short-lived. Even exceptionally preserved latest Proterozoic acritarch assemblages are limited to a handful of leiosphaerids and small acanthomorphs. The extinction correlates stratigraphically with the appearance of diverse Ediacaran-type metazoans; where diverse acanthomorphs and Ediacaran remains occur in the same succession, the former lie stratigraphically beneath the latter. However, correlation to the independent chronostratigraphic record of C and Sr isotopic variation suggests that the two types of assemblage coexisted for a limited interval (Kaufman and Knoll, in press). Acritarchs again diversified rapidly during the Early Cambrian (Volkova et al., 1983; Moczydlowska, 1991; Figure 4H).
Estimates of Evolutionary Tempo
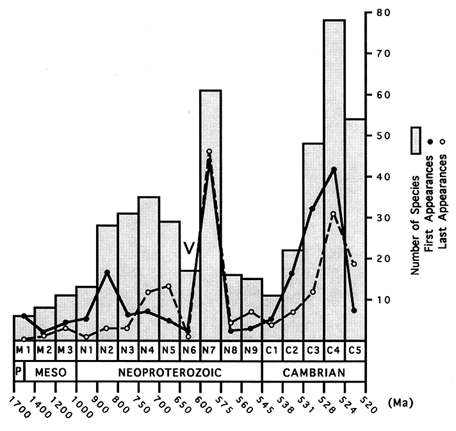
Figures 1 and 2 depict assemblage and total diversity through the interval from 1700 to 520 Ma ago. (Figure 3 plots total diversity data on a linear time scale to show more clearly the length of the initial interval of low diversity and the rapidity of subsequent changes.) The similarity of the figures is not surprising, given the wide facies and geographic distributions of many taxa. Species richness began low and rose only slightly during the first 700–800 Ma of the acritarch record—an interval longer than the entire Phanerozoic Eon. A burst of first appearances 900–800 Ma nearly doubled both assemblage and total diversity, bringing them to a new level that would persist with limited change until the Varanger ice age. The figures show diversity peaking 750–700 Ma ago and then declining to a minimum during the Varanger interval. However, intervals N5 and N6 are the most poorly sampled of the entire period under consideration. Taxa whose currently known last appearance is in N4 or N5 may well be discovered in closer proximity to Varanger strata, while some of the many acritarch taxa whose first known appearance is in N7 may be found in earlier intervals. For example, the large acritarchs Papillomembrana compta and Ericiasphaera spjeldnaessi, both conspicuous components of N7 assemblages, occur in clasts of the Biskopås Comglomerate, Norway, that underlie Varanger tillites (Vidal, 1990). On the other hand, few pre-Varanger taxa occur in the beautifully preserved assemblages that characterize N7, and in places like northwestern Canada, assemblages deposited just before the ice age contain only characteristically pre-Varanger taxa (Allison and Awramik, 1989). Thus, the marked change in assemblage composition across the Varanger interval is probably a stable feature of the record, and the extinctions inferred from the figures may have been concentrated in a brief interval before or during the Varanger ice age.
The high diversity of immediately post-Varanger acritarch assemblages is apparent from the figures. A burst of first appearances lifted both assemblage and total species richness to their Proterozoic maxima, and an ensuing maximum in last appearances subsequently reduced diversity to levels resembling those of the Mesoproterozoic and earliest Neoproterozoic. Seventy-five percent of recorded species disappeared, including most if not all large morphologically distinctive forms.
The first four intervals of the Early Cambrian exhibit sharp increases in numbers of first appearances; species richness within assemblages eclipsed its Proterozoic maximum in C3 (ca. 531–528 Ma), and total diversity peaked one interval later (C4, ca. 528–524 Ma ago). Last
TABLE 2 Species richness, rates of origination, and rates of extinction for Proterozoic and early Cambrian acritarchs
|
Interval (Ma ago) |
N |
FA |
FA/Ma |
FA/sp/Ma |
LA |
LA/Ma |
LA/sp/Ma |
|
M1 (1700–1400) |
6 |
6 |
0.02a |
0.003a |
0 |
0.00 |
0.000 |
|
M2 (1400–1200) |
8 |
2 |
0.01 |
0.001 |
2 |
0.01 |
0.001 |
|
M3 (1200–1000) |
11 |
5 |
0.03 |
0.004 |
3 |
0.02 |
0.002 |
|
N1 (1000–900) |
13 |
5 |
0.05 |
0.005 |
1 |
0.01 |
0.001 |
|
N2 (900–800) |
28 |
16 |
0.16 |
0.009 |
3 |
0.03 |
0.002 |
|
N3 (800–750) |
31 |
6 |
0.12 |
0.004 |
3 |
0.06 |
0.002 |
|
N4 (750–700) |
35 |
7 |
0.14 |
0.004 |
12 |
0.24 |
0.008 |
|
N5 (700–650) |
28 |
5 |
0.10 |
0.004 |
13 |
0.26 |
0.010 |
|
N6 (650–600) |
17 |
2 |
0.04 |
0.002 |
1 |
0.02 |
0.001 |
|
N7 (600–575) |
60 |
44 |
1.76 |
0.060 |
46 |
1.84 |
0.060 |
|
N8 (575–560) |
16 |
2 |
0.13 |
0.008 |
4 |
0.26 |
0.017 |
|
N9 (560–545) |
15 |
3 |
0.20 |
0.015 |
6 |
0.40 |
0.030 |
|
C1 (545–538) |
11 |
5 |
0.70 |
0.070 |
4 |
0.60 |
0.060 |
|
C2 (538–531) |
22 |
16 |
2.30 |
0.185 |
7 |
1.00 |
0.080 |
|
C3 (531–528) |
48 |
32 |
10.70 |
0.400 |
12 |
4.00 |
0.150 |
|
C4 (528–524) |
78 |
42 |
10.50 |
0.198 |
31 |
7.75 |
0.150 |
|
C5 (524–520) |
54 |
7 |
1.75 |
0.035 |
18 |
4.50 |
0.089 |
|
N, total species richness; FA, first appearance; LA, last appearance; FA (LA)/Ma, first (last) appearance per Ma; FA (LA)/Sp/Ma, first (last) appearance per species per Ma. For the calculation of per species rates of origination and extinction, standing diversity was taken to be the geometric mean of diversity at the beginning and end of each interval, making the simplifying assumption that all extinctions took place at the ends of intervals. a All species present in M1 are counted as first appearances, but some may have originated earlier. Thus, calculated rates of first appearance for M1 may be too high. |
|||||||
appearances also increase throughout these intervals and exceed first appearances at the end of the Early Cambrian.
Table 2 shows calculated rates of cladogenetic evolution for each of the intervals under consideration. For intervals M1 though N1, both total and per taxon rates of first and last appearances are low, indicating not only that diversity was low but also that constituent species were long lasting. (The calculated rates of first appearance for M1 may be misleading, in that all species are recorded as first appearances. The presence of simple acritarchs in rocks that may be older than 1700 Ma indicates that at least some of these forms may have originated earlier.) By 900–800 Ma ago (N2), total rates of origination had increased by an order of magnitude to a level at which they remained for the duration of the pre-Varanger Neoproterozoic. Interestingly, after an increase during interval N2, per taxon rates of origination returned to levels comparable to earlier intervals; both total and per taxon extinction rates increased toward the Varanger ice age.
Another order of magnitude increase in origination and extinction
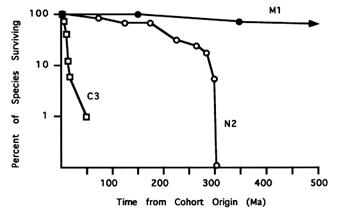
Figure 5 Cohort survivorship curves for species that originated during intervals M1 (1700–1400 Ma), N2 (900–800 Ma), and C3 (531–528 Ma). The abscissa denotes time since cohort origin.
rates attended the brief post-Varanger diversification event, after which terminal Proterozoic turnover returned to average Neoproterozoic levels. During the peak of the Cambrian acritarch radiation, origin and extinction rates both increased to levels an additional order of magnitude above the Neoproterozoic mean.
Cohort survivorship provides another means of evaluating evolutionary tempo (Van Valen, 1973; Raup, 1978, 1985). A comparison of the three cohort survivorship curves in Figure 5 shows that species originating in early Cambrian interval C3 turned over much more rapidly than those of Neoproterozoic cohort N2, which in turn decayed more quickly than Paleo- and Mesoproterocoic cohort M1. Very rough estimates of mean species duration and half-life (median species duration) confirm the order of magnitude increases in tempo between the Mesoand Neoproterozoic and again between the Neoproterozoic and Early Cambrian (Table 3). Thus, relative to earlier intervals, Cambrian acritarch assemblages contain more and more species that survive for shorter and shorter periods of time. Mean species duration and half-life for Cambrian acritarchs is similar to values computed for both younger protistan groups with good fossil records and Phanerozoic animal taxa (Table 3).
Discussion
How seriously should we take these figures? The general problems of sampling, data set size, and age estimation have already been noted. Imprecisions could easily alter estimates of tempo by a factor of two.
TABLE 3 Estimates of species durations
|
Taxonomic group |
Half-life, Ma |
Mean duration, Ma |
|
Acritarch cohort M1 |
1390a |
1960a |
|
Acritarch cohort N2 |
75 |
102 |
|
Acritarch cohort C3 |
5.4 |
7.7 |
|
Planktonic foraminifera |
5 |
7 |
|
Dinoflagellates |
9 |
13 |
|
Diatoms |
5.5 |
7.9 |
|
All invertebrates |
7.7 |
11.1 |
|
Half-life (median species duration) and mean duration were calculated according to Raup (1978, 1985). Data for Phanerozoic protists and invertebrates are from Van Valen (1973) and Raup (1985). a Of course, species that originated <1700 Ma ago cannot have true durations that exceed this age. Durations presented were calculated from the mean extinction rate (the slope of the cohort survivorship curve), which is very low. |
||
However, it is unlikely that any combination of sampling, taxonomic, or geochronometric errors exerts a major control on the principal pattern revealed by this analysis—that of doubling in diversity and ten-fold increase in rates of origination and extinction near the Mesoproterozoic–Neoproterozoic boundary and again in the Early Cambrian. Indeed, this exercise quantifies what we have long known as biostratigraphers—that acritarch-based correlation is difficult among Mesoproterozoic successions, somewhat easier in the Neoproterozoic, and precise in the Lower Cambrian.
Comparisons with Previous Diversity Estimates. The diversity trends noted here are broadly similar to those outlined a decade ago by Vidal and Knoll (1983), indicating an overall stability of pattern despite substantial increases in the numbers of species and assemblages described. There is one difference between the two estimates, however, and it is a major one; Vidal and Knoll's compilation lacks any inkling of the short-lived diversity peak in N7. That peak first appears in the literature in 1988 in a figure by Zang (1988), who discovered highly diverse N7 acritarchs in the Pertatataka Formation, Australia. Since then, even more diverse assemblages have been recognized in rocks of this age (Figure 1). The N7 spike cautions us that despite the overall stability of Proterozoic and early Cambrian acritarch diversity trends, short-lived diversification and extinction episodes may be missed at current levels of temporal sampling density.
A different view of Proterozoic protistan diversity was presented by Schopf (1992), who showed a broad diversity peak 1000–850 Ma ago, followed by a strong and continuing decline until the end of the eon.
Schopf's compilations of mean assemblage diversity for plankton and eukaryotes emphasize the inferred early Neoproterozoic diversity peak even more strongly. This discrepancy arises for at least three reasons: (i) Schopf's estimates of species richness for early Neoproterozoic assemblages from Russia significantly exceed those accepted here, (ii) most of the fossils that determine the diversity levels of intervals N4 to N7 in the present paper do not appear in Schopf's data set, largely because of recent discovery, and (iii) Schopf's calculation of mean assemblage diversity is swamped by low diversity assemblages of limited paleobiological value. For these reasons, I believe that the diversity trends shown in Figures 1–3 of the present paper better reflect the known record of early protists.
Intimations of Mode? As noted above, the increase in acritarch diversity and tempo near the Mesoproterozoic–Neoproterozoic boundary coincides with the appearance of identifiable red, green, and probable chromophyte algae in the record. Branching patterns in molecular phylogenies of the eukaryotes suggest that these algal taxa, along with stramenopiles (ciliates, dinoflagellates, and plasmodia), fungi, and the ancestors of animals, diverged rapidly relatively late in the history of the domain (Sogin et al., 1989). The paleontological data suggest that the radiation implied by molecular phylogenies occurred near the Mesoproterozoic–Neoproterozoic boundary; phylogenetic data, in turn, suggest possible explanations for the acceleration of evolutionary tempo documented by the fossils.
Nuclear introns, multicellular development that includes coordinated growth and cellular differentiation, and life cycles in which classical meiosis plays a prominent role are all characters displayed by higher eukaryotes but not earlier branching clades (Cleveland, 1947; Margulis et al., 1989; Tibeyrence et al., 1991; Palmer and Logsdon, 1991). The evolutionary relationships among these features are poorly understood, but possibly not coincidental. Either sexual life cycles or the exon shuffling made possible by introns could increase genetic variation and, thereby, accelerate evolutionary tempo (Schopf et al., 1973; Knoll, 1992a). This would be true of nuclear introns whether they first evolved at the time of higher protistan differentiation (Palmer and Logsdon, 1991) or were simply retained more readily in lineages characterized by sexual life cycles (Hickey, 1982).
Given the population genetic possibilities of such changes, it is surprising that the greater increase in acritarch diversity and tempo is concentrated at the beginning of the Cambrian Period. At this time, there is no evidence of genetic reorganization. New faster evolving clades may enter the acritarch record, but groups such as the prasino-
phytes that appear to have been important on both sides of the Proterozoic–Cambrian boundary also document the acceleration of cladogenetic tempo. Of course, the sharp increase in acritarch diversity and turnover coincides with a comparable evolutionary burst in animals. The nearly simultaneous radiation in two such phylogenetically, developmentally, and trophically disparate groups suggests the importance of ecology in determining the tempo of Cambrian (and later) evolution. Evolving animals would have contributed in several ways to the complexity of environments perceived by acritarch-producing protists: for example, through predation, the disturbance of pre-existing physical environments, the creation of new physical environments, and the alteration of nutrient fluxes in marine platform and shelf waters. Diversifying protists would have had reciprocal effects on animals. Diversity levels reached by Early Cambrian animals and protists were later eclipsed by continuing diversification, but the increased rates of turnover established at this time have persisted for the past 500 Ma (Table 2; Van Valen, 1973; Raup, 1985).
This is interesting in light of evidence that turnover in Phanerozoic marine communities may be coordinated among species and concentrated at times of environmental disturbance represented sedimentologically by sequence boundaries (Brett et al., 1990; Morris et al., 1992; Miller, 1993). This suggests that the basal Cambrian increase in the biological complexity of environments may have lowered the response thresholds of populations to physical fluctuations, perhaps by decreasing population sizes and effective niche breadth.
The short-lived acritarch radiation in N7 stands out as anomalous. Is this when faster evolutionary tempo was established in protists, only to be cut off by mass extinction? Might it correspond to an epoch of cryptic animal diversification that presaged the Ediacaran faunas of the next interval? Is the acritarch diversification causally related to oceanographic changes that accompanied the end of the Varanger glaciation, and if so, why aren't comparable changes observed in the wake of earlier Neoproterozoic ice ages?
Conclusions
We still glimpse early biological history through a glass darkly, but broad patterns are beginning to come into focus. These patterns suggest that on the time scale of eukaryotic evolution as a whole, evolutionary tempo has increased episodically. Morphological diversity and turnover rates were low for the earliest recorded period of early protistan evolution, an interval that lasted longer than the entire Phanerozoic Eon. Near the Mesoproterozoic–Neoproterozoic bound-
ary, the morphological diversity and turnover rates of acritarch-producing protists increased significantly, apparently as part of a larger increase in eukaryotic diversity that included heterotrophs as well as algae. Most notably, the Proterozoic and Early Cambrian record of acritarchs suggests that radiating animals had a profound effect on both diversity and turnover within clades already present in marine communities, implying an important role for ecology in fueling the Cambrian explosion and, perhaps, earlier protistan diversification.
Summary
In rocks of late Paleoproterozoic and Mesoproterozoic age (ca. 1700–1000 million years ago), probable eukaryotic microfossils are widespread and well preserved, but assemblage and global diversities are low and turnover is slow. Near the Mesoproterozoic–Neoproterozoic boundary (1000 million years ago), red, green, and chromophytic algae diversified; molecular phylogenies suggest that this was part of a broader radiation of ''higher" eukaryotic phyla. Observed diversity levels for protistan microfossils increased significantly at this time, as did turnover rates. Coincident with the Cambrian radiation of marine invertebrates, protistan microfossils again doubled in diversity and rates of turnover increased by an order of magnitude. Evidently, the Cambrian diversification of animals strongly influenced evolutionary rates within clades already present in marine communities, implying an important role for ecology in fueling a Cambrian explosion that extends across kingdoms.
I thank Zhang Yun, Yin Leiming, Kathleen Grey, Zang Wenlong, Malcolm Walter, Tadas Jankauskas, Nina Volkova, Tamara German, Alexei Veis, Vladimir Sergeev, Nicholas Butterfield, and—especially—Gonzalo Vidal and Malgorzata Moczydlowska for access to and discussions about Proterozoic and Cambrian microfossils. George Miklos, Kenneth Campbell, and J. William Schopf provided helpful criticisms of an earlier draft. This work was sponsored in part by National Aeronautics and Space Administration Grant NAGW-893 and National Science Foundation Grant BSR 90-17747.
References
Allison, C. W. & Awramik, S. M. (1989) Organic-walled microfossils from earliest Cambrian or latest Proterozoic Tindir Group rocks, northwest Canada. Precambrian Res. 43, 253–294.
Aseeva, E. A. (1983) Vendian microfossils of the Ukraine, in Vend Ukrainy [The Vendian of the Ukraine], eds. Velikanov, V. A., Aseeva, E. A. & Fedonkin, M. A. (Naukova Dumka, Kiev, pp. 102–127).
Bowring, S. A., Grotzinger, J. P., Isaachsen, C. E., Knoll, A. H., Pelechaty, S. M. & Kolosov, P. (1993) Calibrating rates of Early Cambrian evolution. Science 261, 1293–1298.
Brett, C. E., Miller, K. B. & Baird, G. C. (1990) Paleocommunity temporal dynamics: the long-term development of multispecies assemblages. Paleontol. Soc. Spec. Publ. 5, 178–209.
Butterfield, N. J., Knoll, A. H. & Swett, K. (1990) A bangiophyte red alga from the Proterozoic of arctic Canada. Science 250, 104–107.
Butterfield, N. J., Knoll, A. H. & Swett, K. (1994) Paleobiology of the Neoproterozoic of arctic Canada. Foss. Strata, in press.
Cleveland, L. R. (1947) The origin and evolution of meiosis. Science 105, 287–288.
Compston, W., Williams, I. S., Kirschvink, J. L., Zhang, Z. & Ma, G. (1992) Zircon U-Pb ages for the Early Cambrian time-scale. J. Geol. Soc. 149, 171–184.
German, T. N. (1990) Organic World Billion Year Ago (Nauka, Leningrad).
Han, T. M. & Runnegar, B. (1992) Megascopic eukaryotic algae from the 2.1-billion-year-old Negaunee Iron-Formation, Michigan. Science 257, 232–235.
Harland, W. B., Armstrong, R. L., Cox, A. V., Craig, L. E., Smith, A. G. & Smith, D. G. (1990) A Geologic Time Scale 1989 (Cambridge Univ. Press, Cambridge, U.K.).
Hickey, D. (1982) Selfish DNA: a sexually-transmitted nuclear parasite. Genetics 101, 519–531.
Jankauskas, T. V., ed. (1989) Mikrofossilii dokembriya SSSR [Precambrian Microfossils of the USSR] (Nauka, Moscow).
Jankauskas, T. V. (1982) Microfossils from the Riphean of the Southern Urals, in Stratotip rifeya: Paleontologiya, Paleomagnetizm [Stratotype of the Riphean: Paleontology, Paleomagnetism], ed. Keller, B. M. (Nauka, Moscow), pp. 84–120.
Kaufman, A. J. & Knoll, A. H. (1994) Neoproterozoic variations in the C-isotopic composition of seawater: implications for stratigraphy and biogeochemistry. Precambrian Res., in press.
Keller, B. M. & Jankauskas, T. V. (1982) Microfossils in the Riphean stratotype section in the Southern Urals. Int. Geol. Rev. 24, 925–933.
Knoll, A. H. (1992a) The early evolution of eukaryotes: a geological perspective. Science 256, 622–627.
Knoll, A. H. (1992b) Vendian microfossils in metasedimentary cherts of the Scotia group, Prins Karls Forland, Svalbard. Palaeontology 25, 751–774.
Knoll, A. H. (1994) Archean and Proterozoic paleontology, in Palynology: Principles and Applications, ed. Jansonius, J. & MacGregor, D. C. (Am. Assoc. Stratigraphic Palynologists Found., Tulsa, OK), in press.
Knoll, A. H. & Swett, K. (1987) Micropaleontology across the Precambrian-Cambrian boundary in Spitsbergen. J. Paleontol. 61, 898–926.
Knoll, A. H. & Walter, M. R. (1992) Latest Proterozoic stratigraphy and Earth history. Nature (London) 356, 673–678.
Knoll, A. H., Swett, K. & Mark, J. (1991) Paleobiology of a Neoproterozoic tidal flat/lagoonal complex: the Draken Conglomerate Formation, Spitsbergen. J. Paleontol. 65, 531–570.
Margulis, L., Corliss, J. O., Melkonian, M. & Chapman, D. J., eds. (1989) Handbook of Protoctista (Jones & Bartlett, Boston).
Miller, W. (1993) Benthic community replacement and population responses. N. Jb. Geol. Paläont. Abhandlangen 188, 133–146.
Moczydlowska, M. (1991) Acritarch biostratigraphy of the Lower Cambrian and the precambrian/Cambrian boundary in southeastern Poland. Foss. Strata 29, 1–127.
Moczydlowska, M., Vidal, G. & Rudavskaya, V. A. (1993) Neoproterozoic (Vendian) phytoplankton from the Siberian Platform, Yakutia. Palaeontology 36, 495–521.
Morris, P. J., Ivany, L. C. & Schopf, K. M. (1992) Paleoecological stasis in evolutionary theory. Geol. Soc. Am. Abstr. Prog. 24, A313.
Palmer, J. D. & Logsdon, J. M. (1991) The recent origins of introns. Curr. Opin. Genet. Dev. 1, 470–477.
Peat, C. R. Muir, M. D., Plumb, K. A., McKirdy, D. M. & Norvick, M. S. (1978) Proterozoic microfossils from the Roper Group, Northern territory, Australia. BMR J. Aust. Geol. Geophys. 3, 1–17.
Raup, D. M. (1978) Cohort analysis of generic survivorship. Paleobiology 4, 1–15.
Raup, D. M. (1985) Major features of the fossil record and their implications for evolutionary rate studies, in Rates of Evolution, eds. Campbell, K. S. W. & Day, M. F. (Allen & Unwin, London), pp. 1–14.
Schopf, J. W. (1992) Patterns of Proterozoic microfossil diversity; an initial, tentative, analysis, in The Proterozoic Biosphere, eds. Schopf, J. W. & Klein, C. (Cambridge Univ. Press, Cambridge, U.K.), pp. 529–552.
Schopf, J. W. & Klein, C., eds. (1992) The Proterozoic Biosphere (Cambridge Univ. Press, Cambridge, U.K.).
Schopf, J. W., Haugh, B. N., Molnar, R. E. & Satterthwait, D. F. (1973) On the development of metaphytes and metazoans. J. Paleontol. 47, 1–9.
Semikhatov, M. A. (1991) General problems of Proterozoic Stratigraphy in the USSR. Sov. Sci. Rev. G 1, 1–192.
Sergeev, V. N., Knoll, A. H. & Grotzinger, J. P. (1994) Paleobiology of the Mesoproterozoic Billyakh Group, northern Siberia. J. Paleontol. , in press.
Simpson, G. G. (1994) Tempo and Mode in Evolution (Columbia Univ. Press, New York).
Sogin, M., Gunderson, J., Elwood, H., Alonso, R. & Peattie, D. (1989) Phylogenetic meaning of the kingdom concept: an unusual ribosomal RNA from Giardia lamblia. Science 243, 75–77.
Summons, R. E. & Walter, M. R. (1992) Molecular fossils and microfossils of prokaryotes and protists from Proterozoic sediments. Am. J. Sci. 290-A, 212–244.
Tappan, H. (1980) The Paleobiology of Plant Protists (Freeman, San Francisco).
Tibeyrence, M., Kjellberg, F., Arnaud, J., Oury, B., Darde, M. & Ayala, F. (1991) Are eukaryotic microorganisms clonal or sexual? A population genetics vantage. Proc. Natl. Acad. Sci. USA 88, 5129–5133.
Van Valen, L. (1973) A new evolutionary law. Evol. Theory 1, 1–30.
Veis, A. F. & Semikhatov, M. A. (1989) Lower Riphean Omakhtin association of microfossils in eastern Siberia, composition and conditions of formation. Izv. Akad. Nauk SSSR Ser. Geol. 5, 36–54.
Veis, A. F. & Vorobyeva, N. G. (1992) Riphean and Vendian microfossils of the Anabar Massif. Izv. Akad. Nauk SSSR Ser. Geol. 8, 114–130.
Vidal, G. (1976) Late Precambrian microfossils from the Visingsö Beds, southern Sweden. Foss. Strata 9, 1–57.
Vidal, G. (1981) Micropaleontology and biostratigraphy of the Upper Precambrian and Lower Cambrian in East Finnmark, northern Norway. Norges Geol. Unders. Bull. 362, 1–53.
Vidal, G. (1990) Giant acanthomorph acritarchs from the Upper Proterozoic in Southern Norway. Palaeontology 33, 287–298.
Vidal, G. & Ford, T. D. (1985) Microbiotas from the late Proterozoic Chuar Group (northern Arizona) and Uinta Mountain Group (Utah) and their chronstratigraphic implications. Precambrian Res. 28, 349–389.
Vidal, G. & Knoll, A. H. (1983) Proterozoic plankton. Mem. Geol. Soc. Am. 161, 265–277.
Vidal, G. & Peel, J. S. (1993) Acritarchs from the Lower Cambrian Buen Formation in North Greenland. Bull. Grønl. Geol. Unders. 164, 1–35.
Volkova, N. G. (1990) Acritarchs and other plant microfossils of the East European Platform, in The Vendian System, eds. Sokolov, B. S. & Iwanowski, A. B. (Springer, Berlin), Vol. 1, pp. 155–164.
Volkova, N. G. et al. (1983) Plant microfossils, in Upper Precambrian and Cambrian Palaeontology of the East-European Platform, eds. Urbanek, A. & Rozanov, A. Yu. (Wydawnictwa Geologiczne, Warsaw), pp. 7–46.
Walter, M. R., Du, R. & Horodyski, R. J. (1990) Coiled carbonaceous megafossils from the Middle Proterozoic of Jixian (Tianjin) and Montana. Am. J. Sci. 290-A, 133–148.
Yan, Y. & Zhu, S. (1992) Discovery of acanthomorphic acritarchs in the Baicaoping Formation of Yongli, Shanxi, and its geological significance. Acta Micropalaeontol. Sinica 9, 278–282.
Yin, L. (1987) Microbiotas of the latest Precambrian sequences. Strat. Palaeontol. Systemic Bound. China Precamb.-Camb. Bound. 1, 415–494.
Zang, W. (1988) Dissertation (Australian National Univ., Canberra, Australia).
Zang, W. (1990) Sinian and Early Cambrian floras and biostratigraphy on the South China Platform. Palaeontograph. Abteilung B 224, 75–119.
Zang, W. & Walter, M. R. (1992) Late Proterozoic and Cambrian microfossils and biostratigraphy, Amadeus basin, central Australia. Mem. Assoc. Aust. Palaeontol. 12, 1–132.
Zhang, Z. (1986) Clastic facies microfossils from the Chuanlinggou Formation (1800 Ma) near Jixian, North China. J. Micropalaeontol. 5, 9–16.
| This page in the original is blank. |






















