
Two
Linking Pattern to Process: A Regional-Scale Approach
A research program to understand the role of human activities in altering biodiversity must involve studies at relatively large spatial and temporal scales. The open nature of marine systems (Box 4) dictates a regional-scale approach that involves studying multiple separate sites within an appropriately large geographic region. Physical (together with chemical and geological) oceanographic processes provide the dynamic backdrop against which all biological processes take place in the ocean. Thus, biological and physical considerations should be used to delimit regional-scale systems. Water motion affects biology by acting as a transport mechanism for organisms and their propagules, as a dynamic boundary between regimes, and as a force to which organisms must adapt or respond, for example, in their feeding and locomotor activities (Vogel, 1981; Nowell and Jumars, 1984; Denny, 1988, 1993).
Delineating the boundaries of geographic regions thus involves both biological and physical criteria—criteria that define the maximum spatial and temporal scales generally required to characterize adequately the processes that control biodiversity maintenance at a local scale. This means, for example (as explained below) that it is imprudent to study long-term prospects for biodiversity in the Chesapeake Bay independently of other Atlantic coastal plain estuaries, or to assume that the coral reefs of Florida are independent of other reef tracts in the greater Caribbean area (see Box 5).
Fundamental research questions concerning the creation, maintenance, and regulation of biodiversity have generally been studied experimentally on very small-scales (centimeters to hundreds of meters). Consequently, much is known about mechanisms of interaction and the proximate causes of local dynamics, much less about how the sum total of species at a site indirectly affects individual
|
Box 4: Marine ecosystems have many attributes distinct from those of terrestrial ecosystems that have important implications for understanding biodiversity. DISTINCTIVE FEATURES OF MARINE ECOSYSTEMS Marine and terrestrial ecosystems differ in significant ways that suggest that the ocean may respond to human perturbations in a fundamentally different manner from the land. Some of the unique attributes of marine ecosystems are listed below:
|
species, and almost nothing about interactions among communities at different sites (e.g., S.A. Levin, 1992). Changes in climate and the flux of nutrients, plankton, or larval recruits all depend on regional oceanographic processes, often largely independent of local events (e.g., Butman, 1987; Roughgarden et al., 1988; Underwood and Petraitis, 1993). There are also nagging questions about minimum population sizes and critical areas for survival of species in the sea (e.g., Nee and May, 1992). None of these issues can be addressed at a local scale.
Moreover, understanding larger-scale phenomena is central to any scientifically rational plan for biodiversity conservation and maintenance.
The Spatial Continuum
''... the problem is not to choose the correct scale of description, but rather to recognize that change is taking place on many scales at the same time, and that it is the interaction among phenomena on different scales that must occupy our attention."
S.A. Levin (1992, p. 1947)
Biodiversity depends on processes operating at many different spatial, temporal, and organizational scales (S.A. Levin, 1992). In general, these scales are broadly overlapping, with processes interacting among scales (Fig. 1). The most relevant scales for studying particular species or dynamical interactions among species will not be the same as for others, and hence there is no single correct set of scales for viewing a system. Rather, there must be awareness of the selective filter a particular perspective imposes on observed dynamics and of how information is transferred across scales (Ricklefs, 1987, 1990; Underwood and Petraitis, 1993).
A "patch" is an initially relatively uniform portion of a habitat whose limits
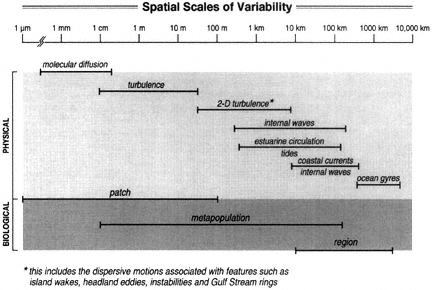
Figure 1
The spatial scales of variability associated with physical and biological processes (adapted from NRC, 1987).
are defined by relatively abrupt changes in the abundance of common species (Paine and S.A. Levin, 1981), or by the physical phenomena contributing to patch formation and maintenance (Fig. 1). This is the scale over which individual organisms interact with each other and with their immediate environment. Patches in the pelagic environment are three-dimensional and mobile, whether they consist of actively moving nekton (Hamner et al., 1983; Boudreau, 1992) or passively advected plankton (Haury et al., 1978; Fasham, 1978; Yoder et al., 1981). In pelagic systems, patches are found on almost every scale of observation (Owen, 1989; Powell and Okubo, 1994), and measures of patchiness seem to change continuously across scales (e.g., S.A. Levin et al., 1989). Defining and understanding characteristic patch sizes for plankton thus remain a focus of considerable interest, and although patchiness in the pelagic environment is less well-understood than in some benthic systems, new acoustic and imaging technologies are increasing the ability to measure patchiness (see Box 12). For intertidal and sublittoral benthic communities, distinct patches are much more evident and may impose a hierarchical organization to the system (Paine and S.A. Levin, 1981). In these systems, interindividual and interspecific interactions occur to a larger extent within patches, and processes such as dispersal, disturbance, and predation impose interpatch correlations (Sousa, 1985).
The collection of patches that interact or are otherwise connected constitute a "metapopulation" (Gilpin and Hanski, 1991). Note that this includes empty patches of suitable habitat, for example, in a mussel bed or on a coral reef, where a group of mussels or corals has been recently consumed by sea stars or destroyed by moving debris during violent storms (Connell, 1978; Paine and S.A. Levin, 1981). The subsequent successional events within each patch, and the movements of organisms among patches, can be integrated to characterize the demography of patches at the site. Physical processes operating over a wide range of time and space scales may affect metapopulation dynamics (Fig. 1). The theory of metapopulations is still in its infancy, and few marine populations have been studied at this scale. Communities are the sum total of metapopulations in a given habitat at a site.
A "region" comprises all the sites within a biogeographic province whose limits are defined by the relative homogeneity of the biota, as well as by unifying geographic features and oceanographic processes (Fig. 2). Ecological connections between sites within a region depend on fundamentally different processes from those between patches within sites (Butman, 1987; Roughgarden et al., 1988; Underwood and Petraitis, 1993). For example, the exchange of nutrients, food, and colonists between one site and another all depend on patterns of coastal and ocean currents that are largely independent of local events (e.g., see Box 5). Such processes can be modeled as a metapopulation of sites or metacommunities (Gilpin and Hanski, 1991; D.S. Wilson, 1992). There are, however, almost no data showing how changes in biodiversity at one site within a region affect the diversity at another.
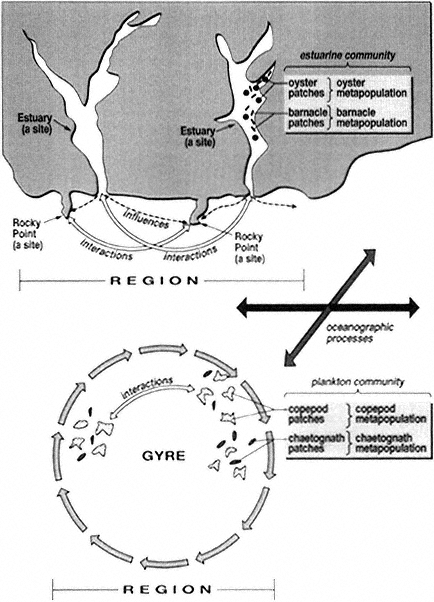
Examples of coastal regional-scale systems include the region that contains all the coral reefs of the Hawaiian Islands, the region that contains all the rocky open coasts of the Pacific Northwest, or the region that contains all the estuaries and enclosed bays of the Gulf of Mexico coastal plain. These regions thus encompass all physical and biological processes that could affect the reefs, rocky shores, or estuaries and bays contained within them. For example, the frequency, magnitude, and spatial extent of freshwater outflow from major rivers or local estuaries may affect coastal communities, and, likewise, offshore circulation is one factor controlling the residence time of water (containing larvae and nutrients) within bays and estuaries.
Within pelagic open-ocean systems a region could be specifically defined by a physical-oceanographic feature, by relatively persistent circulation patterns, or by water-property distributions. Thus, physical-oceanographic processes that occur on a variety of spatial scales (see Fig. 1) are critical elements to the definition of a pelagic regional-scale system (Fig. 2). Present-day oceanographic features that are persistent over relatively long time scales (days to months or longer) are likely to have had the most impact on defining, from a physical perspective, contemporary biological boundaries (but see below regarding longer time scales). Well-known examples of persistent physical features in North America include tidal mixing fronts (e.g., Gulf of Maine), gyres (e.g., North Pacific, Bering Sea), upwelling fronts (e.g., coastal Eastern Pacific), and warm-core rings (e.g., North Atlantic).
Temporal Considerations
"Geological history and oceanographic processes are the warp and woof of the biological understanding of any marine habitat."
Dayton et al., (1994, p. 90)
Biological patterns and processes also vary over a range of temporal scales. Time scales associated with speciation may range from thousands to millions of years, depending on such factors as the mode of speciation and the ecology and life-history characteristics of the organisms involved. Yet, time scales for genetic changes within species may be much less—on the order of years to centuries. Almost nothing is known about appropriate time scales for significant genetic changes in microorganisms. But, clearly, ecological interactions among members of a community, and adaptations of organisms to their environment, may develop over very long time scales indeed. In fact, because current regimes and other physical transport characteristics of the marine environment have changed over geologic time due to changing sea level, coastline, and bottom morphology, and water temperature and salinity characteristics, it cannot be assumed that organisms are necessarily optimally adapted to the physical regime in which they presently reside (Valentine and Jablonski, 1993). Any understanding of present-
day patterns of biodiversity therefore must be embedded firmly in an historical perspective, underscoring the importance, for example, of knowing if a given species was introduced or is native to the environment in which it now occurs (Carlton, 1989; Valentine and Jablonski, 1993) and of retrospective analyses in general (discussed in Chapter 6).
Understanding and predicting changes in biodiversity due to effects of human activities requires consideration of the time scales of variation of physical processes relative to biological processes. This consideration defines the relevant regional-scale of study for a given community or habitat. This marine biodiversity initiative is envisioned as a decadal-scale program. Given this time frame and physical considerations, important life-history features of the organisms include, for example, generation time, larval type (direct development or planktonic), dispersal period in the plankton (for pelagic larvae or spores of benthic organisms), and resting stage duration (for dormant benthic stages of planktonic organisms). Generation time, for example, is one criterion dictating the ultimate duration of a study whose goal is to understand changes in biodiversity due to effects of human activities.
Moreover, spatial scales tend to increase as time scales increase. Thus, for example, time scales associated with various physical oceanographic processes define advection length scales for planktonic larval transport, and the frequency and duration of resuspension events for benthic resting stages. Based on larval transport considerations alone, then, regional spatial scales would tend to be smaller for polar compared with temperate benthic invertebrate communities because the proportion of direct developers tends to increase with latitude (Thorson, 1950; Mileikovsky, 1971; Strathmann, 1985).
Defining the Most Meaningful Scales of Study
Ultimately, a variety of criteria, based on all life-history stages of the organisms, the oceanographic processes relevant to these stages, and the critical environmental issues in the region, must be used in defining the maximum spatial and temporal scales of study (e.g., Box 5). Once the maxima have been defined, shrewd insight will be needed to select the most meaningful smaller-scales of study. A compelling terrestrial example is the 12-year study of Tilman and Downing (1994; but see also Givnish, 1994 and Tilman et al., 1994) on the relationship between the biodiversity of vascular plants (manipulated by nitrogen fertilization) and the stability of the grassland ecosystem (in response to drought). This marine biodiversity program would emphasize inclusion of biological and oceanographic processes at the largest relevant scales—recognizing the connectivity among sites within a geographic region—while acknowledging the critical importance of ecological processes and physical-biological coupling at smaller, site-specific scales. This initiative does not recommend studying all processes at all scales within a selected geographic region. Rather, this initiative
|
Box 5: Understanding the origins of profound biological changes at one site will require understanding similar or related changes at the regional level. FLORIDA BAY: THE NEED TO SEEK A BIGGER PICTURE In what may be a system that mirrors the broad range of human alterations to estuaries, there is disturbing evidence that the Florida Bay ecosystem is collapsing. Florida Bay (2,200 km2), and the adjacent Florida Keys, is the only tropical marine ecosystem in the continental United States, with a vast economic value for the state of Florida. Florida Bay and the coral reef tracts of the Keys are connected by coastal currents, and thus the Bay may have a critical influence on the reefs. Likewise, alterations to land runoff and freshwater systems can affect Bay waters.
There is considerable scientific debate over the causes of collapse, and thus of the potential for recovery, or the steps that are necessary to implement and facilitate recovery. Whereas some of these changes could be considered as only "local" effects, it is important to view this ecosystem collapse in terms of broader changes in the Caribbean marine and terrestrial ecosystems. A more narrow focus on only Florida Bay may be doomed to failure. An important further lesson of the Florida Bay situation is that the water systems on land that impact the Bay, and in turn the Keys offshore, also must be understood and managed. |
is designed to accommodate studies at all scales relevant to a specified biodiversity research project.
A Rationale for Studying Several Types of Regional Model Systems
Three biological rationales suggest the need to use a regional-scale approach to study, concurrently, a variety of different types of marine ecosystems.
- Various marine systems present different opportunities to address fundamental questions because the systems vary in their evolutionary history, resulting in different biotas; in their connectivity (among sites within a system and among systems); and in the knowledge and description of their biodiversity.
- Establishing the appropriate regional-scale is simpler for systems whose boundaries are more easily defined, and for which there is greater knowledge of specific anthropogenic stresses and their effects, as well as greater understanding of the role of dispersal mechanisms.
- There are fundamentally different levels of environmental impact in different marine systems: some are now highly impacted (such as most coastal and coral-reef systems), whereas for others there is a grave possibility of future impact (such as open-ocean and deep-sea systems through, for example, human alterations to the global climate) suggesting the advisability of "anticipatory" research.
At larger—possibly the largest—scales, it is clear that distinct regional systems thousands of kilometers apart may have reciprocal influences on each other. Currents carry larvae of some species transoceanic distances, far from their parents (Scheltema, 1986). For example, the larvae of the Caribbean spiny lobster spend 9 or more months traversing the open-ocean pelagic ecosystems of the North Atlantic gyre (Farmer et al., 1989). Thus, an important factor influencing the abundance of adult lobster populations may be very distant oceanographic and biological processes—in an ecosystem not likely to be directly studied by Caribbean lobster ecologists. Likewise, larvae of the American eel undergo planktonic development in the Sargasso Sea, thousands of kilometers from the adult habitat in coastal rivers (Avise et al., 1986; McCleave, 1993). Also, currents associated with periodic El Niño-Southern Oscillations are known to transport warm water and propagules across the Pacific, causing population and community changes (Glynn, 1988).
Although this marine biodiversity initiative focuses on the larger within-regional-scale oceanographic and ecological processes that directly bear on the expression of local, site-specific biodiversity, awareness of the ocean-wide influences between regions remains important to an overall understanding of regional biodiversity. Furthermore, addressing a well-defined set of research questions in regional-scale studies of several different types of marine ecosystems will permit meaningful comparisons of the causes and consequences of changes in biodiversity due to human activities. Such comparisons will greatly enhance predictive capability relative to future human impacts on the marine environment.
Objectives of this Initiative
This marine biodiversity initiative sets a research agenda that represents a fundamental change in the approach to measuring and studying biodiversity in
the oceans by emphasizing an integrated regional-scale research strategy within an environmentally relevant and socially responsible framework. The purposes of this proposed initiative are thus to define a suite of cross-system basic research questions that transcend the typical approach to studying the patterns, processes, and consequences of marine biological diversity. These questions are designed to yield new insights and to enhance understanding of the factors controlling biological diversity, and to increase predictive capability regarding changes in marine biodiversity due to anthropogenic effects in order to provide essential management and conservation guidelines.
The five fundamental objectives of this initiative are:
- to understand the patterns, processes, and consequences of changing marine biological diversity by focusing on critical environmental issues and their threshold effects, and to address these effects at spatial scales from local to regional and at appropriate temporal scales;
- to improve the linkages between the marine ecological and oceanographic sciences by increasing understanding of the connectivity between local, smaller-scale biodiversity patterns and processes, and regional, larger-scale oceanographic patterns and processes that may directly impact local phenomena;
- to strengthen and expand the field of marine taxonomy through training, the development of new methodologies, and enhanced information dissemination, and to raise the standard of taxonomic competence in all marine ecological research;
- to facilitate and encourage the incorporation of (1) new technological advances in sampling and sensing instrumentation, experimental techniques, and molecular genetic methods, (2) predictive models for hypothesis development, testing, and extrapolation, and (3) historical perspectives (geological, paleontological, archeological, and historical records of early explorations) in investigations of the patterns, processes, and consequences of marine biodiversity; and
- to use the new understanding of the patterns, processes, and consequences of marine biodiversity derived from this regional-scale research approach to improve predictions of the impacts of human activities on the marine environment.










