
Four
Regionally Defined Model Systems: Examples of Habitats
As has been stated previously, the processes responsible for biodiversity changes due to anthropogenic effects must be studied on all relevant scales. One of the most important results of metapopulation theory for the conservation of biodiversity is that changes in the abundance of a species at some sites may cause its extinction or increase at other sites as an inevitable result of its vital statistics, natural history, and colonization potential (Nee and May, 1992). Thus conservation and management of marine biodiversity ultimately depend on understanding regional processes. Such a regional approach must simultaneously include relevant small-scale ecological studies and good taxonomy, which are essential to the entire picture. These activities must fit together to provide the necessary links between local and regional perspectives.
Much of the difficulty in studying regional biodiversity is that it depends on both ecosystem and population processes that are traditionally studied at different scales and in different disciplines (S.A. Levin, 1992). On the one hand, biological oceanographers have tended to emphasize ecosystem processes such as primary production operating on the scale of ocean currents and upwelling systems. On the other hand, marine ecologists have tended to emphasize processes affecting populations of species on a local scale (e.g., patches, see Figures 1 and 2). Thus a major goal of this marine biodiversity initiative would be to bring together population and ecosystem approaches at the same scales to understand, when possible, the origin and maintenance of marine biodiversity.
These considerations, along with the rationales discussed in Chapter 2 "Linking Pattern to Process," suggest that it will be necessary to focus on regionally defined model systems. These systems are of the appropriate scale to address
questions concerning the linkages between ecological processes operating at relatively small-scales and oceanographic processes impacting these systems at larger scales. A dedicated regional focus provides the framework for simultaneously conducting basic and applied research in biological oceanography, ecology, and conservation biology aimed at understanding changes in biodiversity due to human activities.
Discussed below are six types of habitats that would be contained within regional model systems. They are described in terms of the intrinsic interest and value of the regions, how they have been or may be impacted by anthropogenic effects, and the compelling opportunities for study. The systems cover the complete range of spatial scales discussed earlier (see also Figure 3), but the degree of understanding of processes that control biodiversity within and across these scales varies widely among the systems. The first three systems (estuaries and bays,
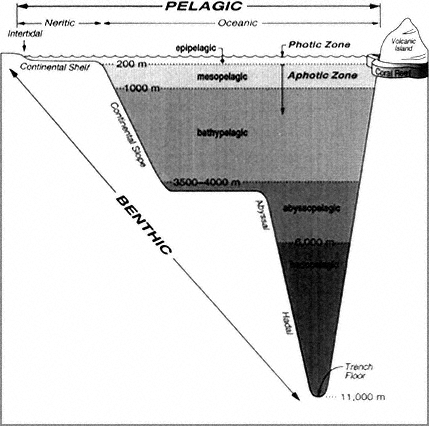
Figure 3
Regions of the marine environment.
coral reefs, and temperate rocky shores) have been and are undergoing heavy impacts from human activities. For the last three systems (shelf-slope systems, the deep sea, and the pelagic open ocean), a sense of urgency relative to what human activities may now be doing or could do in the near future is driven by profound ignorance of how diversity in these systems is created or maintained.
Estuaries and Bays
The majority of people in the world live within 100 kilometers of bays and estuaries (Norse, 1993), and such environments command enormous attention and use. Estuaries have long been associated with:
- some of the world's greatest fisheries for oysters, clams, shrimp, crabs, and fishes. In addition, bays, marshes, and seagrass beds act as well-known ''nursery grounds" for larvae and juveniles of many species (Weinstein, 1979; Boehlert and Mundy, 1988). Yet many of these notable fisheries are at or near collapse because of overfishing, disease, and pollution (e.g., Maurer et al., 1971; Newell, 1988).
- marshes, seagrass beds, and mangroves. Such coastal vegetation is ecologically critical as detrital and nutrient sources driving nearshore production, filters for land runoff, protection from coastal storms, sediment traps, and sediment stabilizers (Fenchel, 1977; Adam, 1990). Marshes also have important aesthetic and recreational value in their "pristine" condition (Teal and Teal, 1969; Chabreck, 1988). They are being rapidly lost, however, because of extensive draining and filling practices (Dahl et al., 1991).
- harbors and marinas. Dredged channels maintain open routes for economically important international shipping. In turn, international shipping means that ballast water is discharged frequently, facilitating the establishment of exotic species (see Boxes 6 and 7) that can significantly change ecosystem structure and function (e.g., Fig. 4). Moreover, floats, docks, and pilings have replaced mud-flats, marshes, and seagrass meadows to provide space for recreational pleasure boats (Zedler, 1994).
- pollution discharge. Estuaries and bays have long received the bulk of human-generated municipal and industrial wastes that enter the oceans from the land, rendering many of them unfit for fisheries production (Kennish, 1992; Schubel, 1994).
These and many other activities in the world's most populated areas adjacent to marine environments mean that estuaries and bays are where the greatest proportion of natural habitat has been destroyed or severely altered (Dahl et al., 1991). Thus, estuaries and bays, and their component marshes, seagrass beds, and mangroves provide striking opportunities for understanding how a profusion of human activities, acting singly and in combination, decrease, maintain, or increase biodiversity (e.g., see Box 5). In many estuaries, the original life is all
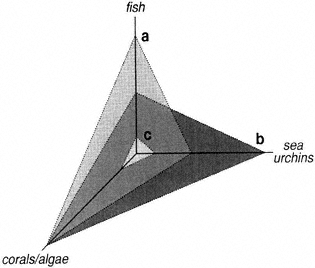
Figure 4
Changes in chlorophyll a concentration with increasing abundances of the introduced clams Potamocorbula amurensis and Mya arenaria in San Francisco Bay.
Source: Alpine and Cloern (1992).
but gone, in terms of species diversity, abundances, and local distributions. In more remote estuaries, little is known regarding the balance between pristine and altered conditions. And, for other estuaries, there appear to be far more species now than ever before (e.g., 200 introduced marine, brackish, and freshwater animal and plant species have been added in 150 years to the San Francisco Bay and delta region [Carlton and Cohen, pers. comm., 1994]), but little is known about the structural or functional consequences of this increased diversity, or of the addition of so many species to a single system at such a rapid rate.
In addition, the naturally lower diversity of most estuaries provides an opportunity to study the relationship between diversity and ecosystem function over a range of systems (see Box 9).
Coral Reefs
"At the rate things are going, there may be very few Caribbean corals to be affected by global climate change (Smith & Buddemeier 1992)."
Jackson (1994, p. 59)
Coral reefs are renowned for their remarkable diversity (Connell, 1978). Reef organisms display extraordinary specializations, intensive predator-prey evolutionary pathways, and competitive interactions within and among species
(e.g., Jackson, 1991). Because this diversity is expressed in a spectacular array of form and color, and because of increasing threats to the survival of this diversity (Hallock et al., 1993), coral reefs have attracted considerable tourism and public interest. Moreover, modern coral reefs, representing approximately 6,000 years of growth during the most recent period of sea-level rise, are the oldest and largest biogenic structures in nature bearing detailed paleoecological and climate records (Dunbar and Cole, 1993; S.V. Smith and Buddemeier, 1992). Whereas global climate change, and the potential associated effects of sea-level rise, increasing water temperature, and increasing ultraviolet (UV-B) radiation, are possible long-term threats to reefs, there is an immediate need to manage the relentless impact of explosive human population growth on reef habitats (D'Elia et al., 1991; Norse, 1993).
Reefs provide good examples of the importance of linkages between habitats—reef biodiversity is dependent on adjacent ecosystems for feeding areas and nursery grounds and as buffers against land runoff of sediments and nutrients (Kuhlmann, 1988; Ogden, 1988). Runoff from terrestrial environments is one of the most severe problems affecting reefs around the world. Sediment load from agriculture or forest clear-cutting, freshwater, and industrial activity are known to be damaging reefs (Grigg, 1984, 1994; Hodgson, 1989; Richmond, 1993; Sebens, 1994) either by killing colonies, preventing settlement of new recruits, interfering with sexual reproduction, or all three. Eutrophication from agricultural fertilizers and human sewage is a particular problem, first because organic enrichment causes faster growth in weedy macroalgae than in corals, overgrowing and killing them (S.V. Smith et al., 1981), and second because corals are adapted to live in a nutrient-poor environment, and thus overfertilizing alters the productive but very fragile relationship between the corals and their algal symbionts (Falkowski et al., 1993). In addition, reefs hold a significant portion of the fisheries resources of developing tropical countries, and they are very sensitive to overfishing, especially the removal of large predators and herbivorous fishes (see Boxes 2 and 6).
Biotic and abiotic disturbances on reefs that shift reef composition from framework builders (corals) to nonframework builders (algae) have particularly dramatic effects on biodiversity. One example is provided in Figure 5 that illustrates how the delicate natural balance (a) between fishes, the ratio of coral to large algae, and the herbivorous sea urchin Diadema antillarum in Caribbean reefs has been tipped first one way (b) by overfishing and then another way (c) by mass mortality of the urchin due to an unknown pathogen (Jackson, 1994).
Excellent opportunities exist in coral reef systems to look directly at the dynamic interface between the natural patterns, processes, and consequences of biodiversity, and the increasing pressures from human activities. A broad range of observed transitions between different reef communities exhibiting differing levels of impacts and thus threshold effects—effects that may be irreversible over the scale of a human lifetime (Knowlton, 1992)—offers an irresistible comparative menu for study.
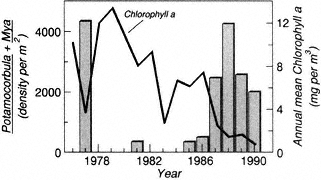
Figure 5
Hypothesized response of Caribbean coral reef communities to overfishing and sea urchin disease.
Source: Jackson (1994).
Temperate Zone Rocky Shores
Rocky intertidal zones, and the shallow sublittoral kelp forests just below them, have provided the conceptual framework for research on most other benthic ecosystems (Connell, 1961; Paine, 1980). They therefore offer one of the best possible opportunities for rapid advance in understanding of the dynamics of regional systems.
Many temperate rocky shores have been very extensively altered by the virtual removal of large vertebrate predators such as sea otters (Estes and Palmisano, 1974). Human foraging on intertidal shores and, in more recent decades, recreational tidepool tourism have resulted in some of the more dramatic demonstrations of how direct removal of large numbers of invertebrate and vertebrate carnivores and herbivores, combined with sustained trampling, can locally obliterate the fauna and flora of a rocky shore (Beauchamp and Gowing, 1982; Bally and Griffiths, 1989). Thirty years of visits by hundreds of thousands of school children to the tidepools of what is now a "marine reserve" in central California have resulted in the complete local absence of seashore snails and crabs that are considered to be the most typical representatives of Pacific marine life (R. Breen, pers. comm., 1994). In Chile, the best experimental examples of the devastating effects of human extraction of animals on a marine community come from the rocky intertidal zone (see Box 13).
Rocky shore systems, because of their geographic ubiquity and experimental tractability, remain prototype systems for exploring the connection between biodiversity and ecosystem function. Biodiversity issues at all trophic levels, especially at the microbial level, remain unexplored. The interaction between nearshore oceanography and larval transport is basic to understanding the presence, absence, and distribution of species. This interaction, set against the high
variability of dispersal capability among species, seems certain to generate variation in community composition at large spatial scales. The extent of this variability, its relation to whether particular sites are larval sources or sinks, and biodiversity as a consequence of local interaction remain major topics in metapopulation dynamics and conservation biology. Marine, nearshore conditions provide an appropriate environment to investigate the interplay between connected populations in open systems.
Continental Shelves and Slopes
"The canneries themselves fought the war by getting the limit taken off fish and catching them all ... It was the same noble impulse that stripped the forests of the West and right now is pumping water out of California's earth faster than it can rain back in."
Steinbeck (1954)
Continental shelves represent the great interface between the continents and open oceans. Directly and indirectly impacted by the natural and human effects that operate on land margins, the shelves are the only "open ocean" most people will ever see or know—and yet the shelves are separated by boundary currents from most of the sea. The concept of shelf waters representing "the ocean" is reinforced by the popularization since the 1950s of "food from the sea," resulting in shelves being portrayed primarily as sites of most of the major world fisheries, located on fishing banks, in upwelling zones, and on broad shallow platforms.
Shelf waters have suffered habitat alteration and changes in biodiversity due to overfishing and the extensive physical damage caused by the deployment of mobile fishing gear (Graham, 1955; Hutchings, 1990). Regional "hot spots" include dramatic shifts in community structure in northeast American shelf fish communities (NOAA, 1992) and the Eastern Bering Sea shelf, where a critical environmental concern is the decline in abundance of certain marine mammal and seabird populations (Alverson, 1992; Pascual and Adkison, 1994), with the Steller sea lion being particularly threatened. Increasing stresses in offshore systems from inshore pollution are of escalating concern but remain poorly understood. Other stresses arise from oil spilled in the sea during tanker transport and operations (NRC, 1985), and to a lesser extent, from oil and gas exploration on the "outer continental shelf," the subject of extensive environmental impact studies since the 1960s (NRC, 1985; Boesch and Rabalais, 1987).
The seemingly distant and more immune slope waters are no longer far away and no longer immune. Deep-water fisheries—and their attendant physical effects (e.g., habitat alteration due to dredging and trawling)—have entered slope waters (Robertson and Grimes, 1983; Polovina and Ralston, 1986). Deep oil and
gas mining will soon commence on slopes at and below 2,000 meters in the Gulf of Mexico (R. Carney, pers. comm., 1994).
Shelf-slope systems thus provide abundant opportunity for examining the impact of anthropogenic activities—of which fisheries-related effects have been and are the most conspicuous—within a regional-scale context. The extent of spatial heterogeneity of shelf-slope systems requires particular attention, striking as it does at the heart of assumptions in ocean management about seemingly "large and monotonous" habitat systems. In particular, research approaches should be undertaken that examine the contribution of several different areas of the shelf to the recruitment of key species, specifically incorporating new techniques for larval tracking (L.A. Levin, 1990; L.A. Levin et al., 1993; see also Boxes 11 and 12). Shelf waters also provide opportunities to study multiple steady states in community composition, the reversibility of diversity shifts, and how long such reversals could or do take, species redundancy, and altered energy budgets.
Pelagic Open Ocean
The open oceans of the world cover nearly three-quarters of the Earth's surface and have been studied for well over 100 years; yet, there are continuing discoveries of new higher taxa and new communities. These include a recently discovered group of microorganisms that in some areas may contribute almost as much to oceanic primary production as all previously known primary producers (Fig. 6; Chisholm et al., 1988, 1992; R.J. Olson et al., 1990; S.W. Chisholm, pers. comm., 1994). The pelagic ecosystems that contain the highest diversity are the epipelagic and mesopelagic strata between the surface and approximately 1,000 meters, within which vertically migrating plankton and micronekton provide active biological linkages.
The public's view of the open sea as a vast homogeneous body of water is belied by the complexity of oceanic subsystems: fronts and eddies, upwelling and downwelling regions, boundary currents, and large ocean gyres (Longhurst, 1981). Such subsystems offer exciting opportunities for the study of biodiversity patterns and the processes that maintain them (Angel, 1993). In regions of intensive fishing pressure over large spatial and temporal scales, for example, there are unique opportunities to investigate "top-down" controls where sufficient historical data are available (discussed in Chapter 6, "Retrospective Analysis: Importance of an Historical Perspective").
Unlike other marine and terrestrial ecosystems, the pelagic realm is not dominated by substrate-related processes. Thus, the ecosystems of the open oceans provide a test of the generality of paradigms developed elsewhere, the resolutions of which are essential for a balanced understanding of marine biodiversity. For instance, studies from some substrate-dominated ecosystems have led to the paradigm that biodiversity is directly related to environmental hetero-
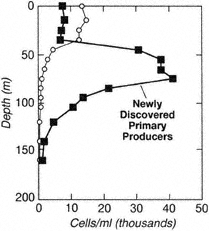
Figure 6
Total numbers of prochlorophytes (squares), a newly discovered group of marine, free-living, primary producers in the picoplankton, greatly exceeded those of the other group of primary producers in this size range, the cyanobacteria or blue-green algae (circles), in samples from the euphotic zone off of southern California. Source: Chisholm et al. (1988)
geneity (e.g., Tilman and Pacala, 1993). Yet, in the open ocean, the most diverse communities occur in environments of very low physical variability (McGowan and Walker, 1993). Likewise, studies of intertidal and coral reef ecosystems have shown the importance of predation in maintaining the diversity of the prey species ("top-down" control) (e.g., Paine, 1980, 1992; Jackson, 1994). This has not been demonstrated in open-ocean ecosystems, which are usually assumed to be controlled by nutrient input ("bottom-up" control). In fact, some of the most exciting recent, large-scale experimental research in the oceans has been on the role of iron as a limiting element controlling primary productivity (Ditullio et al., 1993; Martin et al., 1994).
The fluid nature of pelagic environments shifts natural variability towards slowly developing, long-lived fluctuations. Many biological interactions occur over short temporal scales which, when coupled with the large spatial extent of many pelagic environments, result in subsystems that are similar in all but a few scales of environmental processes. Comparisons between such systems can provide information akin to that derived from very large manipulative experiments. For instance, the eastern and western basins of the North Pacific central gyres may be contrasted to examine the influence of increased mesoscale variability and consequent nutrient input on the biodiversity of the western basin. Comparison between the subarctic gyres of the Atlantic and Pacific may show how the depth of winter mixing, which is known to influence the composition of the primary producers, is transmitted through the ecosystem—another scale of "bottom-up" control.
Many open-ocean ecosystems have, however, been less influenced by recent anthropogenic changes than have their coastal counterparts. Because of their relatively pristine nature and the relatively low levels of natural short-term fluctuations, these pelagic systems potentially could provide some of the first oppor-
tunities to evaluate ecosystem responses to the slowest, but most pervasive, effects of human activities, such as global warming, increased ultraviolet radiation, and the gradual accumulation of pollutants. In order to recognize, and ultimately to predict, such responses, it will be necessary to identify the functionally significant organisms in the pelagic ecosystems and to establish their responses to the anticipated anthropogenic agents. Therefore the open ocean should provide excellent opportunities to relate biodiversity concerns, traditionally the domain of population biologists and taxonomists, to issues of ecosystem organization and function.
Deep Sea
"The frequency with which the deep ocean is popping up as a fanciful solution to waste and resource problems is extremely worrisome. I see four origins for this trend. First, it is the product of our success in defending better known habitats like reefs and bays. Second, it is the persistent belief among nonbiologists that the abyss is the desert proposed by Issacs (seep and vent communities are still referred to as oases). Third, the distant deep ocean is the ultimate 'not in my backyard.' Finally, the vastness of the abyss gives rise to a wasteland management policy which views insults to relatively small areas as fully acceptable."
R. Carney (pers. comm., 1994)
The number of species contained within the deep sea—the least-studied marine habitat—has been a subject of recent, intensive debate (J.F. Grassle and Maciolek, 1992; May, 1992; Poore and Wilson, 1993). There is no question, however, that the previous notion of a global deep-sea bottom that is uniformly featureless has been shattered over the last two decades by countless discoveries of unique, sometimes bizarre, and highly diverse deep-sea communities (e.g., Butman and Carlton, in press). Each major ocean basin has a distinctive fauna, and bottom assemblages vary according to latitudinal gradients (Rex et al., 1993) and topographic features such as basins, canyons, and areas of strong currents (J.F. Grassle, 1989). Discrete, ephemeral patches of food (Billett et al., 1983; Wolff, 1979; Suchanek et al., 1985; Gooday and Turley, 1990) or biogenic structure (C.R. Smith et al., 1986; Thistle and Eckman, 1990), and defaunated patches produced by disturbance (C.R. Smith and Hessler, 1987; Snelgrove et al., 1994), are common and introduce a measure of small-scale spatial heterogeneity. At larger spatial scales, disturbances generated by upwelling regions, episodic strong currents (bottom boundary currents, canyons), slumping (trenches or steep slopes) or heavy sedimentation (e.g., Mississippi plume, or offshore of Cape Hatteras) introduce large-scale patterns (greater than 1,000 square kilometers) that appear to obliterate the patterns of discrete, widely separated, ephemeral patches (J.F.
Grassle, 1989; Schaff et al., 1992; Blake and J.F. Grassle, in press) or gradual community changes with depth (Carney et al., 1983) and sediment grain-size diversity (Etter and J.F. Grassle, 1992).
Although long considered inaccessible and difficult to sample, better quantitative sampling and improved methods of experimentation are continually improving the ability to study the deep-sea biota (see Box 12). The surprisingly high diversity of benthic invertebrates in the deep sea—hundreds of species co-occurring within a square meter of ocean floor—provides a remarkable platform from which to renew empirical, experimental, and theoretical attempts to explain the causes and patterns of global diversity (Rex et al., 1993).
Although the deep sea is vast and remote, humans are still having an effect on this environment. A small, steady increase in abyssal temperature of 0.32°C in 35 years has been attributed to global climate change (Parrilla et al., 1994). Increases in pollutants from atmospheric sources have been observed on the deep-sea floor (La Flamme and Hites, 1978; Takada et al., 1994). Ocean dumping (waste disposal) and deep-sea mining are clear causes for concern. Changes in upper-water column trophic structure—directly or indirectly, for example, through pollution, overfishing, or global climate change—potentially affect deep-sea trophic dynamics (e.g., see Box 1). Because time-series information on biological diversity is unavailable for any deep-sea area, unknown changes currently may be in progress, and thus, the deepest oceans of the world are not immune to human actions. Underscoring these actual and potential changes is the concern that anthropogenic effects on deep-sea communities may be especially severe because deep-sea organisms may be more poorly adapted to change (J.F. Grassle et al., 1990).
Increasing technological sophistication for studying the deep-sea environment (see Box 12) will permit biodiversity issues to be examined experimentally at traditional, necessarily small spatial scales (cm2 to m2). Deep-sea mining technologies and waste-disposal experiments can be designed to produce well-replicated and well-controlled experiments at scales of tens to hundreds of km2. The integration of these approaches should promote better understanding of how and which processes generate and maintain biodiversity in a global environment containing one of the richest species complements.











