The Chemistry of Sexual Selection
THOMAS EISNER AND JERROLD MEINWALD
Chemical dependencies are fundamental in nature. Animals as a group depend on plants for that most basic of metabolites, glucose, which plants produce through photosynthesis. Countless organisms, ourselves included, have vitamin and other dietary requirements. Insects synthesize steroidal hormones but can produce these only from other steroids, such as cholesterol, which they need to obtain with the diet (1). In recent years there has been considerable interest in animals, mostly insects, that depend on exogenous compounds for defense (2). The strategy has human parallels. Many of the medicinals we use against parasites and pathogens are obtained from nature. In insects, the acquired compounds protect primarily against predators. The monarch butterfly, for instance, is distasteful to birds by virtue of cardenolides that it sequesters from its larval foodplants (3).
We here summarize work we have done with a moth, Utetheisa ornatrix, that has a dependence on certain plant alkaloids. The moth uses the compounds for defense and for production of a pheromone that plays a decisive role in sexual selection. The species has a broad range, extending through North America east of the Rockies and southward into Brazil, Argentina, and Chile. Our studies were done mostly with populations of the moth from central Florida.
Thomas Eisner is Schurman Professor of Biology and director of the Cornell Institute for Research in Chemical Ecology and Jerrold Meinwald is Goldwin Smith Professor of Chemistry at Cornell University, Ithaca, New York.
DEFENSE
Utetheisa, like many other Lepidoptera of the family Arctiidae, is aposematic. White, with pink hindwings and black and yellow markings (Figure 1A), it is highly conspicuous on the wing. It flies as readily in daytime as at night. We suspected the moth to be unpalatable, and we were able to prove this in experiments with orb-weaving spiders.
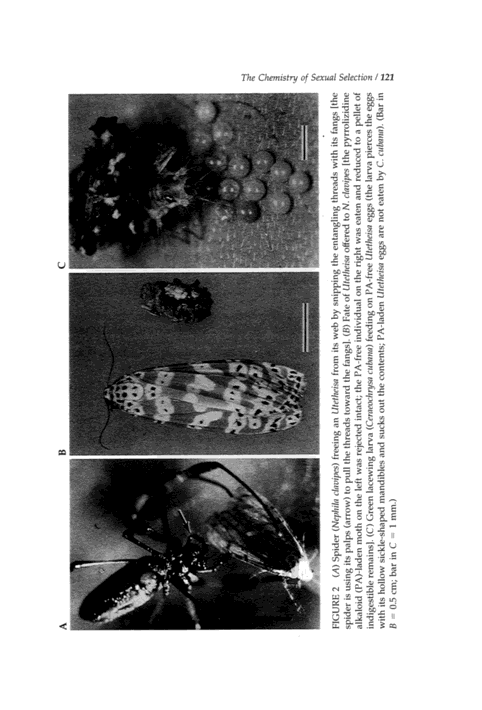
We knew from previous work that moths are protected from entanglement in spider webs by their investiture of scales. Instead of sticking to webs as "naked" insects typically do, they simply lose scales to points of contact with the orb and flutter loose (4). Utetheisa, in contrast, becomes instantly quiescent when it flies into a web. The spider converges on the moth and inspects it, but then, almost invariably, sets it free. During the inspection the moth sometimes emits its defensive froth (Figure 1C), but it is released even if it withholds the effluent. Spiders such as Nephila clavipes cut the moth from the web by snipping the entangling threads with their fangs (5, 6) (Figure 2A), while others, such as Argiope florida, free the moth by pulling it from the web (T.E., unpublished observations).
We knew Utetheisa to feed on poisonous plants as a larva (Figure 1B). The plants, of the genus Crotalaria (family Leguminosae), were known to contain pyrrolizidine alkaloids (henceforth abbreviated as PAs), intensely bitter compounds potently hepatotoxic to mammals (7). Other species of Utetheisa were known to sequester PAs (8). We found this to be true for U. ornatrix as well. Adult Utetheisa raised on Crotalaria spectabilis, one of the principal foodplants available to the moth in the United States, contain on average about 700 µg of monocrotaline (1), the principal PA in that plant (9, 10).
In the laboratory, we succeeded in raising Utetheisa on two alternative artificial diets, one made up with Crotalaria seeds and containing the PA monocrotaline (CS diet), the other based on pinto beans and devoid of PA (PB diet). On the assumption that the PB diet-raised moths, which we proved to be PA-free, would be vulnerable to predation, we took moths

A

B

C
FIGURE 2 (A) Spider (Nephila clavipes) freeing an Utetheisa from its web by snipping the entangling threads with its fangs [the spider is using its palps (arrow) to pull the threads toward the fangs]. (B) Fate of Utetheisa offered to N. clavipes [the pyrrolizidine alkaloid (PA)-laden moth on the left was rejected intact; the PA-free individual on the right was eaten and reduced to a pellet of indigestible remains]. (C) Green lacewing larva (Ceraeochrysa cubana) feeding on PA-free Utetheisa eggs (the larva pierces the eggs with its hollow sickle-shaped mandibles and sucks out the contents; PA-laden Utetheisa eggs are not eaten by C. cubana). (Bar in B = 0.5 cm; bar in C = 1 mm.)
from both cultures into the field and offered them to N. clavipes. The spiders consistently freed the moths raised on CS diet [whose PA content matched that of field-raised Utetheisa (10)] but killed and consumed the PA-free controls (Figure 2B). We also tested directly for the deterrency of PA. We added crystalline monocrotaline to edible items (mealworms) ordinarily consumed by N. clavipes and found that by doing so we could render such items relatively unacceptable to the spiders (6).
Adult Utetheisa tend to be rejected also by birds (blue jays, Cyanocitta cristata; scrub jays, Aphelocoma coerulescens; T.E., unpublished observations), as might be expected, given their aposematism, but there is no definitive evidence that the unacceptability is due specifically to the PAs.
Tests with larval Utetheisa showed these to be rejected by wolf spiders, but only if the larvae had fed on Crotalaria or CS diet. Larvae raised on PB diet proved consistently palatable to the spiders (11).
COURTSHIP
Utetheisa, like many other insects, court at dusk (Figure 1D). The female initiates the behavior. Positioned on vegetation (often on branches of Crotalaria itself), she emits a sex attractant that drifts downwind and lures males (12). Caged virgin females, placed outdoors, attract males (Figure 3A). The glands that produce the pheromone are a pair of long, coiled tubes, opening close together near the abdominal tip (Figure 3C). Extraction of the glands led to the characterization of a long-chain polyene (3), which proved electrophysiologically active on male antennae
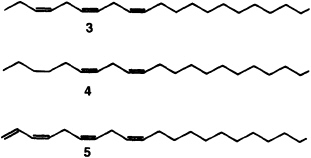
[electroantennogram (EAG) tests] and attractive to males in the field (12) (Figure 3B). The pheromone has since been found to contain two additional polyenes (4 and 5) (13).
We found female Utetheisa to emit their attractant in pulses (1.5 ± 0.2 pulses per s) (12) (Figure 3D), as some other moths have since also been shown to do (14, 15). Pulsation frequencies are similar in the various
species, indicating that there is no informational specificity associated with the temporal modulation of the signals. Views differ as to the function of pheromone pulsation in moths (12, 14-17). A reasonable suggestion, based on mathematical modeling, is that it increases the efficiency of signal output. By pulsing, females can presumably increase the range over which they are able to attract males from downwind (18).
SEXUAL SELECTION
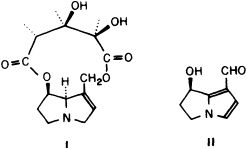
Courtship in Utetheisa involves more than mere attraction of males by females. Once a male locates a female, the pair does not at once proceed to mate. The male first flutters around the female, hovering beside her or circling her at close range, while at the same time occasionally thrusting his abdomen against her. It is only after one or more such thrusts that the female parts her wings and presents her abdomen for mating. We analyzed this behavior, which takes place in darkness, from videotapes taken under infrared illumination (19). We noted that when the male executes his abdominal thrusts, he everts from the abdominal tip a pair of brushes that he ordinarily keeps tucked away in pouches (Figure 1 E and F). The brushes, called coremata, are secretory. Each consists of a tuft of modified scales, associated with glandular tissue at the base, and wetted by secretion. Extraction of the brushes revealed presence of a compound, hydroxydanaidal (HD; 2) (19), previously found in coremata of other Utetheisa species (20). The structure of HD suggested that the compound was derived from PA. We predicted that Utetheisa raised on PA-free diet should have HD-free coremata, which turned out to be the case (19).
We showed that corematal excision rendered males less acceptable to females (mock-operated males were not thus handicapped). This proved that the coremata had a function, but did not in itself provide evidence for the role of HD. However, we found that PB diet-raised males, whose coremata were normal except for lack of HD, were also relatively unsuccessful in courtship (19).
Further data showed HD to have a direct stimulatory effect on the female. When quiescent females were stroked with excised coremata, they tended to present their abdomen, but only if the coremata contained HD. Moreover, to elicit maximal effect, the HD had to be of the absolute configuration [R(-)] in which the compound occurs in the coremata (19). Predictably, female Utetheisa were found to have antennal chemoreceptors highly sensitive to the R(-) isomer of HD (21).
We wondered about the nature of the corematal message. Was HD simply the male's way of announcing his presence to the female, or was the molecule conveying more subtle information? Specifically, we asked whether the derivation of a pheromone from phytotoxin might have
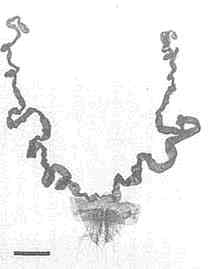
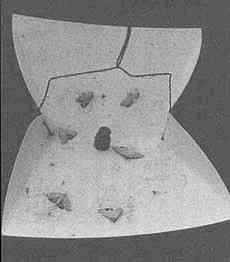
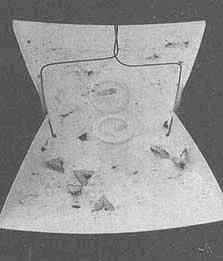

FIGURE 3 Male attraction in Utetheisa. (A) Virgin females, confined to small screened cages affixed to a tray placed outdoors, have attracted a number of males at dusk (the males have become trapped in the sticky coating of the tray). (B) Test comparable to the preceding one, but using synthetic compound 3 of the female's pheromone in lieu of females (the compound is in rubber cup in center of tray). (C) Glandular source of female pheromone (the glands, ordinarily more tightly coiled, are in posterior part of the female's abdomen). (D) Simulation of pulse emission pattern of female pheromone [a fluidics system has been used to generate a pulsed, visibly marked (titanium tetrachloride), plume of air]. (Bar in C = 0.5 mm.)
special adaptive significance. We postulated that HD could provide the male with a means for proclaiming his PA content, a parameter that could be variable, and which could provide the female with a basis for exercising mate choice (19, 22).
We knew that larval Utetheisa, in their later instars, feed predominantly on the seeds of Crotalaria, the parts of the plant richest in PA. We knew further that PA was a strong phagostimulant that drove larvae in their quest for food (10, 23). Moreover, field observation had told us that the seeds of Crotalaria were a variable resource, for which the larvae might need to compete at times. We also knew that adult Utetheisa differed substantially in their PA load in nature and that under experimental conditions their PA load varied in proportion to the seed content of the larval diet (9, 24). It remained to be seen whether the male's HD content was an indicator of his PA content. Chemical analyses proved that it was (24). Our postulate that HD could provide the female with a means of assessing the male's PA load, as well as possibly his larval competitive ability, a potentially heritable trait, was strengthened. But, as we were to learn, we were not being imaginative enough.
PARENTAL INVESTMENT
When we analyzed Utetheisa eggs, we discovered that they too contain PA, suggesting that they were being endowed, for their chemical protection, by the mother. However, we determined that eggs could contain PA even when the mother was herself PA-free. All that was necessary was for the father to be PA-laden. These results, together with others, established that the male transfers PA to the female at mating, and that both parents bestow PA upon the eggs (25). The allocation is uneven. By mating males and females laden with different PAs (usaramine and monocrotaline, respectively) and analyzing the eggs for both PAs, we were able to determine that the eggs receive only about a third of their PA from the father (25). However, these data pertained to once-mated females, when in fact female Utetheisa mate with a number of males over their life-span (mean number = 4-5 males; more than 10 partners have been recorded) (26). On average, therefore, eggs might receive more than a third of their PA from males.
The eggs are effectively protected by their alkaloidal endowment. Both coccinellid beetles and ants are deterred by PA dosages commensurate with levels naturally prevailing in Utetheisa eggs (Utetheisa eggs contain an average 0.8 µg of PA). With both predators, PA proved most effective as the N-oxide, the predominant form in which PA occurs in Utetheisa (25, 27). Also deterred were larvae of green lacewings (Ceraeochrysa cubana), which rejected eggs from moths raised on CS diet while accepting those
from individuals raised on PB diet (T.E., unpublished observations) (Figure 2C).
We were forced to review our interpretation of the corematal message. HD, it seemed, could serve not only for proclamation of alkaloid load and of a genetic capacity, but for advertisement of a nuptial gift. We postulated that the magnitude of the male's PA offering should be proportional to his PA load, and we found this to be the case (24).
We determined further that in nature male PA content is proportional to male mass (9, 26), indicating that by favoring males of high PA content females could be selecting also for males of large size. Larger males transfer larger spermatophores, and thus more nutrient for investment in eggs (with each mating beyond the first, the female is able to produce, on average, an extra 32 eggs, an equivalent of upward of 10% of her basic output) (26, 28). The advertisement implicit in the corematal signal could thus be for both PA and nutrient (26).
The correlation of large size and PA load in Utetheisa could have a simple reason. Crotalaria seeds are rich not only in PA but in nutrient. Larvae competing successfully for seeds could therefore inevitably be destined to achieve large adult size.
SPERM SELECTION
A question of interest concerned the paternity of Utetheisa offspring. Given that females are promiscuous, do males have assurance of fathering offspring when they mate? Is there sperm mixing in multiply mated females, or do the sperm of some males ''win out" over those of others?
By means of enzymatic markers we checked into offspring paternity of twice-mated females. We found the progeny in most cases to be sired almost exclusively by the larger of the two males. Factors such as duration of copulation, mating order, or between-mating interval were not determinants of male "success." Nor was male PA content, which in our laboratory-raised Utetheisa did not correlate with male size (we attribute this lack of correlation to the fact that our artificial larval diets were equally nutritious, whether PA-laden or PA-free) (26).
We have evidence that the female herself controls the mechanism by which one set of sperm is favored over the other. If females are anesthetized so as to inactivate their muscles, including presumably the many pumping muscles of their reproductive organs, the normal routing of sperm is inhibited. The anesthesia does not immobilize the sperm, indicating that it is not the sperm themselves that are in control of their fate (26). The reproductive system of female Utetheisa is a complex labyrinth of ducts and chambers. While we do not know precisely how
the various components of the system operate, we have hypotheses on how the female might selectively retain or expel sets of sperm.
We also have evidence of how the female assesses the size of her mating partners. She appears to do so indirectly, by gauging the size of their spermatophore, for which purpose she may use stretch receptors that female moths are known to have in the chamber (bursa) in which spermatophores are deposited (29). Male Utetheisa can be caused to produce inordinately small spermatophores if they are mated relatively recently beforehand. If such mated males are placed in competition with physically smaller males, whose spermatophores may now be the relatively larger ones, they tend to "lose out" (30). We predict from this that males, in nature, may space their matings days apart. To regain the capacity to produce full size spermatophores takes a male about a week (30).
The female strategy is an interesting one. By accepting multiple partners she can accrue multiple gifts, to her obvious benefit, since she can thereby promote both her fecundity and the survivorship of her offspring. By discriminating between sperm, she is able to select for traits that in the genetic sense have long-range payoff. By favoring sperm of large males the female is essentially reinforcing, after copulation, the choice mechanism that she already exercised in the precopulatory context. Postcopulatory assessment provides the female with the option of taking corrective action. If on a given evening she accepted a male of moderate size and PA content, she can still discriminate genetically against that male by utilizing the sperm of a larger, more PA-laden and therefore genetically superior individual, that she is able to lure on a subsequent night. But the earlier mating is canceled in a genetic sense only. Nutrient and PA that the female receives from the losing male are utilized by her, as are all gifts that she obtains from males (26, 28).
We feel that we may have a tentative answer to the question of why smaller males, of lesser PA content, appear not to "lie" in the context of courtship. Could such males not masquerade as "desirable" by producing exaggerated levels of HD? Perhaps their failure to do so is a reflection of the fact that they would be "found out" unless they also produced outsized spermatophores. Smaller males, even if able to convert extra PA into HD for inflation of their chemical message, may lack the extra nutrient needed for inflation of the spermatophore. By putting the male to the test by way of a second criterion after mating, the female has the means to check on liars.
ADDITIONAL FINDINGS
We discovered in the laboratory that Utetheisa larvae deficient in PA can make up their chemical shortfall by resorting to cannibalism (10, 23).
They attack both pupae and eggs, and they appear to be driven to cannibalism not so much by hunger as by the PA deficiency itself. Moreover, they target specifically eggs and pupae that are PA-laden rather than PA-free. Possession of high systemic PA loads could therefore, under some circumstances, be endangering to Utetheisa, rather than beneficial. We do not know whether in nature Utetheisa are ever seriously at risk from cannibalism, although it is interesting to note that Utetheisa pupate out of reach of larval attack, in secluded sites away from the foodplant (23). The danger may be real for eggs, however, which are laid on the leaves of Crotalaria and therefore exposed to larvae.
A further finding concerns the stereochemistry of HD and its derivation from PA. Both HD and the primary PAs (monocrotaline, usaramine) that we know to be available to Utetheisa in the field are of the same (7R) stereochemical configuration. It was therefore not surprising to find that Utetheisa is unable to convert a PA of opposite (7S) stereochemistry (heliotrine) into HD. However, we found another arctiid moth, the Asian species Creatonotus transiens, which also produces HD in its coremata, to be able to use 7R and 7S PAs interchangeably for HD production (31). We are tempted to conclude that Creatonotus, unlike Utetheisa, has dietary access to PAs of both stereochemical configurations in its environment.
THE BROADER PERSPECTIVE
Parental bestowment of defensive substances upon eggs may be more widespread in insects than generally suspected. Insect eggs, by virtue of immobility alone, are highly vulnerable, and it makes sense that they should be protected. Utetheisa's strategy of utilizing substances of exogenous origin for egg defense is not without parallel. Nor is the strategy of paternal involvement in the provisioning process.
In Apiomerus flaviventris, an assassin bug (family Reduviidae), the female alone provisions the eggs. She procures a terpenoid resin from plants and applies this to the eggs, thereby protecting these from ants (32). Blister beetles (family Meloidae) protect their eggs with cantharidin. The compound is biosynthesized by the beetles, sometimes by the males alone, which transfer it to the females, for incorporation into the eggs (33). Cantharidin is also utilized by Neopyrochroa flabellata, a fire-colored beetle (family Pyrochroidae). In this insect, the cantharidin is procured by the male from an unknown exogenous source and is also transmitted by way of the female to the eggs. Interestingly, the courting male advertises his possession of cantharidin by exuding a small amount of the compound as secretion from a cephalic gland. The female feeds on this secretion prior to mating and rejects males unable to provide such proof of "worth" (31).
The closest parallel to the Utetheisa strategy is exhibited by danaine butterflies (family Nymphalidae). In one of these, the queen butterfly (Danaus gilippus), which we have studied in detail, the adult male visits plants that produce PA, and he ingests PA from these sources. He then transfers the PA to the female at mating, and she bestows virtually the entire gift upon the eggs. Remarkably, the male produces a pheromone, danaidone, which he derives from PA, and which he "airs" in the context of courtship by everting two glandular brushes that secrete the compound. Males deficient in danaidone tend to be rejected in courtship (6, 34). Work done on other danaines, as well as on the related ithomiines, suggests that comparable behavior may be widespread in these butterflies (6, 35-38).
Paternal provisioning may also involve bestowal of inorganic compounds upon the eggs. Many butterflies and moths drink extensively at water sources, a phenomenon known as "puddling." The behavior is sex-biased and involves for the most part males. It had long been suspected that the benefit from puddling is sodium sequestration (39), and this has now been demonstrated (40,41). It is also becoming clear that the acquired sodium is transferred in part to the female at mating (40, 41), and that the female transmits the gift to the eggs (41). Whether prior to mating males advertise their ionic merits by proclaiming in some fashion that they are "worth their salt" remains unknown.
SUMMARY
The moth Utetheisa ornatrix (Lepidoptera: Arctiidae) is protected against predation by pyrrolizidine alkaloids that it sequesters as a larva from its foodplants. At mating, the male transfers alkaloid to the female with the spermatophore, a gift that the female supplements with alkaloid of her own and transmits to the eggs. Eggs are protected as a result. The male produces a pheromone, hydroxydanaidal, that he derives from the alkaloid and emits from a pair of extrusible brushes (coremata) during precopulatory interaction with the female. Males rendered experimentally alkaloid-free fail to produce the pheromone and are less successful in courtship. The male produces the pheromone in proportion both to his alkaloid load and to the amount of alkaloid he transfers to the female. The pheromone could thus serve as an indication of male "worth" and provide a basis for female choice. Utetheisa females are promiscuous and therefore are able to accrue multiple nuptial gifts (alkaloid and nutrient, both transmitted with the spermatophore). They use sperm selectively, favoring those of larger males. Larger males in nature are also richer in alkaloid. Females therefore reinforce after copulation the choice mechanism they already exercise during courtship.
We are greatly indebted to the many associates who collaborated in our research on Utetheisa and to the staff of the Archbold Biological Station, Lake Placid, FL, where much of our field work was done. The study was supported largely by National Institutes of Allergy and Infectious Diseases Grants AI02908 and AI12020, by Hatch Grants NYC-191424 and NYC-191425, and by unrestricted funds from the Schering-Plough Research Institute and the Merck Research Laboratories.
REFERENCES
1. Clayton, R. B. (1964) J. Lipid Res. 5, 3-19.
2. Rosenthal, G. A. & Berenbaum, M. R. (1992) Herbivores: Their Interactions with Secondary Plant Metabolites (Academic, New York), Vols. 1 and 2.
3. Brower, L. P., Nelson, C.J., Fink, L. S., Seiber, J. N. & Calhoun, B. (1988) in Chemical Mediation of Coevolution, ed. Spencer, K. C. (Academic, New York), pp. 447-475.
4. Eisner, T., Alsop, R. & Ettershank, G. (1964) Science 146, 1058-1061.
5. Eisner, T. (1982) BioScience 32, 321-326.
6. Eisner, T. & Meinwald, J. (1987) in Pheromone Biochemistry, eds. Prestwich, G. D. & Blumquist, G. J. (Academic, Orlando, FL), pp. 251-269.
7. Mattocks, A. R. (1972) in Phytochemical Ecology, ed. Harborne, J. B. (Academic, New York), pp. 179-200.
8. Rothschild, M. (1973) in Insect/Plant Relationships, ed. van Emden, H. F. (Blackwell, London), pp. 59-83.
9. Conner, W. E., Roach, B., Benedict, E., Meinwald, J. & Eisner, T. (1990) J. Chem. Ecol. 16, 543-552.
10. Bogner, F. & Eisner, T. (1991) J. Chem. Ecol. 17, 2063-2075.
11. Eisner, T. & Eisner, M. (1991) Psyche 98, 111-118.
12. Conner, W. E., Eisner, T., Vander Meer, R. K., Guerrero, A., Ghiringelli, D. & Meinwald, J. (1980) Behav. Ecol. Sociobiol. 7, 55-63.
13. Jain, S. C., Dussourd, D. E., Conner, W. E., Eisner, T., Guerrero, A. & Meinwald, J. (1983) J. Org. Chem. 48, 2226-2270.
14. Schal, C. & Cardé, R. T. (1985) Experientia 41, 1617-1619.
15. Conner, W. E., Webster, R. P. & Itagaki, H. (1985) J. Insect Physiol. 31, 815-820.
16. Conner, W. E. (1985) in Perspectives in Ethology, eds. Klopfer, P. H. & Bateson, P. P. G. (Plenum, New York), Vol. 6, pp. 287-301.
17. Baker, T. C., Willis, M. A., Haynes, K. F. & Phelan, P. H. (1985) Physiol. Entomol. 10, 257-265.
18. Dusenberry, D. B. (1989) J. Chem. Ecol. 15, 971-977.
19. Conner, W. E., Eisner, T., Vander Meer, R. K., Guerrero, A. & Meinwald, J. (1981) Behav. Ecol. Sociobiol. 9, 227-235.
20. Culvenor, C. C. J. & Edgar, J. A. (1972) Experientia 28, 627-628.
21. Grant, A. J., O'Connell, R. J. & Eisner, T. (1989) J. Insect Behav. 2, 371-385.
22. Eisner, T. (1980) in Insect Biology and the Future, eds. Locke, M. & Smith, D. S. (Academic, New York), pp. 847-878.
23. Bogner, F. & Eisner, T. (1992) Experientia 48, 97-102.
24. Dussourd, D. E., Harvis, C. A., Meinwald, J. & Eisner, T. (1991) Proc. Natl. Acad. Sci. USA 88, 9224-9227.
25. Dussourd, D. E., Ubik, K., Harvis, C., Resch, J., Meinwald, J. & Eisner, T. (1988) Proc. Natl. Acad. Sci. USA 85, 5992-5996.
26. LaMunyon, C. W. & Eisner, T. (1993) Proc. Natl. Acad. Sci. USA 90, 4689-4692.
27. Hare, J. F. & Eisner, T. (1993) Oecologia 96, 9-18.
28. LaMunyon, C. W. (1993) Ph.D. dissertation (Cornell Univ., Ithaca, NY).
29. Sugawara, T. (1979) J. Comp. Physiol. 130, 191-199.
30. LaMunyon, C. W. & Eisner, T. (1994) Proc. Natl. Acad. Sci. USA 91, 7081-7084.
31. Schultz, S., Francke, W., Boppré, M., Eisner, T. & Meinwald, J. (1993) Proc. Natl. Acad. Sci. USA 90, 6834-6838.
32. Eisner, T. (1988) Verh. Dtsch. Zool. Ges. 81, 9-17.
33. McCormick, J. P. & Carrel, J. E. (1987) in Pheromone Biochemistry, eds. Prestwich, G. D. & Blumquist, G. J. (Academic, Orlando, FL), pp. 307-350.
34. Dussourd, D. E., Harvis, C. A., Meinwald, J. & Eisner, T. (1989) Experientia 45, 896-898.
35. Ackery, P. R. & Vane-Wright, R. I. (1984) Milkweed Butterflies: Their Cladistics and Biology (Cornell Univ. Press, Ithaca, NY).
36. Brown, K. S. (1984) Nature (London) 309, 707-709.
37. Boppré, M. (1986) Naturwissenschaften 73, 17-26.
38. Schneider, D. (1992) Naturwissenschaften 79, 241-250.
39. Arms, K., Feeney, P. & Lederhouse, R. (1974) Science 185, 372-374.
40. Pivnick, K. A. & McNeil, J. N. (1987) Physiol. Entomol. 12, 461-472.
41. Smedley, S. R. (1993) Ph.D. dissertation (Cornell Univ., Ithaca, NY).
42. Meinwald, J. & Eisner, T. (1995) Proc. Natl. Acad. Sci. USA 92 , 14-18.