
20
Social Facilitation and Inhibition of Eating
John M. de Castro1
Not Eating Enough, 1995
Pp. 373–392. Washington, D.C.
National Academy Press
INTRODUCTION
The human is a social animal whose behavior is profoundly affected by social influences. "Of all the stimulation that impinges on the organism in its lifetime, stimulation from social sources is most important" (Zajonc, 1980, p. 50). Most human behavior is shaped or controlled by social forces. In fact, the act of soldiering itself is primarily a social phenomenon.
The social context can determine which behaviors are emitted. An individual may exhibit quite different behaviors in public than in private. For that matter, an individual's public behavior may vary considerably in different social contexts. A soldier may be quiet and reflective when alone; boisterous, outgoing, and rebellious with buddies; or obedient and retiring in the presence of a superior officer. In addition, these responses may be very different depending on whether a soldier is in the barracks on the base, at a private party off the base, in a church at home, or engaged in a firefight.
Not only which behaviors are emitted but also their magnitude is largely determined by the social context. In general, the presence of other people tends to increase the expression of simple behaviors, such as well-learned motor responses, and it tends to interfere with complex behaviors, such as learning new material or responses (Zajonc, 1980). This effect of social context on the magnitude of behavior is generally referred to as social facilitation or inhibition. Social facilitation of behavior has been defined as "increments in the frequency or intensity of responses already learnt by the individual, shown in the presence of others usually engaged in the same behavior" (Crawford, 1939, p. 432).
In order for social effects to occur, the other individuals must in some way be involved with the subject. This involvement could simply mean that the other individual provides an audience, or the other individual could be a co-actor, actively engaged in a behavior along with the subject. Alternatively, the other individual may be behaving while the subject is the audience. This may evoke modeling or imitation by the subject, or it could provide vicarious learning. The mere presence of another is not enough. If the other person is blindfolded and the subject knows there will be no interaction, then social facilitation does not occur (Cottrell et al., 1968).
SOCIAL FACILITATION OF FOOD INTAKE
Simple behaviors tend to be increased in magnitude by social influences. Because eating is a very simple behavior, it would be expected that more would be eaten when dining occurs with others present. Such an effect has long been known to occur with animals, who eat more in the presence of other animals than when they eat alone. Bayer (1929) demonstrated this phenomenon when he allowed a chicken to completely satiate by eating as much wheat as it wanted. he then introduced a hungry chicken who began to eat. The first chicken, although just satiated, immediately began to eat again. The same phenomenon has been replicated in pigs (Hsia and Wood-Gush, 1984).
The general phenomenon of social facilitation of eating has subsequently been demonstrated in a large variety of species, including chickens (Rajecki et al., 1975; Tolman and Wilson, 1965), fish (Welty, 1934), rats (Harlow, 1932; Hoyenga and Aeschleman, 1969), gerbils (Forkman, 1991), puppies (James, 1960), and primates (Harlow and Yudin, 1933). As an example, the results of the Harlow (1932) study with rats are summarized in Figure 20-1. Harlow fed rats either in pairs or alone on alternate days. As shown in Figure 20-1, the animals always ate more in pairs than when alone.
Even though the animal research clearly demonstrated that social facilitation was a robust phenomenon, it was still believed that somehow people were different. Indeed Harlow (1932, p. 12) wrote "…in the presence of individuals like ourselves…eating is influenced, probably not so much as
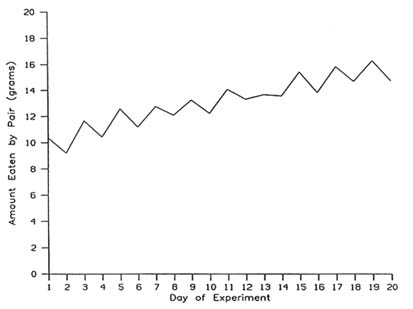
FIGURE 20-1 Mean daily intake (g) by pairs of rats fed on alternating days either together (odd-numbered days) or alone (even-numbered days). SOURCE: Adapted from Harlow (1932).
to quantity as to appreciation. A good meal tastes better if we eat it in company of friends." Food intake by humans was looked upon as fundamentally the different from animals, that is, animals feed and humans dine! Researchers believed that "in man, the time of meals, as well as the choice of foods, their presentation, the cooking style, and the quantities presented…escape the direct control exerted in animals" (Bellisle, 1979, p. 164). As a result, the lively research on social facilitation of food intake in animals was not paralleled by similar research in humans.
Laboratory Studies of Social Facilitation of Eating in Humans
In spite of this thinking, laboratory findings have emerged which suggest that social facilitation also affects the eating behavior of humans. Frank (1944) demonstrated that subjects would eat simply in response to an instruction to eat, even when told that they were participating in an experiment on persuasion and that it did not matter whether they ate or not. Much like Bayer's chickens, humans, when paired with someone who eats a large amount of food, markedly increase their intake. Nisbett and Storms (1972) invited subjects to eat crackers in a taste test. Subjects tasted the crackers alone or paired with a
confederate (model) who ate either 1 or 20 crackers. Normal-weight subjects ate 29 percent less with the low-intake model and 25 percent more with the high-intake model than when alone. Using a similar modeling technique, Conger et al. (1980) found an even larger effect: subjects exposed to a high-intake model ate 86 percent more crackers than those exposed to a low-intake model. In addition they demonstrated a comparable inhibitory effect. Subjects paired with someone who ate no crackers decreased their intake by 42 percent.
Polivy et al. (1979) described a similar modeling effect. Subjects who had fasted for 5 hours were asked to fill themselves with sandwich quarters in preparation for a "taste test." When the subjects were paired with a confederate who ingested eight sandwich quarters, they ate 57 percent more than when the confederate only ate two quarters. The influence of the model persisted into the taste test, with the result that subjects exposed to the high-intake model ate 31 percent more nuts than those exposed to the low-intake model. Using a similar modeling technique, Goldman et al. (1991) demonstrated 50 percent greater intake with a high-intake model than with a low-intake model in subjects who were deprived of food for 24 hours. Hence, in the laboratory, the food intake of a subject can be profoundly influenced by the food intake of a companion even when the subject is extremely hungry.
The snack intake of a subject appears to be affected not only by the intake but also by the nature of the companion. De Luca and Spigelman (1979) had a nonobese or obese model always eat the same amount of candy while filling out a questionnaire. Obese subjects tended to eat more candy when paired with the obese model than when with the nonobese model, and nonobese subjects were unaffected by the model's weight. Furthermore gender also appears to influence a subject's response to eating with a companion. During a "get-acquainted" session in the lab, female subjects ate 75 percent less when accompanied by a desirable male than when accompanied by an undesirable companion (Mori et al., 1987). A comparable effect was not apparent for males. Hence, in the laboratory, the snack intake of a subject can be greatly affected by the nature of a companion.
The amount people eat has been observed to be affected by the mere presence of other people eating with them. In the laboratory, subjects, regardless of gender, ate 94 percent more ice cream in groups than when eating alone (Berry et al., 1985). In more naturalistic settings, subjects' food consumption has also been observed to be affected by the presence of other people. Edelman et al. (1986) compared the amount eaten by subjects in a cafeteria to that ingested in isolated conditions. They found that both obese and nonobese people ate 48 percent more in the cafeteria than when alone. Krantz (1979) performed a naturalistic observation in a university cafeteria of the effect of eating with others on the intake of obese and nonobese subjects. Obese subjects purchased less food when accompanied by others than when alone. In contrast, nonobese subjects did the opposite; they purchased more food when accompanied by companions than when alone.
Social Facilitation of Spontaneous Eating in Free-Living Humans
The laboratory evidence makes a compelling case that social influences can produce strikingly large increases or decreases in the amounts of food ingested. However, this evidence is from studies of adjunctive intake, snacking, or, at best, a single lunch meal. This raises the question whether these results, mainly obtained under artificial conditions, can be generalized to the natural, everyday intake of normal people and whether similar results would be obtained over sustained periods of time.
The Diet Diary Technique
For a number of years this author's laboratory has been using the diet diary technique to measure the spontaneous, natural intake of free-living normal adult humans (de Castro, 1987a, b; 1990; 1991a, b, c; 1993a, b; 1994; de Castro and Brewer, 1992; de Castro and de Castro, 1989; de Castro et al., 1986, 1990; Redd and de Castro, 1992). Seven-day diaries of food intake have been collected from adults, children, adolescents (de Castro and Goldstein, in press), and elderly humans (de Castro, 1993a) eating normally in their natural environment. Subjects record for each meal when it starts and ends, exactly what they eat or drink, their subjective states, and where they eat the meal. Most importantly from the standpoint of investigating social influences on food intake, they also record the number and nature of other people eating with them.
Self-report methods in general, and specifically the diet diary technique, have been believed to be inaccurate and unreliable. Indeed, the 24-h recall procedure for acquiring dietary information has been found to produce fairly inaccurate results (Brown et al., 1990; Dubois and Boivin, 1990; Krantzler et al., 1982; Larkin et al., 1991; Mullenbach et al., 1992; Myers et al., 1988). However, the diary self-report method has important differences in that subjects record every item they either eat or drink at the time it is consumed. This method minimizes the distortion of memory and improves the reporting of details and the estimation of quantities. It has been reported to be a reliable procedure with good agreement being found between separate records collected after a delay as long as 2 years (Adleson, 1960; Block, 1982; Heady, 1961; Livingstone et al., 1990; St. Jeor et al., 1983).
Evidence exists that subjects' self-reports of their intake are truthful reports of the foods they actually ingest. Surreptitious measurements of the actual amount of food consumed at meals have been found to be in close agreement with diary records (Eagles and Longman, 1963; Gersovitz et al., 1978; Krantzler et al., 1982). To check the validity of self-reports, this lab has each subject identify two individuals with whom they will be eating during the recording period. These individuals are later contacted and asked to recall or
verify what the subject ate at the meals where they were present. After 1,015 verifications, in no case has the observer reported that the subject ate something that was not recorded in the diary or reported that the subject did not eat a recorded food.
Evidence also exists that the amounts of food reported in diaries underestimate actual intakes by about 20 percent (Bandini et al., 1990; Goran and Poehlman, 1992; Lissner et al., 1989; Livingstone et al., 1990, 1992; Mertz et al., 1991; Prentice et al., 1986). However, this is only a problem when the absolute amounts of nutrients ingested are required. In most research applications, the quantities ingested by one group or condition are related to those ingested by another group or condition. The error created by underestimation would be expected to affect all subjects equally and thus would not change relative values.
A number of sources of unsystematic error could affect diet diary records of intake. However, these sources of error would be expected to add to error variance and thereby tend to obscure relationships rather than produce systematic differences between intakes associated with different groups or conditions. "Random inaccuracy may lead to false negative conclusions…by reducing true associations but will not generate misleading correlations" (Livingstone et al., 1990, p. 708). That significant systematic and subtle relationships have been discerned in many prior projects (de Castro, 1987a, b; 1990; 1991a, b, c; 1992; 1993a, b; 1994; de Castro and Brewer, 1992; de Castro and de Castro, 1989; de Castro and Elmore, 1988; de Castro and Goldstein, in press; de Castro and Kreitzman, 1985; de Castro et al., 1986, 1990) in spite of the error attests to the robustness of the phenomena observed and indicates that the technique is sensitive enough for most research purposes.
The Presence of other People at Meals
To investigate whether social facilitation influences people's intake outside of the laboratory, this author reanalyzed the diet diary data that were collected in prior studies (de Castro, 1987a, b; 1990; 1991a, b, c; 1993a, b; 1994; de Castro and Brewer, 1992; de Castro and de Castro, 1989; de Castro et al., 1986, 1990). The amounts ingested in meals eaten alone versus those eaten with other people present were reviewed. Results showed that the meals eaten with other people present were on the average 44 percent larger than the meals eaten alone (de Castro and de Castro, 1989) and included larger amounts of carbohydrate, fat, protein, and alcohol. In addition, this result was found to be an orderly phenomenon, the number of people present had a significant positive correlation with the amount eaten in the meal (Figure 20-2). This phenomenon, called a social correlation, indicates that the more people that are present at the meal, the more will be eaten (Figure 20-2, left panel). In addition, the correlation was equivalent even when meals eaten alone were
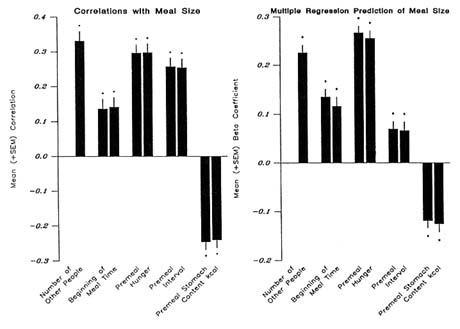
FIGURE 20-2 Mean ± SEM Z transformed correlation coefficients (left panel) and mean ± SEM ß coefficients (right panel) from the multiple linear regressions predicting the amount ingested in the meals on the basis of the number of other people present at the meal, the minute of the day that the meal was begun, the self-rating of hunger at the start of the meal, the time since the last meal, and the estimated contents of the stomach at the start of the meal. The first bar of each pair represents the multiple regressions containing the number of people as a predictor, and the second bar represents the regressions without the number of people included. indicates that the mean is significantly (P <0.05) different from zero as assessed with a t test.
excluded. Hence, not only are meals eaten with other people larger than meals eaten alone, but the greater the number of people present, the more that is ingested.
To investigate whether the influence of the presence of other people was primary or secondary to other salient influences on meal size, multiple linear regressions were performed that used the number of people present as a predictor of meal size along with four other predictors that are known to be related to the amount eaten at a meal: time of day (de Castro, 1987b), hunger self-ratings (de Castro and Elmore, 1988), premeal interval (de Castro and Kreitzman, 1985), and premeal estimated stomach content (de Castro et al., 1986). Results of these regressions were compared to similar regressions that did not use the number of people present as a predictor. The mean univariate correlations and the ß (standardized regression) coefficients from these
analyses are presented in Figure 20-2. The four-factor multiple regression without the number of people present accounted for 40.7 percent of the variance in meal size. When the number of people was added, the regression accounted for 52.4 percent of the variance. Taking into consideration social facilitation did not alter the influence of any of the other four factors. The ß coefficients for these factors were the same regardless of whether or not the number of people was included. Hence, the presence of other people would appear to have a major effect on food intake, which is independent of other salient influences.
Social Correlation as an Artifact
It is possible that this relationship between the presence of other people and meal size is artifactual. The positive correlation could result from a covariation produced by a third factor, the time of day. Breakfast is the smallest meal of the day and may be eaten with the fewest other people present, while dinner is the largest meal and may have the greatest number of other people present. The social correlation may also occur as a result of alcohol intake, which may increase the caloric content of meals eaten with other people. Additionally, snacks are small and generally eaten alone, while meals are in general larger and more likely to be eaten with others. Another possible explanation for the correlations is that meals eaten in restaurants may be larger and eaten with more other people than meals eaten at home, which, in turn, may be larger and more social in nature than meals eaten elsewhere. Still another possibility is that meals eaten on weekends may be larger and eaten with more other people present than are meals eaten on weekdays.
This author investigated these potential artifactual explanations by isolating meals that occurred under specific conditions and demonstrated that, although the covariances exist, they do not account for the social correlation. Strong, positive, and significant correlations between meal size and the number of other people present—social correlations—were found separately for meals eaten during breakfast, lunch, or dinner; eaten in restaurants, at home, or elsewhere; accompanied by alcohol intake or without alcohol; eaten only as snacks or only as meals (de Castro et al., 1990); or eaten as meals during weekdays or during weekends (de Castro, 1991a). This finding suggests that the social correlation results from a true social facilitation of eating and is not an artifact of time, place, alcohol, or snacks. This facilitation is an important determinant of eating regardless of whether alcohol is ingested with the meal, whether the food is consumed as a snack or a meal, and when or where the food is eaten.
The Orderly Relationship of Social Facilitation to Intake
To look more systematically at this relationship, meals eaten alone or with one, two, three, four, five, six, or seven or more other people were separated, and average meal sizes were calculated (de Castro and Brewer, 1992). As seen in Figure 20-3, there is an orderly relationship between the number of people present and meal size. One other person present at the meal was associated with a 33 percent increase in meal size while 47 percent, 58 percent, 69 percent, 70 percent, 72 percent, and 96 percent increases were associated with two, three, four, five, six, and seven or more people, respectively. The size of the effect is remarkable. The magnitude of these differences is much larger than those obtained in prior research with physiological (de Castro, 1987a; de Castro and Elmore, 1988), age (de Castro, 1993b), circadian (de Castro, 1987a), seasonal (de Castro, 1991b), or psychological (de Castro and Elmore, 1988) variables, which suggests that the most salient factor associated with short-term food intake in humans is social facilitation.
The orderliness of the effect is also remarkable. It has been shown with many human behaviors that social facilitation can in general be adequately described by a power function (Latane, 1981). De Castro and Brewer (1992) found this to be true for social facilitation of meal size, which can be best
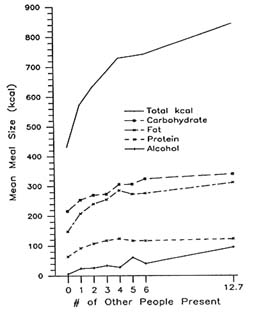
FIGURE 20-3 Mean meal size in total kcal (solid) or kcal attributable to carbohydrate (0), fat (*), protein (X), or alcohol (+) intake, as a function of the number of people present at the meal.
represented by the power function, Meal Size = 485 N23, where N equals the total number of people present including the subject. Hence social facilitation of food intake is an orderly, lawful process that is similar to other social facilitation phenomena and likewise can be fairly well described by a power function.
Social Facilitation and Increased Intake
Even with all this evidence, because of the observational nature of the research, it is not acceptable to conclude that the presence of other people is the cause of the increased intake. To establish causation, this laboratory actively manipulated the number of other people present by instructing subjects to eat only by themselves for a 5-d period, to eat normally for another 5-d period, and to eat only with other people for a third 5-d period. The order of these periods was randomized. In comparison to the normal instruction period, the subjects ingested on average 212 kcal, 11 percent, less per day when they were instructed to eat alone (Redd and de Castro, 1992). This result suggests that the presence of other people is indeed the cause of the increase in intake at meals.
In contrast to the ''eat alone" instruction, when instructed to eat all meals socially, the subject did not eat significantly more overall or significantly larger meals than normal. This lack of an influence of increasing the number of people present at meals can be explained on the basis of the power function relationship between the number of people present and the amount ingested. There were on average 1.3 people present at the meals under the normal condition. In contrast, there were on average 1.7 people present at meals in the "eat socially" instruction condition. As is apparent from Figure 20-3, an increase from 1.3 to 1.7 people would not be expected to produce much of an increase in intake. This suggests that social facilitation may be of limited use in attempting to increase intake, unless the individual eats primarily alone or unless large numbers of other people can be added to the individual's eating context.
Note that the magnitude of the effect of eating alone, 11 percent, is considerably smaller than the magnitude of the social facilitation effects observed in unmanipulated contexts as reported above. In fact, the meal sizes reported during the manipulated alone condition were 20 percent larger than the alone meals during the normal condition (Redd and de Castro, 1992). This result might indicate that separating naturally occurring meals that just happen to be eaten alone from those that happen to be eaten with others may overestimate the impact of social facilitation of eating. Alternatively it might suggest that subjects compensate, increasing meal size in the alone condition to bring overall intake to more nearly normal levels. Research that looks at
when people eat alone for periods longer than 5 days is needed to address this issue.
THEORIES OF SOCIAL FACILITATION OF EATING
Research to date has demonstrated that social facilitation is a ubiquitous and salient influence on food intake that operates in the real everyday environments of normal people causing increased consumption of nutrients. Of all the myriad of stimuli that affect the ad libitum food intake of humans, social facilitation is the most powerful yet to be discovered (de Castro and de Castro, 1989). The question remains as to how the presence of other people may operate on the individual to influence the amount eaten in a meal. There are a number of theoretical explanations.
Social facilitation of eating may increase intake by producing imitation (Tolman, 1968) or by calling an individual's attention to food by observing a companion's activities. This hypothesis suggests that an individual alters intake in the direction of a companion's intake, eating more with a high-intake companion and less with a low-intake companion. Indeed, subjects in the presence of a high-intake model tend to eat large amounts and in the presence of a low-intake model eat very little (Conger et al., 1980; Nisbett and Storms, 1972; Polivy et al., 1979) even after a 24-h food deprivation (Goldman et al., 1991). However, this hypothesis would predict that women should eat more in the presence of relatively high-intake males and men should eat less in the presence of relatively low-intake females. Using the data obtained with the diet diary technique this author demonstrated that females eat more with males than with females as predicted by the hypothesis. However, contrary to the predictions of the hypothesis, males exhibited social facilitation, increasing intake, regardless of whether they ate with males or females (de Castro, 1994). Hence, this hypothesis cannot explain social facilitation at least in males.
The presence of other people might induce an aroused state that leads to greater consumption (Zajonc, 1965, 1980). This explanation postulates that the presence of other people increases the individual's drive level, which facilitates the emission of dominant responses. Eating in this enhanced drive state would be expected to result in an increase in the rate at which the individual emits the dominant response of ingesting food. Indeed, a pecking companion increases the peck rate of a chick (Tolman, 1964). Analyses of the diet diary data fail to support this hypothesis. If the presence of other people increased arousal level, then it would be expected that the energization of eating behavior would result in a faster rate of ingestion. In fact, no differences have been found between the rate of intake for meals eaten alone and those eaten with other people present (de Castro, 1990). Results for the analyses of the duration and rate of intake with differing numbers of people present are presented in Figure 20-4. As shown, the rate of intake does not increase with
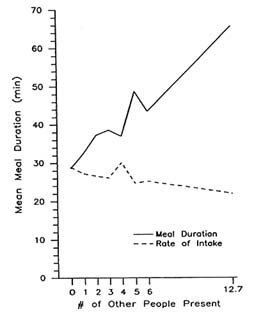
FIGURE 20-4 Mean meal duration (solid line) in minutes and the rate of intake (dashed line) in kcal/min, as a function of the number of people present at the meal.
social meals and may in fact decrease with large groups (de Castro and Brewer, 1992). Hence, an aroused state hypothesis cannot explain social facilitation of intake.
Social facilitation might operate by inducing an emotional response, such as increased anxiety or elation, which may produce an increase in intake (Harlow, 1932; Harlow and Yudin, 1933). This hypothesis would predict an increase in self-reported anxiety or elation with meals eaten socially. Indeed, Harlow (1932) observed greater emotionality in rats fed with others than when fed alone. However, this hypothesis, like the increased arousal hypothesis, predicts an increase in the rate of intake. Also, this hypothesis predicts that when eating with a companion, the more emotionally arousing the companion, the more will be eaten. Hence, eating with people well known to the subject and with whom the subject is comfortable, such as family and friends, should produce the least effect on intake. Analyses of the diet diary data again fail to support this hypothesis. Meals were separated according to the type of companion present with the subject: friend, family, spouse, work associate, or other. Average meal sizes for meals eaten with and without each of these companion types and meals eaten alone are presented in Figure 20-5. As shown, contrary to the hypothesis, family and friends had the greatest impact
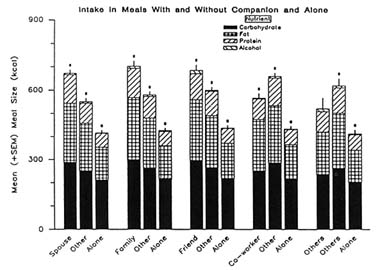
FIGURE 20-5 Mean (± SEM) amounts (kcal) ingested per meal of carbohydrate (solid portion of each bar), fat (cross-hatched portion), protein (hatched upward) and alcohol (hatched downward) for meals eaten with a particular companion type (first bar of each set of three), with others but not that companion type present (second bar of each set of three), and alone (third bar). The above the bar indicates a significant difference (P <0.05) between the total meal sizes as assessed with a t test. The above the first bar signifies the "with companion-with other" comparison, above the second bar signifies the "with other-alone" comparison, and above the third bar signifies the "with companion-alone" comparison. SOURCE: Reprinted from Physiol. Behav., 56(3), J.M. de Castro, Family and friends produce greater social facilitation of food intake than other comparisons, 445–455, 1994, with kind permission from Elsevier Science Ltd., The Boulevard, Langford Lane, Kidlington OX5 1GB, U.K.
on eating (de Castro, 1993a). In addition, the increased emotionality explanation predicts that when social facilitation occurs it should be accompanied by increased elation and/or anxiety, and when social facilitation does not occur, there should be no change in elation or anxiety. In fact, the self-ratings obtained for meals eaten with spouse, family, or friends, where social facilitation occurred, indicated less or equivalent anxiety. In addition, meals eaten with coworkers and others, where no social facilitation was apparent were eaten with higher levels of elation and anxiety (de Castro, 1993a). Hence, an increased emotionality hypothesis cannot explain social facilitation of intake.
Another potential explanation for social facilitation is that the presence of other people may increase perceived hunger (Tolman, 1968). A completely sated chick has been observed to begin eating again when placed in the presence of a hungry chick in the process of eating (Bayer, 1929). This hypothesis predicts that self-rated hunger would be higher at the beginning of socially facilitated meals but should be about the same at the end. In fact, in
all cases of social facilitation, people eat to a significantly higher level of satiety at the end of the meal than when alone (de Castro, 1993a). In addition, when meals are eaten with friends, where marked social facilitation occurred, the levels of premeal hunger are equivalent to the levels observed when the subject eats alone (de Castro, 1993a). Hence, an increased hunger hypothesis also cannot explain social facilitation of intake.
Social facilitation might operate by producing disinhibition; the presence of a companion relaxes the individual, thereby releasing behavior from inhibition (Rajecki et al., 1975). Observing someone else eating may remove constraints on eating that otherwise would limit the amount ingested. Social factors might also act by distracting the individual from the eating process and thereby release cognitive restraints. This notion would predict a greater level of calmness during social meals and a release of restraints on eating. The results of the analyses of the diet diary data tend to support this hypothesis, which predicts that the better known the companion, the greater the relaxation and thus the greater the facilitation of intake. Indeed, this author found that social facilitation was greatest when the subjects self reported greater calmness. This calmness occurred when subjects ate with friends, family, or a spouse. In contrast, when subjects self-reported greater anxiety and less calmness while eating with work associates, classmates, or other companions, social facilitation had the least impact on intake (de Castro, 1994; see Figure 20-5). Hence, the disinhibition hypothesis is a viable explanation of social facilitation of eating.
Finally, the presence of other people might quite simple extend the amount of time spent at a meal and thus increase the amount eaten (de Castro, 1990). Verbal interactions that occur during social meals may simply cause a person to linger over the meal and as a result eat more. This notion predicts that the rate of intake should be the same regardless of the social conditions, but the duration of the meal would be extended when other people are present. Indeed, the diet diary data indicate that the duration and not the rate of intake increases when meals are eaten socially (de Castro, 1990; de Castro and Brewer, 1992; see Figure 20-4). Furthermore, this hypothesis is also supported by diet diary findings in which social facilitation only occurs when eating with friends, family, or a spouse (de Castro, 1994). Hence, the time extension hypothesis is also a viable explanation of social facilitation of food intake.
Although the theories are highly speculative, it is possible that both disinhibition and time extension occur and account for social facilitation of intake. Time is extended, regardless of companion type, resulting in increased intake with all companions. Examination of the diet diary data with multiple regression suggests that even when the number of people present is considered, the presence of spouse, family, and friends continue to be associated with higher intake (de Castro, 1994). With spouse, family, and friends, the subject should feel the most comfortable. According to the disinhibition model, these people would tend to have a relaxing effect on the subject and thereby increase
intake by releasing inhibitions. The fact that where anxiety levels are the highest—during meals with coworkers and other companions—social facilitation is the smallest tends to support such an interpretation (de Castro, 1994). Hence, the best current available explanation for social facilitation involves a general time extension working in conjunction with a companion-specific disinhibition.
CONCLUSIONS AND RECOMMENDATIONS
Social facilitation of food intake has a number of practical consequences. It may be useful for decreasing intake when it is too high. Because eating alone reduces intake by over 200 kcal/d (Redd and de Castro, 1992), it may be useful as a dietary strategy in weight-loss programs. In theory, all other things being equal, this 10 percent reduction in intake should lead to the loss of 1 lb (0.5 kg) of fat every 18 days, and, if maintained, a 20-lb (9.1-kg) loss at the end of a year. However, either behavioral or physiological compensation for the lowered intake may occur over time, reducing the effectiveness of the eating alone manipulation. Longer-term studies than those present in the literature are needed to assess the usefulness of this manipulation as a dietary strategy.
Social facilitation of intake may also be useful for increasing intake when it is too low. Soldiers in the field under-ingest nutrients, and social facilitation may be useful in promoting greater food intake. Three different strategies are suggested by the research: eating in groups, modeling, and orders. Whether or not any of these strategies would work is speculative, and they have not, to the author's knowledge, been tried for prolonged periods of time or under anything like field conditions.
The research suggests that eating in groups has a major impact on intake, but its effectiveness for soldiers in the field depends on the situation and conditions. If soldiers are in a situation where they mainly eat alone, then social facilitation is likely to be of more benefit than if they already eat socially. This notion is suggested by the power function, which indicates that maximum increase in intake is obtained when going from eating alone to eating with one other person. Less and less benefit is obtained as more and more people are added to the eating context. Eating in groups may still be of benefit, but the magnitude of the effect would be expected to be greatly reduced.
The research also suggests that eating in groups would only be of benefit when the soldiers have time available to extend the duration of meals. If eating time must be short due to the field situation, then social facilitation would probably not have much effect. In addition, the research suggests that social interaction and conversation may be necessary, and conditions in the field may not be conducive to these kinds of interactions. The research also indicates that
eating with strangers would be ineffective. In sum, eating in groups may facilitate food intake if the meals are not already eaten socially, if time is available, if conversation is possible, and if friends are involved. However, once again, there are no studies of the long-term effectiveness of eating in groups. Behavioral or physiological compensation for the increased intake over time is a distinct possibility. Longer-term studies need to be carried out.
The research regarding modeling is not as well developed as that for eating in groups, and it has much larger gaps. Therefore, proposing field application is extremely speculative. In application, soldiers could be designated as high-intake models and instructed to eat large amounts in the presence of low-intake soldiers. Alternatively, soldiers who normally eat a lot could simply be paired with those identified as under-ingesters. Research to date has been confined to laboratory settings and has involved only single meals or snack intake. Whether similar methods would be effective in increasing overall intake for even brief periods in naturalistic settings is unknown. The only naturalistic data comes from diet diary studies which indicate that when females eat with a higher intake, male partner, an increment in food intake occurs. Again, the research is simply too sparse and lacks investigations of the long-term effectiveness of modeling as a facilitator of intake to reach any firm conclusions. Such studies need to be performed.
A final alternative is simply to order the soldiers to eat more. This method might be effective if a "facilitator" was designated whose job it was to order the soldiers to eat at mealtime and to prompt soldiers to eat more when they stop eating. The research on compliance suggests that this technique could be effective, however, such a scheme could have such negative consequences as producing anger and resentment, and the research on long-range effectiveness is absent.
In summary, the research on social facilitation and inhibition of intake clearly demonstrates that social factors can profoundly affect the amount of food ingested on a short-term basis. Eating in groups, modeling, and ordering all result in large increases in intake. There are no data to suggest that these strategies would not be effective in the field with appropriate conditions and implementation. However, there is also no direct evidence that such strategies would be effective, particularly over an extended period of time. Due to the magnitude of the effects, these strategies should be viewed as promising, and further research performed to ascertain whether this positive effects can be realized.
REFERENCES
Adleson, S.F. 1960. Some problems in collecting dietary data from individuals. J. Am. Diet. Assoc. 36:453–461.
Bandini, L.G., D.A. Schoeller, H.N. Cyr, and W.H. Dietz 1990. Validity of reported energy intake in obese and nonobese adolescents. Am. J. Clin. Nutr. 52:421–425.
Bayer, E. 1929. Beitriige zur Zweikomponententheorie des Hungers. Z. Tierpsychol. 118:283–349.
Bellisle, F. 1979. Human feeding behavior. Neurosci. Biobehav. Rev. 3:163–169.
Berry, S.L., W.W. Beatty, and R.C. Klesges 1985. Sensory and social influences on ice cream consumption by males and females in a laboratory setting. Appetite 6:41–45.
Block, G. 1982. A review of validations of dietary assessment methods. Am. J. Epidemiol. 115:492–505.
Brown, J.E., T.M. Tharp, E.M. Dahlberg-Luby, D.A. Snowdon, S.K. Ostwald, I.M. Buzzard, S.D.M. Rysavy, and S.M.A. Wieser. 1990. Videotape dietary assessment. Validity, reliability, and comparison of results with 24-hour dietary recalls from elderly women in a retirement home. J. Am. Diet. Assoc. 90:1675–1679.
Conger, J.C., A.J. Conger, P.R. Costanzo, K.L. Wright, and J.A. Matter 1980. The effect of social cues on the eating behavior of obese and normal subjects. J. Pers. 48:258–271.
Cottrell, N.B., D.L. Wack, G.J. Sekerak, and R.H. Rittle 1968. Social facilitation of dominant responses by the presence of an audience and the mere presence of others. J. Pers. Soc. Psychol. 9:245–250.
Crawford, M.P. 1939. The social psychology of vertebrates. Psychol. Bull. 36:407–446.
de Castro, J.M. 1987a. Macronutrient relationships with meal patterns and mood in the spontaneous feeding behavior of humans. Physiol. Behav. 39:561–569.
1987b. Circadian rhythms of the spontaneous meal patterns, macronutrient intake, and mood of humans. Physiol. Behav. 40:437–446.
1990. Social facilitation of duration and size but not rate of the spontaneous meal intake of humans. Physiol. Behav. 47(6):1129–1135.
1991a. Social facilitation of the spontaneous meal size of humans occurs on both weekdays and weekends. Physiol. Behav. 49(6):1289–1291.
1991b. Seasonal rhythms of human nutrient intake and meal patterns. Physiol. Behav. 50(1):243–248.
1991c. Weekly rhythms of spontaneous nutrient intake and meal pattern of humans. Physiol. Behav. 50(4):729–738.
1992. Age related changes in natural spontaneous fluid ingestion and thirst in human. J. Gerontol. 47(5):P321–P330.
1993a. The effects of the spontaneous ingestion of particular foods or beverages on the meal pattern and overall nutrient intake of humans. Physiol. Behav. 53(6):1133–1144.
1993b. Age related changes in natural spontaneous food intake and hunger in humans. Appetite 21:255–272.
1994. Family and friends produce greater social facilitation of food intake than other companions. Physiol. Behav. 56(3):445–455.
de Castro, J.M., and E.M. Brewer 1992. The amount eaten in meals by humans is a power function of the number of people present. Physiol. Behav. 51(1):121–125.
de Castro, J.M., and E.S. de Castro 1989. Spontaneous meal patterns in humans: Influence of the presence of other people. Am. J. Clin. Nutr. 50:237–247.
de Castro J.M., and D.K. Elmore 1988. Subjective hunger relationships with meal patterns in the spontaneous feeding behavior of humans: Evidence for a causal connection. Physiol. Behav. 43:159–165.
de Castro, J.M., and J.J. Goldstein In Press Eating attitudes and behaviors of pre- and postpubertal females: Clues to the etiology of eating disorders. Physiol. Behav.
de Castro J.M., and S.N. Kreitzman 1985. A microregulatory analysis of spontaneous human feeding patterns. Physiol. Behav. 35:329–335.
de Castro, J.M., J. McCormick, M. Pedersen, and S.N. Kreitzman 1986. Spontaneous human meal patterns are related to preprandial factors regardless of natural environmental constraints. Physiol. Behav. 38:25–29.
de Castro, J.M., E.M. Brewer, D.K. Elmore, and S. Orozco 1990. Social facilitation of the spontaneous meal size of humans is independent of time, place, alcohol, or snacks. Appetite 15:89–101.
De Luca, R.V., and M.N. Spigelman 1979. Effects of models on food intake of obese and nonobese female college students. Can. J. Behav. Sci. 11:124–129.
Dubois, S., and J.F. Boivin 1990. Accuracy of telephone dietary recalls in elderly subjects. J. Am. Diet. Assoc. 90:1680–1687.
Eagles, J.A., and D. Longman 1963. Reliability of alcoholics' reports of food intake. J. Am. Diet. Assoc. 42:136–139.
Edelman, B., D. Engell, P. Bronstein, and E. Hirsch 1986. Environmental effects on the intake of overweight and normal weight men. Appetite 7:71–83.
Forkman, B.A. 1991. Social facilitation is shown by gerbils when presented with novel but not with familiar food. Anim. Behav. 42:860–861.
Frank, J.D. 1944. Experimental studies of personal pressure and resistance: I. Experimental production of resistance. J. Gen. Psychol. 30:23–41.
Gersovitz, M., J.P. Madden, and H. Smicikalas-Wright 1978. Validity of the 24-hour dietary recall and seven-day record for group comparisons. J. Am. Diet. Assoc. 73:48–55.
Goldman, S.J., C.P. Herman, and J. Polivy 1991. Is the effect of a social model on eating attenuated by hunger? Appetite 17:129–140.
Goran, M.I., and E.T. Poehlman 1992. Total energy expenditure and energy requirements in healthy elderly persons. Metabolism 41(7):744–753.
Harlow, H.F. 1932. Social facilitation of feeding in the albino rat. J. Genet. Psychol. 43:211–221.
Harlow, H.F., and H.C. Yudin 1933. Social behavior of primates 1. Social facilitation of feeding in the monkey and its relation to attitudes of ascendance and submission. J. Comp. Psychol. 16:171–185.
Heady, J.A. 1961. Diets of bank clerks: Development of a method of classifying the diets of individuals for use in epidemiological studies. J. R. Stat. Soc. (Series A), pt. 3, 124:336–361.
Hoyenga, K.T., and S. Aeschleman 1969. Social facilitation of eating in the rat. Psychosom. Sci. 14:239–240.
Hsia, L.C., and D.G. Wood-Gush 1984. Social facilitation in the feeding behaviour of pigs and the effect of rank. Appl. Anim. Ethol. 11(3):265–270.
James, W.T. 1960. The development of social facilitation of eating in puppies. J. Gen. Psychol. 96:123–127.
Krantz, D.S. 1979. A naturalistic study of social influences on meal size among moderately obese and nonobese subjects. Psychosom. Med. 41:19–27.
Krantzler, N.J., B.J. Mullen, H.G. Schultz, L.E. Grivetti, C.A. Holden, and H.L. Meiselman 1982. The validity of telephoned diet recalls and records for assessment of individual food intake. Am. J. Clin. Nutr. 36:1234–1242.
Larkin, F.A., H.L. Metzner, and K.E. Guire 1991. Comparison of three consecutive-day and three random-day records of dietary intake. J. Am. Diet. Assoc. 91(12):1538–1542.
Latane, B. 1981. The psychology of social impact. Am. Psychol. 36:343–355.
Lissner, L., J-P. Habicht, B.J. Strupp, D.A. Levitsky, J.D. Haas, and D.A. Roe 1989. Body composition and energy intake: Do overweight women overeat and underreport? Am. J. Clin. Nutr. 49:320–325.
Livingstone, M.B., A.M. Prentice, J.J. Strain, W.A. Coward, A.E. Black, A.E. Barker, P.G. McKenna, and R.G. Whitehead 1990. Accuracy of weighed dietary records in studies of diet and health. Br. Med. J. 300:708–712.
Livingstone, M.B., A.M. Prentice, W.A. Coward, J.J. Strain, A.E. Black, P.S.W. Davies, C.M. Stewart, P.G. McKenna, and R.G. Whitehead 1992. Validation of estimates of energy intake by weighted-dietary record and diet history in children and adolescents. Am. J. Clin. Nutr. 56:29–35.
Mertz, W., J.C. Tsui, J.T. Judd, S. Reiser, J. Hallfrisch, E.R. Morris, P.D. Steele, and E. Lashley 1991. What are people really eating? The relation between energy intake derived from estimated diet records and intake determined to maintain body weight. Am. J. Clin. Nutr. 54:291–296.
Mori, D., S. Chalken, and P. Pliner 1987. "Eating lightly" and the self-presentation of femininity. J. Pers. Soc. Psychol. 53:693–702.
Mullenbach, V., L.H. Kushl, C. Jacobson, O. Gomez-Martin, R.J. Prineas, L. Roth-Yousey, and A.R. Sinaiko 1992. Comparison of 3-day food record and 24-hour recall by telephone for dietary evaluation in adolescents. J. Am. Diet. Assoc. 92(6):743–745.
Myers, R.J., R.C. Kresges, L.H. Eck, C.L. Hanson, and M.L. Klem 1988. Accuracy of self-reports of food intake in obese and normal-weight individuals: Effects of obesity on self-reports of dietary intake in adult females. Am. J. Clin. Nutr. 48:1248–1251.
Nisbett, R.E., and M.D. Storms 1972. Cognitive and social determinants of food intake. Pp. 190–208 in Thought and Feeling: Cognitive Alteration of Feeling States, H. London and R.E. Nisbett, eds. Chicago: Aldine Publishing Co.
Polivy, J., C.P. Herman, J.C. Younger, and J. Jaeger 1979. Effects of a model on eating behavior: The induction of a restrained eating style. J. Pers. 47(1):100–117.
Prentice, A.M., A.E. Black, W.A. Coward, H.L. Davies, R.R. Goldberg, P.R. Murgatroyd, J. Ashford, M. Sawyer, and R.G. Whitehead 1986. High levels of energy expenditure in obese women. Br. Med. J. 292:983–987.
Rajecki, D.W., R.F. Kidd, D.A. Wilder, and J. Jaeger 1975. Social factors in the facilitation of feeding in chickens: Effects of imitation, arousal, or disinhibition? J. Pers. Soc. Psychol. 52:510–518.
Redd, E.M., and J.M. de Castro 1992. Social facilitation of eating: Effects of instructions to eat alone or with others. Physiol. Behav. 52(4):749–754.
St. Jeor, S.T., H.A. Guthrie, and M.B. Jones 1983. Variability on nutrient intake in a 28 day period. J. Am. Diet. Assoc. 83:155–162.
Tolman, C.W. 1964. Social facilitation of feeding behavior in the domestic chick. Anim. Behav. 12:245–251.
1968. The role of tie companion in social facilitation of animal behaviour. Pp. 33–54 in Social Facilitation and Imitative Behaviour, E.C. Simmel, R.A. Hoppe, and G.A. Milton, eds. Boston: Allyn and Bacon.
Tolman, C.W., and G.F. Wilson 1965. Social feeding in domestic chicks. Anim. Behav. 13:134–142.
Welty, J.C. 1934. Experiments in group behavior of fishes. Physiol. Zool. 7:85–128.
Zajonc, R.B. 1965. Social facilitation. Science 149:269–274.
1980. Compresence. Pp. 35–60 in Psychology of Group Influence, P.B. Paulus, ed. Hillsdale, N.J.: Erblaum Press.
DISCUSSION
MARY MAYS: When we first heard about John de Castro's studies, we decided to see if we could get the effect in the lab Herb Meiselman and I agree that it is nice to get both field-based and lab-based studies agreeing, and this has never really been done in a systematic way.
So, we fed people—either alone or in groups of four—a spaghetti, salad, and dessert dinner. They were all the same sex. When we began, we put people in groups where they did not know one another because we did not want interference. We got no effects at all, no enhancement.
Then when we decided to test friends in both male and female groups, we got a 50 percent increase in both genders.
However—and this is something we did not make a lot of—it all practically increased dessert consumption. I know you did not find any differences in the nutrient composition, but practically all the social effects were increased consumption of dessert.
JOHN DE CASTRO: These are interesting findings, and they fit well with the comparison type finding I have reported. However, social facilitation may be affecting different components of the meal in different contexts.




















