
Status of Ecological Knowledge Related to Policy Decision-Making Needs in the Area of Biodiversity and Ecosystems in the United States
WALTER V. REID
World Resources Institute
CONTENTS
|
Biodiversity conservation, |
329 |
|||
|
Ecosystem services, |
330 |
|||
|
Resource management goals, |
331 |
|||
|
Where Can Improved Ecological Knowledge Best Aid Decision-Making? |
||||
Scientific understanding of the structure and function of ecological systems has advanced tremendously over the past three decades. During this time, ecology and evolutionary biology have changed from descriptive disciplines into experimental and theoretical sciences with an ever-growing capability for prediction. As a result, ecological knowledge has become more and more useful to policy-makers. The utility of ecological information has been further enhanced by advances in research tools and technologies—particularly the development of sophisticated mathematical models of ecological systems and the growing use of geographical information systems—that help to provide information to decision-makers in forms tailored to the decisions that they face.
Still, the limits of scientific understanding are obvious. Even where general trends, such as wildlife population declines or changing stream quality, are clear, scientists are often unable to determine the impact of a specific action on those trends with any precision (or even whether the trends are a consequence of previous human actions or are natural). Problems of cumulative effects, lack of site-specific ecological knowledge, and the natural variability of ecological systems conspire to add substantial uncertainty to almost all uses of scientific knowledge in environmental decision-making. As a consequence, we must place as much emphasis today on techniques and policies for coping with uncertainty as we do on efforts to reduce that uncertainty.
In this paper, I assess the influence of recent advances in ecological knowledge on environmental policy decision-making, the current status of policy-relevant ecological knowledge, and key opportunities where advances in knowledge (or technologies) would improve decision-making.
Ecological knowledge influences three general aspects of environmental policy. First, choices about basic societal goals are made based on knowledge—or assumptions—about how the world works. Consequently the overarching goals we set for environmental management are based, in part, on our knowledge of ecological systems. For example, it was long assumed that biological communities were highly co-evolved equilibrium systems of organisms. Based on that assumption, a common goal of resource management has been to maintain certain systems in their ''natural" state. But research that has found that biological communities are neither highly co-evolved nor regulated around equilibrium calls into question the very meaning of a natural state. We can't objectively define a natural state in the absence of equilibrium states because we have no way of knowing what the structure and function of a system would have been if humans had never intervened. While we might set a goal of minimizing human intervention in certain systems, our current knowledge of ecology suggests that we should not set a goal of maintaining a system in a "natural state."
Second, ecological knowledge undergirds our ability to predict the ecological—and economic and social—consequences of human impacts, such as filling a wetland, changing stream flows, or introducing chemicals into the environment. Until recently, virtually all environmental management has been reactive rather
than anticipatory, simply because we did not have sound information that would enable prediction of consequences of our actions. In effect, our management of resources has been a series of large scale—and often irreversible—experiments from which we often learned little because of the lack of information on baseline conditions or monitoring of trends. Now, with more sophisticated experimental research and modeling tools and information, we are better able to base decisions on anticipatory planning.
Third, ecological knowledge helps to define the types of policy tools and resource management practices that can be used to achieve various social and ecological objectives. For example, between the 1950s and 1970s, fisheries management relied on deterministic fisheries population models that were used to calculate harvest levels based on the maximum sustained yield of the population. As knowledge of fisheries population dynamics has grown, managers now realize that the non-linear nature of the population dynamics of many species, combined with the substantial uncertainty surrounding factors influencing recruitment and survival, requires different approaches to fisheries management—specifically, reliance on adaptive management techniques and highly conservative quotas.
ESTABLISHING GOALS FOR ENVIRONMENTAL MANAGEMENT
Do we know enough about ecological systems to be confident of the environmental management goals that we set? As our understanding of ecological systems has increased, we have refined environmental management goals and, in some cases, substantially changed them. For example, many of the regulations that were initially issued under the Clean Air Act and Clean Water Act in the early 1970s were based on limited knowledge of dose-response curves, environmental thresholds, and relative risks. With further research and experience with the consequences of various pollutants, those goals have been modified. More generally, though, how firm is the science on which we base our choice of environmental goals today?
Biodiversity conservation. Advances in scientific understanding of the importance of biodiversity and the threat that it faces is contributing to significant rethinking of biodiversity conservation objectives and goals. When the Endangered Species Act (ESA) was enacted in 1973, the goal of biodiversity conservation was straightforward—to save endangered species from extinction. The goal become substantially more complex, however, as scientists began to recognize the value of protecting all levels of biological diversity—genes, species, and biological communities. It became still more complex as the full magnitude of the species extinction problem in the U.S. was revealed. Where it may have seemed conceivable to protect all of the nation's species from extinction based on the knowledge available in the late 1960s, it is clear now that this would be extremely difficult and costly—not a surprising conclusion given the extensive habitat transformation that has taken place in the U.S.
Yet although current knowledge leads us to question aspects of the biodiversity policy goals implicit in legislation like the ESA, it is not yet clear what a new set of goals should be. For example, there is still no clarity on what the objective of conservation of biodiversity above the level of species should be. Is our goal simply to protect representative samples of biological communities in their own right, to protect samples of communities that also protect the widest diversity of species, or to protect communities and ecosystems that provide valuable ecosystem services? The problem is even more confusing when we consider that the composition and structure of communities will change through time as climate changes. Should we seek to protect the "arena" in which different distinctive communities exist, rather than attempting to protect "vignettes" of existing communities? If so, how would we choose those arenas? Similarly, although complete protection of all biodiversity is clearly an unrealistic goal, we are unable to determine what is realistic or desirable because we can't accurately estimate the social, economic, and ecological consequences of its loss.
Ecosystem services. We have long recognized the important role that ecosystems play in providing such "free" services as flood control (e.g., wetlands), water purification, maintenance of soil fertility, nutrient cycling, regulation of the micro-climate and regional climates, protection of coastal zones, dispersal and breakdown of wastes and cycling of nutrients, control of crop pests and disease vectors, and pollination of crops. And, to some extent, we have attempted to protect some of these services through legislative and regulatory actions. For example, Section 404 of the Clean Water Act, and the no-net-loss-of-wetlands policies of recent administrations, both seek to protect wetlands and the associated services that they provide. Similarly, requirements to maintain buffer strips along stream and river corridors help protect streams from sedimentation and changes in temperature and water flow.
More generally, however, we have established relatively few goals related to the protection of ecosystem services. This is due, in part, to the fact that with a few exceptions like wetlands and riparian habitat, our understanding of the relationship between various human pressures or changes in the structure or composition of ecological systems and the services they provide is very limited. Thus, even when the maintenance of the service is of obvious value, we are unable to define a management goal that will ensure the protection of the service. In some cases, where the maintenance of the service is particularly important we may set overly conservative goals. For example, protection of stream flows is often seen as essential for city or agricultural water supplies, and strict protection of the watershed is often the preferred means of ensuring those stream flows. In reality, changes could be made in an ecosystem (e.g., the addition or removal of species, harvesting of certain species, some transformation in the community structure) without any consequence for stream flows. But because in all but the most intensively studied systems, ecologists would have difficulty identifying what specific
changes could or could not be made without disrupting the hydrology, we tend to set conservative goals when protection of the stream flow is a priority.
Without better understanding of how various ecological changes affect ecosystem services, we will be hard pressed to set goals for the protection of those services. One attempt to circumvent this problem is to define goals based on the need to protect the "integrity" or "health" of ecosystems (Noss 1990, Karr 1991). Although intuitively appealing, these goals face even greater problems than defining goals based on the maintenance of specific ecosystem services. Although it is entirely appropriate for political or educational reasons to seek to maintain a "healthy ecosystem," just as we seek a "strong economy,'' the former provides no more guidance to the biological manager than the latter provides to the economist. Establishing when an ecosystem is "sick" or disintegrating is entirely subjective. Is the eastern deciduous forest more healthy today than it was before climate change led to the migration of oak and hemlock into the community 10,000 years ago? Did the loss of the chestnut decrease the biological integrity of the system? Like "ecosystem services," concepts like health or integrity are fundamentally anthropocentric—we define the health of an ecosystem based on the characteristics and services that we want the ecosystem to provide.
Resource management goals. The development of the field of population biology in the 1950s fostered the emergence of the fields of wildlife, forestry, and fisheries management as professional disciplines. Population models enabled the determination of quantitative goals for resource harvesting. Concepts like "maximum sustained yield" and "optimum sustained yield" became the guiding principles of resource management. Clearly, though, no single harvested species exists in isolation from other species in the community, and virtually all ecosystems provide far more products and services than just the few harvested species. As a result, harvesting at the optimum sustained yield for one species may prove suboptimal for the benefits we obtain from the ecosystem if the management practice depletes other resources. For example, there was considerable concern in the 1980s that the Southern Ocean krill fishery would have negative impacts on other populations dependent on that krill, such as whales, seals, and marine birds. Thus, today's management goals must often be multi-species or ecosystem-wide goals.
Yet how such an ecosystem-wide management goal should be defined is not clear. We know that we should consider values other than just those of the harvested species—such as the value of ecosystem services and various values associated with biodiversity (e.g., ethical, existence, option, and use values), but we typically don't have good estimates of those values for any given ecosystem, we don't know how changes in the population of a harvested species might affect those values, and we don't know what relative weight to give the various types of values. Indeed, ecosystem management goals must necessarily have political as well as scientific dimensions. For example, while scientists and economists could, in theory, determine the economic consequences of continued forest harvest in
the Pacific Northwest as a result of impacts on tourism, fisheries, and so forth, they are not in a position to weigh the ethical values of conserving a species like the Northern Spotted Owl against various economic values associated with continued loss of its habitat.
ASSESSING ENVIRONMENTAL IMPACTS
One of the few generalizations that can be derived from the past two decades of ecological research is that few generalizations can be made about the dynamics of ecological systems. This conclusion represents a significant change from the beliefs of researchers in the 1950s and 1960s, when studies suggested that ecological systems functioned in a state of dynamic equilibrium and that communities were tightly co-evolved assemblages of species. Considerable effort was devoted to the elaboration of underlying rules that would transform ecology into a predictive science. Today, there are relatively few general theories that researchers would be willing to apply to communities or to ecological interactions in a predictive manner. This change has come about as scientists came to realize that most communities appeared to be non-equilibrium systems with non-linear dynamics (Botkin 1990, Buzas and Culver 1994).
With few predictive theories at hand, considerable emphasis must be placed on empirical studies of specific systems to develop the ability to assess the consequences of change in those systems. Our ability to evaluate the impacts of any given perturbation depends strongly on the availability of long-term data on the particular system and in particular on the availability of experimental information bearing on the change being assessed. Even where scientists can predict the direct consequences of a specific perturbation on certain species (for example, the impact of a pesticide on a particular species), we are unable to predict the secondary effects of those changes on the structure and function of the ecosystem without much more detailed information.
The status of our knowledge about the impacts of human-caused changes differs depending on the type of perturbation involved:
Pollution and toxic chemicals
Our ability to predict the direct effect of a number of different classes of toxic chemicals on various wildlife populations is quite good. For many families of pesticides, for example, we have a good understanding of the mechanism by which the chemical works, and can predict which species might be affected by related chemicals and how they might be affected. Moreover, laboratory studies can often be used to identify particularly threatening chemicals relatively quickly and cheaply.
More worrisome, however, are chemicals that do not exhibit direct toxic effects on plants and animals but may have chronic effects. For example, a growing body of evidence suggests that a broad array of "estrogenic" chemicals may be influencing the reproductive physiology of many wildlife species (Colborn
and Clement 1992, Kelce et al. 1995). Even where evidence of effects exists, there are substantial difficulties in distinguishing these effects from natural variation due to the lack of baseline information. Given the rapid growth in synthesized chemicals being released into the environment, and the potentially profound impacts on ecological and human social systems if some of these materials were responsible for chronic effects that changed basic life history characteristics of the species involved, there is a substantial need for further research on these potential risks.
The effects of many other widespread air and water pollutants—such as nitrogen in water or sulfur dioxide in air—on ecological systems has received considerable study over the past two decades. Although site-specific predictions of the impacts of such pollutants are often uncertain, the general effects of those pollutants in different types of ecosystems can often be predicted with reasonable accuracy. As the experience with the Exxon Valdez oil spill in Prince William Sound in Alaska demonstrates, however, considerable uncertainty and inaccuracy can surround the evaluation of impacts of even widespread pollutants like oil. In this particular case, the widespread initial predictions of an ecological disaster proved far too pessimistic. While the spill may still have substantial long-term impacts on the ecological systems, it is unlikely that it will be possible to distinguish those impacts from natural fluctuations since the baseline of data in the region is not extensive.
Habitat loss and fragmentation
One ecological generalization that has withstood the test of time is that the number of species found in a given habitat varies with the size of the habitat (see Figure 1). Consequently, when habitat is lost the number of species present will be reduced. While this species-area effect does allow a general assessment of the likely consequences of habitat change, it cannot provide detailed assessments of impacts on biodiversity. The actual impact of habitat conversion depends strongly on which areas are lost. Because species with restricted ranges are sometimes clustered in "hot spots" of diversity, protection of those hot spots could greatly reduce the effect of habitat loss on species diversity.
For a given species for which detailed demographic and life history information is available, we are able to make projections of the probability of survival of the species when its habitat is lost by use of Minimum Viable Population methodologies (Shaffer 1981). Insufficient demographic information is available for the vast majority of species, however, to allow such calculations.
The study of the impacts of fragmentation on ecological systems is still a relatively new field. Fragmentation can have significant effects both due to the increased ratio of edge habitat to core habitat (edge habitats have a different micro-climate and different species composition), and due to the access edge habitat may provide for certain species into core regions that would be otherwise inaccessible, as well as the barriers it may create for the dispersal and migration of
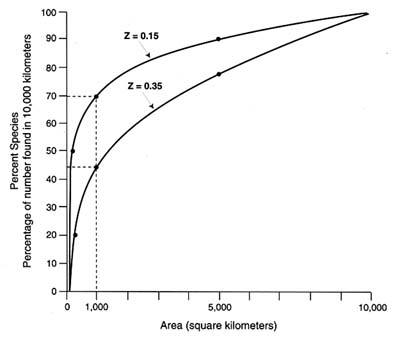
FIGURE 1 Species-area curve. SOURCE: M.L. Shaffer. 1981. "Minimum Population Sizes for Species Conservation." BioScience 31.
other species. Fragmented habitats are clearly far less desirable for conservation purposes than continuous habitats, but the specific impacts of fragmentation on a given species can only be determined through careful study of that species.
Species addition or removal
The loss of keystone species from ecological communities can have profound effects on the system because these species play important functional or ecological roles. Keystone species can include (i) predators, herbivores, parasites, and pathogens that maintain diversity among competing organisms by reducing the abundance of dominant competitors and preventing competitive exclusion; (ii) mutualists that link the fate of many species, such as specialized pollinators; and (iii) species that provide resources critical to the survival of dependent populations during periods of low resource availability. It is not possible, however, to determine which species serve "keystone" roles in a biological community without detailed experiments. In particular, neither trophic level, abundance, nor body size of a species is a good predictor of keystone status.
Introductions of certain species can also profoundly change ecosystem services. For example, the introduction in southwestern U.S. of the deep-rooted Tamarix tree, which has the ability to draw water from unsaturated alluvial soil—an ability missing in native trees in the region—has altered ecosystem services
related to groundwater levels and stream flows. While it is extremely difficult to predict the consequences of an introduction, certain types of species are clearly higher risk than others. Weedy ("r"-selected) species that have large numbers of offspring, rapid growth, and high dispersal abilities, are more likely to become established in new habitats than "K"-selected species with lower population growth rates.
While the removal of a keystone species will, by definition, have profound impacts on a community and the services it provides, the removal or addition of some species in communities appears to have little demonstrable effect either on other species or on an ecosystem process. In these situations, other species compensate for the absence of the target species, at least over the short term. (It is not known for sure if all functions of the species in question are compensated for; in fact, it is rarely understood what the full range of functions is for each species.) As is the case with keystone species, it is not possible to predict based on general information about the biology of a species whether its loss would have little effect on the ecosystem. However, the one group of species whose loss would tend to have relatively little impact contains species that are currently in danger of extinction, simply because the primary impact of their loss would already have been felt when the species was originally reduced in population size to the point of endangerment.
Thresholds may sometimes exist in the response of ecosystem services to changes in abundance or distribution of species. In many of the world's biomes, ecosystems can exist in two or more alternative states that differ widely in rates of productivity or some other ecosystem services. Semiarid grassland can remain productive as long as grazing density is modest, but when density rises above a threshold the ecosystem shifts rapidly to a state of low productivity. Often, large and sustained reductions in exploitation are necessary to return ecosystems to high-productivity states, and in some cases, recovery from the undesirable state is not possible. At present, scientific knowledge is insufficient to be able to predict either the presence or the level of thresholds in ecosystem behavior.
Climate change
We currently have insufficient knowledge to make accurate assessments of the potential consequences of human-induced climate change on ecological systems. A few general predictions can be made: species found only high on mountains will be threatened with loss; protected areas with significant elevational or latitudinal gradients will be more secure than those without; species restricted to coastal regions will face significant threats from sea level rise, etc. By virtue of more complete knowledge of the requirements of crop plants, our ability to make predictions of impacts is somewhat better for agro-ecosystems than for natural systems. But our predictive ability is seriously constrained due to the high regional uncertainty of predictions from climate models and the lack of detailed autecological information (information on the interaction of a species and its physical environment) on many important species.
The predictive accuracy of climate models is quite low at a regional scale—the very scale where climatic factors become ecologically relevant. Until more precise regional estimates of changes in rainfall, temperature, seasonality, and weather disturbance frequencies are available, only general impact assessments are possible. But even when more detailed regional information becomes available, assessments will be hindered by the lack of information on the response of individual species to changes in climate. Because communities are not deterministic equilibrium assemblages of species, any climatic changes will have different consequences for different species. For tree species, which are some of the most structurally significant species in many North American communities, we do have relatively good paleoecological information on their rates of dispersal in response to past climate changes. Dynamic forest models also allow some prediction of how changes in climatic factors will alter forest community composition (Botkin 1990). For most other species, however, changes in response to climate shifts cannot be accurately predicted.
Moreover, the effect of changing distributions of species on various ecosystem services is even less certain. For example, changes in species distribution as a result of climatic changes will alter patterns of disease incidence. Such an effect can be seen in evidence that links the outbreak of cholera in South America in 1991 to El Ni ño (Epstein et al. 1993, Stone 1995). The warming of the waters off the coast of South America may have stimulated growth of a plankton harboring the cholera bacterium. While the increased frequency of El Ni ño in recent years cannot be conclusively tied to human-caused changes in climate, the example demonstrates how ocean current changes that are likely to occur in the event of global warming could have substantial effects on human health. Current models of changes in the distribution of disease vectors under likely future climates suggest that developing countries will see an increase in malaria, schistosomiasis, sleeping sickness, dengue, and yellow fever. The outbreak of the hantavirus in the southwestern U.S. has also been linked to weather conditions, possibly also associated with El Ni ño.
Introduction of genetically engineered organisms
Introductions of new cultivated varieties of crops (developed either through traditional breeding methods or through genetic engineering) into natural or agroecosystems can pose significant risks, such as the introgression of genes into wild populations, weediness, and pathogenicity. Genetic material from introduced plants, animals, and micro-organism can be transferred into wild populations of related species through the formation of fertile hybrids. The new genetic material can then potentially alter the ecological interactions of that wild relative. For example, a disease or frost resistance gene transferred into a wild weedy relative of the crop could extend the range of that wild relative. To date, most gene flow in agricultural systems has taken place between crops and their weedy relatives, but in aquatic systems, substantial gene flow occurs between hatchery-reared fish and wild populations.
Current ecological knowledge can help minimize risks from the introduction of transgenic species. For example, risks are clearly higher in regions where wild relatives of the species are present.
These five examples demonstrate that substantial knowledge gaps exist that prevent a clear assessment of the ecological effects of any particular pressure or perturbation on a system. For certain types of well-studied perturbations, such as the application of the pesticide DDT, scientists can make detailed predictions of consequences. For certain well-studied species, such as deer, we can also make precise assessments of the impact of any change in the population of that species. But, by and large, unless a particular system has been the subject of intense research and monitoring, the current status of ecological knowledge only enables us to identify a list of potential consequences of a given action and to identify the set of data or experiments that would enable that uncertainty to be reduced.
MANAGING RESOURCES
Ecological knowledge helps to define the types of management practices and policy tools available for resource management. Current management practices have been influenced by advances in ecological knowledge, particularly increased understanding of (i) the natural variability of ecological systems; (ii) the non-linear nature of many ecological interactions; and (iii) the site-specific nature of many ecological interactions and processes (Ludwig et al. 1993).
For decades, wildlife and fisheries managers relied on demographic models that assumed far less natural variability in populations and more linear population responses than is now recognized to generally be the case. The knowledge available at the time led to the design of management techniques like calculating the maximum sustainable yield for a population and then setting harvest levels based on those estimates. Now, we recognize that the non-linear aspects of the population dynamics of most species place insurmountable limits to the precision of population projections. As a consequence, managers increasingly set goals for population management based on a safe minimum standard rather than a prediction of the optimum sustained yield.
Natural variability of ecological systems also makes it exceedingly difficult to evaluate the consequences of any particular management intervention. Adaptive management practices have been developed to reduce uncertainty and improve resource management through time. Adaptive management is a technique for managing biological systems so as to simultaneously reduce uncertainty about the functioning of the systems and respond to the changing social, biological, and physical environment. The principal elements of adaptive management are (i) management interventions are made in an experimental manner so that the outcome of the intervention can be used to reduce uncertainty about the system; (ii) sufficient monitoring prior to and during the intervention enables detection of the results of the management intervention and thereby allows managers to learn
from past experience; and (iii) based on the feedback to managers, communities, and other constituencies, management interventions are then refined.
Recognition of the inherent variability in natural systems has also led to greater emphasis being placed on the need for long-term monitoring. The recent detection of a dramatic drop in phytoplankton abundance off the coast of Southern California was made possible only due to a monitoring program initiated in 1951 by the California Cooperative Fisheries Investigations (CalCOFI) (Roemmich and McGowan 1995). In contrast, the causes of coral bleaching episodes in the Caribbean remain controversial, in part because of the lack of long-term monitoring of water temperatures in the region. Without long-term records, it is difficult to distinguish human-caused changes from natural changes in most systems. Even with long-term records, however, conclusive evidence of human-caused influences typically also requires some form of experimental verification.
Finally, because of the predictive limits implicit in the fact that ecological communities are not tightly co-evolved assemblages of species existing in an equilibrium state, a premium is placed on site-specific information about the identity and biology of the species that occur in a given area. Resource managers are typically constrained by the absence of inventories of species in most regions of the United States.
To some extent, the need for inventories could be diminished if we had better knowledge of the complementarity of patterns of distribution of various groups of species. It has long been assumed, for example, that the pattern of diversity of plants would be a good predictor of the pattern of diversity in other groups of terrestrial organisms, because of the importance of the physical structure provided by plants for the diversity of other taxa in the community. We know that the distribution of plant diversity does not completely mirror other groups (Prendergast et al. 1993), but there are relatively few studies that seek to determine just how coincident patterns actually are.
WHERE CAN IMPROVED ECOLOGICAL KNOWLEDGE BEST AID DECISION-MAKING?
Setting Goals
The greatest need for improved ecological knowledge currently relates to its role in setting environmental goals. We are in the midst of a fundamental rethinking of environmental goals, but while our science is good enough to call into question some of our past goals, we cannot yet be certain of their replacements. In the past we set narrowly defined goals for species protection, wetlands conservation, or sustainable harvests. Yet progress toward these narrow goals did not always achieve appropriate balance among the many benefits we received from living systems and were often in conflict. Worse still, many of our environmental management goals were designed to be reactive to threats to the environment and,
as a consequence, did not serve well as means to anticipate and prevent environmental threats. For example, we sought to prevent endangered species from going extinct, but paid less attention to the need to prevent species from becoming endangered.
It is fashionable now to speak of ecosystem management as the new environmental management paradigm. This may be true, but it is not a clear environmental management goal. Ecosystem management is a natural outgrowth of the recognition that the products and services we obtain from living systems are interdependent. It also is in keeping with the need to develop anticipatory policies to replace our current reactive regulations. But until we develop specific goals for "ecosystem management," it will be all things to all people. Advances in knowledge bearing on the following questions would significantly aid in the development of sound environmental management goals:
1. What is the distribution of biodiversity in the U.S.? How much complementarity exists between regions of high conservation value for genetic, species, and community diversity? How sensitive is the protection of biological diversity to the area protected? How much opportunity is there to protect biodiversity in disturbed landscapes?
One of the most difficult challenges for sustainable resource management in the U.S. is the protection of biological diversity. Our current focus on protecting endangered species from extinction needs to be complemented by efforts to prevent species from becoming endangered. This can be done most efficiently by protecting critical ecosystems from transformation. We still do not know the extent to which the protection of certain hot spots of biological diversity would protect the nation's biodiversity or the extent to which biodiversity can be protected in disturbed landscapes. We also do not know how effectively the protection of hot spots of species diversity also serves to protect community and genetic diversity. Finally, for many lesser-known taxa such as many invertebrates and fungi, we do not know enough about their status or distribution to know whether they are being adequately protected under current conservation policies. The answers to these questions bear on both issues related to management strategies (how can we most efficiently protect biodiversity) and to the goals themselves (what and how much are we trying to protect).
2. What is the economic value of various services we obtain from ecosystems and what is the economic value of biological diversity? How should we weigh instrumental with intrinsic values of diversity?
Decisions about the protection of many ecosystem services are fundamentally economic. The loss of the flood protection service of wetlands, for example, can be partially offset by the construction of dams and levees. However, because we do not have good estimates of the costs incurred by many environmental changes, they are not factored into the cost-benefit calculus of decision-making.
Particularly given the political climate that stresses greater and greater attention to such cost-benefit calculations, it is imperative that the ecological values be well represented. Consider three examples. The commitment to the goal of no-net-loss of wetlands is admirable. However, given the substantial benefits that wetlands provide in the form of flood control it may well be the case that an economically justified goal should be net gain in wetlands. Similarly, in the Pacific Northwest, the amenity value of relatively undisturbed ecosystems has a high economic value, demonstrated by the rapid growth in "footloose" industries in the Northwest. A more accurate calculus of the economic costs and benefits of resource conservation in the Northwest would probably give greater weight to the goal of biodiversity conservation. Finally, although the protection of habitat in rapidly growing urban areas is often criticized as being economically unsound, the maintenance of greenbelts and open space in urban areas is often found to have significant economic benefits. Again, better calculus of the economic values of ecosystem services and natural systems could often lead to the incorporation of those values as one goal of management. This would aid in defining goals not only for the protection of ecosystem services but also for multi-species resource management.
3. How can planning tools be made to be more interactive and accessible to a broader cross-section of the public?
Clearly, the goals of resource management cannot and should not be based only on economic considerations. Many values of biodiversity, for example, involve ethical or moral considerations and thus cannot be incorporated into cost-benefit calculus. Moreover, the protection—or use—of particular resources may have profound cultural significance that would be difficult to appreciate in studies of valuation. (How else to explain the surprisingly strong support for grazing-land management policies that benefit a tiny fraction of people at great cost to taxpayers and to the environment?)
Geographic information systems provide a tool that with sufficient development could be used by a broader array of civil society to learn about both the values of the resources in their region and the consequences of various management options. These tools would enable people to better understand the consequences of various options for both the short-term and the long-term economic and ecological outlook in a region. Without such tools, neither the civil society nor its representatives in government are able to adequately weigh the pros and cons of different resource management options.
Assessing Impacts and Managing Resources
The obstacles to enhanced ability to predict the consequences of human impacts on specific systems and the ability to successfully manage resources are similar. Most significantly, they relate to need for significantly improved site-specific
inventory and research and to the need for long-term monitoring (ESA 1991, NRC 1993).
4. What species and communities occur in specific ecosystems and regions? What are their ecological requirements? How does the diversity of species influence various ecosystem services? What species play particularly important—keystone—roles in the system? How does fragmentation influence the key species? What methods can be used to successfully restore degraded habitats or ecosystem services?
Site-specific information is of fundamental importance for successfully managing biological systems and for predicting the consequences of changes in those systems. This does not mean that we need to know everything about all systems. But it does mean that unless we have certain basic knowledge we will have limited ability to manage systems or to prevent unwanted changes. Perhaps the most fundamental need is to improve understanding of what species and resources are present in the system (NRC 1993).
5. What are the long-term trends in the structure and function of given biological communities and what is the variability in various measures?
There are only two ways of obtaining information about human impacts on systems. First, the perturbation can be made in a controlled experiment. Second, and much less conclusive, the effect of a perturbation can be compared to the long-term behavior of the system. While experimental ecology will continue to provide the primary means of teasing apart the workings of ecological systems, we must turn to long-term monitoring as the only practical means of identifying unexpected impacts and enabling adaptive management in situations where the replication of an experiment is impossible. However, the history of ecological research and funding, driven as it is by proposals to test specific hypotheses, is fundamentally in opposition to the need for monitoring and long-term ecological research.
6. What are the chronic effects of chemicals being released into the environment on plant and animal populations?
Sufficient evidence of endocrine disruption of wildlife populations caused by chemicals released into the environment now exists to warrant a substantial research effort to pin down the causality and to determine what classes of chemicals represent the greatest threats. (The potential impacts on human reproduction also warrant study.) The combination of the potentially dramatic effect of these chemicals on ecological systems and the rapid growth in the release of these chemicals makes this a high priority concern.
7. How will species in various ecosystems respond to changes in temperature, precipitation, disturbance, and CO2levels as predicted under climate models?
The ability of human societies to adapt to global changes will be linked in no small part to our success in ensuring that ecological systems adapt to change (NAS 1991). As predictions of changes in weather patterns become more refined, ecologists need to be in a position to model the responses of ecological systems. This will enable us to identify ecosystem services that might be threatened or species whose conservation will require which management interventions, and to evaluate potential economic costs or public health risks from ecological changes.
8. What environmental indicators can be developed that bear on the achievement of environmental management goals?
Indicators of the status of ecological systems in relation to some agreed goal or target, or of the value of various ecosystems or services, both provide an important means of communicating the status of ecological systems to decision-makers and the public, and present the opportunity for designing new conservation policies based on market incentives rather than regulatory approaches. With better indicators of the value of different types of resource management practices for the conservation of biodiversity, for example, it would be possible to create incentive (e.g., tradable permits, impact fees, tax breaks) that would encourage landowners to manage resources in a fashion that protects biodiversity.
CONCLUSIONS
Consider a classic case where ecological knowledge is called on to aid environmental decision-making: a proposal to build a housing development on natural habitat near an urban area. Should the development be allowed? What will its impact be on the environment? Are there alternative sites where the development could be placed with less impact? What information is needed to aid in the decision? What changes can be made to minimize the adverse impact?
Based on the analysis of this paper, the following insights on the role of scientific and technical information bear on the case. First, prior to assessing the impacts we need to clarify what the goals of environmental protection are for the region. The science is fairly clear on some aspects of those goals: the need to protect important services such as stream flow and flood control, the need to protect the diversity of species in the region. Some other goals, such as the need to protect representative samples of communities, may be less clear. Is the intent to actually protect the natural community that exists or simply to ensure that a representative sample of the bioregion is protected, recognizing that the species composition may change through time? More work is needed to clarify environmental goals based on recent scientific findings.
At the level of actual impact assessment, a qualitative assessment and evaluation of alternatives can be made with relatively little site-specific knowledge. Certain critical habitats, such as riparian communities, wetlands, steep slopes, and so forth, need to be protected to maintain ecological services in any region.
Relatively little site-specific information is needed to also allow a proposal to be modified to minimize its impact—minimizing fragmentation of habitats, avoiding migratory corridors, using native species in landscaping, avoiding riparian or wetland habitats, etc. But beyond these general statements, as assessment requires site-specific information on at least the identity, distribution, and range of the various species and community types. Ideally, sound planning also requires time-series information and knowledge of the ecological interactions in the ecosystem. Obviously, that ideal situation is rarely found. At best, impact assessments can only (i) identify critical resources (e.g., habitats of rare species) and (ii) weigh the relative risk of various alternatives. It is rare that any precise information can be given on the actual amount of impact on an ecosystem that would take place, much less the economic consequences of those impacts.
We do know enough, however, to design human interventions in ecosystems in a manner that will help us learn more about those ecosystems, through adaptive management. But for this approach to succeed, the financial resources need to be available for long-term studies and monitoring. Such long-term research, with its—by definition—delayed payoff, tends to be highly vulnerable to budget cuts.
The nation's biodiversity and ecological systems are resources that are growing in importance as they become increasingly imperiled and as society places ever greater values on the diversity of biological assets for tourism, recreation, ecological services, and quality of life, not just on their value as extractive resources. But our policies and actions still do not reflect this growing value—land is still ''developed" only when we've altered the natural communities, even though the wisest economic development of much of our land now is to keep it in a natural or semi-natural state. Ecological knowledge can help improve our management decisions. But because we still undervalue ecological systems as resources, we also undervalue the need for the inventory, monitoring, and research that will enable us to anticipate and prevent their loss and degradation.
BIBLIOGRAPHY
Botkin, D.B. 1990. Discordant Harmontes. Oxford University Press, New York, N.Y.
Buzas, M.A., and S.J. Culver. 1994. Species pool and dynamics of marine paleo communities. Science 264: 1439–1441.
Colborn, T., and C. Clement, eds. 1992. Chemically Induced Alterations in Sexual and Functional Development: The Wildlife/Human Connection . Princeton Scientific Publishing, Princeton, N.J.
Cullota, E. 1995. Will plants profit from high CO2 ? Science 268:654–656.
Epstein, P.R., T.E. Ford, and R.R. Colwell. 1993. Marine ecosystems. The Lancet 342:1216–1219.
ESA [Ecological Society of America]. 1991. The sustainable biosphere initiative: an ecological research agenda. Ecology 72(2):371–412.
Karr, J.R. 1991. Biological integrity: a long-neglected aspect of water resource management. Ecological Applications 1:66–84.
Kelce, W.R., C.R. Stone, S.C. Laws, L.E. Gray, J.A. Kemppainen, and E.M. Wilson. 1995. Persistent DDT metabolite p, p'-DDE is a potent androgen receptor antagonist. Nature 375:581–585.
Ludwig, D., R. Hilborn, and C. Walters. 1993. Uncertainty, resource exploitation, and conservation: lessons from history. Science 260:17, 36.
NAS [National Academy of Sciences]. 1991. Policy Implications of Global Warming. National Academy Press, Washington, D.C.
Noss, R.F. 1990. Can we maintain biological and ecological integrity? Conservation Biology 4:241–243.
NRC [National Research Council]. 1993. A Biological Survey for the Nation. National Academy Press, Washington, D.C.
Prendergast, J.R., R.M. Quinn, J.H. Lawton, B.C. Eversham, and D.W. Gibbons. 1993. Rare species, the coincidence of diversity hotspots and conservation strategies. Nature 365:410–432.
Roemmich, D., and J. McGowan. 1995. Climatic warnings and the decline of zooplankton in the California Current. Science 267:1324–1326.
Shafter, M.L. 1981. Minimum population sizes for species conservation. BioScience 31:131–134.
Stone, R. 1995. If the mercury soars, so may health hazards. Science 267:957–958.


















