
8 Population Biology of the Elderly
James R. Carey and Catherine Gruenfelder
Introduction
An important, yet widely overlooked incongruity in ecological and evolutionary literature concerns the gap between the emphasis on understanding age-structured populations (Caswell, 1989; Charlesworth, 1994) and the elderly as a subgroup of the population. This disparity implies that age structure is important, but only within a restricted subset of younger age classes. Indeed, the elderly are often viewed as expendable and thus as possessing little real or potential ability to contribute to the fitness of the population (Williams, 1957).
There are several likely reasons for historical neglect of the elderly in behavioral ecology literature. First, it is widely known from field life tables that most individuals in the wild die young and even fewer survive to older ages (Deevey, 1947). Thus, it is argued that their role is negligible, since the elderly subgroup of animals in the wild is not only frail but also is only a small minority of the total population.
Second, the evolutionary theory of aging (Williams, 1957) suggests that older individuals contribute little to fitness of populations because, according to the Lotka model (Lotka, 1907), the force of selection decreases with age (Rose, 1991; Kirkwood, 1985). One of the problems with the use of the Lotka model to define fitness is that it does not consider some situations where the elderly may contribute substantially to the fitness of the population. For example, it does not consider circumstances in which only older adults, particularly females, overwinter and regenerate the population in the spring (Izquierdo, 1991), in which the elderly provide infant care, or in which the older matriarchs or patriarchs provide critical leadership and cohesion.
Third, senility is often used as the physical and behavioral criterion for defining and identifying the elderly in the wild—i.e., those individuals that are decrepit, moribund, and near death. Unfortunately, this anecdotal "aging" of wild individu-
als is subject to substantial error. Only in the last several decades have detailed longitudinal data been systematically gathered on individuals of known ages and greater emphasis been placed on understanding age structure of populations (Finch and Ricklefs, 1991). Researchers have found that the variability in the rate of aging in wild animals is as great as that in humans, and in most species, it is virtually impossible to determine the exact age of an individual by its appearance or a biomarker. This is particularly true at older ages. Indeed, "physiologic heterogeneity" is a consistent characteristic of the elderly population (Timiras, 1994).
The broad objective of this paper is to extract important facts about aging and social and community structure in nonhumans and to reassemble them on a foundation of ecology and population biology. Just as certain morphological traits in animals, such as antler type, leg length, wing shape, visual acuity, gut length, and ear design, have been successfully related to the ecology and life histories of different animal species, we expect longevity and mortality patterns to be interpretable in the same context. We believe that this approach will provide insight into questions such as: What aspects of natural history favor the evolution of greater longevity? To what extent is life span a function of ecology and social structure? Is slowing of mortality at older ages a consequence of selection for certain adaptive traits at younger ages?
We organized this paper into three broad sections and a discussion. In the first section we introduce foundational principles we believe are important for understanding the relationship between longevity and life history of animals. These include biodemographic concepts and behavioral characteristics, such as altruism, dominance, territoriality, learning, and culture. In the second section we present a synopsis of roles and life histories of the elderly in three selected animal groups, including elephants, cetaceans, and primates, to provide specific biological context. In the third section we explore the concept of extended longevity as a preadaptation for the evolution of eusociality in wasps. This concept is important because it broadens the evolutionary scope from microevolution, as is discussed by Rose (1991) and Kirkwood (1985), to macroevolution concerned with speciation and the evolution of broad taxonomic groups (Strickberger, 1996). In the discussion we attempt to synthesize the general findings of the natural history and roles of the elderly, discuss some common life-history patterns associated with extended life span, and suggest future directions.
Foundational Concepts
The Elderly: An Integration of Biodemographic Concepts
Defining the Elderly
Life span of animals is not an orderly unfolding of precisely timed events from fertilization to death, with the elderly as a distinct, definitive stage. There exists no objective physiological landmark for aging, such as occurs with sexual
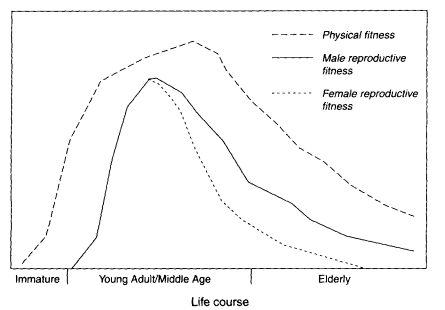
Figure 8-1
Physical and reproductive fitness generally peak during middle age and tend to steadily decline throughout the elderly stage (shown here to be the last third of the life course).
maturation in most animals. Its onset occurs at some indeterminate point following maturity, and its progression follows timetables that differ with each individual (Timiras, 1994). Consequently, a consistent characteristic of the elderly in nature is the wide range of frailty and robustness; therefore, it is impossible to specify for any species the age at which the elderly stage begins. However, as an operational definition, we consider the elderly in nature as those individuals who survive to the later half to third of the life span (Figure 8-1). Perhaps a more realistic framework, but one that would not serve the current purposes, would be to consider functional, rather than chronological, markers to define old age (Silverstone, 1996).
Life-Course Premises
Many of the premises outlined by Riley (1979) for the life course and demography of older humans are general and thus relevant for understanding the life course, population dynamics, and aging of populations for any species. These premises are as follows: (1) Aging for all animals is a continual process of development, maturation, and growing old. Thus, no single stage of an individual's life can be understood apart from its antecedents and consequences. (2) Aging consists of several sets of processes, including biological, social, and
experiential processes, all of which are systematically interactive over the life course. (3) The life-course pattern of any particular individual is affected by changes in the group and in the environment. (4) Not all individuals survive to old age. However, the importance of the elderly to the overall dynamics and welfare of the population may be disproportionately greater than their numbers.
Consequences of Long Life
Long life has several consequences for individuals in animal populations (Riley and Riley, 1986). First, long life prolongs opportunity to accumulate social and biological experiences. For example, long-lived individuals in social species may possess unique knowledge, such as the location of water during a drought or how to manufacture and use tools. Second, long life maximizes an individual's opportunities to complete or to change role "assignments." For example, a long-lived animal can ascend, lose, and re-attain alpha status within a troop, serve as a grandparent "caregiver"; become a forager after having been a colony "nurse"; become a breeder, by inheriting breeding territory, rather than remain a nest helper. Third, greater longevity prolongs an individual's relationships to others, including mates, parents, offspring, and associates. This feature increases the structural complexity of an individual's social networks, such as kinships, alliances, and communities. For example, when a monogamous pair of birds survives over multiple years, the relationship can become highly complex, leading to large clutch sizes and increased breeding success not seen in short-lived species (Coulson, 1966). In general, these consequences demonstrate that longer-lived individuals can significantly influence change within their group.
Relevant Behavioral Principles
The elderly often exhibit behaviors fundamental to behavioral ecology. This section provides background on several of these concepts that frequently appear in literature on the elderly and thus are basic to understanding the broader nature of the roles of elderly individuals.
Altruism
Altruism, defined by Wilson (1975) as "self-destructive behavior performed for the benefit of others," is relevant to understanding the role of the elderly in nature in several social contexts, including care-giving, allomothering, group defense, and nest care. Altruism is also apparent among the elderly in some nonsocial species. For example, altruism is demonstrated by older, foul-tasting Saturnid moths that sacrifice themselves to "teach" predators that younger counterparts are repugnant (Blest, 1963).
Hamilton (1964) proposed the primary theoretical background for explaining the evolution of altruistic behavior. Hamilton's theory based altruism on the
fundamental kinship relations of donor and recipient (Rosenberg, 1992). Altruism has since been generalized to include any form of nonselfish behavior and serves as the cornerstone for much of sociobiology (Wilson, 1975). In this sense, altruistic behavior includes care-giving to kin other than an individual's direct offspring. Feeding, protecting, warning, defending, and teaching kin, in proportion to their consanguinity, emerges as an adaptationally optimal strategy for older individuals.
Many forms of altruism are evident within social species. Maxim (1979) lists four altruistic roles of aging in primates: (1) defense of territory, (2) defense against predation, (3) control of troop movement, and (4) aunt behavior. These activities are of little or no direct benefit to the elderly individuals but do serve to directly benefit the kinship community. Further, Medawar (1957) comments that altruistic behavior of grandparents should be indirectly selected for because, although grandparents are infertile and their behavior is of no benefit to themselves, "grandmotherly indulgence" does benefit the community. Hamilton (1966) also proposed that postreproductive life spans may have evolved when the older animal benefits its younger kin. For example, old postreproductive female langurs ardently defend their troops and protect troop infants to a much greater degree than younger females (Hrdy, 1981). Also, Norris and Pryor (1991) suggest that many postreproductive, yet still lactating, females in dolphin societies may be present as a source of nourishment to offspring of younger mothers while those mothers dive to great depths for prey. Diamond (1996) supports the occurrence of altruistic postreproductive females by stating that aging human females can, evidently, "do more to increase the number of people bearing her genes by devoting herself to her existing children, her potential grandchildren, and her other relatives than by producing yet another child."
An example of altruism in a nonsocial species is given by Blest (1963). Blest found that cryptic New World Saturnid moths, which rely on camouflage to escape detection but are quite palatable to predators, die shortly after completing reproduction. The aposematic subgroup of New World Saturnid moths, which rely on their terrible taste to discourage predators, exhibit a long postreproductive period. Postreproductive survival of the tasty moths is detrimental to conspecifics because a predator can establish a search image of the cryptic species by feeding on palatable individuals. However, a predator's unpleasant experience with an aposematic individual would benefit its kin and other conspecifics. Thus, Blest argued that the sustained physical fitness of postreproductive aposematic moths contributes to the survival of the younger reproductives.
Dominance and Leadership
To dominate is to possess priority of access to the necessities of life and reproduction (Wilson, 1975). Understanding the governance of animal societies by dominance hierarchies provides the fundamental basis for studying the behav-
ioral dynamics of social groups. Because social groups are often headed by older individuals, such as in primate societies, it follows that understanding dominance relations is basic to understanding the role of the elderly in nature.
Allee (1931) addressed the issue of stable hierarchies and argued that dominance hierarchies created cooperative groups that were better able to compete with other groups. One of the classic studies on chickens, from which the concept of ''pecking order" was derived, was by Guhl and Allee (1945), who found that chickens with stable hierarchies had individuals that consumed more food and laid more eggs than chickens without hierarchies. Dugatkin (1995) provides further support to this study by demonstrating that subordinate chickens do not challenge superiors but often prefer to wait their turn. Dugatkin shows that this complacency occurs not because subordinates fear damage and retribution but that group selection has "chosen" genotypes that will not contest authority. Closely related to dominance is leadership, which is initiative and control of an activity, usually by one individual. Whereas dominance alone suggests suppression of group activity, even though it may stabilize the social interactions, an individual who is also a leader can contribute to group survival by synchronizing and stimulating group activity (Greenberg, 1947).
Wilson (1975) lists several "special properties" of dominance order that have important bearing on understanding the role of the elderly in social species. First, "the peace of strong leadership" refers to how dominant animals of some primate societies, including those of gorillas, chimpanzees, macaques, spider monkeys, and squirrel monkeys, use their power to terminate fighting among subordinates. Species organized by despotisms also live in peace due to the universally acknowledged power of the tyrant (e.g., the queen bumblebee). Second, "social inertia" refers to the intrinsic stability of a dominance hierarchy. An animal that attempts to change its position in a fixed dominance hierarchy is less likely to succeed than if it made the exertion during the formative, fluid stages of the hierarchy (Guhl, 1968). Third, societies structured as "nested hierarchies" are partitioned into units and exhibit dominance both within and between components. For example, in some primate societies, hierarchies exist both within and between family lines.
Territoriality
A territory is an area occupied either directly by overt defense or indirectly through advertisement (Wilson, 1975). This area usually contains a scarce resource (e.g., steady food supply, shelter, space for sexual display, a site for laying eggs or bearing offspring, etc.). The adaptive advantage of territorial behavior is that successful defense of a territory increases the chances of an individual to secure its necessary share of environmental resources, to mate, and to increase survival of offspring (Kluijver and Tinbergen, 1953). Individuals resident in an area have more to gain from retaining the territory than do intruders from taking
it. This is true because residents invest a great deal of energy and time into mastering their area and resources and may well have dependent young. A challenger is devoid of these properties and thus has nothing to lose (Gosling, 1982).
Age is virtually never the sole criterion for achieving dominance or acquiring territory. However, there is likely to be a strong association with age because of two factors. First, the territory holder almost always triumphs in one-on-one encounters. Therefore, territory holders often have held their territories for a long time and thus tend to be older. Second, there is a positive relationship between age and dominance of subordinates, who are continually competing for "next-inline" status.
Learning and Culture
Learning, defined as any change of behavior of individuals due to experience (Cavalli-Sforza and Feldman, 1981; Alcock, 1993), is found in virtually all animal species from the simplest bacteria and protozoa to complex mammals, such as cetaceans, primates, and elephants. The mechanisms of learning can occur at different levels. Self-teaching occurs when a young individual eats a food that tastes bad and subsequently avoids it. Birds acquire songs and repertoires through imitation. Teaching occurs when inexperienced young of predator species accompany their parents on a hunt to acquire skill. The cumulative learning experiences of groups constitute unique cultures that can often be transmitted from one generation to the next (Alcock, 1993). It is widely believed that selection favors specific learning abilities as solutions to specific ecological problems (Alcock, 1993). For example, solitary animals gain useful flexibility by adjusting their diets to whatever nourishment is available. Also, many animals that hoard food must remember where their food cache is located. The ability of social animals to adjust to the behavior of others (e.g., who is dominant to whom, what one individual did to another individual, etc.) may greatly affect their reproductive success (Alcock, 1993). Information that can only be transmitted through learning includes home range, migratory pathways, location of food sources, tool construction and use, acceptability of food items, parental-care skills, and social relationships, such as pair bonding and alliances (Bonner, 1980).
The Elderly In Nature: Selected Case Studies
Although there are few papers published on the elderly in nature, the biological literature contains a surprising amount of information on nonhuman elderly that has never been compiled and synthesized. The life history and field biology of a large number of endangered or economically important species in all major taxonomic groups has been studied and documented in meticulous detail. This includes longitudinal studies of marked individuals or monitoring, for decades,
individuals in laboratories and in zoos. The purpose of this section is to introduce the concept of the role of the elderly in nature, using information extracted from the biological literature on elephants, cetaceans, and primates. We begin each subsection with background information on the biology and social structure of each group to provide ecological context for the information on aging and the roles of the elderly.
Elephants
Ecological Background
Elephants belong to the order Proboscidea, which includes the extinct mastodon and mammoth. There are only two extant species, the African elephant and the Asian elephant, although there are a number of subspecies within each group (Estes, 1991). The African forest elephant lives in the West African rain forest, whereas the savanna race inhabits lowland forests, low-lying swamps, flood plains, and woodland. Elephants also live in subdesert terrain where gallery forest or neighboring mountains provide shade, browse, and water. These animals must frequently move large distances for food and water due to seasonal variations in food distribution and abundance. Elephants are mixed feeders, concentrating on grasses in the rainy season and on woody plants in the dry season (Buss, 1961, 1990). Thus, they wander widely across the savanna during the rains, yet remain near forest and water sources at other times.
Both African and Asian elephants appear to have identical social structures, of which the family is the core unit (Figure 8-2). The family consists of closely related females, including the matriarch, several younger, yet mature, females, and their calves (Eltringham, 1982). Juvenile males remain with their group until they reach sexual maturity, at which time they are driven away by the matriarch (Buss, 1990). Adult males are either solitary or remain on the distant periphery of the herd. Elephant populations are migratory within large home ranges. Activity, direction, and rate of movement are determined by the matriarch, which is typically the largest (and oldest) cow. Typically, the matriarch leads the group, while another large (and old) female brings up the rear (Douglas-Hamilton, 1987). If disturbed, the group clusters around the matriarch awaiting instruction and protection. If the matriarch is shot, the family becomes a frantic, disorganized mob and, most often, falls prey to the same fate as their leader (Eltringham, 1982). Female leadership and experience play a crucial role in elephant social organization, and thus, female life span extends well past the reproductive years (Laws. 1970).
Elephants do not exhibit seasonal breeding (Poole and Moss, 1989). A female elephant is in estrus for only 3 to 6 days at a time and may conceive only once every 3 to 9 years (Poole and Moss, 1989). Thus, given the short female receptive period and the high investment in each offspring, it is crucial for a
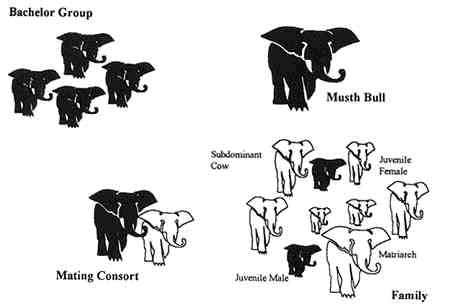
Figure 8-2
The social organization of elephants includes a central family core consisting of the matriarch, other adult females, sexually mature and immature female offspring, and sexually immature juvenile males. The sexually mature juvenile males roam in isolated bachelor groups. Adult males will form mating consorts with adult females. Old musth bulls remain solitary and away from the family unit.
female to find a high-quality mate. Usually a female will prefer to mate with an old male that will chase away the younger males and allow the female to rest, a necessity for successful elephant reproduction (DiSilvestro, 1991).
Reproduction, Life Span, and Aging
Females are sexually mature at 10 years, the gestation period is 22 months, and interbirth intervals last up to 9 years (Buss and Smith, 1966). The close bond between mothers and their offspring can endure for 50 years. Males reach sexual maturity at 17 years (DiSilvestro, 1991), at which time they leave the family unit. At 30 years, a male is almost full size (3.7 meters at shoulder) and first experiences an annual condition of heightened sexual motivation, musth (Owens and Owens, 1992). There is only one musth male in an elephant population at one time because musth males are dominant and can suppress this breeding condition in other males (DiSilvestro, 1991).
Although reproduction is continuous throughout an elephant's lifetime (Laws et al., 1975), the peak of female elephant fecundity occurs at 30-40 years and
steadily declines thereafter to 37 percent of the peak at 46-50 years, 25 percent at 51-55 years, and 0 percent after 55 years (Laws, 1966). Laws et al. (1975) estimated menopause in African elephants at 55 years, although Sukumar (1989) reported a pregnant 60-year-old cow.
The prime of life for an elephant is about 30-40 years (DiSilvestro, 1991). Old age begins around 50, and most elephants die by 60 (Fatti et al., 1980), although some have lived into their 70s (Freeman, 1980). Sikes (1967, 1971) believes that the upper limit of life span is 65 years and is determined by molar-teeth deterioration, so that old individuals cannot eat and survive. Elephants past 50 years usually have the temporal region of the skull depressed and sunken. This physical trait is a sign of old age but not an indication of failing health.
Major Roles of the Elderly
The first major role of the elderly in elephant populations is leadership. Elephant size determines dominance and is also age-dependent because elephants continue to grow throughout their lives. Therefore, older females tend to be the largest of their sex in the herd and, because elephants' social organization is matriarchal, the oldest females are usually the leaders (DiSilvestro, 1991; Redmond, 1991). Despite disease and deterioration, aging females still have an important role in elephant society, and the movements of the herd are adaptable to an aging animal (Freeman, 1980). If the matriarch is injured or killed, the family unit is lost and the animals are easily disbanded or fall prey to the same fate (e.g., poaching) in their confusion (Eltringham, 1982). When the family is approached by predators or other threatening situations, if the matriarch runs, the family runs; if she attacks, the family attacks (Hanks, 1973, 1979). The matriarch will also stand guard over any injured animal in her group, roaring loudly over the body while attempting to upright the animal.
Laws et al. (1975) noted that because of the overwhelmingly matriarchal structure of elephant populations, selective culling of old or barren cows is probably disadvantageous. It removes from the population the adult females with the lowest reproductive rate, which may be helpful for population control but, more seriously, removes the leaders. The elimination of their accumulated experience must add appreciably to the disturbance factor. Moreover, the removal of matriarchs may well lead to formation of larger groups with unfavorable effects on the habitats due to more intensified localized use.
The second major role of the elderly in elephant populations involves reproduction in males. Older males tend to be physically larger and possess larger, heavier tusks (Laws, 1966; Poole, 1987, 1994). Both traits have major influence on male mating success. Females prefer to mate with old bulls because consequently, offspring will inherit genes of long-lived, healthy adults, and old musth bulls will protect her from the harassment of young bulls (Lawley, 1994). After their matings, the older bull stays with the younger cow and guards or escorts her
until the end of her estrous cycle, which provides her with some days of calm, free from male attentions, in which she hopefully will conceive (Owens and Owens, 1992; Lawley, 1994).
The importance of old bulls to the success of elephant reproduction eluded many early observers of elephant behavior when considering the impact of the harvest of the "big tuskers" in ivory trade (DiSilvestro, 1991). The old bulls no longer associated with the herd were interpreted as past their prime. However, they were in their prime, although solitary as almost all adult males are, and in search of estrous females from many different herds (Poole and Moss, 1981; DiSilvestro, 1991).
The third major role of elderly elephants involves essentially the collective knowledge of the matriarchs. In peaceful times, the matriarch decides where the family goes, when to move, and where and when to sleep. Seasonal movements are guided by the matriarch because she has retained knowledge of the optimal or long-forgotten food and water reserves from her long life (Shoshani, 1991). This she teaches to the younger animals (Eltringham, 1982). The solution to many problems of elephant daily life are learned rather than instinctual, making the matriarch, above all, the ultimate repository of essential survival skill that younger animals simply would not have had time to learn on their own (Lawley, 1994). As Laws et al. (1975) notes, "We feel that the dependence of a family unit upon its matriarch even in the most extreme of situations illustrates the great importance of the acquisition of experience and learning made possible by the evolution of longevity. We consider that this situation represents an evolutionary development that is now probably essential for the stability of elephant communities." The matriarch is a reservoir of "collective wisdom of the community" (Eltringham, 1982).
Primates
Ecological Background
Nonhuman primates are composed of 185 species comprising four main branches: prosimians (including lemurs, lorises, pottos, and bush babies), New World monkeys (including capuchins, marmosets, and night squirrel monkeys), Old World monkeys (including mandrills, guenons, baboons, and colobus monkeys), and great apes (including chimpanzees, orangutans, and gorillas) (Estes, 1991). With few exceptions (e.g., macaques and langurs in temperate Japan and China) primates are confined to the tropics, and 80 percent live in rain forests (Estes, 1991). Most nonhuman primates are arboreal or cliff-dwelling; only the gorilla sleeps on the ground (Estes, 1991).
Small primates, such as pottos and bush babies, feed primarily on small arthropods, but larger ones must rely on new leaves, buds, shoots, seed pods, and fruit. All monkeys and apes are sociable, and the majority live in female-bonded
groups. Females never leave their home group, whereas males emigrate and have to join or take over another group before they can reproduce (Wrangham, 1980). Consequently, primate groups are typically female-biased (Clutton-Brock and Harvey, 1983; Clutton-Brock et al., 1977).
Mating systems differ among primate species (Figure 8-3). For example, langurs form large groups of unrelated individuals, consisting entirely of females and also one male that has dominant reproductive access. Other males remain on the periphery of the group awaiting a chance to challenge the dominant male. However, chimpanzees form groups that include more than one male that has access to breed. Within the large group, there is a coalition of several males that roams overlapping female ranges. Each female and her young offspring occupy a distinct range. This mating system allows males to have access to more females, yet no single male has ultimate control. Most primates breed relatively late in life, give birth to only one offspring at a time, have long gestation periods, and, in some species, breed at only one particular time of year.
Dunbar (1988) summarizes the six main types of social organizations in primates that include: solitary territorial animals, monogamous pairs, matrilines, one-male territorial troops, multi-male societies, and one-male harems. First, solitary territorial animals, such as the pottos, inherit their home ranges from their mothers. Female offspring often settle near their mother, who may donate part of her home range to successive female offspring. Males roam from female range to female range. Second, for monogamous primates (e.g., one lemur species and
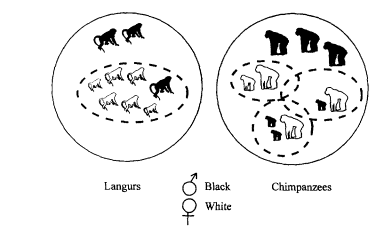
Figure 8-3
Mating systems differ among primate species. Langur females live in small groups, and a single male may be able to prevent other males from joining the group. In chimpanzees, brothers defend a large joint territory, giving them access to more females, each of whom, with her juvenile offspring, lives in her own separate range within the larger territory.
possibly one guenon species), an obligatory pair bond is essential for offspring survival. Third, matrilines, or female kinship groups, consist of families of closely related females and one reproductively dominant male. Females forage separately at night but sleep together with the male during the day. This form of social organization is common in bush babies. Fourth, one-male territorial troops prevail among higher primates and are common in the blue monkey and black-white colobus. The resident male is the dominant member of the troop, and members may not necessarily be genetically related. Fifth, multi-male societies. typified by the savanna baboon, are found in species that forage in large troops in undefended home ranges. Social relations are complex and reflect kinship bonds, male coalitions, and male-female alliances. Finally, one-male harems are exemplified best in the hamadryas baboon and the gorilla. In this system a male may own two or more females for a period of time and remain in a separate bachelor group otherwise.
Reproduction, Life Span, and Aging
Most primates breed relatively late in life, give birth to only one offspring at a time, have long gestation periods, and, in some species, breed at only one particular time of year. A study by Caro et al. (1995) revealed that a significant postreproductive period in old females occurs in most nonhuman primate species. especially gorillas and chimpanzees.
The life spans of most nonhuman primates have been estimated conservatively, in that the recorded age represents the time in captivity. Thus the ages of animals captured in the wild are underestimated by their age at entry into captivity; the ages of those still living are underestimated by the time they will continue to live. Harvey et al. (1987) provide maximum longevity records of over 100 major primate species including the following: long-haired chimpanzee (Pan troglodytes; 44.5 yr), gorilla (Gorilla gorilla; 39.3 yr), (Cercopithecus aethiops; 31 yr), stumptail macaque (Macaca arctoides; 30 yr) hanuman langur (Presbytis entellus; 20 yr), and mouse lemur (Microcebus murinus; 15.5 yr). Cutler (1976) notes that the maximum life span potential of primates is related to their postnatal developmental rate, length of general vigor, length of reproductive period, and time available for learning and teaching.
It is difficult to determine the age of primates in the wild, although the very old and senile individual can be identified by erosion and tartarization of teeth, deepening creases in facial skin, balding, changes in skin pigmentation and hair color, and weight loss (Hrdy, 1981).
Major Roles of the Elderly
The statement by Hrdy (1981) summarizes the beliefs of many scientists regarding elderly primates: "If one were to ask a primatologist how old animals
behave in the wild the answer might well be 'They don't,' for it has long been thought that very few primates in natural habitats could evade predators and disease long enough to grow old. As a consequence little attention has been given to old animals in the vast literature on primate behavior."
The social rank of macaque females increases with age. For example, a daughter of a high-ranked female may adopt a home range overlapping her mother's range. When the mother dies, the young female gains access to resources from which she would otherwise be excluded (Clutton-Brock, 1991). Some studies show that feeding and reproductive skills also increase with age (Lack, 1968; De Steven, 1978), and the costs of rearing offspring may be greater for inexperienced adults than for older individuals (Clutton-Brock, 1991). Older females tend to be reproductively less active and may lead solitary lives, displaced from prime food sources and roosting sites. In contrast, older females of the gray-cheeked mangabey are fully integrated into the group but tend to be less active in grooming, embracing, and other individual social acknowledgments. Chalmers and Rowell (1971) observed that among old females, grooming decreased, while being groomed increased. Kinship ties in the macaque, Macaca fascicularis, preserve the rank of aging females, and only those that are senile drop in rank (Hrdy, 1981).
There is considerable evidence suggesting that leadership is a primary role of older individuals in primate groups. Old silverback gorillas make troop decisions including when and where to forage, rest, and sleep and arbitrate disputes within their harems (Stewart and Harcourt, 1987). However, the type of leadership is conditional upon the sex, ecological or social context (e.g., foraging, defense, etc.), and the mating system and overall life history of the species. In squirrel and toque monkeys, langurs, and gorillas, aging females are the most active in leading the troop movements.
A second important role of elderly females is care-giving. In geladas, aging females maintain their rank in association with their related female groups (matrilines) and act as "aunts" cuddling, carrying, and disciplining others' infants (Maxim, 1979). Researchers discovered that the presence of the grandmother of female vervets (small gray African monkey) in a captive socially living colony had a profound impact on the mortality rate of newborns. Specifically, (1) the presence of the infant's grandmother determined the level of risk; and (2) infant mortality for female vervets with grandmothers present was only 32 percent, whereas infant mortality for vervets without their grandmothers present was over double at 73 percent (Fairbanks and McGuire, 1986). In these circumstances, selection could be expected to favor females that terminate their reproductive life before the natural mortality rate rises steeply, particularly if reproduction would substantially increase the risk of mortality.
Even though the reproductive fitness is low in the elderly, their physical fitness may be relatively high, and thus their contributions to the group to which they belong may be in ways other than reproduction. For example, Hrdy (1981)
suggested that in Japanese macaques the rank of the older females has a great potential effect on her genetic contribution to subsequent generations because her daughters inherit her rank. She referred to these as the "nepotists." Despite increasing feebleness with age, old females in high- and middle-ranking lines are still deferred to by their younger and more vigorous descendants (Figure 8-3). In contrast to the nepotistic system of macaques, hanuman langurs of India have a different breeding system, where females are more closely related to one another than females in a macaque troop. Hrdy (1981) showed that older, postreproductive langurs are low ranking but participate vigorously in defending the troop, advancing troop interests, and protecting and caring for infants and juveniles of close relatives. She referred to these as the "altruists." The concept of inclusive fitness offers an explanation for a ranking system that favors youthful females of high reproductive value combined with a defense system in which old females take great risks (Hamilton, 1964).
The rise and fall by age in the dominance hierarchy of three adult male chimpanzees from the long-term study by Goodall (1986) is shown in Figure 8-4. Several points merit comment. (1) It was not possible for either a young or a female chimpanzee to occupy the alpha position. (2) However, age alone does not assure a high position within the group. For example, Jomeo lived for 26 years but was always low ranking despite his superior weight and aggressiveness; one individual (Figan) occupied the alpha position for nearly a decade, and Evered occupied the
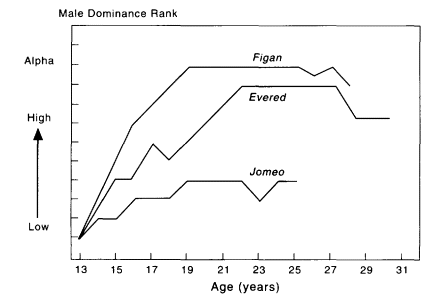
Figure 8-4
The rise and fall in social rank of three adult male Kasakela chimpanzees studied by Goodall (1986) in Gombe National Park, Kenya over a 25-year period.
beta position or a high rank also for nearly a decade. This provides stability to the entire group. (3) Also the beta position was held for a sustained period; the ranking males were almost certainly past their physical prime in their late 20s and early 30s and yet they maintained their rank out of ''habit" and tradition.
One of the most important implications of the detailed longitudinal studies of individuals by Goodall and her coworkers is that they provide critical historical perspective on individuals at their older ages. For example, Goodall (1986) notes that, even when young, Figan showed signs of the qualities that would take him to the top of the male hierarchy. This included exploiting temporary ill health in older males and developing close supportive relationships with members of his family (e.g., high-ranking mother). Evered was motivated to attain high social rank but was at a disadvantage because he had no close ally. In contrast, Jomeo was apparently not motivated to attain high social status despite his apparent physical capabilities.
Cetaceans—Whales and Dolphins
Ecological Background
Cetacea consists of two extant suborders—Mysteceti, baleen whales, and Odontoceti, toothed whales (Carroll, 1988). Although both have evolved from a common archeocete ancestor (Carroll, 1988; Novacek, 1994; Gingerich et al., 1994), they drastically differ not only in feeding morphology but also in behavior (Figure 8-5; Table 8-1). Baleen whales are generally filter feeders, surviving on
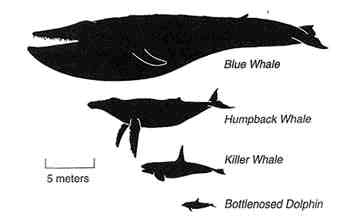
Figure 8-5
Body shape and scale for four species of cetaceans. Baleen whales, (e.g., the blue and humpback whales) tend to be larger than toothed whales (e.g., the killer whale and the bottlenosed dolphin). Not only do these groups differ by size, but also by life history.
TABLE 8-1 Comparative Life-History Traits of the Cetaceans
|
Life-History Trait |
Baleen Whales |
Toothed Whales |
|
Social organization Structure |
None |
Communities with well-defined subgroups |
|
Length of associations |
Fleeting |
Long-lasting |
|
Migration |
Seasonal |
None |
|
Feeding |
Filter feeders |
Active predators |
|
Foraging |
Individual grazing |
Cooperative hunting |
|
Communication |
Only for mating |
Highly developed, multipurpose language |
|
Mating/reproduction |
Seasonal |
Continuous |
|
Care of offspring |
Mother only |
Mother and other adults |
|
Juvenile development |
Rapid |
Normal |
|
SOURCE: Based on Tyack (1986). |
||
small planktonic crustacea and other small oceanic organisms, while toothed whales are active hunters of larger prey, including octopodes, sharks, and other whales. However, a fundamental difference between these groups lies in social organization. Baleen whales have virtually no social structure, and the only relations between individuals (usually mother-calf bonds or sexual encounters) are short-lived, never lasting more than 1 year and more often lasting only several hours (Tyack, 1986). In contrast, toothed whales generally live in complex social communities, consisting of adult females and young of both sexes, while coalitions of adult males travel between communities for access to females (Tyack, 1986; Wells, 1991). Other behavioral differences between the suborders, including migration, foraging, communication, mating, and offspring care, can be seen as artifacts of their social organizations. Therefore, analysis of the dichotomous life histories of baleen and toothed whales may give insight into why they live as long (or as briefly) as they do, and what the elderly members of their species contribute to the population.
Aging and Life Span
Age of cetaceans is usually determined by counting the annual layers of cementum or dentine in the teeth (Kasuya and Marsh, 1984), but this may be unreliable due to bone deterioration, especially in baleen whales (Tyack, 1986). However, the main obstacle to determining age of these animals is lack of specimens to study. Some detailed whaling records are kept, but for the most part, these are either inaccurate or not concerned with age. The only other source of information is from dead cetaceans beached on shore or recent captive specimens (Hayley, 1978). Thus, until recently when researchers have begun to track marine organisms for extended time periods, there have been only sketchy estima-
TABLE 8-2 Developmental Parameters of the Three Main Groups of Cetaceans
|
Parameter |
Baleen Whale |
Toothed Whale |
Dolphin/ Porpoise |
|
Gestation period |
11 mo |
15-16 mo |
10.5-12 mo |
|
Intercalf interval |
1-2 yr |
3-5.8 yr |
1-3 yr |
|
Weaning |
|
|
|
|
Age |
7-11 mo |
Mean 1-5 yr |
6-20 mo |
|
|
|
Max 13-15 yr |
|
|
Length |
8.8-12.8 m |
2.7-7 m |
1-2 m |
|
Sexual maturity |
|
|
|
|
Age |
4-5 yr |
13-17 yr |
2.5-11.5 yr |
|
Length |
11.8-23.3 m |
3.7-10.8 m |
1.4-2.4 m |
|
Adult |
|
|
|
|
Maximal age |
82 yr |
54.5 yr |
27.5 yr |
|
Maximal length |
31 m |
15 m |
3.8 m |
|
Note: For species in which male and female data were supplied, the average was used. SOURCE: Adapted from Tyack (1986: Table 1 ) baleen whale maximum age is averaged from Table 8-3 in this paper. |
|||
tions of life span. However, these rough estimates are sufficient for examining general overall trends.
Specifically, investigators have attempted to define several life-history parameters for both baleen and toothed whales (Table 8-2). Baleen whales reach a length and age that is almost twice that of toothed whales. The ratio of longevity to length clearly increases from baleen whales to toothed whales and then increases dramatically in dolphins (Table 8-3). Specifically, the ratio of longevity to length ranges from 1.5 yr/m to 2.6 yr/m in baleens, from 1.8 yr/m to 2.9 yr/m in toothed whales, and reaches a maximum of 3.8 yr/m in the striped dolphin. The sperm whale (classified Odontoceti) shows a value much closer to those of Mysteceti, consistent with recent studies indicating sperm whales might be an intermediate of the two suborders (Milinkovitch et al., 1994).
Major Roles of the Elderly
It seems that toothed whales and, specifically, dolphins live much longer than would be predicted from their length. Thus, it can be construed that the occurrence of elderly organisms within odontocete communities is probably much greater than among the solitary baleen whales. From a fitness perspective, these old organisms would not seem to contribute to the reproductive fitness of a population; however, this may not be the case.
The sociality of toothed cetaceans has lead to the development of a specialized division of labor among the age classes. Therefore, it is apparent that the
TABLE 8-3 Relation of Longevity to Length in Cetaceans
|
Species |
Common Name |
Maximum Longevity (yr) |
Length (m) |
Ratio (yr/m)a |
|
Baleen whales |
|
|
|
|
|
Balaenoptera acutorostrata |
Minke whale |
47 |
28 |
1.7 |
|
Balaenoptera borealis |
Sei whale |
74 |
47 |
1.6 |
|
Balaenoptera musculus |
Blue whale |
110 |
67 |
1.7 |
|
Balaenoptera physalus |
Fin whale |
114 |
45 |
2.6 |
|
Eschrichtius robustus |
Gray whale |
70 |
46 |
1.5 |
|
Megaptera novaeangliae |
Humpback whale |
77 |
46 |
1.7 |
|
Toothed whales |
|
|
|
|
|
Globicephala macrorhynchus |
Pilot whale |
49.5 |
17 |
2.9 |
|
Orcinus orca |
Killer whale |
50 |
24 |
2.1 |
|
Physeter catadon |
Sperm whale |
77 |
44 |
1.8 |
|
Dolphins |
|
|
|
|
|
Stenella coeruleoalba |
Striped dolphin |
34 |
9 |
3.8 |
|
Tursiops truncatus |
Bottlenosed dolphin |
35 |
9.8 |
3.6 |
|
aThis ratio was calculated by dividing longevity by length. Note: For species in which male and female data were supplied, the average was used. SOURCE: Hayley (1978); pilot whale longevity from Leatherwood and Reeves (1978): data for striped dolphin length from Reilly (1978). |
||||
older animals in a group perform specific tasks not done by the younger animals. However, this is not apparent in the relatively less intelligent baleen species, in which parental care of young and social learning of offspring is not emphasized (Tyack, 1986). There are several roles that elderly odontocetes perform in their societies: guardian, aunt, and nurse.
First, both old male and old female dolphins are documented guarding or "baby-sitting" younger animals in their home groups. For example, in bottlenosed dolphins, an older male will not just pair with a juvenile, but will watch over that juvenile while the rest of the group forages (Wells, 1991). In a study by Johnson and Norris (1994), a very old Hawaiian spinner dolphin male was frequently seen with juveniles and in shore coves while the rest of the herd cruised the coast. This behavior suggests alloparenting because the old male and juveniles rest while most adults forage for food. This pairing can benefit the young dolphins in several ways: the young are protected from predators that wait to attack solitary members of the group, they are monitored so that they do not swim to unfamiliar waters and lose track of the herd, and possibly, they are taught adult behaviors from the old, experienced male. Norris and Pryor (1991) assert that a pilot whale's "principal biological contribution might be to learn, remember, and transmit what pilot whales need to know." By exercising these parent-like duties, the older male is allowing the younger parents to forage, an activity that the old male may not be able to do as efficiently. Therefore, there is a reciprocity in work allocation that requires the active involvement of all age classes (Tyack, 1986).
Old females in a group perform similar child care duties as old males. Specifically, elderly female dolphins act as "aunts" to other dolphin mothers before, while, and after a calf is born. During the birth, the aunt will be a midwife and even break the umbilical cord. After the calf is born, the aunt remains with the mother and calf, serving as a guardian to the young dolphin when the mother forages and rests (Tyack, 1986). In Atlantic bottlenosed dolphins, the grandmother will remain close to her daughter during pregnancy and assist in childbirth. Once the calf is born, the grandmother becomes a guardian to guide the infant through its early years (Caldwell and Caldwell, 1966). In killer whales, older females are thought to nurture the young of other females, creating a matriarchal bonding system (Brault and Caswell, 1993). Also, the oldest female in a Hawaiian spinner dolphin pod acts as a protector, herding the young away from dangerous obstructions (Johnson and Norris, 1994).
The high prevalence of postreproductive females within dolphin communities is significant. In a study on killer whales, a large proportion of females that had not bred for a long time were suggested to be postreproductive (Bigg, 1982, and cited in Kasuya and Marsh, 1984). Among all species, the only value possessed by the female sex is believed to be the ability to bear offspring. Once a mother has surpassed her reproductive prime, she becomes less and less important until she eventually dies. Therefore, it would seem that a menopausal female or one that has ceased to be fertile would certainly be ostracized from her group. However, within the odontocete cetaceans, postreproductive females play an active part in rearing young.
Not only do these females help watch, defend, and teach the younger animals, but, in fact, the postreproductive females nourish these young. Postreproductive female Hawaiian spinner dolphins can produce milk, enabling them to help the younger mothers feed their calves (Johnson and Norris, 1994). Also, in pilot whales, females have an exceptionally long postreproductive period, ceasing ovulation at about 40 years, although they often live until age 63. In Japanese fisheries, 25 percent of females captured are postreproductive and 25 percent of the postreproductive females are lactating. This information implies that females have switched their energies from bearing offspring to nurturing the ones already born, although whether these cetaceans suckle only their own offspring is unclear (Tyack, 1986). Continuous lactation of postreproductive pilot whales allows these whales to nurse other mothers' calves, which are unable to make the deep dive for food that is so characteristic of this species (Norris and Pryor, 1991). Thus, nonreproducing, yet lactating, females can be an important reservoir of nourishment for the young in the community, allowing younger mothers to put more energy into group activities in which older mothers may be less efficient.
In short, contrary to widely held beliefs, older animals that have passed their "prime" do exist in the wild and not just in extraordinary circumstances. This is evident in Cetacea, where the older males and females are essential to the optimal function of their community.
Extended Longevity As A Preadaption For Eusociality
Success or failure in evolving particular mechanisms or structures often depends simply on the presence or absence of a previously existing structure. physiological process, or behavior pattern that is already functional in another context and available as a stepping-stone to the attainment of a new adaptation. This structure, process, or pattern is referred to as a preadaptation (Wilson, 1975: Hölldobler and Wilson, 1990). Preadaptations are often pervasive forces in the histories of all species, creating multiplier effects—such as fins evolving into limbs, limbs into forelegs, and forelegs into grasping appendages with digits. Preadaptations may reach all the way to brood care and advanced sociality by way of extended longevity—a requisite for the existence of both behaviors. Because of the importance of the concept of preadaptation in general and extended longevity in particular, the objective in this section is to examine the preadaptive role of extended longevity in the context of the evolution of eusociality in wasps.
Evolution of Eusociality in Wasps
Wasps are in the insect order Hymenoptera, which is thought to have evolved about 200 million years ago (Edwards, 1980). This order contains a wide variety of species including the sawflies, parasitic hymenopterans, and most extant social insect species, including social wasps, ants, and bees. Most social wasps, such as the yellow jackets and paper wasps, are in the super-family Vespoidea, the majority of which construct a nest of papery material and progressively provision (i.e., feed as needed) their young. Vespoidea also contains a large number of solitary wasps, such as potter wasps and spider wasps, which first construct nests, find suitable prey, and sting and paralyze the prey. Then these insects bring the immobile prey back to the nest, lay an egg, seal the nest, and then repeat the process. This overall strategy is referred to as mass provisioning.
Extended Longevity in Wasps
One of the prerequisites for the evolution of eusociality in insects is generation overlap (i.e., the parents are alive when the offspring emerge as adults). Indeed, Evans and West-Eberhard (1970) conclude that an important factor explaining the lack of significant social evolution in sphecid wasps is the relatively short female life span, so that the adult lives of mothers and daughters rarely overlap. Significant preadaptations for this overlap include increased female longevity, multiple generations, and adult overwintering. The advantage of adults living throughout the winter is that female wasps can begin nesting earlier in the favorable season and be ready to mate as soon as the winter passes, rather than complete adult development, emerge, and mate as overwintering immatures would do (Matthew, 1991). The fact that virtually all social wasps and bees
Evolutionary Steps to Eusociality
Evans (1958) outlined 13 grades of social evolution in wasps, starting with the simplest and, presumably, most primitive and ending with worker-caste differentiation in large, complex colonies. These grades are said to follow the subsocial route, through which a single foundress gains enough longevity to coexist in the same nest as her female offspring (Hölldobler and Wilson, 1990). The parasocial route to eusociality arises when members of the same generation use a composite nest and cooperate in brood care. Recent evidence suggests eusociality appears to arise far more commonly from solitary or semisocial precursors favoring the familial route (Danforth et al., 1996). For brevity, we collapse Evans' 13 steps into the following five phases (Figures 8-6 and 8-7):
Phase I Solitary The ancestral type for the evolution of social wasps is thought to have been a parasitoid. A parasitoid is a hunter, searching for prey that it then
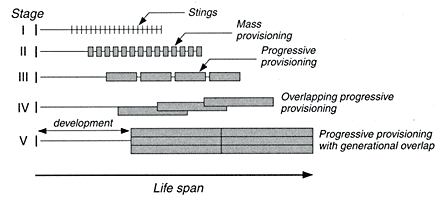
Figure 8-6
Schematic diagram of wasp evolution through five stages. Width of the horizontal bands indicates the duration of the association between parent and offspring. Vertical bands or blocks represent length of association between parent and individual offspring; for example, stage I shows short duration of association between a solitary parasitoid wasp and its offspring. The parasitoid lays eggs in a sequence of hosts but provides no parental care, thus "abandoning" each offspring directly after oviposition. Stage III indicates a longer association with progressive provisioning for each offspring; the parent wasp remains longer with each juvenile. Stage IV shows complete overlapping of generations. Note also that developmental time from egg to adult steadily increases from stage I to V (see also Figure 8-7).
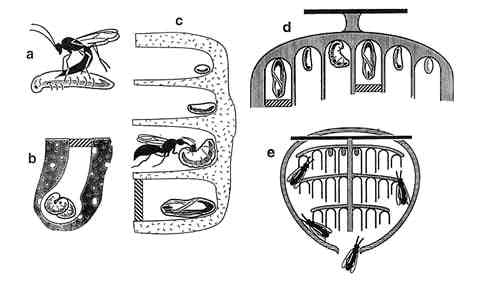
Figure 8-7
Likely stages for the evolution of eusociality in wasps (condensed from Evans, 1958). (a) Solitary parasitoid wasp laying egg in host. (b) Mass-provisioned chamber (paralyzed prey in sealed chamber with wasp egg). (c) Wasp feeding developing larva a piece of masticated caterpillar. (d) Foundress female lays egg in empty chamber and progressively provisions until daughters mature and can help. (e) Eusocial colony with distinct worker castes and age polyethism for nest construction, maintenance, nursing, and defense. SOURCE: a-c adapted from figure in Cowan (1991), d and e adapted from figure 14.43b and figure 14.43c, respectively, in Wenzel (1991).
temporarily paralyzes so that the prey can become a live incubator for its egg. The wasp egg hatches into a larva inside the prey, the larva consumes the prey, exits, and pupates.
Phase II Mass Provisioning An early subphase of mass provisioning occurs when the female paralyzes a prey, carries it to a previously constructed nest, lays her egg, and seals the nest. A later subphase occurs when the female brings multiple prey to the nest. She may have contact with some of her feeding offspring before sealing the nest and after delivering the last prey.
Phase III Progressive Provisioning The female brings more prey to the burrow, as needed, for the developing larva. She may remain in the nest with her offspring, adding some protection for the young. Later, females may begin to feed macerated food directly to the larvae. Here the degree of parental care strongly increases the bond between adult and offspring. The assumption is that progressive provisioning arose from delayed provisioning (Evans, 1966). Among
sphecids, Evans (1966) suggests that progressively provisioning Bembix live two to four times as long as mass-provisioning Gorytes. As females spend longer periods sitting in the protected nest, different selective factors become important. Mortality due to accident and predation is likely to be reduced, there is selection to reduce rates of senescence, life span increases, and mothers are more likely to be alive when some of their offspring reach adulthood (Cowan, 1991).
Phase IV Generation Overlap/Primitive Eusociality The female's life is prolonged and overlaps that of the progeny that remain with her in the nest to form an extended family. Females at this phase lay eggs into an empty cell, a trait thought important in permitting evolution of the extended brood care of social wasps (Evans, 1958; Carpenter, 1991). The young females may construct more cells, lay eggs, and care for their own larvae. However, some females occasionally care for the brood of another female. These behaviors are important steps because the evolution of sociality is not possible without overlap of generations and, at least, rudimentary division of labor. Jayakar and Spurway (1966) believe that the ability to care for more than one cell and larva simultaneously is an important evolutionary step toward eusociality.
Phase V Caste Differentiation Extensive overlap of generations occurs because of extended life spans and because a clear division of labor and caste systems begins to emerge. Wasps have opportunities for fitness gain by associating with relatives of different generations (Cowan, 1991). Differential feeding of larvae occurs. Early progeny are workers, but some intermediate forms are present between these and true females. A single queen, usually the original foundress, is much more long lived than the workers and monopolizes oviposition, referred to as queen control. As long as the colony remains small and the life spans of reproductive individuals and helpers do not differ significantly, helpers may be expected to resist evolutionary specialization as workers because this would reduce or eliminate their chances to replace their mother as the sole reproductive female (Alexander et al., 1991).
As eusocial colonies become large and long-lasting, mothers become increasingly specialized as reproductives, and offspring specialize as workers and soldiers. As this happens, daughters have fewer opportunities to become replacement reproductives, both because the mother lives longer and because so many other individuals are available to replace the mother as well. Thus the identity of a queen's replacement becomes a sweepstakes, with each individual having a chance of, perhaps, one in 50,000 or one in a million (Alexander et al., 1991). One consequence of increased longevity as a preadaptation to sociality is that the stage is set for the evolution of large colonies that have certain advantages over smaller ones, including the ability to organize labor more efficiently, integrate colony activities, defend the nest more aggressively, and exert homeostatic control over physical conditions within the nest (Jeanne, 1991).
Evolution of Life Span in Eusocial Workers
Few uses of the disposable-soma theory of senescence (Kirkwood, 1985) are more apt than when this concept is applied to worker castes in social insects. The fact that honeybee workers are expendable is apparent from their sting and aggressive defensive behavior. Because honeybees' stings are barbed, their stings and adjacent tissues remain anchored in the skin of intruders, thereby creating a chemical alarm that guides other guards to the enemy (Seeley, 1985). Thus the gerontological question is not whether workers are "disposable," but rather how long they should live relative to the "needs" of the colony.
Insights into this question can be gained by considering the relationship of the number of births in a monogynous (single queen) colony (bq) , daily death rate of workers (dw), and maximal colony size (N*) as given in the equation (Carey, 1993):
It is evident from this simple equation that colony size can increase by either increasing daily birth rate or decreasing daily death rate of workers. For example, if bq = 100 eggs/day and dw = 0.10, then N* = 1,000. However, if either dw decreased to 0.01 or bq increased to 1,000, then the maximal colony size, N*, would increase to 10,000 individuals. A more complete range of tradeoffs between birth and death rates for a fixed maximal colony size is given in Table 8-4. Primitive social-insect colonies probably maintain a modest colony size by evolving an increased life span of the as-yet-undifferentiated "worker" caste. This is because high egg production in queens requires a highly evolved system of food provisioning and propholaxis (liquid food exchange). At the early stages of the evolution of eusociality, it is probably easier to extend life span in all colony members. However, in more advanced groups, when queen longevity is several fold greater than workers, the worker life span is probably determined by needs of colony defense, predictability of food resources, and colony-maintenance requirements.
TABLE 8-4 Maximum Size of Social Insect Colonies Given Different Worker Life Expectancies and Queen Daily Egg Production
|
|
Queen Daily Egg Production |
|||
|
Life Expectation |
100 |
1000 |
10,000 |
100,000 |
|
20 days |
2,000 |
20,000 |
200.000 |
2,000,000 |
|
50 days |
5,000 |
50,000 |
500.000 |
5,000,000 |
|
100 days |
10,000 |
100,000 |
1,000,000 |
10,000,000 |
|
200 days |
20,000 |
200,000 |
2,000.000 |
20,000,000 |
|
SOURCE: Carey (1993; tables 5-12). |
||||
Discussion
General Themes
The overall theme of this paper is that, as a subgroup, the elderly play important roles in the cohesion and dynamics of groups, populations, and communities. They serve as care-givers, as guardians, as leaders, as stabilizing centers, as teachers, as sexual consorts, and as midwives. The elderly are at an evolutionary forefront when their brood-care behavior contributes to population fitness, which, in turn, may lead to evolution of sociality in insects and perhaps to other evolutionary pathways in other species. Virtually all behaviors exhibited by the elderly have an evolutionary basis, yet evolutionary aspects of behavior are seldom considered in the context of aging biology. Perhaps the time has come to expand the scope of gerontology and demography of aging to include behavioral to better incorporate the contributions of the elderly to the fitness of populations.
Another theme of this paper, particularly concerned with longevity as a preadaptation for the evolution of eusociality, is that it is important to distinguish between micro- and macroevolutionary aspects of aging and longevity change. Clearly, the evolutionary scale of change for fruit fly selection experiments (Rose, 1991) involving the cost of reproduction is different from changes involving the emergence of major taxonomic groups, new structures, physiological processes, or major adaptive shifts (i.e., eusociality). It is important to recognize differences in evolutionary scales because the macroevolutionary scale provides context for broad differences in life spans, which may shed light on more subtle, microevolutionary differences. Cost of reproduction tradeoffs or metabolic-rate differences may help explain longevity constraints in microevolutionary settings but provide little insight into why, for example, bats and birds live longer than similarly sized mammals. Explanations for these life span differences may have more to do with macroevolutionary causes, such as the evolution of sociality or changes in ecological factors, than with proximate factors, such as reproductive costs.
Biodemographic Perspectives on Life Span
In many species a substantial amount of energy is devoted to offspring care. Demographically, this results in reduction of birth rate. For example, Van Lawick-Goodall's (1976:86) description of the chimpanzee's early life is informative:
The infant does not start to walk until he is six months old, and he seldom ventures more than a few yards from his mother until he is over nine months old. He may ingest a few scraps of solid food when he is six months, but solids do not become a significant part of his diet until he is about two years of age and he
TABLE 8-5 Partial List of Life-History Traits Associated with Extended Longevity in Animals
|
Long gestation or developmental period |
|
Small number of offspring per pregnancy, brood, or litter |
|
Long interbirth intervals |
|
Long period of offspring dependency |
|
Intense parenting or brood care |
|
Strong social bonds |
|
Protracted learning period and increased learning potential |
|
Home base, nest, or territory as locus |
|
Monogamy/pair bonding |
|
Enhanced sibling relationships |
continues to nurse until he is between four-and-a-half and six years old. Moreover, while he may travel short distances...when he is about four years old, he continues to make long journeys riding on his mother's back until he is five or six.
As Lovejoy (1981) notes, the extreme degree of parental investment has profound demographic consequences.
When the basic life-history patterns of humans are considered, noting that chimpanzees often live 30-35 years in the wild and that species with intensive parental care usually live substantially longer than species with little or no parental care, all the evidence suggests that the evolved life span of primitive humans was several fold greater than the life expectancy of 15-25 years (Hayflick, 1994). For example, if the maturation period of early hominids was 15 years, the birth interval was 5 years, and survival to adulthood was 30 percent to 50 percent (even with intensive parental care), then a female must live 35 years just to replace herself and her mate and 50 years to adequately care for her last-born child (i.e., the mother's death would substantially reduce the survival chances of several dependent offspring, and even the survival chances of other members of the group if she is involved in cooperative care of the young). If probability of survival to adulthood was less than 30 percent and/or a percentage of the females were sterile, then the last age of reproduction would increase to 40 years and life span would increase to 55 years. This value is consistent with predictions of life spans based on correlations between longevity, brain weight, and adult size (Hayflick, 1994). Our approach combines biological and demographic logic with perspectives on the life histories of other species. Table 8-5 contains a list of life-history traits that appear to associate with increased longevity in many of these species.
Gerontological and Demographic Implications
The perspectives provided by exploring the natural history and the roles of the elderly in nonhuman populations have the potential to greatly expand the
scope of gerontology and the demography of aging by providing important ecological and biological context for these disciplines. Specifically, we have identified four different demographic and gerontological concepts that will benefit from further inquiry into the role of the elderly in nature. First, perspectives on the natural history of the elderly will provide a broader and deeper biological foundation for life-course analysis in demography, including the integration of concepts in demographic sociology (Riley and Riley, 1986), those in sociobiology (Wilson, 1975), and those in life-history theory (Roff, 1992).
Second, emphasis on research on the elderly in nonhuman species may encourage evolutionary biologists to reevaluate the evolutionary theories of aging and senescence. Theories may need to be reformulated to account for situations where the elderly contribute a disproportionate share of genes to the next generation. Emphasis on the importance of the elderly at the population level will also provide important context for studies concerning adaptive demography or the optimal age/stage/size composition of various social groups (Oster and Wilson, 1978), as well as for the rate of genetic change in age-structured populations (Rose, 1991; Charlesworth, 1994).
Third, the concept ''roles" of the elderly provide an organizational theme for interdisciplinary aspects of aging. For example, retirement has been referred to by social demographers as a "roleless role," lacking content and sure rewards (Riley and Riley, 1986). But social scientists have no biological foundation upon which to construct theories of social roles, just as many biologists have long thought that very few animals in natural habitats could evade predators and disease long enough to grow old.
Fourth, in humans, the role of the elderly at the oldest ages is becoming almost synonymous with the role of elderly women. The aging society is increasingly becoming a society of older women, often widows. Our future studies will examine whether this feminization of the elderly is general, whether there are gender-specific roles of the elderly, and, if so, how and why these exist. Indeed, infant and juvenile care given by older "aunts" appears to be fairly common across many species. Thus, the literature on inclusive fitness theory (Hamilton. 1964) can be brought to bear on the development of aging theory and the behavioral traits of the elderly. These concepts have not yet been introduced to the gerontological literature; they are clearly fundamental to a deeper understanding of how older caregivers help perpetuate their own genes.
Acknowledgments
We thank Lynn Kimsey and Robert Jeanne for their insights and help with wasp evolution, and Claudia Graham and Shin-min Tsai for graphics. We are particularly indebted to the undergraduates who enrolled in our University of California at Davis research course on the role of the elderly in nature: Karen Abalos, Harold Amogan, Sophie Betts, Alicia Cook, Kimberly Day, Hoa Giang,
Sandra Lara, Sarah Mertz, Sakura Nakahara, Phuoc Nguyen, Janel Rodas, Nancy Toy, Jennifer Schmidt, Duc Vo, and Elmer Yee. We also thank Stephen Helfand for his suggestions. The research was supported by the National Institute on Aging (Grant AG08761-01) and a Duke University Pilot Project Program grant from the Center for Demographic Studies.
References
Alcock, J. 1993 Animal Behavior: An Evolutionary Approach, 5th ed. Sunderland, MA: Sinauer Associates, Inc.
Alexander, R.D., K.M. Noonan, and B.J. Crespi 1991 The evolution of eusociality. In P.W. Sherman, J.U.M. Jarvis. and R.D. Alexander, eds., The Biology of the Naked Mole-Rat. Princeton, NJ: Princeton University Press.
Allee, W.C. 1931 Animal Aggregations: a Study in General Sociology. Chicago: University of Chicago Press.
Bigg, M. 1982 An assessment of the killer whale (Orcinus orca) stocks off Vancouver Island. British Columbia. Report of the International Whaling Commission 32:655-670.
Blest, A.D. 1963 Longevity, palatability and natural selection in five species of New World saturniid moth. Nature 197:1183-1186.
Bonner, J.T. 1980 The Evolution of Culture in Animals. Princeton, NJ: Princeton University Press.
Brault, S.. and H.C. Caswell 1993 Pod-specific demography of killer whales (Orcinus orca). Ecology 75(5):1444-1544.
Buss, I.O. 1961 Some observations on food habits and behavior of the African elephant. Journal of Wildlife Management 25(2):131-148.
1990 Elephant Life: Fifteen Years of High Population Density. Ames, IA: Iowa State University Press.
Buss, I.O., and N.R. Smith 1966 Observations on reproduction and breeding behavior of the African elephant. Journal of Wildlife Management 30(2):375-385.
Caldwell, M.C., and D.K. Caldwell 1966 Epimeletic (care-giving) behavior in Cetacea. In K. Norris, ed., Whales, Dolphins, and Porpoises. Berkeley, CA: University of California Press.
Carey. J.R. 1993 Applied Demography for Biologists, with Special Emphasis on Insects. New York: Oxford University Press.
Caro, T.M.. D.W. Sellen, A. Parish, R. Frank, D.M. Brown. E. Voland, and M. Borgerhoff Mulder 1995 Termination of reproduction in nonhuman and human female primates. International Journal of Primatology 16(2):205-220.
Carpenter, J.M. 1991 Phylogenetic relationships and the origin of social behavior in the Vespidae. In K.G. Ross and R.W. Matthew, eds., The Social Biology of Wasps. Ithaca. NY: Comstock Publishing Associates.
Carroll, R.L. 1988 Vertebrate Paleontology and Evolution. New York: W.H. Freeman and Company.
Caswell, H. 1989 Matrix Population Models. Sunderland, MA: Sinauer Associates, Inc.
Cavalli-Sforza, L.L.. and M.W. Feldman 1981 Cultural Transmission and Evolution: A Quantitative Approach. Princeton, NJ: Princeton University Press.
Chalmers. N.R., and T.E. Rowell 1971 Behavior and female reproductive cycles in a captive group of mangabeys. Folia Primatologica 14:1-14.
Charlesworth, B. 1994 Evolution in Age-Structured Populations, 2nd ed. London: Cambridge University Press.
Clutton-Brock, T.H. 1991 The Evolution of Parental Care. Princeton, NJ: Princeton University Press.
Clutton-Brock, T.H., and P.H. Harvey 1983 The functional significance of variation in body size among mammals. Special Publication, American Society of Mammalogists 7:632-663.
Clutton-Brock, T.H., P.H. Harvey, and B. Rudder 1977 Sexual dimorphism, socionomic sex ratio and body weight in primates. Nature 269:797-800.
Coulson, J.C. 1966 The influence of the pair-bond and age on the breeding biology of the kittiwake gull, Rissa tridactla. Journal of Animal Ecology 35:269-279.
Cowan, D.P. 1991 The solitary and presocial Vespidae. In K.G. Ross and R.W. Matthew, eds., The Social Biology of Wasps. Ithaca, NY: Comstock Publishing Associates.
Cutler, R.G. 1976 Evolution of longevity in primates. Journal of Human Evolution 5:169-202.
Danforth, N.J., L. Neff, and P. Baretto-Ko 1996 Nestmate relatedness in a communal bee, Perdita texana (Hymenoptera:Andrenidae), based on DNA fingerprinting. Evolution 50:276-284.
Deevey, E. S., Jr. 1947 Life tables for natural populations of animals. Quarterly Review of Biology 22:283-314.
De Steven, D. 1978 Clutch size, breeding success and parental survival in the tree sparrow (Iridoprocne bicolor). Evolution 34:278-291.
Diamond, J. 1996 Why women change. Discover 17:131-137.
DiSilvestro, R.L. 1991 The African Elephant: Twilight in Eden. New York: John Wiley and Sons, Inc.
Douglas-Hamilton, I. 1987 African elephants: population trends and their causes. Oryx 21:11-24.
Dugatkin, L.A. 1995 Formalizing Allee's ideas on dominance hierarchies: an intrademic selection model. The American Naturalist 146(6):954-960.
Dunbar, R.I.M. 1988 Primate Social Systems. London: Christopher Helm.
Edwards, R. 1980 Social Wasps. Their Biology and Control. East Grinstead, England: Rentokil Limited.
Eltringham, S.K. 1982 Elephants. Poole: Blandford Press.
Estes. R.D. 1991 The Behavior Guide to African Mammals, Including Hoofed Mammals, Carnivores, and Primates . Berkeley, CA: University of California Press.
Evans, H.E. 1958 The evolution of social life in wasps. Proceedings of the 10th International Congress on Entomology, (Montreal, 1956) 2:449-457.
1966 The Comparative Ethology and Evolution of the Wasps. Cambridge, MA: Harvard University Press.
Evans, H.E.. and M.J. West-Eberhard 1970 The Wasps. Ann Arbor, MI: University of Michigan Press.
Fairbanks, L.A., and M.T. McGuire 1986 Age, reproductive value, and dominance-related in vervet monkey females: cross-generational influences on social relationships and reproduction. Animal Behavior 34:1710-1721.
Fatti, L.P., G.L. Smuts, A.M. Starfield, and A.A. Spurdle 1980 Age determination in African elephants. Journal of Mammology 61(3): 547-551.
Finch, C.E.. and R.E. Ricklefs 1991 Age structure of populations. Science 254:779.
Freeman, D. 1980 Elephants: the Vanishing Giants. Hong Kong: Bison Books Limited.
Gingerich. P.D., S.M. Raza, M. Arif, M. Anwar, and X. Zhou 1994 New whale from the Eocene of Pakistan and the origin of cetacean swimming. Nature 368:844-847.
Goodall, J. 1986 The Chimpanzees of Gombe. Cambridge, MA: Belknap Press.
Gosling, L.M. 1982 A reassessment of the function of scent marking in territories. Zeitschrift fur Tierpsychologie 60:89-118.
Greenberg, B. 1947 Some relations between territory, social hierarchy, and leadership in the green sunfish (Lepomis cyanellus). Physiological Zoology 20:267-299.
Guhl, A.M. 1968 Social inertia and social stability in chickens. Animal 16(2.3):219-232.
Guhl, A.M., and W.C. Allee 1945 Some measurable effects of social organization in flocks of hens. Physiological Zoology 17:320-347.
Hamilton, W.D. 1964 The genetical evolution of social behavior. Journal of Theoretical Biology 7:1.
1966 The moulding of senescence by the natural selection. Journal of Theoretical Biology 21(1):12-45.
Hanks, J. 1973 Population dynamics of the African elephant (Loxodonta africana). Journal of Zoology (London) 169:29-38.
1979 The Struggle for Survival: The Elephant Problem. New York: Mayflower Books.
Harvey, P.H., R.D. Martin, and T.H. Clutton-Brock 1987 Life histories in comparative perspective. In B.B. Smuts, D.L. Cheney, R.M. Seyfarth, R.W. Wrangham, and T.T. Struhsaker, eds., Primate Societies. Chicago: University of Chicago Press.
Hayflick, L. 1994 How and Why We Age. New York: Ballantine Books.
Hayley, D.H. 1978 Cetaceans. In D. Hayley, ed., Marine Mammals of Eastern North Pacific and Arctic Waters. Seattle, WA: Pacific Search Press.
Hölldobler, B., and E. O. Wilson 1990 The Ants. Cambridge, MA: Belknap Press of Harvard University Press.
Hrdy, S.H. 1977 The Langurs of Abu. Cambridge, MA: Harvard University Press.
1981 "Nepotists" and "altruists": the behavior of old females among macaques and langur monkeys. In P.T. Amoss and S. Harrell, eds., Other Ways of Growing Old: Anthropological Perspectives. Stanford, CA: Stanford University Press.
Izquierdo, J.I. 1991 How does Drosophila melanogaster overwinter? Entomolgia Experimentalis et Applicata 59:51-58.
Jayakar, S.D., and H. Spurway 1966 Re-use of cells and brother-sister mating in the Indian species Stenodynerus miniatus (Sauss.) (Vespidae: Eunenidae). Journal of the Bombay Natural History Society 63:378-398.
Jeanne, R.L. 1991 The swarm-founding Polistinae. In K.G. Ross and R.W. Matthews, eds., The Social Biology of Wasps, Ithaca, NY: Comstock Publishing Associates.
Johnson, C.M., and K.S. Norris 1994 Social behavior. In K.S. Norris, B. Wursig, R.S. Wells, and M. Wursig, eds., The Hawaiian Spinner Dolphin. Berkeley, CA: University of California Press.
Kasuya, T., and H. Marsh 1984 Life history and reproductive biology of the short-finned pilot whale. Globicephala macrorhynchus, off the Pacific Coast of Japan. Report of the International Whaling Commission (Special Issue) 6: 259-310.
Kirkwood, T.B.L. 1985 Comparative and evolutionary aspects of longevity. In C.E. Finch and E.L. Schneider. eds., Handbook of the Biology of Aging, 2nd ed. New York: Van Nostrand Reinhold Co.
Kluijver, H.N., and L. Tinhergen 1953 Territory and the regulation of density in titmice. Archives Néerlandaises de Zoologie, Leydig 10(3):265-289.
Lack, D. 1968 Ecological Adaptations for Breeding in Birds. London: Methuen and Company.
Lawley, L. 1994 The World of Elephants. New York: Freidman/Fairfax Publishers.
Laws, R.M. 1966 Age criteria for the African elephant, Loxodonta a. africana. East African Wildlife Journal 4:1-37.
1970 Elephants as agents of habitat and landscape change in East Africa. Oikos 21:1-15.
Laws, R.M., I.S.C. Parker, and R.C.B. Johnstone 1975 Elephants and their Habitats: The Ecology of Elephants in North Bunvoro, Uganda . Oxford: Clarendon Press.
Leatherwood, S., and R.R. Reeves 1978 Porpoises and Dolphins. In D. Haley, ed., Marine Mammals of Eastern North Pacific and Arctic Waters. Seattle, WA: Pacific Search Press.
Lotka, A.J. 1907 Relation between birth rates and death rates. Science 26:12-22.
Lovejoy, C.O. 1981 The origin of man. Science 211:341-350.
Matthew, R.W. 1991 Evolution of social behavior in Sphecid wasps. In K.G. Ross and R.W. Matthew, eds., The Social Biology of Wasps. Ithaca, NY: Comstock Publishing Associates.
Maxim, P.E. 1979 Social behavior. In D.M. Bowden, ed., Aging in Nonhuman Primates. New York: Van Nostrand Reinhold Company.
Medawar, P.B. 1957 The Uniqueness of the Individual, 2nd ed. New York: Dover Publications, Inc.
Milinkovitch, M.C., A. Meyer, and I.R. Powell 1994 Phylogeny of all major groups of cetaceans based on DNA sequences from three mitochondrial genes. Molecular Biology and Evolution 11(6): 939-944.
Norris, K.S., and K. Pryor 1991 Some thoughts on grandmothers. In K. Pryor and K.S. Norris, eds., Dolphin Societies: Discoveries and Puzzles. Berkeley, CA: University of California Press.
Novacek, M.J. 1994 Whales leave the beach. Nature 368:807.
Oster, G., and E.O. Wilson 1978 Caste and Ecology in the Social Insects. Princeton, NJ: Princeton University Press.
Owens, D., and M. Owens 1992 The Eye of the Elephant. Boston, MA: Houghton Mifflin Company.
Poole, J.H. 1987 Elephants in musth, lust. Natural History November: 46-55.
1994 Sex differences in the behavior of African elephants. In R.V. Short and E. Balaban, eds., The Differences Between the Sexes. Cambridge: Cambridge University Press.
Poole, J.H., and C.J. Moss 1981 Musth in the African elephant, Loxodonta africana. Nature 292:830-831.
1989 Elephant mate searching: group dynamics and vocal and olfactory communication. Symposia of the Zoological Society of London 61:111 - 125.
Redmond, 1. 1991 With elephants underground. In R. Orenstein, ed., Elephants: Saving the Gentle Giants. Toronto, Canada: Key Porter Books Ltd.
Reilly, S.B. 1978 Pilot whale. In D. Hayley, ed., Marine Mammals of Eastern North Pacific and Arctic Waters. Seattle, WA: Pacific Search Press.
Riley, M.W. 1979 Introduction. Life-course perspectives. In M.W. Riley. ed., Aging From Birth to Death: Interdisciplinary Perspectives Vol. I. AAAS Symposium 30. Boulder. CO: Westview Press.
Riley, M., and J. Riley, Jr. 1986 Longevity and social structure: the added years. Daedalus 115:51.
Roff, D. 1992 The Evolution of Life Histories. New York: Chapman and Hall.
Rose, M. 1991 Evolutionary Biology of Aging. New York: Oxford University Press.
Rosenberg, A. 1992 Altruism: theoretical contexts. In E.F. Keller and E.A. Lloyd. eds., Keywords in Evolutionary Biology. Cambridge, MA: Harvard University Press.
Seeley, T. 1985 Honeybee Ecology. Princeton, NJ: Princeton University Press.
Shoshani, J. 1991 Last of a noble line. In R. Orenstein, ed., Elephants: Saving the Gentle Giants. Toronto, Canada: Key Porter Books Ltd.
Sikes. S.K. 1967 The African elephant, Loxodonta africana: a field method for the estimation of age. Journal of Zoology (London) 154:235-248.
1971 The Natural History of the African Elephant. New York: American Elsevier Publishing Company, Inc.
Silverstone, B. 1996 Older people of tomorrow: a psychosocial profile. The Gerontologist 36:27-32.
Stewart, K.J., and A.H. Harcourt 1987 Gorillas: variation in female relationships. In B.B. Smuts, D.L. Cheney, R.M. Seyfarth, R.W. Wrangham, and T.T. Struhsaker, eds., Primate Societies. Chicago: University of Chicago Press.
Strickberger. M.W. 1996 Evolution. 2nd ed. Boston, MA: Jones and Bartlett Publishers.
Sukumar. R. 1989 The Asian Elephant: Ecology and Management. Cambridge: Cambridge University Press.
Timiras. P.S. 1994 Physiological Basis of Aging and Geriatrics, 2nd ed. Boca Raton, FL: CRC Press.
Tyack. P. 1986 Population biology, social behavior, and communication in whales and dolphins. Trends in Ecology and Evolution 1(6):144-150.
Van Lawick-Goodall, J. 1976 Continuities between chimpanzee and human behavior. In G.L. Isaac and E.R. McCown, eds., Human Origins: Louis Leakey and the East African Evidence. Menlo Park, CA: Benjamin.
Wells, R.S. 1991 The role of long-term study in understanding the social structure of a bottlenose dolphin community. In K. Pryor and K.S. Norris, eds., Dolphin Societies: Discoveries and Puzzles. Berkeley. CA: University of California Press.
Wenzel. J.W. 1991 Evolution of nest architecture. In K.G. Ross and R.W. Matthews, eds., The Social Biology of Wasps. Ithaca, NY: Comstock Publishing Associates.
Williams, G.C. 1957 Pleiotropy, natural selection and the evolution of senescence. Evolution 11:11-21.
Wilson, E.O. 1975 Sociobiology: the New Synthesis. Cambridge. MA: Harvard University Press.
Wrangham, R.W. 1980 An ecological model of female-bonded primate groups. Behavior 75:262-300.


































