
2 Trajectories of Mortality at Advanced Ages
James W. Vaupel
Benjamin Gompertz (1825) proposed that the force of mortality—the hazard of death—increased exponentially with age for humans, at least as a serviceable approximation over the range of adult ages for which he had data. Various subsequent researchers, especially in biology and gerontology, have viewed Gompertz' observation as a law that describes the process of senescence in almost all multicellular animals at all ages after the onset of reproduction. As a rough approximation at younger adult ages, Gompertz' exponential formula does capture the rise in mortality in a great variety of species (Finch, 1990).
Until recently, it was impossible to determine whether this exponential rise continued to advanced ages. For humans, the scattered data available suggested that mortality decelerated at the highest ages, but questions about data reliability precluded strong conclusions. For other species, virtually nothing was known about mortality at advanced ages because the populations studied had been too small to permit dependable estimates of death rates at ages that only a small fraction of the starting cohort reached (Carey et al., 1992).
A research team of demographers and biologists—including James R. Carey, James W. Curtsinger, Thomas E. Johnson, Vaino Kannisto, Pamela Larsen, Hans Lundström, A. Roger Thatcher, Anatoli I. Yashin, myself, and others—has now succeeded in peering into the remote regions of survival for five species—Homo sapiens, Drosophila melanogaster, Ceratitis capitata (better known as the medfly), Anastrepha ludens (the somewhat larger Mexican fruit fly), and Caenorhabditis elegans (a nematode worm). This chapter summarizes the key research findings that pertain to the trajectory of mortality at advanced ages.
The basic finding is that mortality decelerates at older ages. For some species, such as humans, death rates keep on going up with age up to advanced ages but the rate of increase slows down. For other species, such as medflies, death rates reach a plateau and then fall dramatically. Why mortality decelerates at older ages, in all the species for which large, careful studies have been conducted, is a puzzle. The chapter concludes with an explanation of why this is a puzzle and what might be the key to an answer.
The research reported here is based on collaborative efforts by scientists from different backgrounds. Hence, the chapter constitutes a case study of the topic I was asked to address—namely, "demographers, ecologists, and evolutionary biologists: what can we learn from each other?"
Theories About Mortality At Advanced Ages
Medvedev (1990) reviews more than 300 theories of aging; most could be used to derive testable predictions about the trajectory of mortality at advanced ages. These theories, however, are, for the most part, simply sets of ad hoc assertions derived from or consistent with a few observations and largely based on strongly held prior beliefs. For example, Buffon, the eighteenth century French naturalist, hypothesized that each species has a characteristic maximum life span and that this maximum life span is six or seven times the duration of the period of growth. A long stream of variants on this theme culminated with Fries (1980), who popularized the conjecture that all individuals are born with a genetically determined maximum life span. Individuals who do not die prematurely die of senescence shortly before their fixed allotment of life.
In the 1950s and 1960s, Medawar (1952), Williams (1957), and Hamilton (1966) developed a more cogent theory, based on evolutionary arguments, of why mortality increases at adult ages. Partridge (in this volume) and Rose (1991) review the development of this theory. Charlesworth (1994) summarized it as follows: at older ages "mutation pressure overpowers selection," leading to "mutational collapse." The basic idea is that there will be little or no pressure of evolutionary selection against mutations that have detrimental effects at older ages but neutral or positive effects at younger ages. The elderly have little impact on evolution because only a fraction of a cohort live to old age and few, if any, of them produce offspring. For humans and other species that care for their children and grandchildren, the elderly may have some evolutionary role, but this role diminishes with advancing age. The prediction of this line of thinking is that the age-trajectory of mortality should shoot up at postreproductive ages, especially for species in which parents do not help their progeny (Curtsinger, 1995b).
Until recently, the evolutionary theory of aging largely rested on theoretically plausible arguments that had not yet been empirically tested; Rose (1991) reviews the limited empirical results. Although there have been some important recent experiments (such as Hughes and Charlesworth, 1994: Charlesworth and
Hughes, 1996), the theory still substantially outpaces available observations. Furthermore, even the theory is based largely on reasoned speculation rather than on well-specified mathematical models with compelling implications.
Empirical Findings
Demographers have long been interested in the shape of the age-trajectory of human mortality. The general nature of this trajectory from birth to age 80 or so is well known: mortality is high right after birth, falls to a very low level around puberty, and then rises more or less exponentially except for some excess mortality among adolescents and young adults. Due to the painstaking efforts of Kannisto (1994, 1996), Lundström (1995; Vaupel and Lundström, 1994), Thatcher (1992), and others, reliable data on mortality after age 80 are now available for Japan and 13 Western European countries. When these data are pooled, it is possible to accurately estimate the age-trajectory of human mortality up to about age 107 for females and age 105 for males. Reasonable estimates can be made up to about age 110, and shakier "guesstimates" can be boldly ventured up to age 120 (Thatcher et al., 1997). Figure 2-1 clearly shows that human mortality does not increase exponentially after age 80. Mortality decelerates, reaching perhaps a maximum or ceiling around age 110. Whether mortality is slowly increasing, level, slowly decreasing, or rapidly decreasing after age 110 is uncertain.
These results are based on data on some 70 million humans who reached age 80, some 200,000 who celebrated their 100th birthday, and I exceptional person, France's Jeanne Calment, who reached age 122 on February 21, 1997.
Humans are animals. Almost all animals show signs of aging; for almost all animals death rates tend to rise after the age of maturity (Finch, 1990). Even researchers who are only interested in people may benefit from biological insights from studies of other species, because these insights may cast light on the biology of humans. The following paragraphs describe some recent findings about age-trajectories of mortality for insects and worms.
The largest nonhuman population followed to natural death consisted of 1.2 million medflies studied in a laboratory near Tapachula, Mexico. These flies were held in cages, each holding several thousand flies. As reported by Carey et al. (1992), the trajectory of mortality rises, peaks, and then falls to a low level around which it hovers until the last fly died at an age of 171 days (compared with an average life span in the experiment of 21 days). The results up to day 100 are shown in Figure 2-2.
Weismann (1889), one of the forerunners of the evolutionary theory of aging, argued that "there is no reason to expect life to be prolonged beyond the reproductive period; so that the end of this period is usually more or less coincident with death" (quoted in Rose, 1991). As noted earlier, Charlesworth (1994) and others who helped develop the evolutionary theory of aging have made
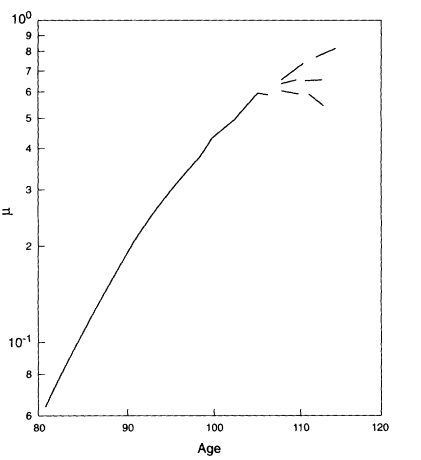
Figure 2-1
Mortality at ages 80 and over, females, 1980-1992. Pooled data are from 14 countries (Japan and 13 Western European countries) with the most reliable information. µ = force of mortality. SOURCES: Kannisto (1994, 1996); Lundström (1995); Vaupel and Lundström (1994); and Thatcher (1992).
similar assertions. In the wild there is little egg-laying among medflies after the first month of adult life, so a sharp rise in mortality might be expected around age 30 days. Many of the medflies, however, survived many months after the usual cessation of reproduction.
Following up on this pathbreaking initial study, the Carey laboratory completed life span analyses on an additional 1.6 million medflies, raised under a variety of conditions. Again, mortality decelerated at older ages. This result was replicated in a study of I million Anastrepha ludens, the Mexican fruit fly, which is somewhat larger and somewhat longer-lived than the medfly. For this species,
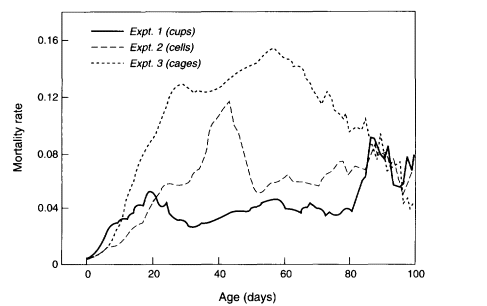
Figure 2-2
Smoothed age-specific death rates for three medfly mortality experiments. The total number of individuals remaining alive at age 100 days for experiments 1 (cups), 2 (cells), and 3 (cages) was 307, 31, and 62, respectively. SOURCE: Carey et al. (1992).
mortality peaked around 2 months of age and then roughly leveled off (Boe et al., 1995).
In James Curtsinger's laboratory, the life spans of tens of thousands of Drosophila melanogaster have been studied in various controlled experiments. These experiments all involve genetically identical strains of flies. From the earliest reported findings (Curtsinger et al., 1992) on, the experiments have shown a deceleration of mortality at older ages (e.g., Promislow et al., 1996), with strong indications that the mortality rate at older ages is "approximately constant and independent of age" (Curtsinger et al., 1992). Figure 2-3 shows a typical pattern of mortality for one of the isogenetic strains analyzed.
In Thomas Johnson's laboratory, a cohort of 180,000 nematode worms of the species Caenorhabditis elegans were followed until the entire population had died (Brooks et al., 1994). The worms were of the same genotype—that is, they were genetically identical, like identical twins. Analysis of these data revealed a definite deceleration in mortality around day 8 or 9, which is the age when reproduction for this genotype falls to very low levels (Vaupel et al., 1994). As shown in Figure 2-4, a two-stage Gompertz model fits the data fairly well. From day 5 to 8 mortality increased at a rate of 0.58, which is more than twice the rate of 0.21 that prevails afterwards. A smooth curve that levels off at the highest ages also fits the data well.
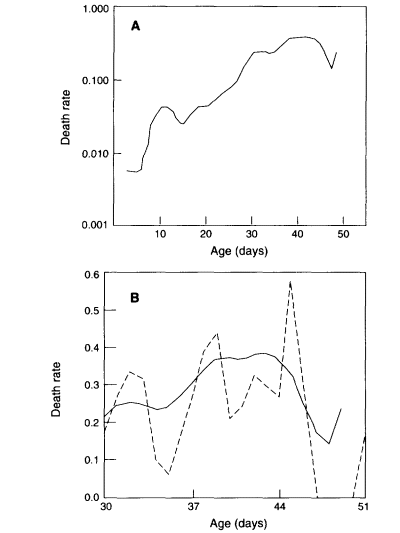
Figure 2-3
(A) Semilogarithmic plot of death rates for 5751 male Drosophila melanogaster from inbred line. (B) Detail of mortality rate (arithmetic scale) of last 1906 deaths. Dashed line: Observed daily probabilities of death (five final deaths not shown). Solid line: Death rates estimated by smoothing.
SOURCE: Curtsinger et al. (1992).
Pamela Larsen directs a nematode laboratory at the University of Southern California. She has compared the mortality trajectory of the so-called wild-type nematode, the genotype usually studied, with mutant strains that differ from the wild type by a point mutation at a single locus known as DAF2 (Larsen, 1993), As shown in Figure 2-5, her experiments show a sharp rise in mortality in the wild-type strain, but a much slower increase, with signs of a rough leveling off, in the DAF2 genotypes. What is remarkable is the radical change in the age-trajectory of mortality produced by a point mutation. [Larsen's results look quite different from Johnson's results, as shown in Figure 2-4, but they are not inconsistent. Larsen's experiments involved thousands, rather than hundreds of thousands, of worms. As demonstrated in Carey et al. (1992), the deceleration of mortality may not become apparent until a very large experiment is done. Furthermore, Johnson's experiments were conducted under much harsher conditions than Larsen's experiments, which explains the difference in life expectancy.]
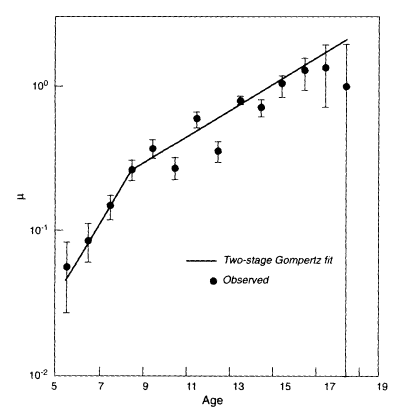
Figure 2-4
Nematode mortality. µ = force of mortality.
SOURCE: Vaupel, Johnson, and Lithgow (1994).
For all five species for which reliable data are available on large populations—humans, medflies, Mexican fruit flies, Drosophila, and nematode worms—mortality tends not to accelerate, but rather to decelerate, at older ages. This finding also appears true in some other species for which smaller populations were studied, including the bruchid beetle Callosobruchus maculatus (Tatar et al., 1993) and perhaps the flour beetle Tribolium confusum and the common house fly Musca domestica (Wilson, 1994). Death rates are such that many individuals in these various species survive well past the ages when reproduction usually ceases or drops to very low levels. Madame Jeanne Calment, who died at age 122, had lived seven decades after menopause. For the flies and worms, which do not care for their progeny, extraordinarily long spans of life have been observed in the ''postreproductive" period when fertility is very low. Nematodes, for instance, have very few progeny after the
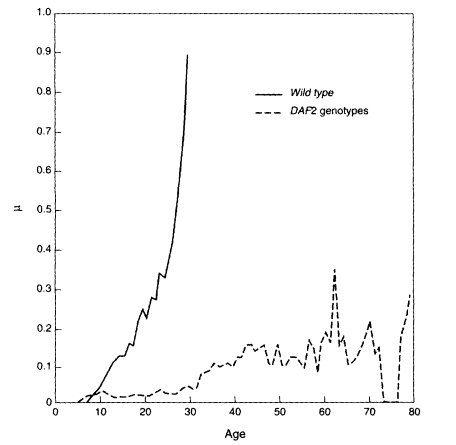
Figure 2-5
Nematode mortality. µ = force of mortality. SOURCE: Larsen (1993).
eighth day of their lives, but Pamela Larsen's longest-lived worm wriggled around for some 96 days.
Mechanisms
Mortality deceleration came as a surprise, indeed as a shock, to many biologists and gerontologists. (Demographers, on the other hand, had long suspected that death rates probably increased more slowly at older ages, although the data available until recently did not permit a definitive conclusion.) A spate of possible explanations were quickly developed. The following paragraphs review the main possibilities.
In most large animal experiments, populations are kept in confined areas of fixed size. As population size declines with age, population density decreases. If lower densities are salubrious, then the observed leveling off and decline in mortality with age might be an artifact of such changes in density (Graves and Mueller, 1993; Nusbaum et al., 1993; Kowald and Kirkwood, 1993). However, populations of medflies raised in solitary confinement in individual cells also showed a deceleration of mortality at older ages (Carey et al., 1992). Further medfly experiments were done in which density were systematically varied and changes in density were statistically controlled (Carey et al., 1995). The level of mortality increased at higher densities, but regardless of density, mortality rose, leveled off, and then fell with age. In addition, Drosophila experiments were painstakingly conducted in which density was held constant by replacement of older flies (Khaezeli et al., 1995). These experiments "show a deceleration of mortality at older ages that is not attributable to density effects" (Curtsinger, 1995a).
All populations are heterogeneous. In some of the experiments described above, strong efforts were made to ensure that all individuals in the population were genetically identical and that environmental conditions were highly similar for all individuals. Even in these experiments, however, individuals differed from each other, in size, in weight, and, more generally, in robustness and vitality. Some individuals are frailer than others . . . and the frail tend to die first. This creates a fundamental problem—indeed it seems to me the fundamental problem—for demographic analyses in general and for analyses of age-trajectories of mortality in particular. The individuals alive at older ages are systematically different from the individuals alive at younger ages. The age-trajectory of mortality reflects both the underlying age-trajectories of mortality for individuals in the population and the effects of compositional change as the frailer individuals drop out of the population.
Some of my colleagues and I have devoted a lot of thought to this problem over the past two decades (e.g., Vaupel et al., 1979; Vaupel and Yashin, 1985; Vaupel and Carey, 1993). Several researchers suggested that deceleration observed in medfly and Drosophila mortality might be attributable to heterogeneity
(Olshansky et al., 1993; Kowald and Kirkwood, 1993; Brooks et al., 1994), and this possibility was noted in both the original Carey et al. (1992) report on medfly mortality and the original Curtsinger et al. (1992) report on Drosophila mortality. It can be mathematically proven that compositional change (resulting from the higher mortality of the frail) decreases the rate of increase in the age-trajectory of mortality. In fact, even if death rates are increasing with age for every individual in a population, the age-trajectory of mortality can level off or even decline as a result of compositional change. The Gompertz exponential formula could hold for individual medflies, even though death rates for cohorts of medflies fall substantially at older ages.
Vaupel and Carey (1993) show this. They also show, however, that the required degree of heterogeneity is very large. To fit an underlying Gompertz model to the medfly data, they had to divide the population into subgroups that had vastly different chances of death. The frailest subgroup had 5 billion times the chance of death at any age as the most robust subgroup. Although there may be considerable heterogeneity in relative risks of death in a population, it seems unlikely that the degree of heterogeneity is this large. Natural selection will tend to eliminate subgroups with very high mortalities. Such subgroups could, in theory, be blessed with counterbalancing advantages: they could be extremely fertile or they could tend to produce a few very robust individuals. Further research is warranted, but at present it seems implausible that all of the observed deceleration of mortality at older ages is an artifact of heterogeneity.
If the deceleration of mortality is not completely attributable to compositional change, then there has to be some deceleration at the individual level. It seems plausible that some of the observed deceleration is due to behavioral and physiological changes that occur with age and that are associated either with declines in reproductive activity or with repair mechanisms that compensate for damage at younger ages (Finch, 1990; Carey et al., 1992; Kowald and Kirkwood, 1993; Tatar et al., 1993; Carey and Liedo. 1995). More research is required on the importance of these effects and on the extent to which the deceleration of mortality at older ages can be attributable to changes at the individual level.
More generally, research is needed on the mechanisms that produce various details of the pattern of mortality seen in large populations. For instance, what accounts for the prominent hump in medfly mortality around day 30 (as shown in Figure 2-2)? This hump not only appears in the initial population of 1.2 million but also appears in the large populations of several million medflies studied since (Carey and Liedo, 1995; Carey et al., 1995). Mueller et al., (1997) analyze this "mortality surge." Their research shows that careful attention to the details of the bends and bulges of mortality trajectories can lead to deeper understanding of the determinants of survival and longevity. Horiuchi and Wilmoth (e.g., in press) have done some interesting work along these lines for humans.
Bio-Reliability Theory
Living organisms are complex systems; reliability engineers and systems analysts have learned a great deal about the failure of complex systems. Even the simplest bacterium is vastly more complicated than the most elaborate machinery. Living organisms have repair and homeostatic programs that far excel those in any mechanical device. Nonetheless, it may prove useful to merge perspectives from reliability engineering and evolutionary theory, from systems analysis and biology, from the study of populations of equipment and the study of populations of people and other forms of life.
Reliability engineers often work with stochastic models in which various states represent the "health" of the system, as determined, for instance, by which components are still working and which components are broken. LeBras (1976) and Gavrilov and Gavrilova (1991) provide some examples that they interpret as models of aging. Yashin et al. (1994) show that the LeBras model and a related model presented by the Gavrilovs produce exactly the same age-trajectory of mortality as does the frailty model proposed by Vaupel et al. (1979). In particular, the various models all lead to a deceleration and eventual leveling off of mortality.
Statisticians interested in stochastic models of failure have considerably generalized this line of thinking. Consider a homogeneous Markov chain in continuous time with a single absorbing state, which can be interpreted as failure or death. Such a Markov chain can have many states and many branches from each state; it can include return to healthier states (as the result of recovery or repair). Suppose there is some initial distribution of individuals among the nonabsorbing states. Then time until death is said to follow a phase-type distribution. Under a variety of fairly general conditions, the age-trajectory of mortality for such a system will eventually level off. Under somewhat more restrictive but still fairly general conditions, the age-trajectory of mortality can rise, decelerate, and then level off. Other more restrictive conditions can produce other patterns, including mortality trajectories with bumps and declines such as those observed for medflies (Figure 2-2). Aalen (1995) suggests that such models should find greater application in biostatistics; he provides some examples as well as a lucid review of the basic ideas.
Many biologists seem to believe that it is possible for organisms to approximate the "one-hoss shay" of Oliver Wendell Holmes that ran perfectly until one day when all of its pieces fell apart simultaneously (e.g., Fries and Crapo, 1981; Dawkins, 1995). This is one of the suppositions used to justify the belief in species-specific maximum life spans. Reliability theory, however, strongly suggests that one-hoss shays are impossible to construct—unless there is a built-in time bomb. Empirical data on the failure of equipment is consistent with the theory. Equipment is often designed such that there is a high probability of survival up until some point. The Pioneer space probe, for instance, was de-
signed to reach Mars: consumer durables may be designed to last until the end of the warranty period. Such designs, however, generally result in substantial spans of life after the target age. The Pioneer space probe was still functioning when it escaped the Solar System; some washing machines and refrigerators continue to function years after their warranties have expired. A body design that gives an organism a good chance of surviving long enough to reproduce may be a sufficiently robust design that some of the organisms can survive long thereafter (Hayflick, 1994).
Automobiles are popular pieces of complicated equipment. They are sufficiently standardized that it is meaningfully possible to count their numbers on an age-specific (model-year) basis. Using automobile registration data gathered by the various states of the United States, R.L. Polk and Company did so for July 1, 1941, and then for July 1 of every year from 1947 to the present. These data, which are closely analogous to population count data collected by national statistical offices and census bureaus, have been assembled and analyzed by Vaupel and C.R. Owens (unpublished work). Using standard demographic methods, they estimated, among other summary statistics, the age-trajectory of mortality for automobiles in various cohorts and in various periods, as shown in Figures 2-6 and 2-7. Depending on the period or cohort, it was possible to compute central death rates—the number of deaths in an interval divided by the average number of individuals alive in the interval—from age 1 to an age up to age 17. Several aspects of the age-trajectories of automobile mortality are intriguing and are addressed by Vaupel and C.R. Owens.
What is important here is the deceleration and leveling off of mortality at older ages. If the titles and legends of Figures 2-6 and 2-7 were erased, an unsuspecting observer might think the trajectories pertained to Drosophila or nematode worms or some other living species. The trajectories are also somewhat similar to the human pattern at older ages.
The deceleration and leveling off of automobile mortality is partially due to heterogeneity—the composition of the automobile fleet changes with age because some kinds of cars are built better and last longer than others. As shown in Figure 2-8, however, even if a single make of car were chosen and a single model year were followed, there is still evidence of deceleration and leveling off. Among cars of the same make and model year—roughly analogous to genotype, although there are different models of the same make—there are certainly differences in "phenotype" (i.e., in observable characteristics of individuals), arising from various errors made in constructing particular cars, from the environment in which the car is driven, from the temperament of the car's owner, etc. Lemons driven on dirt roads by careless, impetuous drivers will tend to be selected out.
Beyond the effects of such compositional change, there are almost certainly "physiological and behavioral" changes that contribute to mortality deceleration. Older cars are repaired. Older cars may be driven less frequently and more
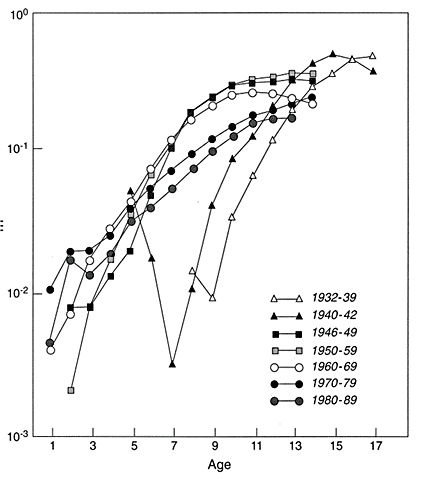
Figure 2-6
Central death rates for automobiles in various cohorts. m = central death rate.
carefully. As cars age, their owners may also age and become more stolid and responsible.
On a more general and speculative level, the question arises—is mortality a property of living organisms or a property of complicated systems? When it comes to death, how do people and worms differ from Chevrolets? In particular, is the deceleration and leveling off of mortality a fairly general property of complicated systems (as suggested by phase-type models and by the automobile data)? Better understanding of these questions may lead to new insights into aging and survival. Judicious caution is required, however, because living organ-
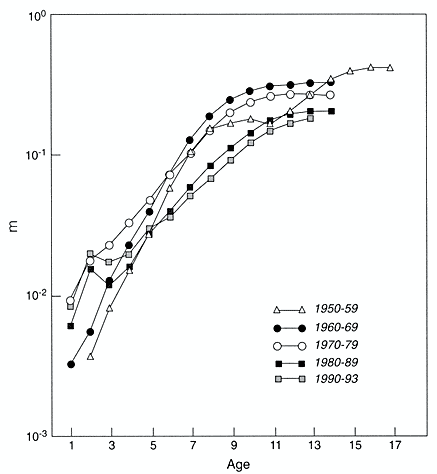
Figure 2-7
Central death rates for automobiles in various periods. m = central death rate.
isms are vastly more complicated than equipment and fundamentally different along many key dimensions.
In sum, although living organisms and equipment share only a few common attributes, demographers and biologists may nonetheless be able to learn something from reliability engineers. There is already a splendid example of this—the statistical methods used to analyze survival data were to a considerable extent developed by reliability engineers interested in analyzing failure-time data. Phase-type models, the long postmission life of space probes, and empirical data on the mortality of automobiles suggest the range of potential opportunities for cross-fertilization.
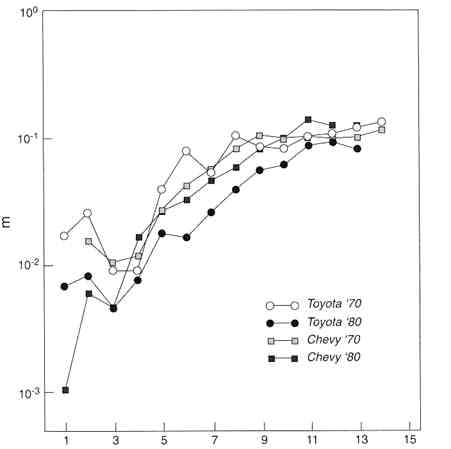
Figure 2-8
Central death rates for Chevrolets and Toyotas. 1970 and 1980 model years. m = central death rate.
The Evolutionary Theory Of Aging Revisited
The discussion so far has addressed a number of explanations for the observed deceleration of mortality in the five species for which large populations have been studied. The deceleration does not appear to be an artifact of declining population densities in the experimental populations. The deceleration is probably partially, but only partially, attributable to compositional change in heterogeneous populations. The deceleration is probably partially attributable to behavioral and physiological changes at the individual level. A deceleration of mortality is also observed in large populations of automobiles—and this decel-
eration is almost certainly due to a combination of compositional change and changing behavior. It seems clear that various genetic, environmental, behavioral, bio-reliability, and population-heterogeneity mechanisms interact to determine the age-trajectory of mortality in general and its deceleration in particular. The nature of these mechanisms and their relative importance remain to be explored. Specific, testable hypotheses have to be formulated, and appropriate data sets have to be gathered. Experiments with various animal models (and perhaps even with inanimate machines) will almost certainly provide valuable information to supplement observational studies of human populations.
Perhaps the biggest current challenge is to clarify the mystifying puzzle of why the evolutionary theory of aging does not appear consistent with the empirical observation that mortality decelerates at older ages. "Nothing in biology makes sense except in the light of evolution," Dobzhansky (1973) asserted. Evolutionary thinking, however, appears to lead to the prediction that deleterious mutations should accumulate at the age when reproduction falls to low levels. A black hole of bad alleles (i.e., of mutational forms of genes) should preclude survival much past this age. There appear to be at least two ways out of this quandary; neither is, as yet, very well understood, and there may be other solutions as well.
First, it may not be true that the elderly are unimportant from an evolutionary perspective. As Carey and Gruenfelder (in this volume) put it, the elderly may play important roles in nature. For some species this role may consist of helping offspring and other relatives survive. For other species, very rare instances of fertility at older ages may be crucially important and hence of much greater evolutionary significance than simple versions of Lotka's equation imply. In particular, it may be that some species have to make it through bottlenecks, due perhaps to lack of food, cold weather, or drought. Only a very few, exceptionally robust and long-lived individuals may make it through such bottlenecks. These individuals would be the founders of the new populations: all the members of these new populations would have the founder's longevity genes. Even if there are no bottlenecks, there may be considerable stochasticity in a species' environment such that late reproduction is sometimes favored. Tuljapurkar (in this volume) discusses the importance of such stochasticity.
Second, it may be that "mutation-accumulation" alleles are rare. These alleles have bad effects but only at older ages: at younger ages, they are neutral and have no effects. Such alleles may be rare because it is difficult in practice for mutations to occur that produce effects that first start operating at older ages. Nature is constrained by the materials and processes at hand. It is also possible that such alleles are rare because they are knocked-out by subsequent mutations. Most mutations are harmful; many are so harmful that they prevent a gene from properly functioning. Such knock-out mutations can be expected to eventually destroy any allele that does not contribute in some way to an organism's evolutionary fitness. If mutation-accumulation alleles are rare, they may not be of
much significance, especially for biological processes that involve many genes in a redundant way that reduces the harmful effects of any particular allele.
"Antagonistic-pleiotropy" alleles (i.e., genes that have multiple effects that pull in different directions) may also be rare. In particular, such alleles are supposed to come in two varieties: some are helpful at younger ages and harmful at middle and older ages, whereas others are harmful at younger ages and helpful at middle and older ages. The older ages in this model do not count for much, if at all, because few individuals survive to older ages, and those that do have low fertility. A tradeoff of good and bad effects between younger and middle ages is, however, theoretically possible. Mutations may arise that lead to the existence of two alleles at the same locus, with opposite age-specific impacts. All species have to make tradeoffs between fertility and survival, between early and late fertility, between early and late survival, etc. Mutations that produce pairs of antagonistic-pleiotropy alleles may be the mechanism that enables alternative choices about such tradeoffs. Both of the two competing alleles may, however, not survive many generations: one may be driven to extinction as the other goes to fixation (Curtsinger et al., 1994). If this were the case, one would observe that species make tradeoffs, but one would only occasionally observe a genetic locus with two or more antagonistic-pleiotropy alleles with different age-specific effects.
The research of Carey, Partridge, Tuljapurkar and Rose (all of whom have chapters in this volume), as well as Curtsinger (1995a,b), Abrams (1993), and others, is helping clarify the mysterious fact that many individuals survive well past the normal end of reproduction. In addition to the ideas adumbrated above, other possible explanations may prove important. At present, this question appears to be a wide-open, exciting issue on the frontier of knowledge.
Conclusion
This chapter was designed to serve two purposes. My charge was to address the question of what demographers, ecologists, and evolutionary biologists can learn from each other. Actually, demographers, ecologists, and evolutionary biologists have learned a lot from each other about the trajectories of mortality at advanced ages. In explaining what they have learned, the potential for this kind of biodemographic collaboration can be conveyed in a concrete, persuasive way. What has been learned—and what could be learned through further collaboration—is fascinating: outlining this was the dual, complementary purpose of this chapter, the other side of the same coin.
Demographers, ecologists, and evolutionary biologists focus on different topics, have memorized different facts and jargon, and have been indoctrinated with different concepts. They share, however, an interest in populations. Although many demographers rigidly restrict demography to the study of human populations, the methods and concepts of demography are useful in studying
many kinds of populations, from insects and other forms of life (Carey, 1993) to Chevrolets. If nothing in biology can be understood except in the light of evolution, an equally valid generalization is that nothing in evolution can be understood except in the light of demography. Lotka's demographic equation, which relates survival and fertility to population growth, is the fundamental equation in most evolutionary theorizing in general and in the evolutionary biology of aging in particular.
As Rose (1991) stated, ''From these basic features of the demography of populations (i.e., the age-trajectories of survival and fecundity), the crucial evolutionary determinants of aging follow." The research of demographers, ecologists, and evolutionary biologists reviewed in this chapter shows it is still not fully understood how the crucial evolutionary determinants of aging follow from the underlying demography of survival and fertility. Nonetheless, the point is well taken. Deeper understanding of the demography of populations—through research on genetic, environmental. behavioral, bio-reliability. and heterogeneity forces and constraints, including emphasis on the role of the elderly in nature (Carey and Gruenfelder, in this volume)—will almost certainly provide new insights and perhaps the crucial insights into the mechanisms that drive the processes of aging and survival.
References
Aalen. O.O. 1995 Phase type distributions in survival analysis. Scandinavian Journal of Statistics 22:447-463.
Abrams. P. 1993 Does increased mortality favor the evolution of more rapid senescence? Evolution 47:877-888.
Boe. C.. C. Gruenfelder. P. Liedo. and J.R. Carey 1995 Mortality Deceleration and Longevity in the Mexican Fruit Fly (Anastrepha ludens). Manuscript. Department of Entomology, University of California, Davis, CA 95616.
Brooks, A. G.J. Lithgow. and T.E. Johnson 1994 Mortality-rates in a genetically heterogeneous population of Caenorhabditis elegans. Science 263[5147]:668-671.
Carey, J.R 1993 Applied Demography for Biologists. New York: Oxford University Press.
Carey, J.R., and P. Liedo 1995 Sex mortality differentials and selective survival in large medfly cohorts. The Gerontologist 35:588-596.
Carey, J.R.. P. Liedo, D. Oro/co. and J.W. Vaupel 1992 Slowing of mortality rates at older ages in large medfly cohorts. Science 258:457-461.
Carey. J.R., P. Liedo, and J.W. Vaupel 1995 Mortality dynamics of density in the Mediterranean fruit fly. Experimental Gerontology 30:605-629.
Charlesworth, R. 1994 Evolution in Age-Structured Populations, 2nd ed. New York: Cambridge University Press.
Charlesworth, R., and K.A. Hughes 1996 Age-specific inbreeding depression and components of genetic variance in relation to the evolution of senescence. Proceedings of the National Academy of Sciences U.S.A. 93(12):6140-6145.
Curtsinger, J.W. 1995a Density and age-specific mortality [comment]. Genetica 96(3):179-182.
1995b Density, mortality, and the narrow view. Genetica 96(3):187-189.
Curtsinger, J.W.. H.H. Fukui, D.R. Townsend, and J.W. Vaupel 1992 Demography of genotypes: Failure of the limited life span paradigm in Drosophila melanogaster. Science 258:461-463.
Curtsinger, J.W., P. Service, and T. Prout 1994 Antagonistic pleiotropy, reversal of dominance, and genetic polymorphism. American Naturalist 144:210-228.
Dawkins, R. 1995 God's utility function. Scientific American 273:80-85.
Dobzhansky, T. 1973 Nothing in biology makes sense except in the light of evolution. American Biology Teacher 35:125-129.
Finch, C.E. 1990 Longevity. Senescence, and the Genome. Chicago: University of Chicago Press.
Fries, J.F. 1980 Aging, natural death, and the compression of morbidity. New England Journal of Medicine 303:130-135.
Fries, J.F., and L.M. Crapo 1981 Vitality and Aging: Implications of the Rectangular Curve. San Francisco. CA: W.H. Freeman.
Gavrilow, L.A., and N.S. Gavrilova 1991 The Biology of Life Span. Chur, Switzerland: Harwood Academic Publishers.
Gompertz, B. 1825 On the nature of the function expressive of the law of human mortality. Philosophical Transactions 27:510-519.
Graves, J.L., and L.D. Mueller 1993 Population density effects on longevity. Genetica 91:99-109.
Hamilton, W.D. 1966 The moulding of senescence by natural selection. Journal of Theoretical Biology 12:1245.
Hayflick, L. 1991 Aging under glass. Mutation Research 256(2-6):69-80.
1994 How and Why We Age. New York: Balantine Books.
Horiuchi, S., and J.R. Wilmoth In press Deceleration in the age pattern of mortality at older ages. Demography.
Hughes, K.A., and B. Charlesworth 1994 A genetic analysis of senescence in Drosophila. Nature 367:64-66.
Kannisto, V. 1994 Development of Oldest-Old Mortality. 1950-1990. Odense. Denmark: Odense University Press.
1996 The Advancing Frontier of Survival. Odense. Denmark: Odense University Press.
Khaezeli, A.A., L. Xiu, and J.W. Curtsinger 1995 Effect of adult cohort density on age-specific mortality in Drosophila melanogaster. Journal of Gerontology 50(5):B262-269.
Kowald. A., and T.B.L. Kirkwood 1993 Explaining fruit fly longevity. Science 260:1664-1665.
Larsen. P.L. 1993 Aging and resistance to oxidative damage in Caenorhabditis elegans. Proceedings of the National Academy of Sciences U.S.A. 90(19):8905-8909.
LeBras. H. 1976 Lois de mortalite a age limite. Population 31:655-692.
Lundström, H. 1995 Record longevity in Swedish cohorts born since 1700. In B. Jeune and J.W. Vaupel, eds., Exceptional Longevity: From Prehistory to the Present. Odense. Denmark: Odense University Press.
Medawar, P.B. 1952 An Unsolved Problem in Biology. London: H.K. Lewis.
Medvedev, Z.A. 1990 An attempt at a rational classification of theories of ageing. Biological Reviews: 65:375398.
Mueller, H.G., J.L. Wang, W.B. Capra, P. Liedo, and J.R. Carey 1997 Early mortality surge in protein-deprived females causes reversal of sex differential of life expectancy in Mediterranean fruit flies. Proceedings of the National Academy of Sciences U.S.A. 94(6):2762-2765.
Nusbaum, T.J., J.L. Graves. L.D. Mueller. and M.R. Rose 1993 Fruit fly aging and mortality. Letter to Editor. Science 260:1567.
Olshansky, S.J.. B.A. Cames, and C.K. Cassel 1993 Fruit fly aging and mortality. Letter to Editor. Science 260:1565-1566.
Promislow, D.E.L., M. Tatar, A.A. Khazaeli, and J.W. Curtsinger 1996 Age-specific patterns of genetic variance in Drosophila melanogaster. Genetics 143(2):839-848.
Rose, M.R. 1991 Evolutionary Biology of Aging. New York: Oxford University Press.
Tatar, M.. J.R. Carey, and J.W. Vaupel 1993 Long-term cost of reproduction with and without accelerated senescence in Callosobruchus maculatus: Analysis of age-specific mortality. Evolution 47:1302-1312.
Thatcher, A.R. 1992 Trends in numbers and mortality at high ages in England and Wales. Population Studies 46:411-426.
Thatcher, A.R., V. Kannisto, and J.W. Vaupel 1997 The Force of Mortality from Age 80 to 120. Odense, Denmark: Odense University Press.
Vaupel. J.W., and J. R. Carey 1993 Compositional explanations of medfly mortality. Science 260:1666-1667.
Vaupel, J.W., T.E. Johnson, and G.J. Lithgow 1994 Rates of mortality in populations of Caenorhabditis elegans. Science 266:826.
Vaupel. J.W., and H. Lundström 1994 The future of mortality at older ages in developed countries. In W. Lutz, ed., The Future Population of the World. London: Earthscan.
Vaupel. J.W., K.G. Manton, and E. Stallard 1979 The impact of heterogeneity in individual frailty on the dynamics of mortality. Demography 16:439-454.
Vaupel, J.W.. and A. I. Yashin 1985 Heterogeneity's ruses: Some surprising effects of selection on population dynamics. American Statistician 39:176-185.
Weismann, A. 1889 Essays upon Heredity and Kindred Biological Problems. Oxford: Clarendon Press.
Williams. G.C. 1957 Pleiotropy, natural selection, and the evolution of senescence. Evolution 11:398-411.
Wilson, D.L. 1994 The analysis of survival (mortality) data: Fitting Gompertz. Weibull. and logistic functions. Mechanisms of Ageing and Development 74:15-33.
Yashin, A.I., J.W. Vaupel, and I.A. Iachine 1994 A duality in aging: The equivalence of mortality models based on radically different concepts. Mechanisms of Aging and Development 74:1-14.





















