
5 Evolutionary Biology and Age-Related Mortality
Linda Partridge
Introduction
Analysis and interpretation of age-specific death rates are of considerable interest to both biodemographers and evolutionary biologists. Of special concern to both groups is the age-related decline in survival probability later in life, characteristic of many organisms including humans. This decline is generally taken to be indicative of a decline in intrinsic state, known as aging or senescence. The exact form of the survival curve differs considerably among human populations living in different areas and at different times (Finch, 1990; Wilmoth, in this volume), and we want to understand these differences and to produce a predictive theory of how mortality rates would change with age in different environments and behave in populations of different composition. Such a theory would be especially useful if it could also help predict patterns of age-related morbidity and cause of death.
Age-specific death rates have three main determinants: age-related changes within individuals, the environment in which they are placed, and qualitative differences between individuals. In addition, these three factors can interact with one another in the determination of mortality. Evolutionary biology has a strong theory for the evolution of age-related changes within individuals (e.g., Cole. 1954; Williams, 1957, 1966; Schaffer, 1974; Pianka and Parker, 1975; Charlesworth, 1980 and 1994; Roff, 1992; Stearns, 1992). Theories for the impact of the environment (Gomulkiewicz and Kirkpatrick, 1992; Scheiner, 1993; de Jong, 1995) and for the effects of individual differences (e.g., McNamara and Houston, 1996) are being increasingly explored. Human populations often inhabit an environment very different from those in which their life history evolved. This
considerably weakens the predictive power of evolutionary life-history theory, because interactions between genotypes and environments for life-history traits are pervasive (e.g., Stearns, 1992; Chapman and Partridge, 1996). In addition, much demographic analysis of the impact of aging has relied on various measures of age-specific death rates late in life. The evolutionary theories of aging apply to fertility and survival rates jointly, and from an evolutionary perspective aging is complete when reproduction ceases, unless the old contribute to reproduction by relatives. Deriving any general predictions for death rates alone, especially for postreproductive death rates, is not simple.
The following sections briefly summarize these issues and point to some areas where further work could be profitable. In addition, the broader implications of evolutionary ideas for the aging process are discussed.
Age-Related Changes Within Individuals
Evolution of Life Histories and of Aging
The intrinsic state of individuals changes with age in humans and other organisms. These changes in state are reflected in changes in vital rates: survival and fertility. Evolutionary life-history theory has been directed at understanding the diverse array of age-related changes seen in different organisms.
From an evolutionary perspective, a life history is the combination of age-specific survival probabilities and fertilities characteristic of a type of organism. These produce the life table of survival probabilities to each age x (lx) and the fertilities at those ages (mx). The life history includes traits such as age at first breeding, number of breeding attempts, fecundity at each breeding attempt, and adult life span. The life histories observed in different species and, indeed, populations are extremely diverse, and the forces affecting their evolution are becoming well understood (e.g., Cole, 1954; Schaffer. 1974; Pianka and Parker, 1975; Charlesworth, 1980 and 1994; Stearns, 1992; Roff, 1992; McNamara and Houston, 1996; Tuljapurkar, in this volume). Natural selection is expected to act on the life history so as to incorporate into the population alleles that maximize the intrinsic rate of increase (r), which takes into account changes in the size of the population into which offspring are born or, in a population where numbers do not vary, lifetime reproductive success (R0). To reach this conclusion requires certain simplifying assumptions such as stable age structure, absence of frequency-dependent selection, and absence of any interaction between the effects of male and female genotype upon the fertility of mating pairs. Other more complex fitness criteria can be used if some of these assumptions are violated (Tuljapurkar, in this volume).
Maximization takes place in the face of two forces. The first is the set of constraints on the combinations of different life-history variables that can be realized. Some of these are internal to the organism, in the form of various
physiological constraints such as costs of growth, repair, and reproduction. Others are ecological, in the form of an altered impact of factors such as predation or parasitism in response to an alteration in, for instance, reproductive rate. Life histories evolve by a process of constrained optimization, in which the organism makes tradeoffs between costly competing activities. The environment that the organism inhabits will also affect the evolution of the life history, by imposing characteristic rates of age-specific (or size- or stage-specific) mortality and insults to fertility. These will affect the prospects for future reproduction at each age. In an environment with relatively high, externally imposed adult mortality, for instance, effort expended on reproduction is expected to increase even if survival probability and future fertility are thereby reduced by physiological costs, because prospects for future reproduction are anyway poor for environmental reasons.
The life histories of many organisms include a period of aging or senescence, terms for the deterioration in performance observed later in the adult life span. From an evolutionary point of view, the important manifestations of aging are a drop in survival probability and in fertility. These senescent changes have been very frequently reported, both in nature and in benign environments (Comfort, 1979; Finch, 1990; Rose, 1991; Charlesworth, 1980 and 1994). Aging is a particularly interesting trait from an evolutionary point of view, because taken in isolation it is maladaptive relative to its absence. We have to account for the failure of organisms later in life to achieve the levels of performance of which they were capable in youth. Aging is caused by damage to molecules, cells, organs, and systems. It could therefore be regarded as simply inevitable (Kirkwood, 1977; Kirkwood and Rose, 1991). However, different organisms have very different rates of aging, suggesting that the process is combated to different extents. It is this diversity in resistance to the effects of damage that requires an evolutionary explanation.
The evolution of aging is embedded in the evolution of the life history as a whole. The intensity of natural selection declines over the adult part of the life history because, even in a potentially immortal or indefinitely fecund organism, extrinsic forces such as disease, predation, and accidents impose mortality and damage to fertility. A mutant that is expressed early in adulthood will therefore be subject to strong selection because most of its initial bearers will still be alive and fully fertile. For a mutant expressed later, many initial carriers will already have died or suffered external insults to their fertility at a rate no different from nonbearers of the mutant, so that only the survivors are exposed to selection (Medawar, 1952). For age-specific fertility, selection intensity depends upon the probability of reaching that age, whereas for survival, selection intensity depends both upon survival probability to each age and what is lost by death at different ages, and therefore upon the future prospects for both survival and fertility (Charlesworth, 1980 and 1994).
Aging can therefore evolve as part of the constrained optimization of the life
history, because an allele with beneficial effects in the early part of life but with deleterious effects later on will be more strongly selected through the former than through the latter effect (Williams, 1957). Aging can therefore be accounted for as a deleterious side-effect of the evolution of an optimal life history (Partridge and Barton, 1993a). This theory has become known as the pleiotropy theory of aging (Vaupel, Tuljapurkar, in this volume), although the evolutionary processes at work need not include pleiotropy (Partridge and Barton, 1993a; 1996). Although life-history optimization may result in the evolution of aging, it does not follow that a monotonic decline in the intrinsic state of the organism will occur during the adult period. Under some circumstances, a condition is predicted to improve for part of the adult period, for instance as a result of growth or learning, which could lead to gains in future reproductive prospects through improved survival probability and fertility (Hamilton, 1966; Charlesworth, 1980 and 1994).
Aging is not an inevitable feature of life-history optimization and, for it to appear, the age of expression of the beneficial effects of the mutant must precede the age of expression of the deleterious effects. The alternative is an increase in risk (Partridge and Andrews, 1985; Partridge, 1987; Partridge and Barton, 1993a,b). For instance, a mutant individual that reproduced more at a particular age could be more likely to die at that age because it attracted the attentions of predators or became more likely to have a heart attack. When the organism ceased the risky activity, its phenotype would then be no different from that of an otherwise comparable organism that had not taken the risk. Risk contributes to the constraints under which the life history is optimized, but it does not, by definition, in itself induce aging, because it does not alter the phenotype, contrary to a recent claim (Blarer et al., 1995). An example of a beneficial effect with a delayed cost could be a negative effect of increased reproductive rate on various somatic repair processes, which could, in turn, reduce subsequent survival or fecundity (Kirkwood and Rose, 1991; Abrams and Ludwig, 1995), sometimes known as the "disposable soma theory" (Kirkwood, 1977).
Aging can also evolve as a result of depression of the life history below the optimal, through mutation pressure, the "mutation-accumulation theory" of aging (Vaupel, Tuljapurkar, in this volume). Deleterious mutations with effects on the phenotype that become apparent early in life are more strongly selected against than those expressed later in adult life (Medawar, 1952; Hamilton, 1966; Charlesworth, 1980 and 1994). Later-acting mutations are more likely to escape the attention of natural selection because their bearers are more likely to have died or become subfertile from some other cause, at a rate no different from that of nonbearers of the mutation, before the age at which the phenotypic effect of the mutation becomes apparent. Deleterious mutations with a later effect on the phenotype can therefore reach a higher frequency under mutation-selection balance, and their effects would be apparent as an intrinsic decline later in life.
For the mutation-accumulation theory to be important, new mutations should have deleterious effects on the phenotype that are at least partially specific to
later ages. This does not mean that the genes should be expressed at these ages. For instance, genes active during development could affect the durability of the soma, while genes affecting levels of activity and reproduction could determine the levels of damage that are incurred. The mutation-accumulation theory does not, therefore, rest on the implausible assumption that the late-life decline in state relies on gene expression of an increasing number of deleterious mutations. Rather, effects of variation between alleles and their patterns of expression at all ages become apparent in the phenotypes of old age.
How Can the Rate of Aging Be Measured?
What do these evolutionary theories allow us to predict about age-related changes in death rate in the later part of life in any population, including human populations? This question raises an important empirical issue, which is how the intrinsic deterioration that causes death rate to increase and fertility to decline at later ages can be detected and compared between different populations. There is considerable current interest in measuring the rate of aging in different natural populations, and in using such measures to deduce the evolutionary mechanisms at work (e.g., Nesse, 1988; Abrams, 1991; Promislow, 1991; Gaillard et al., 1994; Carey et al., 1992; Curtsinger et al., 1992; Abrams and Ludwig, 1995), which has led to lively debate about exactly how aging can be detected and its rate measured.
Most measures of the aging rate used thus far have focused on death rates. If intrinsic deterioration is occurring and death rate alone is used as a measure, then death rate clearly must change with age. Two populations that differ from each another in death rates that remain constant with age within each population do not, by this criterion, differ in their rate of aging (Medawar, 1952; Tatar et al., 1993; Blarer et al., 1995). Various measures of the rate at which death rate increases with age have therefore been proposed (Finch, 1990; Finch et al., 1990; Promislow, 1991; Carey et al., 1992) and have been used, for instance, to examine the occurrence of aging in natural populations (Promislow, 1991; Gaillard et al., 1994), to document the rate of change in death rate with age in humans (Finch et al., 1990), or to show an apparent slowing down in the rate of aging at later ages in insects (Carey et al., 1992; Curtsinger et al., 1992), nematodes (Brooks et al., 1994), and in humans (Vaupel, in this volume). An increase in age-specific death rates in theoretical models has been used to claim that spurious evidence for aging can be found in the absence of any intrinsic decline in the organism (Blarer et al., 1995) or that aging can occur as a direct result of an intrinsic improvement in state (McNamara and Houston, 1996). Superficially, some of these findings might be at variance with evolutionary theories of aging. However, they rest, in part, on the use of an inappropriate measure of aging from the evolutionary point of view, based solely on the way that death rates change with age.
A point that has escaped sufficient emphasis is the inevitability of the joint evolution of death and fecundity rates, rather than the separate evolution of the two traits (Partridge and Barton, 1993b, 1996). Both traits are expected to be affected by aging, and in ways that are not independent. One reason is that aging as a by-product of an optimal life history can occur because of a tradeoff necessitated by the direct physiological connection between survival and fecundity. Another is that the intensity of selection on age-specific fecundity depends upon the survival rate to that age, whereas selection upon age-specific survival depends on the fecundity schedule, because this affects what is lost by death at different ages (Charlesworth, 1980 and 1994). The important issues are to determine (1) how we can combine measures of survival and fertility into a single measure of the rate of aging, and (2) how use of this measure, instead of measures of rates of change in age-specific death rates, would affect conclusions about the pattern of change of intrinsic state in relation to age, and their concordance with evolutionary theories of aging.
Aging, the process we are trying to measure, occurs either because of later-expressed costs of processes beneficial earlier in life or because mutation pressure is depressing the life history below the optimal one. Either of these explanations will lead to an intrinsic decline in state, so that aging alone implies a drop in future prospects for offspring production. How can we detect this decline when it can occur against the backdrop of a state that can, over at least some of the adult life span, increase as part of life-history optimization? During this period of increase, aging may nevertheless be present, because intensity of selection starts to decline at the age of first reproduction (Charlesworth, 1980 and 1994), and its presence will mean that state will not improve as much as it would have in the absence of aging. However, in theory, during this part of such a life history, death rates could increase, decrease, or remain constant, despite the fact that intrinsic state is increasing (Hamilton, 1966; McNamara and Houston, 1996). Furthermore, intrinsic state could not itself be used to indicate the extent of the impact of aging. Only when intrinsic state starts to decline, as it must later on in any life history (Hamilton, 1966), can we start to measure and compare rates of intrinsic deterioration.
As a start in this direction we (Partridge and Barton, 1993b) suggested that the product lxmx, where lx is survival from birth to age x and mx is fecundity at age x could be used as a measure of the extent to which aging has occurred by age x. This product measures the contribution of each age to the total progeny output. However, this measure is taken from birth, and it applies only to age x. No indication therefore is given of the prospects for future reproduction from age x onward, given that age x is reached, which is the real test of intrinsic state at age x. A better measure would be Fisher's ''reproductive value":
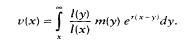
Here, r is the asymptotic rate of population growth, which in many cases could be taken as zero; note that v(0) = 1. The "reproductive value" gives the expected number of descendants that will be produced by an individual of age x over the rest of its lifetime, given that it has already survived to that age. It is therefore a measure taken prospectively from each age of the potential of the organism to produce further offspring, which is the evolutionarily relevant indication of the state of the organism (Partridge and Barton, 1996). Note that, as Hamilton (1966) argued, v(x) does not give the strength of selection on the life history at age x, since that should also account for the probability of surviving to age x. We also ignore complications from the effects of an individual upon the fitness of its relatives. These effects can be very important in many organisms, including humans (Carey and Gruenfelder; Austad, in this volume), and would require modification of the proposed measure of the rate of aging to account for inclusive fitness effects. The outcome should, nevertheless, be qualitatively similar.
The presence of aging in a population can consequently be detected as a late-life decline in reproductive value, and this is what is predicted by evolutionary theories of aging. The measure can also compare the rate of aging in different populations. Reproductive value is a continuous function of age, which means that any two life histories can differ for it in an infinite number of ways. If the reproductive value is always lower in one life history than another, then one can say that life history involves more rapid aging. If, in contrast, they cross over at one or more points, then the implications become more complex, and the rate of aging can be compared only during late-life decline.
Interpretations of a slowing down in the rate of increase in death rates at later ages (e.g., Carey et al., 1992; Curtsinger et al., 1992; Brooks et al., 1994; Vaupel, in this volume) should be viewed in this light. Because all cases so far reported involve postreproductive individuals and most of them are from species where individuals do not contribute to the survival of relatives, the observed effects are selectively neutral and are, therefore, not at variance with evolutionary theories of aging. Postreproductive survival in the absence of inclusive fitness effects is an epiphenomenon from the evolutionary point of view, and changes in mortality rates at these ages could have a variety of explanations (Charlesworth and Partridge, 1997). One could be the inevitable viability of complex machines after the time for which they are engineered (in this case by natural selection) to guarantee to function (Vaupel, in this volume). Postreproductive survival may be increased in unnaturally benign environments, and cessation of reproduction or an associated reduction in activity levels could both contribute to lowering of the impact of damage. Additional factors could be variation in frailty (Vaupel, in this volume), or a tendency for the elderly to inhabit more benign environments. The important point is that evolutionary theories of aging do not necessarily predict Gompertzian-type increases in postreproductive mortality rates. Nor does the mutation-accumulation theory necessarily predict catastrophic increases in mortality when reproduction ceases, especially as mutations that produce aging also
have some small deleterious effect earlier in life (Partridge and Barton, 1993a). The evolutionary theories do predict a monotonic decline to zero in reproductive value at some point in the life history. Re-examination of the reported anomalous cases in which "aging" (measured as mortality rate) appeared in theoretical models where there was either no change (Blarer et al., 1995) or a positive change (McNamara and Houston, 1996) in intrinsic state also shows that use of reproductive value as a measure of aging would have removed the anomaly.
To advance this issue will require the production of explicit theoretical models of the mutation-accumulation and pleiotropy theories that examine outcomes for age-related patterns of mortality in not only the reproductive but also the postreproductive period. Two recent studies (Abrams and Ludwig, 1995; Rose, in this volume) are promising starts in this direction. We also need more data. There is at present a paucity of information about the degree and pattern of age-specificity of fitness effects of new mutations, and of the relevant mutation rates. The timing of the effects of costs of reproduction or of investment in repair processes on future fertility and survival is also poorly known (Partridge and Sibly, 1991). These data are not easy to collect and are a challenge for future work. To understand how postreproductive survival evolves, we need to understand its genetic correlations with survival and mortality patterns during the reproductive period.
Can We Predict Either the Rate of Aging or the Rate of Change in Death Rate?
What can evolutionary theories of aging predict about its rate in different populations? One of the broadest predictions is that there will be a relationship between the impact of external factors on the survival and fertility of the population, on one hand, and the intrinsic rate of aging that it is expected to evolve, on the other (Medawar, 1952; Williams, 1957; Hamilton, 1966; Charlesworth, 1980 and 1994; Tuljapurkar, Rose, in this volume). For instance, higher externally imposed death rates of adults are expected to cause evolution of higher rates of aging for both survival and fertility. However, the precise nature of this prediction and the nature of the measurements necessary to test it have been the subject of some discussion (Abrams, 1993; Promislow, 1991; Blarer et al., 1995).
Population dynamics are one important consideration. If one vital rate is changed by a change in, for instance, the size-specificity of predators, then for the population to remain regulated in numbers, it must compensate in some way. For an alteration in externally imposed factors to impose a permanent change in selection on the life history, the population must not respond by restoring a pattern of age-specific vital rates that leaves the pattern of selection unaltered (Abrams, 1993). The first thing that must be established, therefore, is that a difference in the impact of external factors results in an altered pattern of age-specific vital rates. This external impact must be separated from any intrinsic
changes within the individuals in the study populations. Such studies are extremely difficult in nature. Allocating even differences in natural death rates, in unmanipulated populations, to intrinsic or extrinsic causes is, in general, not possible: are predators catching more great tits in one population because of lack of alternative prey, or because the great tits are distracted by the need to forage for larger broods?
One approach to this problem has been to use differences in life style as an indicator of hazard. For instance, Williams (1957) suggested that the evolution of flight might have resulted in a reduced impact of external hazard. A similar suggestion has been made for bats (Austad and Fischer, 1991). However, information of this kind does not involve any objective measurement of external hazard and does not form the basis for any quantitative statements. The contention about the hazard-reducing effects of flight may well be correct, but there is no direct evidence that it is, and alternative theories for avian longevity have been advanced (Carey and Gruenfelder, in this volume). Another approach has been to use mortality rate in early adulthood as a measure of hazard (Promislow, 1991). However, this mortality rate is affected by intrinsic as well as extrinsic factors. There is therefore a challenge to make the relevant measurements of hazard on natural populations.
The second requirement for a test of the relationship between the impact of environmental hazard and the evolved rate of aging is a measurement of the intrinsic change of state in natural populations and, in particular, its rate of decline in the later part of life. Here, it has been argued that life span under optimal conditions is a suitable measure (e.g., Williams, 1957; Austad and Fisher, 1991; Partridge and Barton, 1993a). There are two objections to this measure. The first comes from the nature of the environments in which such measures are generally made. What is required is an environment with external hazards removed but otherwise similar to the natural one. Typically, measurements of intrinsic change have been made in benign environments, very different from that in which the life history evolved, a problem because life-history traits and the correlations between them are often extremely sensitive to environmental change (e.g., Stearns, 1992; Chapman and Partridge, 1996). The study organism may, through phenotypic plasticity, alter its own intrinsic life history as a proximate response to, for example, changes in nutrition or even the removal of external hazard. The second problem comes from using life span as an indication of the aging rate. Measurement of rate of change of reproductive potential with age is the appropriate measure. Conclusions might be unaltered by its use, but this remains to be seen. Again, these measurements present a challenge for future studies. Although the data from nature are at present equivocal, the results of experimental, laboratory studies support the contention that external hazard is an important determinant of the evolution of both longevity and age-specific fecundity (Rose, 1984; Luckinbill et al., 1984; Partridge and Fowler, 1992; Zwaan et al., 1995).
Predicting the rate of aging for humans is more difficult than for many other organisms because their life history evolved in environments different from those inhabited by most present-day populations. We know rather little about the impact of external hazards on human populations in their evolutionary past, although archaeological, historical, and comparative anthropological data are increasingly being exploited for information on vital rates (Kaplan, in this volume). Disentangling the roles of extrinsic and intrinsic factors in producing these vital rates presents an interesting challenge. Analysis of natural selection on the life histories of humans is complicated by the fact that they can contribute to the care of descendants and other relatives when they themselves have become unable to reproduce (Carey and Gruenfelder, Austad, in this volume). Patterns of personal fecundity do not therefore give an adequate picture of true fitness schedules in the evolutionary past. In any discussion of the implications of evolutionary knowledge for age-related patterns of human death rates, we therefore need to bear in mind that part of the picture is missing, and that this could alter our conclusions. However, the physical frailty of the oldest old may have greatly limited their contribution to the reproductive success of younger kin, so the difficulty may not be too serious.
Environmental Effects
The environment is an important determinant of the form of life histories. We have already seen that the pattern of hazard that it imposes is expected to determine the way in which natural selection acts on the life history. In addition, environmental change can affect the ways in which life-history traits are expressed and the correlations between them. For instance, Drosophila melanogaster reared at lower temperatures develop more slowly and into a larger adult. Environmental effects are not necessarily predictable from evolutionary ones. For instance, D. melanogaster that have evolved at lower temperatures, when reared under similar conditions to higher-temperature populations, show more rapid development to larger adult body size. The developmental and evolutionary effects of temperature on rate of development are therefore in opposite directions, while those on body size are in the same direction (for review, see Partridge and French, in press).
An important issue with environmental effects is their timing. Current environment is frequently important. For instance, female fruit flies (Drosophila) can switch their egg production on and off almost instantaneously in response to altered food supply (Chapman et al., 1994). Data from large cohorts of medflies (Carey et al., 1992) showed that the way in which adult mortality rate changed with age was affected by the kind of container and social environments in which the flies were kept. Environmental effects can also be persistent. In Drosophila, larval density during rearing, to take one of many relevant variables, affects the size, life span. and fertility of adults (e.g., Clark and Feldman, 1981; Zwaan et al., 1991).
Environmental variables can also interact in their effects on the life history. For instance, adult female Drosophila that had been reared under standard conditions were exposed to different levels of nutrition, chosen to span the level that the females usually encountered, and to two levels of opportunities for mating. Excess mating can reduce the life span of female Drosophila (Fowler and Partridge, 1989; Chapman et al., 1995). Nutrition and opportunity for mating interacted in their effect on female lifetime reproductive success, because the cost of mating reduced lifetime reproductive success only at the highest level of nutrition (Chapman and Partridge, 1996).
Life histories, like any other trait evolving under natural selection, are expected to evolve toward a form optimal only for the environments in which selection occurs. There has been a great deal of theoretical and empirical work on how sensitivity to the environment of life-history traits, their "norm of reaction" or "phenotypic plasticity," is expected to evolve for the range of environments naturally encountered (e.g., Gomulkiewicz and Kirkpatrick, 1992; Scheiner, 1993; de Jong, 1995). The conclusions of such models depend, to some extent, upon their assumptions. For instance, increased plasticity might or might not carry a cost. The trait could be one like insect body size, which is fixed at the onset of adulthood, so that plasticity is expressed only once during the life history, or one like egg-laying rate in female Drosophila, which can vary throughout the adult period. Different environments could be encountered equally or unequally frequently. The general theoretical findings are that phenotypic plasticity will evolve under a wide range of circumstances. Further, where different environments are encountered unequally frequently, the pattern of plasticity will produce the most appropriate expression of the trait and minimum additive genetic variance for it in the environment most frequently encountered.
The environments encountered by present-day human populations undoubtedly differ widely from those in which their life history evolved. Factors such as nutrition and disease are probably particularly important. Another peculiarity of human populations is that as their economic status rises, their fecundity declines by design. We are therefore probably dealing with fecundity schedules for both males and females that are very different from those that applied, on average, in the evolutionary past. In humans, changed environment can alter circumstances during development, especially for growth, in a way that is known to affect age at first breeding and adult phenotype. Within the adult period, both fecundity schedule and age-specific death rates are affected by environment. Many peculiarities of modern human populations could be manifestations of adaptive phenotypic plasticity that has evolved in response to the various levels of resources encountered in the past. Some insight into the sorts of plasticity patterns we might expect in response to these variables could, therefore, come from a knowledge of the effects of social and economic structuring in human populations before the demographic transition. The effects on postreproductive survival patterns would be particularly interesting. Whether the effects of the demo-
graphic transition on human life histories are so large as to make them difficult to interpret in evolutionary terms remains to be seen.
Individual Variation
If individuals differ in either their baseline intrinsic state or how it changes with age, the shape of the aging function for the population will partly reflect these individual differences (Vaupel, in this volume). Individual variation can mean that those individuals that become the oldest old do so because they are a nonrandom sample of their cohort.
Individual variation can be genetic or environmental in origin. Our best method of predicting the form of genetic variation comes from evolutionary theories of aging. The pattern of genetic variation predicted is different for the mutation-accumulation and pleiotropy mechanisms (Charlesworth and Hughes, 1996), with the former predicting an increase in additive genetic variance for survival and fertility with age and the latter not doing so, unless by making some restrictive assumptions. We do not yet know the relative importance of these two processes in determining the rate of aging or whether their contributions vary among different kinds of organisms. With mutation accumulation, we expect individually rare alleles at a large number of loci, but that is about all we can say with confidence. Certainly among the oldest old we should expect to see the effects of these rare mutations, and they must be expected to underlie much of any familial resemblance in age at and cause of death, but we cannot at present predict the pattern of individual variation in age at death that they will produce.
Environmental variation could also cause individual variation, perhaps more than that generated by genotype differences. It would be illuminating to obtain data from one or more of the model organisms on what kinds of individuals, in demographic terms, survive to become the oldest old. For instance, some studies with Drosophila have suggested that environmental manipulations that reduce adult fitness (high larval crowding, poor nutrition) can, paradoxically, extend life span (e.g., Luckinbill and Clare, 1986; Chapman and Partridge, 1996). However, at the same time lifetime reproductive success is reduced because of a reduction in fecundity. Furthermore, those individuals with the highest lifetime success produce their offspring, on average, earlier in life and cease reproduction soonest. If these findings generalize to other species, then the oldest old will not include an excess of those individuals that were initially the most robust members of their cohort, but rather of the ones that have been less successful in producing offspring earlier in life. A fuller exploration of these kinds of environmental effects could be worthwhile.
In the context of the oldest old of the human population, one way to get at the effects of past environmental influences would be to exploit natural reversal experiments, where individuals move to a different environment part way through life. Their subsequent survival and fertility can then be compared with those of
individuals who always inhabited the new environment or who always stayed in the old environment. This comparison is analogous to the use of temperature-sensitive mutations to study the effects of mutant gene expression at different ages or of experimental reversal of environmental variables such as reproductive status part way through life. These sorts of experiments raise quite an interesting paradox. In Drosophila this type of reversal experiment has been used to examine the timing of the increase in male mortality caused by sexual activity (Partridge and Andrews, 1985). The results suggested that the elevation of mortality is instantaneous and ceases or commences with the end or onset of elevated sexual activity. However, individual differences in frailty, whether genetic or environmental in origin, could complicate the interpretation of these experiments. If mortality rates were higher among the sexually active than among the celibate males before the reversal of reproductive status occurred, then the survivors of sexual activity would presumably be, on average, less frail than the survivors of celibacy. It is therefore possible that in the above experiment sexual activity did have lingering effects on male death rate, but that these were masked by the effects of the variation in frailty (Prowse and Partridge, 1997). We therefore need to apply some caution when studying the timing of environmental effects on death rate. In addition, these experiments failed to account for any possible effects on fertility, which might alter the conclusions about an absence of intrinsic changes in state.
Interactions
Intrinsic differences between individuals and intrinsic changes caused by aging may interact with the environment, producing a variety of patterns of age-related change in mortality or fertility. Benign environments, defined as those that, on average, improve individual performance, might be expected particularly to favor the frail and the aged, and there are many examples of such changes in relative viability with, for instance, population density. We know rather little about these interaction effects for life histories generally, and they warrant further study.
One of the most important interaction effects for life histories may involve patterns of phenotypic plasticity in relation to individual variation in frailty. There is increasing interest in theoretical analysis of life-history optimization in individuals that differ in state. The issue first became clearly apparent in work on optimization of clutch size in birds, where experimental work with field populations revealed that individual optima varied (e.g., Gustaffson and Sutherland, 1988; Pettifor et al., 1988). The basic finding was that, if brood size was experimentally manipulated, those individuals that laid larger clutches were also more capable of rearing those clutches successfully to fledging. These findings have helped to encourage the development of theoretical analysis of state-dependent life-history optimization (e.g., McNamara and Houston, 1996). Individual varia-
tion in human populations may reflect this type of plasticity and could have a major bearing on individual variation in the aging process. These findings underline the need to understand what kinds of individuals survive to later ages.
Broader Implications Of An Evolutionary Approach For Human Aging
An evolutionary approach to aging has a number of messages in addition to any for age-specific death rates. Any gene that does not have an effect on fertility or survival would be lost by mutation pressure, so all genes must influence one or both of these traits at some ages (Partridge and Barton, 1993a). The deduction is that genetic variation for the rate of aging will be polygenic, making aging a quantitative genetic trait, as has been frequently demonstrated (Hutchinson and Rose, 1991; Hutchinson et al., 1991; Hughes, 1995a,b). We are also not expecting most genes that influence the aging rate to be specialists, in that they did not evolve to their present form to cause aging, which, is, in fact, a deleterious trait. Aging is not like development (Partridge and Harvey, 1993; Martin et al., 1996), which is an orchestrated process under the control of a highly specialized gene hierarchy. Rather, genes active during development will affect the eventual resistance of the adult soma and reproductive processes to damage, while others that determine, for instance, levels of reproduction and activity will affect the rate at which damage is accumulated. Some genes, to be sure, will determine the extent to which damage is repaired, but they may well be the minority.
Another prediction is that, because the rate of aging evolves in response to the external hazards, we would expect the different systems underlying continuing survival and fertility to wear out in parallel, because of the common pattern of natural selection maintaining them in the face of damage. In consequence, we would not expect to find major genes that reduce the aging rate, unless they also had a deleterious effect on some other life-history aspect. Even if a genetic or other kind of intervention ameliorated one aspect of the aging process, we would expect the recipients to become infertile or die as a result of failure of other systems at a time very little later than the controls. The same argument does not apply to single genetic lesions that accelerate the aging process; they can and do cause early death and infertility (e.g., Yu et al., 1996). There have been reports of reduction in the rate of aging through single gene mutations or transgenes (see Johnson, in this volume), which contrasts interestingly with the predictions of evolutionary theory. There could be several explanations. (1) If the whole life history is not measured, perhaps because only survival is examined, there may well be negative, compensatory effects on fertility, preadult survival, or rate of development to adulthood. (2) If the effect of the genetic change is measured in an environment other than that where the life history evolved, complications from gene-environment interaction could result. (3) Particularly where the effects of transgenes are measured, the possibility of mutagenic effects of the
insert, over and above any effects of transgene expression, must be considered. It is essential to control for the insert size, the position of the transgene in the genome, and the genetic background into which it is placed (Kaiser et al., 1997), controls rarely found in published studies.
Conclusions
An evolutionary approach does allow some strong predictions about aging patterns but does not, at present, have strong predictive power for patterns of mortality and morbidity in the elderly members of human populations. However, refinement of theory and data should greatly improve our understanding of these issues, and there may be other ways in which evolutionary thinking can inform future work on these issues. For instance, it would be helpful, at a physiological level, to know what the main tradeoffs are in mammalian life histories. It would be useful to know the cost of repair processes in any organism and how the level of different repair processes interacts with various environmental variables to determine rate of intrinsic deterioration. To this end, it would be extremely illuminating to know whether there are any major "public mechanisms" of aging (Martin et al., 1996), common to all species, in which case work on detailed mechanisms in model species could be directly relevant to humans. It would be useful to look for biomarkers of intrinsic state in natural populations and to learn more about evolution of norms of reaction for life-history traits, to predict the impact of changed environment.
Perhaps one of the most useful contributions of evolutionary thinking in this area is cautionary. We are interested in human senescence because we wish to maintain healthy life as long as possible. Because the rate of aging is affected by genetic variation, it is tempting to apply the kind of thinking to it that is applied to genetic disorders, including exploration of the possibility of genetic intervention. However, everything that we know about the evolution of aging suggests that it is probably the most polygenic of all traits. Fertility and viability are affected by all genes in the genome. The prospects for genetic intervention, therefore, must be explored with some circumspection.
References
Abrams, P.A. 1991 Fitness costs of senescence: The evolutionary importance of events in early adult life. Evolutionary Ecology 5:343-360.
1993 Does increased mortality favor the evolution of more rapid senescence? Evolution 47:877-887.
Abrams, P.A., and D. Ludwig 1995 Optimality theory. Gompertz law, and the disposable soma theory of senescence. Evolution 49:1055-1066.
Austad, S.N., and K.E. Fischer 1991 Mammalian aging, metabolism, and ecology: Evidence from the bats and marsupials. Journal of Gerontology 46:B47-53.
Blarer, A., M. Doebeli, and S.C. Steams 1995 Diagnosing senescence: Inferring evolutionary causes from phenotypic patterns can be misleading. Proceedings of the Royal Society of London, Series B. Biological Sciences 262:305-312.
Brooks, A., G.L. Lithgow, and T.E. Johnson 1994 Mortality rates in genetically heterogeneous populations of Caenorhabditis elegans. Science 263:668-671.
Carey, J.R., P. Liedo, D. Orozco, and J.W. Vaupel 1992 Slowing of mortality rates at older ages in large medfly cohorts. Science 258:457-461.
Chapman, T.. L.F. Liddle, J.M. Kalb, M.F. Wolfner, and L. Partridge 1995 Cost of mating in Drosophila melanogaster females is mediated by male accessory gland products. Nature 373:241-244.
Chapman, T., and L. Partridge 1996 Female fitness in Drosophila melanogaster: An interaction between the effects of nutrition and encounter rates with males. Proceedings of the Royal Society of London, Series B. Biological Sciences 263(1371):755-759.
Chapman, T., S. Trevitt, and L. Partridge 1994 Remating and male-derived nutrients in Drosophila melanogaster. Journal of Evolutionary Biology 7:51-69.
Charlesworth, B. 1980 Evolution in Age-Structured Populations. 1st ed. New York: Cambridge University Press.
1994 Evolution in Age-Structured Populations. 2nd ed. New York: Cambridge University Press.
Charlesworth, B., and K.A. Hughes 1996 Age-specific inbreeding depression and components of genetic variance in relation to the evolution of senescence. Proceedings of the National Academy of Sciences U.S.A. 93(12):6140-6145.
Charlesworth, B., and L. Partridge 1997 Ageing: Leveling of the grim reaper. Current Biology 7:R440-R442.
Clark, A.G., and M.W. Feldman 1981 Density dependent fertility selection in experimental populations of Drosophila melanogaster. Genetics 98:849-869.
Cole, L.C. 1954 The population consequences of life history phenomena. Quarterly Review of Biology 29:103-137.
Comfort, A. 1979 The Biology of Senescence, 3rd ed. Edinburgh: Churchill Livingstone.
Curtsinger, J.W., H. Fukui, D. Townsend, and J. Vaupel 1992 Failure of the limited-life span paradigm in genetically homogeneous populations of Drosophila melanogaster. Science 258:461-463.
de Jong, G. 1995 Phenotypic plasticity as a product of selection in a variable environment. American Naturalist 145:493-512.
Finch, C.E. 1990 Longevity, Senescence and the Genome. Chicago: University of Chicago Press.
Finch, C.E., M.C. Pike, and M. Whitten 1990 Slow mortality rate accelerations during aging in some animals approximate that of humans. Science 249:902-905.
Fowler, K., and L. Partridge 1989 A cost of mating in female fruitflies. Nature 338:760-761.
Gaillard, J-M., D. Allaine, D. Pontier, N.G. Yoccoz, and D.E.L. Promislow 1994 Senescence in mammals: A reanalysis. Evolution 48:509-516.
Gomulkiewicz, R., and M. Kirkpatrick 1992 Quantitative genetics and the evolution of reaction norms. Evolution 46:390-41,.
Gustaffson, L., and W.J. Sutherland 1988 The costs of reproduction in the collared flycatcher Ficedula albicollis. Nature 335:813-815.
Hamilton, W.D. 1966 The molding of senescence by natural selection. Journal of Theoretical Biology 12:12-45.
Hughes, K.A. 1995a Evolutionary genetics of male life history characters in Drosophila melanogaster. Evolution 49:521-537.
1995b The inbreeding decline and average dominance of genes affecting male life-history characters in Drosophila melanogaster. Genetical Research 65(1):41-52.
Hutchinson, E.W., and M.R. Rose 1991 Quantitative genetics of postponed aging in Drosophila melanogaster. I. Analysis of outbred populations. Genetics 127:719-727.
Hutchinson, E.W., A.J. Shaw, and M.R. Rose 1991 Quantitative genetics of postponed aging in Drosophila melanogaster. II. Analysis of selected lines. Genetics 127:729-737.
Kaiser, M., M. Gasser, R. Ackermann, and S.C. Steams 1997 P-element inserts in transgenic flies: a cautionary tale. Heredity 78(1):1-11.
Kirkwood, T.B. 1977 Evolution of ageing. Nature 270(5635):301-304.
Kirkwood, T.B.L., and M.R. Rose 1991 Evolution of senescence: Late survival sacrificed for reproduction. Philosophical Translations of the Royal Society of London, Series B. Biological Sciences 332:15-24.
Luckinbill, L.S., R. Arking. M.J. Clare, W.C. Cirocco, and S.A. Buck 1984 Selection for delayed senescence in Drosophila melanogaster. Evolution 38:996-1003.
Luckinbill, L.S., and M.J. Clare 1986 A density threshold for the expression of longevity in Drosophila melanogaster. Heredity 56:329-335.
Martin, G.M., S.N. Austad, and T.E. Johnson 1996 Genetic analysis of aging: Role of oxidative damage and environmental stresses. Nature Genetics 13:25-34.
McNamara, J.M., and A.I. Houston 1996 State-dependent life histories. Nature 380:215-221.
Medawar. P.B. 1952 An Unsolved Problem of Biology. London: H.K. Lewis.
Nesse, R.M. 1988 Life table tests of evolutionary theories of senescence. Experimental Gerontology 23:445-453.
Partridge, L. 1987 Is accelerated senescence a cost of reproduction? Functional Ecology 1:317-320.
Partridge, L., and R. Andrews 1985 The effect of reproductive activity on the life span of male Drosophila melanogaster is not due to an acceleration of senescence. Journal of Insect Physiology 31:393-395.
Partridge, L., and N.H. Barton 1993a Optimality, mutation and the evolution of ageing. Nature 362:305-311.
1993b Evolution of aging: Testing the theory using Drosophila. Genetica 91:89-98.
1996 On measuring the rate of ageing. Proceedings of the Royal Society of London, Series B. Biological Sciences 263(1375): 1365-1371.
Partridge, L., and K. Fowler 1992 Direct and correlated responses to selection on age at reproduction in Drosophila melanogaster. Evolution 46:76-91.
Partridge, L., and V. French 1996 Why get big in the cold? Pp. 265-292 in Animals and Temperature, L.A. Johnston and A.B. Bennett, eds. Cambridge: Cambridge University Press.
Partridge, L., and P.H. Harvey 1993 Methuselah among nematodes. Nature (News and Views) 366:404-405.
Partridge, L., and R. Sibly 1991 Constraints in the evolution of life histories. Philosophical Transactions of the Royal Society of London, Series B. Biological Sciences 332:3-13.
Pettifor, R.A., C.M. Perrins, and R.H. McCleery 1988 Individual optimization of clutch size in great tits. Nature 336:160-162.
Pianka, E.R., and W.S. Parker 1975 Age-specific reproductive tactics. American Naturalist 109:453-464.
Promislow, D.E.L. 1991 Senescence in natural populations of mammals: A comparative study. Evolution 45:1869-1887.
Prowse, N., and L. Partridge 1997 The effects of reproduction on longevity and fertility in male Drosophila melanogaster. Journal of Insect Physiology 43(6):501-512.
Roff. D.A. 1992 The Evolution of Life Histories: Data and Analysis. New York: Chapman and Hall.
Rose, M.R. 1982 Antagonistic pleiotropy, dominance, and genetic variation. Heredity 48:63-78.
1984 Laboratory evolution of postponed senescence in Drosophila melanogaster. Evolution 38:1004-1010. 1991 Evolutionary Biology of Aging. Oxford: Oxford University Press.
Schaffer, W.M. 1974 Selection for optimal life histories: the effects of age structure. Ecology 5:291-303.
Scheiner, S.M. 1993 Genetics and evolution of phenotypic plasticity. Annual Review of Ecology and Systematics 24:35-68.
Steams, S.C. 1992 The evolution of life histories. Oxford: Oxford University Press.
Tatar, M., J.R. Carey, and J.W. Vaupel 1993 Long-term cost of reproduction with and without accelerated senescence in Callosobruchus maculatus: Analysis of age-specific mortality. Evolution 47:1302-1312.
Yu, C-E, J. Oshima, Y.H. Fu, E.M. Wijsman, F. Hisama, R. Alisch, S. Mathews, J. Nakura. T. Miki, S. Ouasis, G.M. Martin, J. Mulligan, and G.D. Schellenberg 1996 Positional cloning of the Werner's Syndrome gene. Science 272:258-262.
Williams, G.C. 1957 Pleiotropy, natural selection, and the evolution of senescence. Evolution 11:398-411.
1966 Natural selection, the cost of reproduction and a refinement of Lack's principle. American Naturalist 100:687-690.
Zwaan, B., R. Bijlsma, and R.F. Hoekstra 1991 On the developmental theory of aging. 1. Starvation resistance and longevity in Drosophila melanogaster in relation to pre-adult breeding conditions. Heredity 66:29-39.
Zwaan, B., R. Bijlsma, and R.F. Hoekstra 1995 Direct selection on life span in Drosophila melanogaster. Evolution 49:649-659.


















