
III
VACCINES
Bison in captive herds usually are vaccinated using protocols established for cattle, and some elk are vaccinated at feeding grounds. Vaccination in bison and elk is one part of an overall strategy that could be used to control or eliminate B. abortus in the Greater Yellowstone Area (GYA), but much research is needed before current vaccines can be judged adequate for use in those species. The following sections discuss current vaccines and describe the biosafety and efficacy standards that new and existing vaccines must meet if they are to be used effectively as part of a control or eradication program.
EXISTING VACCINES
Four vaccines are used against brucellosis: Brucella abortus strain 19 (S19) and B. abortus strain RB51, Rev 1 against B. melitensis, and strain 2 against B. suis. S19 and RB51 were developed to prevent brucellosis in cattle and have been used in bison and elk. They do not protect completely against infection or abortion in cattle, and data on their limited use in bison and elk suggest a similar degree of protection. Although modern techniques of molecular biology have revealed differences in the arrangement of DNA sequences between these vaccine strains and the virulent field strains, the differences are not reflected in the antibody responses of the host.
Further research on the vaccines is required before adequate data will be available for interpretation of safety and efficacy. If the vaccines prove inadequate, several approaches are available for vaccine development: development of a new or mutant strain of B. abortus designed for use in bison and exploration of the use of adjuvants in association with vaccines. Yellowstone
National Park (YNP) bison are not needed for human consumption so some adjuvants not approved for cattle might be useful in bison and elk.
Strain 19
B. abortus strain 19 is a low-virulence, live vaccine developed for use in cattle. It is cleared from the body of the cow more quickly than is the virulent field strain from which it was derived. S19 lacks the eri gene for erythritol metabolism, but it is uncertain whether that gene is associated with virulence. S19 was the cornerstone of the United States Department of Agriculture program of brucellosis eradication in cattle from the 1930s to 1996. When given to cattle in calfhood, S19 has been shown to be only 67% effective in preventing infection and abortion. It has several disadvantages: it is infectious for and causes disease in humans; when given to pregnant cattle, it infects the placenta and can cause abortion; and it induces serologic responses in vaccinated calves that cannot be discriminated from serologic responses caused by field infections.
S19 in Bison
In commercial bison herds, S19 has been used for calfhood vaccination since the 1960s without important clinical sequelae. Commercial producers using S19 in bison typically use vaccine doses established for cattle. The standard dose of S19 for cattle was originally required to contain at least 10 billion live cells per milliliter (50-billion dose) on initial test and at least 5 billion per milliliter (25-billion dose) at expiration date. A reduced dose of S19 was later established as 0.3-3 billion live cells with age limits of 4-12 months for use. The reduced dose range was based on data for minimal protective doses of 0.09-4.5 billion colony-forming units (CFU) for calves 3-6 months old (Davies et al. 1980) and 0.1-90 billion for calves 4-6 months old (Deyoe 1980).
In South Dakota, which has more bison than any other state, many of the 200 small commercial bison herds are vaccinated according to state regulations. Bison are moved through chutes and vaccinated subcutaneously. One commercial herd of some 5,000 bison in South Dakota (Triple U) was vaccinated with S19 beginning in the 1960s and with RB51 since 1996.
Attempts were made in the 1940s to control brucellosis in YNP bison. A vaccination program with S19 was begun and was believed to have achieved some success in reducing the incidence of brucellosis (Barmore 1968).
Records of that program are not available, and it is not possible to judge the effect of the program on YNP bison.
In bison herds in national and state parks during the 1960s, calfhood vaccination with S19 was part of programs to control brucellosis. The herds were chronically infected with B. abortus, and within a decade of the start of an S19 vaccination and management (including test and slaughter) program, many of the herds were declared free of brucellosis (see Chapter IV). In most cases of S19 use in bison, no abortion, anaphylaxis, lameness, or other sequelae associated with vaccine use have been noted. However, records of vaccine use and effect have not been kept for most herds.
Results of biosafety and efficacy studies of the use of S19 in adult and pregnant bison suggest caution. In some experiments, S19 appeared to be more virulent in bison than in cattle and caused a high incidence of abortion when given to pregnant bison.
Experiments were done with 18 pregnant bison given 1 × 107 CFU of B. abortus strain 2308 in the conjunctival sac. In one experiment, 12 infected bison were placed in contact with 12 susceptible pregnant heifers: six aborted and two had nonviable calves, compared with nine abortions in 12 similarly inoculated cattle. Five of the 12 susceptible cattle became infected (Davis et al. 1990). In a second experiment designed to define pathogenesis, six infected bison were killed, one each week. All tissues were negative at 1 wk, only parotid and supramammary lymph nodes were positive at 2 wk, and most lymph-node samples were positive from 3-6 wk. Thymus was never positive, and spleen was positive only at the sixth week.
At a reduced dose in adult bison, S19 does induce some protection against experimental challenge but also induces a high percentage of abortions (Davis et al. 1991). When adult female bison were vaccinated with S19, abortion and infection rates were reduced in comparison with nonvaccinated bison; however, S19 caused pregnant bison to abort (Davis et al. 1991). S19 vaccine was not effective when administered to (bison) calves (Davis, unpublished data, quoted in Thorne and Herriges 1992). Abortion in bison experimentally vaccinated with S19 has been well documented, and B. abortus has been shown to replicate in epithelia of the trophoblast as it does in cattle and to cause placental inflammation and necrosis, changes that underlie abortion (Davis et al. 1990).
The responses of bison to S19 might differ from those of cattle for several reasons, including increased susceptibility of the host, vaccination of the host at an inappropriate age, and differences in social behavior that favor transmission of B. abortus (F. Enright, LSU, pers. commun., 1997).
S19 in Elk
Nearly 36,000 doses of S19 are estimated to have been given to elk, and the vaccine is considered safe and efficacious. Elk have been vaccinated on Wyoming feeding grounds with a reduced dose of B. abortus S19 given by biobullet (Thorne et al. 1981; Herriges et al. 1991), and a significant decrease in serum antibody titers has been reported (Smith et al. 1995). The Wyoming Game and Fish Department (WGFD) began giving S19 vaccine to elk on WGFD-managed feeding grounds in 1985. More than 35,000 doses of vaccines have been given on 21 elk feeding grounds. At one feeding ground, the prevalence of brucellosis has declined by 50% (P = 0.00001). The decline is attributed to vaccination because other management practices have not changed over 25 yr. No environmental hazard has been associated with S19 vaccine use for elk in Wyoming.
Although elk at Jackson Hole have been vaccinated with S19, serologic studies without culture studies will not give a true picture of the prevalence of brucellosis in the elk population, because the serologic tests cannot differentiate between a titer caused by the vaccine strain and that caused by field strains (D. Ewalt, Nat. Vet. Serv. Lab., pers. commun., 1997).
Strain RB51
B. abortus strain RB51 is a rough mutant of virulent B. abortus strain 2308 that is deficient in O-side chains of lipopolysaccharides on the bacterial surface. RB51 was naturally derived by serial passages on media containing rifampin and by selecting single colonies with rough morphology. The genome of RB51 closely resembles the genome of strain 2308 when examined with most molecular techniques. However, RB51 has a unique genetic rearrangement that differentiates it from strain 2308—one that is stable and has been maintained in all isolates of RB51 (Figure III-1). Genomic restriction-endonuclease patterns produced with pulsed-field gel electrophoresis have also demonstrated a unique ''fingerprint" for RB51 relative to other brucellae (Jensen et al. 1995, 1996). After passage in vitro or in vivo, RB51 retains its resistance to rifampin or penicillin and its susceptibility to tetracycline. A new growth medium has been developed for culture of RB51 (R. Hornsby, Nat. Anim. Dis. Ctr., pers. commun., 1997).
RB51 has replaced S19 as the required vaccine for cattle in the United States. It is genetically stable in bison, as it is in cattle, and does not revert to virulence or to smooth forms after growth in vivo. The dose used commercially
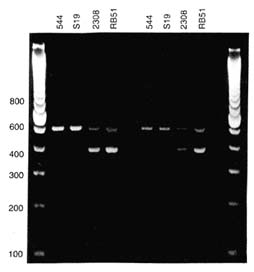
FIGURE III-1. Differentiation of B. abortus vaccine strains S19 and RB51 by a polymerase chain reaction assay (Bricker and Halling 1995). PCR amplification was done on DNA from 4 bacterial strains. DNA was resolved by electrophoresis in agarose, stained, and photographed. Sources of DNA (top) and sizes (in base pairs) of fragments of the 100-bp marker in lane 1 (left).
is 10-34 billion live organisms delivered in 2 mL. At that dose, RB51 has been shown to be protective in cattle when used as a calfhood vaccine between 3 and 10 months of age (Cheville et al. 1996). RB51 studies in bison and elk are shown in Table III-1 (Olsen et al. 1998). It is noteworthy that vaginal, rectal, ocular, and nasal swabs collected from bison 1-18 wk after vaccination in the second Olsen study listed in the table did not contain culturable B. abortus. Numbers of bison that were culture-positive in prescapular lymph node biopsy at 6, 12, 18, and 24 wk after infection were 4 of 4, 3 of 4, 1 of 4, and 0 of 3, respectively. Quantitative data suggested that most bison will clear RB51 within 18-24 wk.
RB51 has tropism for the bison placenta. It has been shown experimentally to cause endometritis and placentitis that result in abortion in pregnant bison. Of eight bison cows given RB51, two aborted (68 and 107 days after vaccination). RB51 was isolated from the cows' reproductive tissue and supramammary lymph nodes and from fetal bronchial lymph nodes and amniotic fluid. The vaccine dose used was similar to that used in cattle and might not be appropriate for bison (Palmer et al. 1996).
RB51 might not be as effective in bison and elk as in cattle, although data on this are not adequate to know. Studies of other species have yielded variable results. Experiments on the efficacy of RB51 against B. ovis in rams showed no protection (Jimenez de Bagues et al. 1995). RB51 was given at 4 × 1010 CFU subcutaneously; 6 months after vaccination, rams were challenged with 3 × 109 CFU of B. ovis and examined 8 wk later. Controls and RB51 vaccinates were found to be 100% infected, compared with 68% infection after use of Rev 1 vaccine in rams.
TABLE III-1 RB51 Vaccine Biosafety Tests
|
Author |
n |
Animals |
Route |
Dose |
Site |
Study |
|
Elk |
||||||
|
Elzer, Davis |
19 |
adult |
oral |
2-3 × 1010 |
North Dakota |
vaccination challenge |
|
Cook/Kreeger |
16 |
calves |
IM biobullet |
1 × 108 |
Wyoming |
vaccination challenge |
|
Cook/Kreeger |
16 |
calves |
IM biobullet |
1 × 109 |
Wyoming |
vaccination challenge |
|
Cook/Kreeger |
10 |
bull calves |
IM |
1 × 109 |
Wyoming |
safety |
|
Thorne |
70 |
pregnant |
IM |
1 × 109 |
Wyoming |
pathogenesis |
|
Thorne |
7 |
pregnant |
IM |
1 × 1011 |
Wyoming |
pathogenesis |
|
Kreeger/Cook |
33 |
mixed* |
oral |
1 × 109** |
Wyoming |
safety |
|
Bison |
||||||
|
Olsen |
6 |
calves |
subcutaneous |
1.2 × 1010 |
Iowa |
vaccination challenge |
|
Olsen |
10 |
calves |
subcutaneous |
1.2 × 1010 |
Iowa |
vaccination challenge |
|
Olsen |
20 |
calves |
subcutaneous |
1.5 × 1010 |
Nebraska |
safety |
TABLE III-1 RB51 Vaccine Biosafety Tests (continued)
Other Vaccines
The B. suis vaccine was developed in China. When given orally, vaccines prepared from B. suis strain 2 have been reported to reduce the incidence of porcine, bovine, ovine, and caprine brucellosis in China (Xie 1986); however, insufficient data are available to judge the usefulness of the vaccine. B. neotomae, a species from the desert woodrat, has been used to immunize caribou and reindeer against brucellosis. Although the extent of infection was reduced slightly, the number of abortions was not reduced (D. Davis, Texas A&M, pers. commun., 1997). Data on this vaccine in bison and elk have not been reported.
Mutants of B. abortus genetically engineered specifically for use in wildlife might prove effective in bison or elk. However, they have not been shown to offer substantial advantages in cattle (Cheville et al. 1993; Elzer et al. 1996). Several genes have been deleted from or added to B. abortus, but even though the resulting mutant survives in vivo, no clear advantage in vaccine efficacy has been established.
EFFICACY
Efficacy is the ability of an intervention to produce the desired beneficial effect. Live vaccines composed of organisms of diminished virulence have been most effective in reducing the incidence of brucellosis, tuberculosis, and diseases caused by most other facultative intracellular bacteria. In contrast, killed vaccines for this group of diseases have not proved efficacious. Several factors must be considered when assessing vaccine efficacy, including strain survival, route, dose, and age.
Strain Survival
When injected, a live vaccine must survive long enough to be immunogenic in the host. In vaccines composed of B. abortus for cattle, bacteria should survive in the lymph node draining the site of inoculation for at least 2 wk.
Four bison vaccinated with RB51 as young calves, allowed to mature, bred, and challenged with 107 CFU of B. abortus during the sixth month of pregnancy showed some degree of protection (Olsen et al. 1997): one aborted at 7 wk after challenge, and three completed a normal pregnancy with culturenegative calves. B. abortus was isolated from the parotid lymph node, bronchial
lymph node, or uterus of two of the three female bison that had normal births.
Route
Vaccines can be delivered in several ways, including subcutaneously by hand injection, subcutaneously by dart, subcutaneously by biobullet, and orally.
Parenteral Injection
Subcutaneous vaccination is the preferred route of vaccination with B. abortus in most species. If it is done properly, and the vaccine is used before its expiration date, S19 and RB51 have been shown to produce an immune response in bison and elk. Conjunctival or intradermal vaccinations of bison and elk with S19 or RB51 have not been reported, but those routes have been used in cattle and were nearly as effective as subcutaneous vaccination, although not as practical.
Biobullet
The biobullet is composed of hydroxypropocellulose filled with a core of lyophilized, freeze-dried vaccine; it also contains stearic acid as a lubricant and calcium carbonate for weight. The 25-caliber bullet is fired from a compressed-air gun with an unrifled barrel. Although reported as inert, the hydroxypropocellulose bullet enters the tissue and produces trauma and some degree of foreign-body stimulation, and that might have some adjuvant effects on the vaccination process.
Oral
Oral vaccination has been tried to some extent in elk and bison. In a Wyoming study, RB51 was not efficacious in elk; there was no significant difference between vaccinates and controls in response to challenge. In contrast, a study of oral RB51 in elk in North Dakota showed some effect (Elzer et al. 1996). Problems with oral vaccination include control of dose per individual and inability to control the population to be vaccinated.
The experimental vaccination of cattle (heifers) with S19 has shown some protection: none of 20 orally vaccinated pregnant heifers challenged orally with strain 2308 at midgestation aborted, whereas 10 of 19 pregnant controls aborted (14 of the 19 were culture-positive)(Nicoletti 1981).
Dose
When any new vaccine is to be used, the dose required for effect and the margin of safety for bison and elk must be determined. Doses for commercial bison and elk follow the doses recommended for cattle. For greatest efficacy, the range of doses of vaccines should be established in bison and elk. High doses of both S19 and RB51 typically produce greater immune responses than low doses. Studies on RB51 in bison have not been extensive, but it appears that the most effective doses are close to those for cattle (Olsen et al. 1997).
Age
In general, young mammals will kill B. abortus and clear the organisms from their tissues more quickly than adults. The practical side of that is that in young calves B. abortus is less likely to be retained into puberty, when it infects the reproductive system. Data are needed in bison and elk differences in response to vaccination of calves, yearlings, adult females, and adult females with multiple inoculations. Data on cattle show little difference in host responses to vaccination with age from 3 to 10 mo (Manthei et al. 1950; Cheville et al. 1996).
Other Factors to Consider
Although it often is claimed that S19 and RB51 vaccines are less effective in bison and elk than they are in cattle, this has not been established clearly. Neither vaccine is 100% effective in cattle, and both have been shown to cause abortion when given in large doses, by inappropriate routes, or to pregnant animals (Palmer et al. 1996). Efficacy studies with similar numbers have not been done in bison and elk.
Wild, free-ranging animals differ from domestic animals in many factors that influence the effects of any vaccine. Among those factors are nutrition, shelter, veterinary care, and environmental stresses, such as food availability
and the presence of geothermal energy. Other, more-subtle factors also could influence vaccine efficacy, such as the exaggerated metabolic changes that occur in bison and elk with handling, which can bring about capture myopathy.
The efficacy of live B. abortus vaccines varies with the age, sex, and genetic factors of the host. Although brucellosis in bison and elk closely resembles the disease in cattle, sheep, goats, and other ruminants, important species differences define the disease in a particular host. Variability in antibody and immune cell responses, in natural surface antigens, and in specific macrophage receptor molecules for B. abortus all are host-specific. For example, in cattle, normal serum has substantial antibrucellar properties, and it could be important to know whether that property exists in bison and elk.
Intercurrent infection at the time of vaccination can have a marked influence on the effect of a vaccine in individual animals. Infection of the host can enhance or diminish the effect of a vaccine, depending on the nature of the etiologic agent. Diseases that stimulate cell-mediated immunity might enhance the effect of some vaccines; for example, severe ringworm in cattle has been shown to increase markedly the antibody response to B. abortus (Cheville et al. 1992).
At the other extreme, systemic viral infections that replicate in the lymphoid system can suppress the effect of vaccines. In cattle, bovine viral diarrhea, bovine leukemia, and infectious bovine rhinotracheitis can have immunosuppressive effects. Similar immunosuppressive viral infections might occur in bison and elk and diminish the effects of vaccines.
Hormonal status, especially the activity of progesterone and other steroids, appears to affect how B. abortus is cleared from the host.
Duration of Immunity
The duration of immunity produced by vaccination of bison or elk is not known. Indirect evidence will come from surveys now under way in a Rhyan et al. study; data will be obtained on abortion and reproductive loss in vaccinated and nonvaccinated herds over time. It will be important to determine whether repeated vaccination is required.
Serologic Responses
RB51-vaccinated cattle that are subsequently infected with virulent B. abortus
develop antibodies that react in standard serologic tests. Experimentally, RB51 vaccinates show increased titers in the standard tube test (STT), but, in contrast with responses in nonvaccinated controls, the titers drop progressively (Figure III-2).
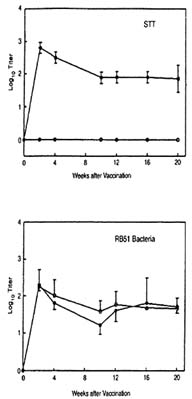
FIGURE III-2. Serologic responses of bison to vaccine RB51 '(-o-, n = 6) or Strain 19 (-•-, n = 3) in the standard tube test (STT) or to RB51 dot-blot test (Yirradiated RB51 antigen). Responses are presented as mean titer log 10.
Vaccination of animals previously infected with virulent wild-type B. abortus could lead to unusual serologic responses. That possibility is important in animals vaccinated with RB51 and other vaccines that do not induce antibodies to lipopolysaccharide components of B. abortus . No data indicate that cattle infected naturally with virulent B. abortus develop an increased serologic titer in the STT after vaccination with RB51. Some data from field tests indicate that calves that have been vaccinated with S19 do not develop increased STT titers when given strain RB51 as adults. Bison probably react in the same manner.
Serologic responses of RB51-vaccinated bison remained negative at all
times in all tests used. However, antibody responses to irradiated RB51 as an antigen were present in dot-blot tests 2 wk after vaccination. At 40 wk, two of six RB51 vaccinates had detectable antibody titers in the test for RB51, but S19 vaccinates did not.
BIOSAFETY OF VACCINES
No clinical disease or evidence of pathologic effect in tissue has been found even when large doses of S19 and RB51 vaccines were given to young bison calves subcutaneously. However, not all criteria of vaccine biosafety have been established or adequately evaluated (Table III-2).
TABLE III-2. Criteria for establishing biosafety in vaccines
|
• Clinical signs of acute disease do not appear after vaccination. • Bacteria are not present in nasal secretions, saliva, or urine. • Bacteria do not persist in the bloodstream for more than 3 days. • Bacteria do not persist in lymph nodes for more than 16 wk. • Evidence of humoral or cellular immunity is present 14 days after infection. • No inflammation or chronic tissue injury appears. • Neither placentitis nor abortion occurs in pregnant animals. • Immunosuppression after 16 wk does not cause recrudescence. • Bacteria recovered after 12 wk growth in the host are genetically identical with the vaccine strain. |
Clinical Signs of Disease
A vaccine should not induce fever, loss of appetite (anorexia), or other clinical signs of disease. Neither bison nor elk vaccinated with S19 or RB51 develop significant clinical signs, and new vaccines also should not induce clinical manifestations. Lameness, which results from persistent infection of the joints (synovitis) with B. abortus , has not been reported. Vaccination-induced anaphylaxis, manifested by rapid development of shock and sudden death, occurs in a very small proportion of cattle vaccinated with S19; anaphylaxis has not been reported in wild mammals vaccinated with S19 or RB51.
Bacteria in Body Secretions
After vaccination, the vaccine strains of B. abortus should not appear in nasal
exudates, tears, saliva, or other body secretions. Only sparse data are available, but S19 and RB51 have not been isolated from nasal swabs, saliva, tears, or urine of bison or elk. Milk has not been examined for excretion of S19, RB51, or other vaccine strains in bison and elk.
Bacteria in the Bloodstream
Live vaccine strains of B. abortus frequently can be isolated from the bloodstream in vaccinated cattle. It is common to be able to isolate the vaccine from large samples of blood for as long as 3 days after vaccination. In the blood, bacteria are typically associated with white cells. S19 and RB51 have been detected in bison in small numbers and only transiently in the bloodstream after vaccination. In one study of 10 bison given RB51, only one had culture-positive blood, and only one sample from that animal was positive (2 wk after vaccination) (Olsen et al. 1997).
Persistence in Regional Lymph Nodes
Live vaccines of B. abortus should not persist in lymph nodes draining the sites of vaccination for more than 16 wk. Replication or persistence beyond that time is associated with localization in the reproductive organs and mammary glands. In cattle, S19 and RB51 persist in lymph nodes after vaccination for times sufficient for development of immunity, but they are cleared before sexual maturity occurs. However, they might not be cleared from host tissues without risk of persistence to adulthood. Most bison vaccinated subcutaneously clear RB51 in 18 to 24 wk (Olsen et al. 1997). Biosafety experiments on RB51 in bison calves showed that replication of RB51 in lymph nodes draining subcutaneous sites of vaccination was greater than that in cattle. Three bison calves given 2.9 × 1010 CFU of S19 at the age of 3 months had S19 12 wk after vaccination but not 16 wk after vaccination (Table III-3). In bison at Ft. Niobrara National Wildlife Refuge, RB51 was present in all four lymph nodes tested 14 and 18 wk after vaccination, 22 wk after in one of four nodes, 26 wk after in three of four, and 30 wk after in none.
Immune Response
Vaccines given subcutaneously are rapidly taken up by the lymphatic system
and induce a prompt antibody response. Antibrucella antibodies appear in the serum of bison about 2 wk after bison are given S19. Cutaneous delayed hypersensitivity after intradermal injection of brucellin or other evidence of cell-mediated immunity should also be detectable at about 2 wk after vaccination.
TABLE III-3 Brucella abortus in lymph nodes of bison after vaccination
|
|
|
CFU/g node tissue (wk after vaccination) |
|||||
|
Vaccine |
n |
1 |
2 |
4 |
6 |
10 |
16 |
|
RB51 |
6 |
667 |
12,769 |
7,337 |
165 |
174 |
9 |
|
|
|
104 |
19,637 |
7,450 |
3,001 |
68 |
33 |
|
S19 |
3 |
104 |
131,967 |
7,746 |
1,216 |
324 |
0 |
|
Source: Olsen et al. 1997 |
|||||||
Absence of Inflammation or Chronic Tissue Injury
No long-term tissue injury should be associated with vaccination; for example, chronic injury to the joints, brain, or other organs should be ruled out in biosafety tests. One consequence of infection with virulent B. abortus is a depletion of lymphocytes in the lymphoid tissues. This tissue destruction should not occur in B. abortus strains used for vaccines, and neither S19 nor RB51 has been shown to produce significant lymphoid destruction (Figure III-3).
Capacity to Induce Abortion
The capacity of a vaccine to be cleared by a calf before sexual maturity is critical with respect to causing abortion. When given to pregnant cattle and bison, all commercial live brucellosis vaccines that are designed for cattle have the capacity to infect the placenta and cause abortion. Placental lesions induced by vaccines cannot be differentiated pathologically from those induced by virulent field strains (Figure III-4). A dose of 5 × 108 CFU of S19 vaccine given to pregnant bison in the second trimester of pregnancy caused abortions in 58% (Davis et al. 1991); the same dose in cattle has been said to
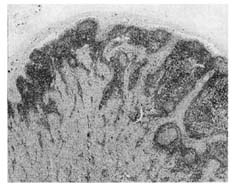
FIGURE III-3. Histology of a superficial cervical lymph node of a bison given Strain 19 vaccine 16 wk previously. No pathologic change. Germinal centers are present and medullary areas are large (Olsen et al. 1997).
induce fewer abortions (Nicoletti 1977). However, those two experiments were not critically compared. In the bison study, one S19-vaccinated cow aborted during her second pregnancy; that suggests that chronic vaccine infections can occur in bison.
Although RB51 can infect the bison placenta, it appears to be less abortigenic than S19 in bison and cattle, perhaps because of its diminished cell wall lipopolysaccharides. At high doses, RB51 vaccine has a tropism for the placenta and can cause abortion. Ten pregnant bison on a Montana ranch were vaccinated subcutaneously with 109 CFU of RB51. Two animals were sacrificed before 68 days; both showed vaccine-induced placentitis. Two abortions occurred—at 68 and 107 days after vaccination. The placenta of the aborting cows had placentitis associated with the presence of RB51 (Figure III-4).
Experimental Recrudescence
One danger in brucellosis research is the failure to detect live organisms when very few are present in tissues. That single B. abortus cells persist in a chronically infected animal in some ''vegetative state" that precludes culture never has been proved. Use of new polymerase chain reaction technology to identify one organism in tissue might be useful. Another technique to show that live B. abortus cells are present is to treat an animal with dexamethasone or some other immunosuppressive treatment that will reactivate or recrudesce bacterial growth (when B. abortus cannot be isolated from chronically infected animals) and thus allow the organism to be detected. That has been used in cattle, but bison studies would be needed to develop a system of reactivation of bacterial growth.
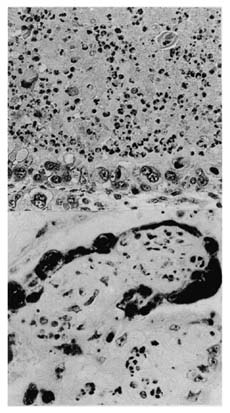
FIGURE III-4. Lesions caused by vaccine RB51 in the placenta of an aborting bison. A. Section of placentome 5 wk after vaccination with Strain RB51. Arcade zone and trophoblast layer contain large epithelial cells bearing B. abortus. B. Strong immunoreactivity for strain RB51 antigen in trophoblast epithelial cell cytoplasm (arrow); alkaline phosphatase (Palmer et al. 1996).
Genetic Stability
In establishing biosafety, the vaccine strain isolated after infection in the animal for several weeks must be genetically identical with the strain in the vaccine; for example, the bacterial strain must not mutate or revert during replication in the host. S19 has been shown to be stable in cattle after field use for more than 6 decades. RB51 has also been proved to be genetically stable in experiments in cattle, goats, and mice.
VACCINATION PROGRAM SUCCESS
The Greater Yellowstone Interagency Brucellosis Committee has agreed that "vaccination of bison and elk would achieve short term goals of brucellosis
control and reduced risk of transmission to cattle." It is unlikely that a vaccination program for bison would succeed without a preliminary or concomitant program for elimination of brucellosis in elk. Re-emergence of brucellosis in a free bison herd is likely to occur if two conditions remain: brucellosis in winter elk feeding grounds is not controlled to prevent reinfection of bison from elk, and bison populations remain high.
Although it has been stated that brucellosis cannot be eradicated from free-ranging wildlife without eradicating the wildlife, it is certain that brucellosis can be eliminated from YNP with combinations of vaccination and culling. However, until a long-term controlled vaccination study is done, no assumptions about whether brucellosis can be eliminated by vaccination should be made. One measure of success would be a reduction in rates of abortion that would directly result in diminished bison-to-cattle and bison-to-bison transmission. Bison are more likely to prevent the long-term elimination of brucellosis from elk than vice versa.


















