
2
Changes to the Biology and Biochemistry of Ecosystems
SUMMARY
The study of large-scale ecosystems has become a rapidly maturing field of science. With the impetus of global change research, such studies have shown major successes over the past decade. Improved fundamental understanding of marine and terrestrial ecosystems and hydrology has already led to practical applications in weather and climate modeling, air quality, and better management and natural hazards responses for water, forest, fisheries, and rangeland resources. The development of spatially resolved global-scale ecosystem models has occurred only during the past five years. Computing capability and remote sensing technology have further driven change in the nature of the field. The capability has emerged not only to model at global scales but also to exploit data at these scales. Models have been developed and rejected based on the use of such data. Historically, large-scale ecosystem studies have also been integrative and multi-disciplinary, with problems often worked from beginning to end with significant interactions with the human dimension components. Some of this experience stems from applied roots in the field and traditional links with agricultural, forestry, and fisheries issues, as well as environmental policy and assessment. In fact, as in atmospheric chemistry (see Chapter 5), there is a rich history of assessment at all spatial scales.
Areas of success in large-scale ecosystem studies include the following:
-
Field and theoretical studies that have laid the foundation to understand the roles of vegetation and soils in weather and climate and that have advanced our methods for interpreting satellite data. Field experiments
-
planned for the Mississippi and Amazon River basins will complete this series of studies.
-
Development of satellite observation techniques, ground-based observations, and models to determine changes in land cover type and spatial and seasonal changes of vegetation.
-
Clarifying the role of nutrients in large-scale interactions of ecosystems with the atmosphere. The effects of nutrients such as nitrogen and phosphorus must now be systematically incorporated into global models of land-atmosphere interactions.
-
Implementation of an ambitious program to measure and model the sources and sinks of CO2 and trace gases from biological and biomass burning sources. This new information will facilitate the development of an observing system to determine trends and patterns of emissions and uptake at continental scales.
-
Oceanic time series observations that have revealed previously unknown year-to-year variations in coupled ocean biology, chemistry, and physics, linked to climate variability.
-
Regional ocean carbon studies that have quantified seasonal marine ecosystem effects on atmosphere-ocean CO2 exchange, and El Niño-related variations in equatorial Pacific sources and sinks of CO 2.
-
Modeling the impacts of climate change and variability on agricultural and forest ecosystems.
Overall, the U.S. Global Change Research Program (USGCRP) has been successful in advancing the science and tools required for space-based assessment of ecosystem change. The synergistic instrument complement consisting of the Earth Observing System (EOS) AM-1 and PM-1 platforms, combined with data from other ocean-sensing satellites, will largely satisfy the satellite data needs of the ecosystems community and will result in a massive improvement in the quality of remote observations. The ground- and ocean-based components of the program have had varying degrees of success. Atmospheric science components (biophysics and trace gases) have had the strongest programs. The more ecological components (vegetation and land cover) and integrative components (ecosystem manipulation experiments) have been supported on a rather ad hoc basis.
The Research Imperatives for the future are as follows:
-
Land surface and climate. Understand the relationships between land surface processes and weather prediction and changing land cover and climate change.
-
Biogeochemistry. Understand the changing global biogeochemical cycles of carbon and nitrogen.
-
Multiple stresses. Understand the responses of ecosystems to multiple stresses.
-
Biodiversity. Understand the relationship between changing biological diversity and ecosystem function.
INTRODUCTION
The ecosystems of the world are critical foundations of human society. People depend on ecosystems extensively for goods and services. Ecosystems provide such commodities as food, construction materials, and pharmaceuticals. In the context of global change, humanity's dependence on the biosphere for climate regulation, air quality, and clean water has also become starkly apparent. Thus, research on ecosystems in global change plays a dual role. First, decisions relating to climate variability, climate change, and other environmental problems require that we understand the impacts of climate, air pollution, and changing ultraviolet radiation on forests, agriculture, livestock, water resources, fisheries, biological diversity, and other critical life support systems. “Impacts research ” builds on the foundation of basic and applied research in agronomy and soil science, forest science, fisheries, ecology, and other well-established disciplines, in a context increasingly influenced by new concerns (climate change, tropospheric pollution, ultraviolet-B). An ambitious effort is under way to conduct a U.S. national assessment of the potential consequences of climate variability and change to provide a detailed understanding of the consequences of climate change for the nation, including the interactive effects of environmental changes to climate, atmospheric chemistry, sea level, water quality, and land use. This chapter describes the research that is under way to provide appropriate links to that activity by emphasizing the scientific aspects of managed ecosystems, especially at the regional scale.
Second, managing global change must also recognize the role that ecosystems play in modifying the atmosphere and hence the ocean-atmosphere-land climate system. We now know that vegetation and soils influence climate by controlling the amount of radiation reflected or absorbed, the evaporation of water, and other direct feedbacks to temperature, precipitation, and weather systems. Terrestrial ecosystems store a great deal of carbon and influence atmospheric CO2 both by releasing carbon as a result of land use (such as deforestation and agriculture) and by taking up carbon (the so-called missing sink). Marine ecosystems also influence oceanic carbon storage, interacting with physical and chemical processes. Ecosystems are also potent sources and sinks of other trace gases such as methane and nitrous oxide. Climate cannot be viewed as a force external to ecosystems: ecosystems participate in the shaping of weather, climate, atmospheric composition, and climate change.
This view of ecosystems, as both responding to and controlling environmental change, is one of the great intellectual and practical contributions of global change research. It has implications for a wide range of issues, from the improvement of weather forecasts (by taking into account the state of vegetation) to
decisions about whether fossil fuel emissions should be limited and by how much.
Ecology contributes a unique perspective to global change research. While the geophysical sciences begin conceptually with a unified physical-chemical view of systems (based in fluid dynamics, thermodynamics, and photochemistry), the underlying paradigm of ecology emphasizes the diversity of ecosystems, resulting from the evolutionary history of organisms, and the soils and landforms or water bodies they inhabit. This perspective, of seeking understanding from the similarities and differences of processes across a range of environments, has become important in interdisciplinary Earth system research.
Planning the Program
At the outset of global change research, ecology as a discipline emphasized organism- to local-scale investigations, and the field 's ability to address problems at even the landscape scale was limited. In addition, collaborations between ecologists, climatologists, and atmospheric chemists only began in the early 1980s. As a result, when the USGCRP and International Geosphere-Biosphere Program (IGBP) began, there was a substantial effort to develop the intellectual infrastructure within ecology to tackle problems at the global scale. A series of National Research Council (NRC) reports on global change and the IGBP planning process paid substantial attention to large-scale ecological issues, and the community also organized many important workshops and meetings. As a result, ecology has become much better prepared to take on science issues at scales from landscape to global on a breadth of issues and using tools not imagined a decade and a half ago.
Some issues have remained over that period. The focus on both ecosystem feedbacks and ecosystem impacts has been consistent in NRC and IGBP guidance throughout this period. However, as the science has evolved, the specific Research Imperatives have evolved substantially. It is worthwhile to review NRC guidance on global change and ecological research. For example, in a 1986 report1 the summary recommendation (given with reference to the IGBP) was that “the initial priority . . . is to obtain additional experimental data, so that new models can be developed to extrapolate ecological responses to environmental changes that have not been experienced in the past.” That report called for “laboratory and field experiments at the organism level and compilation of existing data on population and community patterns. ” It also observed that “experiments are needed on intact ecosystems, using large-scale manipulations ” and that “in the long-term, ecosystem models must be assembled that couple population-community models with process-functional models.”
This agenda had a major influence on the approaches taken by ecologists for both marine and terrestrial ecosystems, and all three of the NRC's recommended agenda areas were pursued in parallel by the community. In addition, long-term
and large-scale observational (as opposed to manipulative) studies became a major component of research, both building on the foundation of the National Science Foundation's (NSF) Long-Term Ecological Research (LTER) program and arising from the increasingly fruitful collaboration with the Earth sciences community (in which observational campaigns play a larger role than in the largely experimental discipline of ecology). By the 1990s the research agenda had come into sharper focus. In 1994 an NRC report, the “Chapin report”2 on terrestrial ecosystem research listed six major research areas:
-
the interactive effects of CO2, climate, and biogeochemistry;
-
factors that control trace gas fluxes;
-
scenarios for managed and unmanaged ecosystems;
-
how global change will influence biodiversity;
-
how global change will affect biotic interactions with the hydrological cycle; and
-
how global change will affect the transport of water, nutrients, and materials from land to freshwater and coastal zones.
The Chapin report (NRC, 1994) also specifies the following needs:
-
experiments that determine ecosystem responses to interactions among elevated CO2, temperature, water, and nutrients;
-
research to predict the role of landscape-scale processes, including land use; and
-
research to determine how changes in species composition affect the functions of ecosystems, which is urgently needed and unlikely to proceed without focused attention.
Finally, the report lists as a major theme “the development and use of comprehensive models of ecological and physical systems” as a means of linking small-scale understanding to large-scale processes.
The NRC was not the only forum in which the role of ecosystems in global change was discussed. The late 1980s to early 1990s saw the rapid development of the IGBP Global Change and Terrestrial Ecosystems Core Project (GCTE) and the Joint Global Ocean Flux Study Core Project (JGOFS), addressing terrestrial and marine ecosystems, respectively. In addition, the International Global Atmospheric Chemistry (IGAC) program became involved in studying biological sources of trace gases, 3 the Biological Aspects of the Hydrological Cycle Core Project began activities in biophysical research, and the Scientific Committee on Problems of the Environment (SCOPE) organized several important collaborations on ecosystems and global change.4 The IGBP elaborated considerably on the science agenda for global change research, and its deliberations have been documented in extensive reports. The IGBP GCTE research plan is in many
ways consistent with priorities enunciated in the Chapin report and earlier NRC documents, though it presents a substantially more detailed vision for studying managed (especially agricultural) ecosystems.
Critical Results and the Development of Large-Scale Ecology
In view of this scientific vision, what were the critical results and advances in ecosystem research during the first decade of the USGCRP? The areas of progress have been diverse, and many critical advances have been highly interdisciplinary. Discussed below are some of the most important areas of progress.
Climate and Ecosystem Change: Evolution of Models and Observations
Techniques and datasets for inferring the behavior of ecosystems from large-scale observations evolved rapidly during the 1990s. The use of inverse modeling a to deduce spatial and, later, temporal patterns of terrestrial and oceanic CO2 exchange produced a qualitative change in perceptions of the likely nature of terrestrial sinks. Although the use of inverse modeling began in the geophysical community, where both inverse modeling techniques and CO2 global observations were developed,5 collaborations to expand use of the technique rapidly grew to include ecologists.6 Inverse modeling showed a sink of CO2 in northern latitudes, through discrepancies between the observed interhemispheric gradient of CO2 and the values predicted based on fossil emissions and characteristics of interhemispheric transport. Whereas initial analyses had reached different conclusions about the distribution of this sink between marine and terrestrial systems, later analyses using 13C in CO2 and measured O 2 indicated a substantial terrestrial sink (see Figure 2.1). This sink has been and remains difficult to quantify or even detect in forest and soil inventory measurements; atmospheric measurements remain the most conclusive evidence for the location of the so-called missing sink. Applications of the inverse methodology over time have also suggested correlations between climate and terrestrial CO2 exchange at hemispheric to global scales. These observations remain preliminary but provide a foundation for future monitoring of global source and sink patterns.
The relationship of terrestrial carbon storage to climate is fundamental to understanding the interactions between climate and ecosystems that may occur during future climate changes. This subject has been addressed for terrestrial systems by a combination of experimental lab and field studies and by observational programs and data synthesis. 7 In addition, there has been vigorous model-
|
a |
Inverse modeling is defined as modeling where the chain of inference runs opposite the chain of causation: in the case of the carbon cycle, sources and sinks are modeled from atmospheric concentrations and transport. |
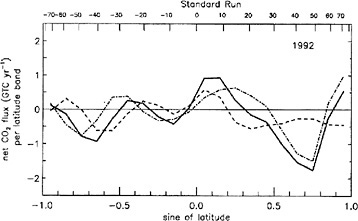
FIGURE 2.1 Latitudinal ocean/land partitioning of the sources and sinks of CO2 versus latitude. The continuous line is the net flux of CO2 after removal of fossil fuels. The dashed line is the net flux of CO2 exchanged with the oceans. The dotted line is the net flux exchanged with land ecosystems. The sum of ocean and land fluxes equals the total net flux of CO2. SOURCE: Ciais et al. (1995b). Courtesy of the American Geophysical Union.
ing of climate effects on both terrestrial and marine ecosystems. 8 Although the response of ecosystem processes to climate has long been of interest, recent work has led to a much more general understanding of temperature and moisture effects on biota and on the interactions of climate effects with internal ecosystem processes such as succession and the nitrogen cycle. 9 This work includes manipulative experimentsb in the lab and field that have led to an improved understanding of microclimatic effects on biological processes and the specific behavior of particular ecosystems, whereas comparative studies and data syntheses have led to better understanding of ecosystem to global patterns. Long-term flux observations have illuminated the effects of climate on carbon storage: measurements over the past 25 years in the Arctic have shown tundra systems shifting from being a sink to a source of CO2 as conditions became warmer and drier. 10 Recent advances in measurement techniques, especially the advent of eddy covariance techniques and their application in long-term studies, have produced unique data on climate effects on net ecosystem exchange (NEE).11 Eddy covariance time series in forests have provided direct observations of the effects of unusually warm, cold, and dry conditions on carbon exchange. 12
Because the temperature (T) responses of respiration (R) tend to be larger than those of photosynthesis (A) (T versus R is exponential, whereas T versus A
|
b |
Manipulative experiments are ones where some variable or variables are deliberately altered and the response of the system is observed (e.g., experiments with artificially elevated atmospheric CO2). |
is saturating), it has been hypothesized that global warming should lead to global net CO2 emissions from ecosystems. 13 Recent studies using USGCRP global datasets have tentatively confirmed this hypothesis for short timescales (<1 year) while suggesting more complex interactions on interannual timescales. 14 Modeling studies have probed this type of relationship quantitatively, suggesting a dependence on rates of mortality, major effects in soils, interactions with the nitrogen cycle, and interactions between physiology and biogeography.15 As observing techniques improve and time series lengthen, it has become increasingly possible to distinguish among alternative hypotheses about how future warming might affect ecosystems. An exciting development is the use of models to understand the emerging time series of climate and carbon exchange at the global scale. 16 Early efforts to model climate effects on ecosystems were extremely hypothetical.17 The models used then had been tested at a limited array of sites, against “typical” or average conditions. The ability of the models to simulate the dynamic response of ecosystems to varying climate had in general not been examined. Currently, models used to project future ecosystem responses are being tested against observed dynamic changes resulting from inter-annual climate variations using data collected at local to global scales.18
Land Surface Processes and Climate
Research on the role of the land surface in climate has been a prominent area of research at the interface of ecology and atmospheric science over the past decade. It has led to dramatic developments in science and in observing systems. Beginning with a few provocative papers on the potential effects of land cover and inhomogeneities in land cover on climate, research on land surface processes has expanded to encompass a large and diverse theoretical and modeling effort validated by a series of highly successful international field campaigns. 19 The field campaigns have in turn led to a number of payoffs.
The use of eddy covariance flux measurements in ecological applications (in which measurement of the vertical winds and concentrations of a gas are measured together) was pioneered and verified in the First ISLSCP (International Satellite Land Surface Climatology Project) Field Experiment (FIFE),20 and the datasets collected during FIFE remain a touchstone for validating land surface models. A key scaling principle for canopies was first combined with land surface models as a result of FIFE collaboration, greatly increasing the simplicity and success of canopy modeling (see Figure 2.2). 21 The validity of “vegetation index”-based estimates of surface conductance was tested against observations during FIFE, and this validation remains a cornerstone of the communities ' confidence in satellite-driven land surface models. Results are just appearing from the Boreal Ecosystem-Atmosphere Study (BOREAS) campaign, which extended the FIFE paradigm to forested ecosystems. These results suggest a strong role for vegetation-atmosphere interactions in northern climates,22 as do results from the
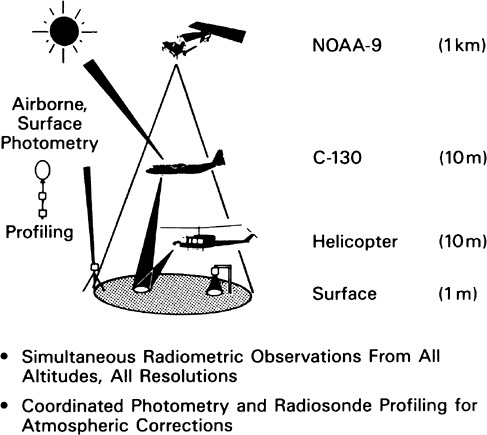
FIGURE 2.2 Range of spatial scales addressed by the First ISLSCP Field Experiment (FIFE). SOURCE: Sellers et al. (1992). Courtesy of the American Geophysical Union.
NSF Arctic System Science program's Land-Atmosphere-Ice Interaction Study, and the importance of disturbance processes (such as large-scale fires) in land surface water and carbon exchange. They are also likely to make major contributions to understanding isotopic exchanges between ecosystems and the atmosphere. 23 Completion of the final planned experiment, the Brazilian-led Large-Scale Biosphere Experiment in the Amazon, will lead to a broad understanding of ecosystem-climate interactions in boreal, temperate, and tropical ecosystems. This work has had, and will continue to have, a major impact on our understanding of paleoclimate, contemporary weather and climate, weather forecasting, and climate projections.24
The need for comprehensive information on the land surface has also spawned a large effort to develop remote sensing algorithms for land surface variables. Major progress has been made in developing satellite algorithms to infer surface resistance to evaporation, temperature, soil moisture, and land cover.25 Experience
in using satellite data was gained using extant satellite systems such as the Thematic Mapper (TM), the Systeme Probatoire pour l' Observatión de la Terre (SPOT), and the National Oceanic and Atmospheric Administration (NOAA) Advanced Very High Resolution Radiometer (AVHRR) systems. These data were successfully combined with algorithm development and evaluation in field campaigns such as FIFE, BOREAS, and the Hydrological and Atmospheric Pilot Experiment.26 The intensive, decade-long effort in this area resulted in the experimental design of two major EOS satellites (AM-1 and PM-1) being optimized for synergistic measurements of land surface variables. The primary platform, AM-1, orbits with a morning overpass time, chosen to minimize cloud contamination for land surface imaging, and includes a synergistic combination of three instruments: the Moderate-Resolution Imaging Spectroradiometer (MODIS), the Multi-Angle Imaging Spectroradiometer (MISR), and the Advanced Spaceborne Thermal Emission and Reflection Radiometer (ASTER). MODIS has capability in the visible and near infrared for remote sensing of vegetation characteristics with high time resolution. MISR takes multiangle measurements that allow determination of albedo and better constrain so-called vegetation indices related to conductance to water and photosynthesis. 27 ASTER and LANDSAT provide high spatial resolution information on land cover. The proposed remote sensing strategy for the physical climate system and carbon cycle studies is shown in Figure 2.3.
Research on land surface processes exemplifies a constructive partnership among many groups: global climate modelers, organism- to ecosystem-oriented bio- and microclimatologists, ecologists, and remote sensing scientists, as well as geographers, plant physiologists, soil scientists, and hydrologists. Having defined the need for improved satellite algorithms early on, the community carried out the necessary theoretical, modeling, and empirical demonstrations of such a capability. These science requirements are now largely executed on the EOS AM-1 spacecraft. AM-1 is a large and expensive mission, but the community has confidence in the quality of its land surface mission. This forms one component of AM-1's full scientific agenda, which also includes cloud, aerosol, atmospheric chemistry, and oceanographic experiments.
Human Use and Modification of Ecosystems
At the beginning of carbon cycle research, the carbon cycle appeared to be roughly in balance, with fossil emissions balanced approximately by ocean uptake and atmospheric accumulation. Beginning in the 1970s, ecologists led by George Woodwell began to make the case that emissions from land use, largely deforestation, had to contribute significant inputs to the atmosphere. As this hypothesis became better and better documented, it became clear that to balance land-use emissions an additional sink process, dubbed the “missing sink,” was required. The significance of land use in the carbon cycle was recognized prior to the USGCRP and the IGBP and has been a major focus of both programs since their
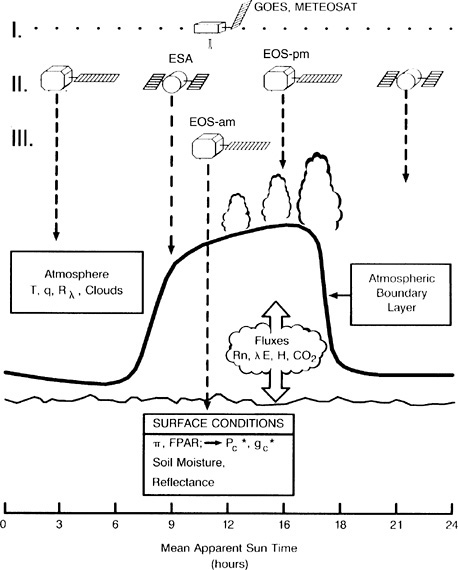
FIGURE 2.3 Satellite remote land-atmosphere interactions. Schematic of the diurnal variation of atmospheric boundary layer height and cloudiness for a humid continental region during the growing season; surface fluxes of net radiation sensible heat flux, water vapor, and CO2 are depicted. The proposed remote sensing strategy is shown for the physical climate system and carbon cycle studies. I. Geosynchronous observations of cloud fields, reflectances, temperatures. (GOES, METEOSAT). II. Two polar platforms with sounding instruments and imagers capable of resolving cloud fields. Platforms are spaced in time to characterize diurnal variations of atmospheric variables (POEM-1, EOS-pm). III. Polar platform with surface imaging payload. A morning crossing time is preferred to minimize cloud contamination (EOS-am). SOURCE: Sellers and Schimel (1993). Courtesy of Elsevier Science-NL.
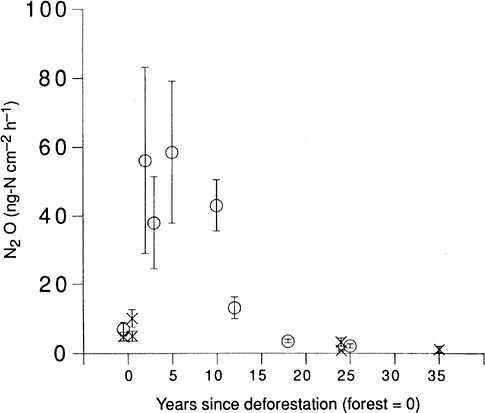
FIGURE 2.4 Average soil emissions (± 1 standard error) of N 2O plotted against pasture age (time since deforestation) for La Selva (C) and Guácimo (O). Forest sites are shown at 0 year. Significant differences were found among site means (analysis of variance, p < .01). SOURCE: Keller et al. (1993). Courtesy of Macmillan Magazines Ltd.
inception, as well as later in the International Human Dimensions Programme for Global Environmental Change. Whereas early attention was given to the carbon cycle, interest increased about the role of land use in altering trace gas budgets through soil processes and biomass burning28 (see Figure 2.4). Indeed, it has long been clear that humans have exerted predominant control over many aspects of ecosystems globally. 29 The fragmentation of ecosystems owing to changing land use, with attendant effects on biotic and biogeochemical function, is another increasing concern central to the concern over the loss of biodiversity.
Research on land-use change has profited enormously from the availability of archived satellite data and improved techniques for satellite data analysis. 30 Early estimates of land-use change were based almost entirely on national statistics and inventory information. 31 Contemporary estimates are increasingly based on satellite information, 32 and techniques for continuing satellite monitoring into
the future are improving. Commercialization of the TM and SPOT satellites and the accompanying expense of acquiring appropriate data over large areas set this effort back, perhaps by a decade. Today, computer technology is available to process large amounts of high-resolution data, but the satellite data remain costly. Retrieval of improved and spatially resolved historical land-use information is needed for retrospective analyses of the carbon cycle and to initialize models for global projections. Global retrieval remains a major problem, which is being addressed, albeit in a piecemeal and somewhat halting fashion at the present. However, understanding contemporary vegetation patterns, patterns of human land use, and the interactions of human and natural ecosystem processes is increasingly emerging as an important research area, and a vigorous community is developing.33 This area of science has a strong natural link to biodiversity research since climate impacts on biodiversity will occur synergistically with other environmental changes.
Biotic Interactions and Global Change
Until recently, the term “global change” generally implied biophysical or biogeochemical change—to the extent that it implied biology at all. Changes in the biota were treated as resulting from changes in climate, CO2, or land use. Climate and chemical changes have the potential to have significant impacts on biodiversity.34 However, it has become evident that biotic changes in and of themselves can represent regionally and globally significant environmental changes. Perhaps the clearest example of such biotic change is biological invasion—the human-mediated transport of plants, animals, and microbes across Earth. The mobility of people and goods has increased the rate of movement of other species by orders of magnitude—and introduced species are now abundant over much of Earth, in coastal marine as well as terrestrial and freshwater ecosystems. 35 The result is homogenization of Earth's biota, with ubiquitous weedy species proliferating at the expense of local species.36 More specific changes include many well-documented invasions that affect the health of humans and other species (most infectious diseases are invaders over most of their ranges). For example, introduced insect pests and microbial pathogens have done more to alter American forests37 than climate change, elevated CO2, and air pollution together—a statement that can safely be extrapolated for decades into the future. But biotic changes may interact with global change to alter biodiversity in hard-to-predict ways. Invading species also can affect the biogeochemical and biophysical properties of the invaded areas. Perhaps the best example is the widespread invasion of fire-tolerant African and Eurasian grasses into North and South America, Africa, Australia, and Oceania.38 Where they invade, these grasses can increase fire frequency and thus affect regional biogeochemistry, simplify vegetation structure and alter biophysical properties of ecosystems, and interact strongly with other components of global change.
Invasion is not the only example of biotic change—human-caused removal of large predators, and sometimes even grazers, is widespread, and it profoundly alters the functioning of aquatic and terrestrial ecosystems.39 Human actions also drive the extinction of species and genetically distinct populations. This is itself an irreversible change, and experimental studies are under way to assess linkages between functional diversity, the characteristics of organisms, and the functional responses of ecosystems to stress.40
System-Level and Long-Term Manipulations
Ecosystem responses to global change forcings include components that interact at a range of temporal and spatial scales, as well as subsequent feedback with the potential to amplify or suppress initial responses. This combination of numerous interactions, multiscale responses, and positive and negative feedbacks is a strong argument in favor of experimental studies on a scale that encompasses a much larger range of significant processes and interactions. Although recent progress in understanding feedbacks at the global to regional scale is still model-centered,41 an increasing number of experimental studies complement modeling studies on whole-ecosystem responses to altered atmospheric CO2, temperature, and water and nutrient inputs. Some of these studies have run or are scheduled to run long enough to provide information or potentially critical feedbacks that develop only as the forcing shapes biogeochemical and ecological phenomena over decades or longer.
These long-term ecosystem-scale experiments have already shown that the intensity of feedbacks and indirect responses can be surprisingly significant at a number of scales. 42 In Arctic tundra, altered nutrition under increased CO2 led to homeostasis of net carbon exchange after only three years. Yet a 4°C temperature increase resulted in persistent carbon storage under elevated CO2. 43 CO2 enrichment under the more fertile conditions of the Chesapeake Bay wetlands leads to continued stimulation of photosynthesis and plant growth, especially below-ground growth, for at least nine years. 44 The contrast between the Arctic and saltmarsh studies shown in these examples suggests a key regulatory role for nutrient limitation; however, shorter-term experiments indicate no consistent modulation of CO2 responses by nutrient availability. 45
Soil warming could drive increased decomposition and mineralization, but it could also lead to increased drought and consequent decreases in primary production, decomposition, and mineralization. With nine years of summertime warming, Arctic tundra primary production changed only slightly, though the increased abundance of deciduous shrubs presages decreased decomposition and future declines in nutrient availability. 46 Warming in montane meadows also stimulated dramatic changes in plant species composition, with increased dominance by sagebrush, indicating likely future changes in carbon storage, nutrient cycling, and biodiversity. 47 Long-term nutrient additions can also stimulate dramatic
changes in plant species composition. In Minnesota old fields, nitrogen additions simulating moderate to heavy atmospheric deposition led to the replacement of C4 by C3 grasses, with important decreases in carbon storage per unit of nutrients as well as changes in seasonality.48
Manipulative experiments have been important in marine as well as terrestrial systems. 49 In some oceanic ecosystems, supplies of major nutrients such as nitrogen and phosphorus persist throughout the growing season. In the late 1980s it was hypothesized that these regions might be iron limited.50 Additions of iron in the equatorial Pacific, together with a tracer allowing the fertilized region to be tracked, increased phytoplankton and microbial activity, affecting both community composition and carbon cycling.51 Experiments of this type, following both natural (e.g., atmospheric deposition events) and artificial fertilization experiments, with major as well as minor nutrients, have long been contemplated as prospects for ocean perturbation studies that can be followed with satellite ocean color data.
Multiple Stresses on Ecosystems
The dominant concern originally motivating global change research was global climate change. As ecologists and their colleagues from other disciplines began to address ecology at regional and larger scales, the importance of multiple large-scale environmental changes became apparent. Changes to climate, air quality, and land use may have synergistic effects, modifying the vulnerability of ecosystems to change. The concept of the metro-agro-plex, a region characterized by dense human populations, urbanization, and intensive agriculture, evolved in this context.52 Examination of the geographies of regional air quality problems, urbanization, population growth, and intensive agriculture indicates that all are taking place in the same regions—for example, in Southeast Asia, the eastern and western seaboards of the United States, and Western Europe. These areas tend to have high air pollution levels, with elevated ozone, carbon monoxide, sulfur, and nitrogen. Urbanization and industrialization lead to air pollution and its precursor inputs, but the proximity of cities and agriculture makes the agricultural system vulnerable to damage from ozone, acidification, and other stresses. However, intensive agriculture is itself a source of trace gases to the atmosphere (NO, NH3, and particulates) that may interact with regional air chemistry, particularly at the margins of the metro-agro-plex. Although air pollution is often thought of as an acute local or regional problem, metro-agro-plexes cover sufficient area and contain a high enough proportion of the world's population that they are properly thought of as a global problem.
Air pollution also has a direct global component. Recent work using observations and modeling has shown that nitrogen pollution has become a global problem and that likely future changes in fossil fuel and fertilizer use will lead to high nitrogen deposition worldwide. 53 Beginning in the 1980s, 54 anthropogenic
nitrogen deposition has been considered to be a potential regulator of the carbon cycle. In the 1990s a number of careful studies 55 evaluated the potential effects of different forms of nitrogen deposition and concluded that global changes to the nitrogen cycle may have a serious impact on the global carbon cycle (see Plate 1). This analysis includes both deleterious effects in the core pollution areas and modest fertilizer effects in the margins of polluted airsheds. 56 It has also been shown, and is likely from first principles, that high N deposition is correlated with high ozone levels (because of the photochemical coupling of NO and O3), leading to a potential multiple-stress situation combining the effects of N loading and oxidant stress.57 This is of particular concern as these stresses, especially in deposition, are also implicated in changes to biodiversity.
Early experiments tend to support a “subsidy-stress” paradigm for N, in which N deposition can lead to increased growth up to a critical level, beyond which deleterious effects dominate. 58 The effects of previous land use may also play a major role in the vulnerability of systems to N stress, with the corresponding prior history of N budget changes and the physiology of current vegetation changing the quantitative relationships between N loading and ecosystem impacts.
CASE STUDIES
The Carbon Cycle
Ecosystem science has played a major role in studies of the carbon cycle. Although there remain substantial uncertainties about the carbon cycle and how it may behave in the future, significant advances have been made. Progress in this area is critical because carbon cycle research forms the basis for setting targets in international negotiations to mitigate climate change.59 Understanding contemporary and possible future fluxes of carbon is the essential underpinning of sound policy to manage radiative forcing of the atmosphere. The development of accurate and reliable measurement techniques for carbon fluxes is a prerequisite for evaluating the success of measures undertaken to comply with the Framework Convention on Climate Change and to monitor international compliance with other treaty measures. Carbon cycle science is thus essential to international decision making.
By the late 1980s the budget of carbon dioxide in the atmosphere was obviously unbalanced (see Table 2.1). Carbon modeling suggested that fossil fuel and cement manufacture sources had essentially been balanced by ocean uptake and atmospheric accumulation.60 However, beginning in the 1970s, ecologists increasingly found that substantial emissions were being made to the atmosphere from land-use change (in the 1980s these sources were largely tropical). The land-use contribution was based on estimates from carbon budgets for land conversion and area converted. Thus, even the definition of the complete set of terms in the carbon cycle (industrial, marine, atmospheric, and terrestrial) required
TABLE 2.1 Average Annual Budget of CO2 Perturbations for 1980 to 1989
reconciliation of oceanography, ecology, and geochemistry. The budgetary imbalance of 1 to 2 gigatons (Gt), or the inability of models to account for the fate of the excess carbon, is referred to as the “missing sink,” and the excess carbon was assumed to be in terrestrial ecosystems because models of ocean uptake of CO2 indicated actual uptake of 2 to 2.5 Gt, based on calibration against isotopic 14CO2 (from thermonuclear bomb testing), 61 and fossil fuel emissions and atmospheric accumulation were known. This information set the stage for remarkable progress.
First, the missing sink, though widely assumed to arise from CO2 fertilization of terrestrial ecosystems, had never been observed in any sense. The quantity was deduced by mass-balance difference, and the assignment of the cause to CO2 fertilization was based on laboratory experiments and circumstantial evidence. Few suitable datasets existed to identify what was assumed to be a ubiquitous but spatially diffuse carbon accumulation in terrestrial ecosystems. In the late 1980s several groups began to estimate spatial patterns of CO2 sources and sinks using inverse modeling (see Figure 2.5). Papers authored by atmospheric scientists, oceanographers, geochemists, and ecologists have explored this methodology and effectively established the existence of a CO2 sink in northern hemisphere terrestrial ecosystems. The initial publications describing the northern hemisphere sink galvanized ecologists who had previously found the missing sink problem poorly posed for empirical research. Thus, the initial inverse work led to the initiation, modification, and reexamination of many studies.62
Work on the biological mechanisms underlying a terrestrial sink has also shown a remarkable and growing interdisciplinary character. The effect of CO2 on photosynthesis, long known from laboratory studies, has been proposed as an explanation for additional ecological uptake of CO2. Early work was largely
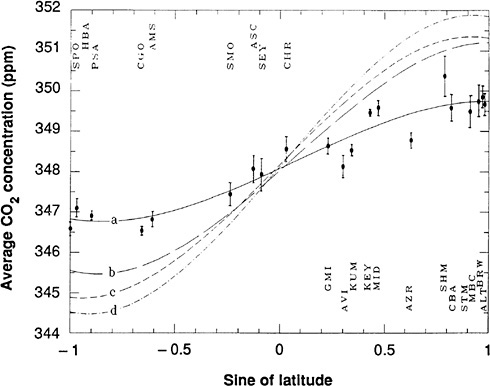
FIGURE 2.5 Observed atmospheric CO2 concentrations at the sites of the NOAA/ GMCC flask network. The error bars represent 1 SD of the annual averages at each site after adjustment to 1987. Curve (a) is a least-squares cubic polynomial fit to the data. The concentration distributions at the NOAA/GMCC sites have also been calculated with the NASA/GISS/General Circulation Model transport fields. Other curves are polynomial fits to the calculated CO2 distributions (not shown) with fossil fuel emissions, seasonal vegetation (no net annual source or sink), tropical deforestation of 0.3 Gt of C per year, and three different cases of ocean uptake: (b) the compilation of CO2 uptake based on the −Ψ CO2 data and empirical transfer coefficients; (c) CO2 uptake based on the same −Ψ CO2 map but calculated with the Liss-Merivat relation for air-sea exchange; (d) an earlier estimate of ocean uptake totaling 2.6 Gt of C per year. SOURCE: Tans et al. (1990). Courtesy of the American Association for the Advancement of Science.
conducted by physiologically oriented ecologists and agronomists and focused on the effects of CO2 on photosynthesis, secondarily on plant growth, and additionally on ecosystem consequences of enhanced CO2 uptake. Although there was widespread skepticism that terrestrial productivity was limited by the availability of carbon—many ecologists believed carbon accumulation to be primarily nutrient limited—most early research focused on the initial physiological effects of increased CO2.
In the mid-1990s the notion that the physiological effects of increasing CO2 might be modulated by nitrogen availability became widely accepted. 63 The argument is that as organic matter accumulates in response to increasing CO2 the associated nitrogen is sequestered from the actively cycling pool that supports plant growth.64 As CO2 fertilization progresses, the ability of plants and soils to store more carbon should become increasingly limited by nitrogen. Although this is difficult to test experimentally (because the effects of decades of environmental change on ecosystem nitrogen budgets is not understood or easily mimicked experimentally), its implications have been examined in a series of modeling exercises whose results have been compared to variations in the spatial and temporal patterns of observed CO2. 65 Once again, these studies have included atmospheric transport modelers, geochemists, ecologists, and, increasingly, atmospheric chemists. This collection of interdisciplinary work has shown, first, that it is difficult to account for more than a fraction of the missing sink (30 to 70 percent) based on CO2 fertilization alone, and, second, that changes in the seasonal cycle of CO2 over the past decades are not consistent with CO2 fertilization being the sole mechanism for the missing sink.66
With the new awareness that nutrients may control carbon cycling, several workers have shown that increasing anthropogenic nitrogen pollution could cause a substantial terrestrial sink of CO2. The magnitude of this sink would depend on the spatial distribution of pollution, the fraction of areas where N pollution is serious enough to reduce productivity, and the fate of N in ecosystems. 67 The N-driven sink could account for some of the spatial and temporal differences between estimated terrestrial uptake and modeled CO2 effects.
The past decade of carbon cycle research has yielded a panorama of extraordinary scientific accomplishment, and yet daunting challenges remain. Identifying source and sink regions, and the controls on these systems, is an extraordinarily difficult scientific problem. However, it is also a pivotal problem and must be attacked. This attack may require a more intensive and coordinated battle plan than the highly successful but ad hoc efforts of the past 10 years.
Modeling Ecosystems Across Multiple Levels of Biological Organization
Early models of terrestrial biogeochemistry lumped terrestrial biota into a small number of compartments, defined as living and dead and characterized by turnover times. Even major biomes were not always distinguished. The effects of changes in vegetation distribution (amply documented in the paleorecord) were not considered. The ways that ecosystem models deal with biota have changed substantially in the past decade. It is now widely recognized that functional differences between vegetation types influence ecosystem processes. It is also recognized that different vegetation types are used differently by humans. These differences to humans are not adequately indexed by net primary productivity (NPP); for example, many western U.S. rangelands have NPPs similar to
montane forests, yet these systems are hardly interchangeable. Contemporary models incorporate a substantial amount of information on species or growth form attributes and simulate behavior that differs in the face of similar stresses depending on biotic and edaphic properties. 68 Although knowledge about the linkage of functional (carbon, water, and nutrient dynamics) to population-community processes is in its infancy at global scales, it is widely viewed as the next major challenge. One influential, if highly simplified paper, suggested that future changes to global ecosystem carbon storage will be dominated by the population processes of mortality, recruitment, and rates of migration or recovery.69
The importance of considering processes across biological levels of organization was highlighted by a recent large-scale international collaboration, the Vegetation and Ecosystem Modeling and Analysis Project.70 VEMAP compared models of biogeochemistry and biogeography for the conterminous United States under current and general circulation model (GCM)-simulated future climates (current climate is defined operationally as the mean over the past few decades). The project covered three models of biogeochemistry and three models of biogeography (vegetation type distribution), developed by leading groups from the United States and Europe. The project was structured as a sensitivity analysis, using factorial combinations of climate (from three GCMs) and direct CO2 effects (simulations were done at 350 and 700 parts per million by volume CO2). Simulations of biogeochemistry (NPP, carbon storage, N turnover) with climate and CO2 change were also conducted, using potential natural vegetation and vegetation distributions from the three biogeography models. VEMAP was motivated by the desire of several ecologists and funders to have state-of-the-art ecological modeling results for input to the 1995 Intergovernmental Panel on Climate Change (IPCC) report. VEMAP 's initial phases were conducted on an accelerated schedule to permit publication of results prior to the IPCC's deadlines.
While there have been several intercomparison studies of ecosystem models, 71 VEMAP was unique in linking biogeochemical and biogeographic processes. VEMAP also contributed greatly to developing consistent input databases and modeling protocols, allowing for both validation of models and their rigorous comparisons. The VEMAP data group provided a “bioclimatology ” of the domain, which included adjustment of the data to account for topographic effects of elevation and rain shadows and the complete suite of variables needed as input for biological models (temperature, rainfall, wind speed, radiation, and humidity). The suite of climate variables was interpolated to preserve physical consistency among variables (e.g., both sunny and rainy days are represented) and was provided at consistent monthly and daily time steps. The climate data were coregistered to a map of potential natural vegetation aggregated into classes consistent with all six models and to soil property information. The GCM climate scenarios were then provided as changes from the base climatology. The database thus provided a “level playing field” that allowed intermodel differences to emerge clearly. In most other intercomparison studies, differences in the
input data used for different models has clouded interpretation of model-to-model differences.
The first VEMAP publications described the study and presented the key results on climate change impacts as input to the IPCC.72 The authors reported that CO2 increases alone (with no climate change) caused modest effects on NPP and carbon storage, with potential leaf-level effects of 30 to 40 percent on NPP in all models scaled back to less than 10 percent by ecosystem-level feedbacks via nitrogen cycling and water availability (see Table 2.2). Modeled responses to climate change and changing CO2 together were usually larger but more variable than to CO2 alone. The responses were amplified because warmer conditions accelerated the model's N cycles, reducing the N limitation of carbon accumulation that occurred under the simulations of doubled CO2 alone. When biotic change was included by linking the projections of vegetation redistribution to the biogeochemical models, the effects grew larger yet and more variable, with some models showing effects of opposite sign, depending on the combination of climate scenario, vegetation scenario, and models used. While the models agreed on current-day stocks and fluxes, their responses to environmental change were variable. As additional environmental perturbations were added to the study, the between-model differences in carbon cycling increased. In the first experiment, responses to CO2 increase alone were modeled; then CO2 plus climate changes were examined. Finally, the effects of CO2 and climate on vegetation distributions and the effects of all three factors (CO2, climate, and biogeography) on carbon stocks and fluxes were evaluated. As these additional factors were added to the experiment, the differences in predictions between models increased. Despite this not unexpected result, all of the biogeochemical models showed that the nitrogen cycle influenced the sensitivity of the carbon cycle to CO2 and climate. 73
The paradox of VEMAP—that the models agreed on NPP and carbon storage under current conditions but simulated divergent behavior under altered conditions—led to a detailed analysis of model mechanisms.74 The analysis showed that, while the models agreed on continental average NPP and carbon stocks (see Table 2.2), they disagreed on spatial patterns within vegetation types and, more fundamentally, predicted different relationships of NPP and decomposition to nutrient and biophysical controls. 75 The differing spatial sensitivity of NPP to water versus nutrient regulation was a clear predictor of response to changes in the modeled climate conditions. Models that had high sensitivity to evapotranspiration (and hence direct climate effects) relative to nutrient limitation behaved differently under altered climate conditions than models with the opposite sensitivity. The validation studies conducted so far provide clear information on why models differ and on the data and experiments needed to establish the correct mechanism.76 Efforts to test the VEMAP models are being coordinated with U.S. and IGBP networks of CO2 fertilization experiments and canopy flux studies.
The VEMAP intercomparison required the modeling groups to agree on a common age for forest stands to avoid having differences in stand age confound
Table 2.2 Results from VEMAP: Biogeochemical Consequences of Changing CO2, Climate, and Vegetation Using Three Biogeochemical Models (Biome-BGC, Century, and TEM), shown with Vegetation Redistribution Simulated Using Three Biogeography Models (MAPSS, DOLY, and BIOME 2). SOURCE: Data from VEMAP Participants (1995)
|
NPP* (Tg C) |
Carbon storage (Gt C) |
||||||
|
Biome-BGC |
Century |
TEM |
Biome-BGC |
Century |
TEM |
||
|
Current climate and CO2 |
3,772 |
3,125 |
3,225 |
118 |
116 |
108 |
Models in general agreement |
|
Doubled CO2 (% change from control) |
+10.8 |
+5.0 |
+8.7 |
+6.5 |
+2.2 |
+8.5 |
Effects are downregulated relative to leaf level potentials |
|
Climate change and CO2 (3 scenarios; % change from control) |
+1.7 to +20.2 |
+14.6 to +23.6 |
+24.6 to +26.5 |
−11 to 27.7 |
+5.9 to +7.2 |
+12.3 to +16.1 |
Effects are larger than CO2 alone but more variable |
|
With vegetation redistribution (3 scenarios of vegetation change; % change from control) |
−0.7 to +21.1 |
+11.3 to +26.0 |
+27.0 to +39.7 |
−8.3 to 39.4 |
−1.8 to +20.4 |
0.0 to +32.3 |
Variability in sign and magnitude of effects increases; effects of changing vegetation and physiology are roughly equal |
|
* Net primary productivity. |
|||||||
model sensitivities. For some of the models, simulating equilibrium conditions implies simulating mature stands, whereas in others standardization implies choosing a disturbance frequency and then presenting model results from a particular phase of the cycle. The consequences of the explicit or implicit handling of disturbance and demography were felt throughout the entire experiment. The assumptions about forest stand conditions affected estimated resource use, biomass turnover, and carbon stocks. The reduction in spatial heterogeneity associated with assuming a single stand age or disturbance regime for each cell within a biome was evident in model-data comparisons. The demographic, physiological, and plant community consequences of disturbance proved of first-order importance, even using the highly aggregated VEMAP-class models.77
Three key findings stand out:
-
First, the biogeochemical consequences of plausible scenarios of vegetation redistribution in response to large-scale environmental changes are as large as the direct effects of climate on biogeochemistry.
-
Second, there are large differences in resource use among species or functionally similar groups of species (“functional types”). Changes in ecosystem function per unit change in resources (water, energy, nutrients) that were caused directly or indirectly by global change may be quite discontinuous, as the biogeographic consequences of climate change interact with the biogeochemical changes. Thus, as species or functional types change in response to climate or other drivers of change, large changes in ecosystem function may occur because different plant species interact with the climate differently.
-
Third, the direct and demographic effects of disturbance and succession have large effects on ecosystem processes at the continental scale. These effects are evident even in the VEMAP models where the processes of disturbance and succession are largely implicit. Present and future disturbance regimes (natural and anthropogenic) will have major effects on future ecosystems and must be included as a critical next step in ecosystem modeling. Such modeling of disturbances is currently a principal focus of the VEMAP group.
The VEMAP study represents a first step in introducing multiple levels of biological organization into large-scale ecosystem modeling. Other efforts are in progress within the IGCP-GCTE program and in numerous investigators' programs. By illustrating quantitatively the sensitivity of ecosystem function to biogeography—functional diversity among plant types, disturbance, and demography—the study emphasizes the importance of linking population/community-level ecology to biogeochemistry, a critical development for the eventual synthesis of global change and biodiversity research. VEMAP provides a partial template for how such integration of information and synthesis of ideas may occur. Key components are the intense and funded interactions of participants
toward an ambitious but focused goal, support for development of the data infrastructure to meet the needs of diverse groups in a consistent fashion, and iteration of increasingly ambitious model experiments along with their rigorous comparison to data. Linking mechanistic models of population and community change to ecosystem function at large spatial scales is a more difficult endeavor but one that is within or just beyond the community's grasp. Finally, VEMAP provided valuable experience in modeling ecosystem responses across multiple levels of biological organization, even though the representations of population, successional, and demographic processes were empirical, simplistic, or implicit.
A RESEARCH AGENDA FOR THE NEXT DECADE
Introduction
The science agenda for ecology is in large part motivated by a series of questions about the effects of ecosystem change on the Earth system, the effects of climate on ecosystems, the consequences of direct human activities on ecosystems, and the feedbacks between climate forcing and ecosystems. Much of the early research on ecosystems and global change focused on the carbon cycle, biophysical effects of the land on the atmosphere, and biogenic sources and sinks of trace gases. Research in these areas must continue, but the questions for the next few years are more focused following a decade of progress. In addition, because questions continue to evolve as new discoveries are made, we attempt to characterize the process as moving from identifying a question to analyzing the resources, collaborations, and infrastructure required to address it. As ecology continues to evolve, techniques for combining individual creativity with highly coordinated resources and technology must likewise evolve.
Research Imperatives and Key Scientific Questions
The preceding discussion indicates there are four Research Imperatives that should guide ecosystem studies in the USGCRP for the coming decade:
-
Land surface and climate. Understand the relationships between land surface processes and weather prediction, and changing land cover and climate change.
-
Biogeochemistry. Understand the changing global biogeochemical cycles of carbon and nitrogen.
-
Multiple stresses. Understand the responses of ecosystems to multiple stresses.
-
Biodiversity. Understand the relationship between changing biological diversity and ecosystem function.
Land Surface and Climate Imperative
Ecosystem-atmosphere interactions that affect climate continue to be an important research topic. Land surface processes are key to understanding regional climate, regional climate change, and climate prediction.
-
How do land surface biophysical processes interact with regional climate and modify patterns of interannual climate variability?
-
How does including knowledge of the land surface state affect weather prediction and seasonal to interannual climate prediction?
-
How might changing patterns of land use affect the climate of the future?
-
How might large-scale atmosphere-ecosystem exchange of water and energy change in a high carbon dioxide world?
Biogeochemical Imperative
Research on biogeochemical cycles is fundamental to global change science. Research for the next decade must focus on the role of the terrestrial and marine biospheres in today's carbon cycle.
-
How is terrestrial carbon storage regulated by land use, changes to marine ecosystems, internal ecosystem processes, and climate, and how might this storage change in response to future environmental changes?
-
What are the consequences of the anthropogenically accelerated nitrogen cycle?
-
Can we quantify the interactive roles of increasing CO2, the changing nitrogen cycle, and land use in terms of present and future terrestrial carbon storage?
-
How will the role of marine ecosystems change with future changes to ocean circulation, temperature, and nutrient/toxic inputs?
Trace gas biogeochemistry contributes to changing tropospheric chemistry, affecting radiatively active greenhouse gases. As research has progressed, the role of changing land use in changing trace gas emissions has come more sharply into focus. Other disturbances such as fire, pest/pathogen outbreaks, and biotic changes due to extinctions or invasions that may accompany changing land use should also be considered.
-
What are the current budgets for the sources and sinks of biogenic greenhouse gases, especially methane and nitrous oxide?
-
How is current environmental change, including land-use change, fertilization, and atmospheric N deposition, affecting the sources and sinks for these gases?
-
How might the sources and sinks change in the future with changing land management, climate, and chemical inputs?
-
As global use of anthropogenic nitrogen increases, is there potential for nitrous oxide emission or methane consumption to change rapidly?
Early work on biogenic trace gases focused on radiatively active trace species. Measurements made during the USGCRP and by IGBP projects have demonstrated that biogenic gases from microbes, plants, and biomass burning influence atmospheric photochemistry and aerosol formation, thereby impacting the atmospheric cycles of ozone, the hydroxyl radical, and reactive nitrogen. These projects have thus led to questions regarding reactive biogenic compounds, including aerosol precursors.78
-
What changes are occurring to the atmosphere-ecosystem exchange of reactive trace gas species (nitric oxide, ammonia, nonmethane hydrocarbons, dimethyl sulfide)?
-
What biological and pyrogenic processes control these exchanges, and how might they change in the future?
-
What is the role of changing biogenic trace gas emissions in the changing photochemistry of the troposphere and stratosphere?
-
Aerosols have become a major issue in climate. What roles do sulfur and organic compounds from biogenic sources, dust from agriculture and other soil disturbances, and biomass burning play in global aerosol forcing?
Multiple Stress Imperative
Work during the past decade has shown that climate and CO2 interact and that changes to the global nitrogen cycle likely affect this interaction. In addition, the susceptibility of regions to climate and biogeochemical change is greatly modified by changes to land cover. Research on global climate change impacts at first emphasized the direct effects of CO2 on climate but has increasingly emphasized interactions of CO2 and climate with the nitrogen cycle as well as interactions with other stresses.79 Research on climate effects on ecosystems should be conducted with the realization that climate change is only one of a number of simultaneous impacts in many systems.
For regional scales the focus is changing from single-factor stresses (e.g., ozone) to consideration of multiple impacts. In areas of dense human populations, where industry, urbanization, and agriculture coexist (the “metro-agro-plexes”), the interactive effects of environmental changes to climate, atmospheric chemistry, sea level, water quality, and land use may dominate, producing consequences different from those expected based on single-factor studies or models. The study of multiple stresses (such as changing climate, air pollution, or water quality) must come to the fore, and large-scale experimental studies (which today focus on, e.g., CO2, soil warming, and ozone exposure) must address interactive effects.
-
How do multiple global changes interact to produce ecosystem responses?
-
What are the interactions of changing land use, climate, nutrient and toxic inputs, and hydrology on ecosystems and their ability to produce goods and services and to sustain biodiversity?
Systems subject to multiple stresses tend to be affected by a combination of large-scale factors (e.g., climate) and regionally unique factors (e.g., air or water pollution, invasive species). Regional studies can be important in studying ecosystems inasmuch as a higher level of physical, biological, and chemical detail can be obtained and modeled at this scale. 80 We must develop a methodology for understanding the implications of multiple environmental changes on interactive human-ecological-hydrological systems.
-
What are the required datasets, theories, and models needed to understand regional coupling of physical and chemical climate, land use, and ecosystems?
-
Can we develop the science needed to manage regional systems subject to multiple stresses to provide ecosystem goods and services while maintaining ecological integrity?
Early discussions of the role of ecosystems in global change focused on terrestrial and pelagic ecosystems. Since then it has become apparent that changes to terrestrial ecosystems caused by climate and land use can substantially alter transfers of material from terrestrial to freshwater systems and ultimately to coastal marine systems. These processes are only crudely quantified.81
-
How do changes to climate and land use affect the transfer of water and materials between terrestrial and freshwater ecosystems?
-
How does global environmental change affect the functioning of freshwater ecosystems?
-
How do changes to terrestrial and hydrological systems alter coastal marine systems?
-
How are coastal marine ecosystems changed by the interaction of climate, large-scale ocean circulation and biogeochemistry, and inputs from the land?
Biodiversity Imperative
As an ecosystem modeling framework has been developed for understanding biophysical and biogeochemical processes at regional to global scales, the interaction of ecosystem function with species ecology has emerged as a topic of increasing urgency. Although not all research on biological diversity lies within the global change research agenda, there is an important interface between the two areas. We must know the relationship of biological diversity to the func-
tional diversity of plants, animals, and microbes. In addition, we must begin to understand the impacts of global change on biodiversity. The vulnerability of biodiversity to climate, nitrogen, and other anthropogenic changes will be conditional on other stresses (e.g., land-use change, toxics). Research on the impacts of global change on biodiversity must be conducted as part of an integrated program, ideally linking global change research to national and international biodiversity efforts such as DIVERSITAS.c
-
How much functional redundancy exists in ecosystems?
-
How does the functional diversity of organisms in ecosystems affect carbon uptake and sequestration, nutrient cycling, biophysical interactions with climate, and trace gas emissions?
-
What information on plant, microbial, and animal function is needed to model the role of organisms in large-scale changes in community composition and ecosystem function?
-
How will climate changes interact with other anthropogenic impacts to alter biodiversity?
-
Are there critical (keystone) species governing large-scale ecosystem function, and can we identify what species could become keystone under changing environmental conditions?
-
Can we identify either systems vulnerable to change as a consequence of biological invasion or species likely to be successful invaders?
-
How might changes in pests and pathogens alter disturbance frequency, including land-use change?
Need for a Crosscutting Research Agenda
The discussion above frames the intellectual direction of global change research on ecosystems. However, experimental and modeling studies on ecosystems tend to be integrative, and many or most field studies will relate to many of the questions above. For example, the well-known long-term flux measurement program at the Harvard Forest, in Massachusetts, has provided insight into climate effects on ecosystem carbon storage and on the exchange of long- and shortlived trace species,82 as well as a site for developing and testing new remote
|
c |
Established in 1991, DIVERSITAS is jointly sponsored by the International Council for Science (ICSU), by several ICSU bodies (the International Union of Biological Sciences, the International Union of Microbiological Societies, the Scientific Committee on Problems of the Environment, and the Global Change-Terrestrial Ecosystems Project of the IGBP) and by the United Nations Educational, Scientific, and Cultural Organization. The goals are to provide accurate information and predictive models of the status of biodiversity and sustainable use of the Earth's biotic resources and to build a worldwide capacity for the science of biodiversity. |
sensing techniques. Soil warming studies have provided insights on carbon-cycle questions, trace gas questions (especially methane), the relative importance of physiological changes to species versus changes in species composition, and potential hydrological changes with global warming. Major field campaigns such as FIFE and BOREAS have provided information on a wide range of scientific problems and disciplines (from mesoscale meteorology to biodiversity). Because of this we describe the components of an ecological research agenda primarily by the type of research (the space and timescales addressed) or the level of ecological organization rather than by identifying research imperatives on an issue-by-issue-by-question basis. The elements of this research agenda (see Table 2.3) are described below.
Components of the Research Agenda
Observe Long-Term Trends and Interannual Variability in the Forcing and Responses of Ecosystems
Long-term measurements and experiments are a crucial component of ecological research, and, given the range of rates of ecosystem change (from seconds to millennia), many of these time series must span years to decades. As discussed later under “Lessons Learned”, observational time series and long-term experi-
TABLE 2.3 Key Measurements and Research Imperatives: Need for a Crosscutting Research Agenda
|
Ecosystems And Physical Climate |
Sources And Sinks Of Biogenic Gases |
Multiple Stresses And Ecosystem change |
Biodiversity And Changing Ecosystem Function |
Key Measurement |
|
X |
X |
X |
X |
Network of sites. |
|
X |
X |
X |
Vegetation index/ocean color. |
|
|
X |
Atmospheric composition.* |
|||
|
X |
X |
Ocean pCO2. |
||
|
X |
X |
X |
X |
Land cover.† |
|
X |
X |
X |
Toxic and nutrient deposition. |
|
|
X |
X |
X |
Large-scale manipulation. |
|
|
* Measurements of moderate to long-lived compounds from a global network (e.g., CO2, 13CO2, N2O, C18O 2, CH4, CO, O2). † May be required at a range of spatial resolutions depending on the question. |
||||
ments have yielded many fruitful insights into ecosystem behavior that are relevant to global change. These achievements were possible because long time series allow small but consistent responses to be observed against a background of considerable variability. 83 Time series observations also allow the direct observation of system responses to trends and major climatic events.84 It is imperative that the scope of time series observations and long-term experiments be strategically broadened. Critical areas for long-term studies include the following.
Experimental manipulations
While much of our understanding of long-term and global- to regional-scale feedbacks is based on models, long-term experimental studies are vital for discovering key interactions and for testing models. Studies of whole-ecosystem responses to altered atmospheric CO2, temperature, and water and nutrient inputs are central to understanding the mechanisms through which environmental changes affect ecosystem processes. Some system-level manipulations have run or are scheduled to run long enough to provide access to potentially dominant feedbacks that develop only as the influence of the forcing penetrates biogeochemical and ecological compartments with time constants of decades or longer. Studies of mature forest stands, of CO2 changes in concert with other changes, and of the effects of diversity remain high priorities. Experimental manipulations of disturbances (fire, insects, and human land use) are also potentially worthy of study. Equally important is the meta-experimental design, the choice of systems in which to conduct system-level manipulations, and the integration of responses across diverse types of ecosystems, as opposed to the specific manipulations performed at a site (the conventional experimental design).
Direct forcing of ecosystem change by human activities
Changes to climate and atmospheric composition are global. The ubiquitous nature of human resource use makes human impacts on ecosystems a globally distributed problem as well, although the specific impacts vary from region to region. Data on human impacts are often collected as a snapshot, but data on land use, pollution, and resource use should be available as time series. Many human impacts are cumulative, and analyses of land-use effects on terrestrial carbon storage show the need for time series data on human impacts. 85 In any program evaluating global changes to ecosystems, direct human impacts must play a central role. 86 Human use of ecosystems also alters their susceptibility to modification by climate and other “natural” processes.87 Understanding ecosystem interactions with the physical environment requires knowing the type and state of ecosystems present, making knowledge of changing community composition and
management imperative. The global distribution of human impacts over time must be known. Key datasets include the following:
-
Land cover. The type and characteristics of global managed and natural vegetation must be known for the present, for the past (as best as can be determined), and from now forward (notably through monitoring, using high-resolution remote sensing). An historical record from the beginning of the industrial era is needed to assess the effects of land-use change on climate and the carbon cycle.
-
Land use. The characteristics of land management, including agricultural practices, forest management practices, irrigation, urbanization, and other direct human manipulations of ecosystems need to be known (historically, to the extent possible) and monitored. This information is important for modeling ecosystems, trace gas emissions, and water quality.
-
Fisheries' practices. It is becoming increasingly evident that fishing dramatically affects marine ecosystems and may alter their interactions with the marine physical environment. Marine ecosystem management practices must be documented as part of global marine ecosystem studies.
-
Atmospheric deposition. Atmospheric deposition has recently emerged as a significant issue in global change research and for over a decade has been viewed as an important ecosystem stress at regional scales. Deposition of important anthropogenic chemical species, especially sulfur species, nitrogen species, and ozone, must be documented.
The above types of data, describing the impacts of humans on ecosystems in a geographic framework, form a central part of the information needed for large-scale investigations in marine and terrestrial ecosystems. They complement other datasets, such as those on climate, soil properties, ocean circulation patterns, and natural vegetation-type distributions that are already widely used in modeling.88 Retrospective and contemporary data on human alterations and use of ecosystems and on collateral impacts are crucial not only for modeling the ecosystems affected but also because these data provide a critical link between ecological and human dimensions research (see Chapter 7).
Observations of atmospheric change
Time series observations of atmospheric CO2 have long been a mainstay of global change research, and, as that time series has lengthened, the degree of insight it provides into the dynamics of the global carbon cycle has continued to increase. Time series measurements of other gases such as methane, nitrous oxide, and carbon monoxide and their isotopic composition are also crucial and have provided insights into their biogeochemistry.89
The current system of international observations emphasizes “background” atmospheric conditions, leading to better coverage of the relatively more uniform marine environments than their terrestrial counterparts. Additional continental stations are required both to improve the analyses' ability to resolve terrestrial influences on atmospheric CO2, CH 4, and other species with more precision and to allow better quantification of regional patterns of trace gas exchange. Whereas background stations can be located at sea level, the high variability of short to moderately long-lived gases (e.g., CO2, CH4, CO) in the lower troposphere over land makes surface observations difficult to interpret. The difficulty arises because trends, seasonality, and biological processes are confounded with the strong diurnal cycle and wind-direction-dependent influence of air pollution near the surface. Ideally, continental measurements should be made on aircraft or from very tall towers; in fact, transmission towers of 400 m or higher are now being used opportunistically90 (see Figure 2.6). Continental measurements are best made as a gradient with height from a tower or aircraft. In addition to using the vertical sampling scheme, continental sites must be located to sample the range of ecosystems and human systems present. The experimental design of the full sampling approach is critical and must be carefully analyzed before an investment in data gathering is made.
FIGURE 2.6 Vertical profile of CO2 difference from 496-m height) predicted for the convective boundary layer. The points show medians and the horizontal bars show the range of the lower and upper quartiles of the afternoon (1500 to 1700 EST) observations taken during June 1993. SOURCE: Bakwin et al. (1995). Courtesy of Munksgaard International Publishers.
Interannual variability and trends in atmosphere-ecosystem exchange
The new technologies of flux measurement by eddy covariance and relaxed eddy accumulation have made it possible to directly measure fluxes of water, CO2, and many other species (e.g., ozone, nitric oxide, isoprene) above plant canopies. Simultaneous measurements of other properties, such as canopy nitrogen, link flux measurements to other ecosystem properties. It is imperative that a carefully designed network of long-term flux measurement sites be implemented.
On a larger scale, satellite measurements of vegetation activity have increasingly proven their utility for observing seasonal and interannual variations in vegetation activity. 91 Global observations of vegetation state are important to understanding climate-land surface interactions, biogeochemical cycles, and hydrology. 92 Although direct observations of canopy gas exchange provide information on ecosystem interactions with climate at local scales, remote observations of vegetation provide a spatial perspective. As the next generation of space-borne sensors and algorithms become available, with improved calibration and atmospheric correction, the detection of variability and secular trends will become more quantitative. Improved algorithms based on radiative transfer theory will allow retrievals of actual vegetation characteristics, as compared to the correlative approaches used today. Ground truth and calibration data for new algorithms must be collected in concert with site-based process studies (see below).
Measurements of ocean color (see Plate 2) as a proxy for phytoplankton biomass and the level of carbon dioxide in solution (partial pressure of CO2 or pCO2) provide information on marine biological activity and the potential for CO2 exchange. Color and pCO2 must be measured to determine their typical seasonal cycle and their responses to interannual climate variability. In situ calibration and validation data are required to fully exploit these data.
Terrestrial and marine ecosystem changes associated with global change
To interpret changes in atmospheric concentrations, fluxes, and surface energy budgets, the dynamics of vegetation and microbial communities must be known. Site-based multivariate measurement sets are required to understand these ecological processes. Plant and phytoplankton productivity (NPP) is a fundamental ecological variable that must be quantified to address a wide range of questions. In terrestrial systems the “allocation” of plant productivity among different plant parts is of fundamental importance. Although theory and experimental data suggest that allocation should respond to climate and other environmental changes, remarkably few observations exist to field validate the predictions of computer or laboratory models. Similar questions arise for marine systems regarding the distribution of NPP among phytoplankton species and trophic levels. Measurements of litter fall arising from leaf, wood, and root
turnover, and of plant mortality are critical for understanding respiration, decomposition, and nutrient cycling. Litter fall, litter chemistry, litter decomposition rates, soil respiration, and nutrient availability are key ecosystem measurements needed to interpret and attribute causation to observed changes in fluxes and storage. In marine systems the export of particulate and dissolved organic matter below the euphotic zone is a primary variable. Measurements of NPP, respiration, and allocation, coupled with net CO2 fluxes determined from eddy covariance, must all be components of an observing strategy for ecosystems under a changing global environment. Coupled measurements of controls over productivity, including climate variables, soil moisture, and nutrient availability, also will be of great value. In addition, as ecosystems change, their emissions of other trace gases may change. Long-term measurements of fluxes for N2O, CH 4, dimethyl sulfide, and other trace species at a network of sites are required to detect changes in trace gas sources and to test process models of trace gas exchange. These measurements are particularly crucial in areas affected by intensifying human impacts (direct land use and changes to nutrient loading).
Summary: Key long-term studies
-
Experimental manipulations of CO2 and other environmental factors.
-
Well-designed observations of atmospheric composition (CO2 and its isotopes, CH4, N2O, O2).
-
Measurement of ocean pCO2 (seasonal cycle and interannual changes).
-
Satellite measurements of global ecosystems.
-
Measurement of fluxes and ecosystem characteristics at a network of sites.
Regional Analyses, Extrapolation, and Observations at Multiple Scales
Both terrestrial and marine ecosystems possess spatial structure at scales from millimeter to global. Ecosystems also interact with the physical environment at a range of scales, from micrometeorological to large-scale circulation of the atmosphere and oceans. Because of the fine structure of ecosystems that is derived from the variability of organisms, patchiness in soil or water properties, and long time-scales of atmospheric processes, large-scale ecosystem studies cannot be applied in their entirety at the global scale.
Our understanding of ecosystem processes has been greatly increased by field projects that have explicitly sought to measure specific processes at multiple spatial scales. At the beginning of global change research on ecosystems, the community's ability to “scale up” or extrapolate process understanding of the organism or measurement plot to higher levels was rudimentary. Field projects organized to address this question, and the use of flux measurements and remote
sensing, led to enormous progress in our knowledge of terrestrial ecosystems at large scales.
Regional-scale studies have been crucial in understanding the role of atmosphere-biosphere exchanges in tropospheric chemistry. Field campaigns such as the Arctic Boundary Layer Expedition series,93 the Canadian Northern Wetlands Studies, and studies of biomass burning (South African Fire-Atmosphere Research Initiative and Transport and Chemistry near the Equator over the Atlantic) transformed our knowledge of tropospheric chemistry. Long-term regional monitoring for atmospheric deposition of acidic species and ozone is key to understanding the regional impacts of atmospheric change on ecosystems. Regional analyses should include both biophysical and biogeochemical interactions of ecosystems and the atmosphere. The biophysical and biogeochemical processes are naturally linked because biophysical processes are critical regulators of trace gas exchange and biogeochemical processes influence biophysical exchanges. Although site measurements of trace gas flux are useful for detecting change and testing models, regional measurements using atmospheric techniques are crucial for constructing budgets. Regional trace gas budgets are naturally estimated in multiscaled studies, such as the Large-Scale Biosphere-Atmosphere Project in Amazonia (see Figure 2.7), where the regional measurements may be tied to local process studies.
Remote sensing should be integral to multidisciplinary studies. Both terrestrial and oceanic remote sensing techniques provide high-resolution information, such as information on land cover (see Plate 3), relevant to regional processes. Integrated regional studies also provide an ideal venue for transferring knowledge about advanced remote sensing capabilities to new user communities, if the research is initiated by interdisciplinary teams.
Modeling at the regional scale is also essential to integrated regional studies. 94 As discussed earlier, it also provides an organizing framework and detailed information complementary to global models. Models provide an encapsulated, or planned, form of disciplinary knowledge in interdisciplinary investigations. The regional scale provides a venue where the critical processes in human-ecosystem-physical interactions may profitably be explored. Models designed for regional application can contain substantially more spatial and biological detail than global models (although this level of detail may only be available in certain regions).
Summary: Key regional measurements
-
High-resolution mapping of land cover (for terrestrial systems) or of physical and optical properties (for aquatic and marine systems).
-
Atmosphere-biosphere exchange of trace gases, toxics, and nutrients.
-
Field programs measuring ecosystem fluxes at organism to regional scales.
FIGURE 2.7 Map showing the research areas in the Large-Scale Biosphere-Atmosphere Experiment in Amazonia. Primary research areas are part of the two ecophysiological and land-use transects. Secondary research areas will be established over the entire Amazon basin. SOURCE: Courtesy of the Large-Scale Biosphere-Atmosphere Project Office, Brazil.
Measurements, Experiments, and Models of Ecological Organization at Genotype to Community Levels
We need to develop techniques and undertake research to determine the variability that functional diversity introduces into the adaptability of ecosystems. This subject is one of many that must be integrated into our understanding of biodiversity, but it must also be addressed explicitly in the context of global change. It should be researched through a partnership between the global change and biological diversity communities. Research on this topic is internationally coordinated by Focus 4 of the IGBP Global Change in Terrestrial Ecosystems
Core Project, by the UNESCO Man and the Biosphere DIVERSITAS program (Programme Element 2) for terrestrial systems, and by the IGBP Global Ocean Ecosystem Dynamics (GLOBEC) program for marine ecosystems. Aspects of ecosystem function that must be addressed include, but are not limited to, carbon exchange, nitrogen cycling, productivity, albedo, and hydrological processes.
Most analyses of global terrestrial ecosystems have lumped plant species into a small number of “functional” types (usually less than 30) relative to the immense diversity revealed by local studies. However, as models and experiments become more sophisticated, it has become apparent that plant species and functional type characteristics may greatly affect processes at large scales. 95 The role of microbial, fungal, and faunal diversity in large-scale ecology is just beginning to receive attention.
In marine ecology the interaction of trophic levels with physical processes is central. Whereas all animals in the sea are affected to some degree by the dynamics of the waters around them, it seems likely that planktonic animals are the most tightly controlled by the physics of the fluid medium. Since the majority of marine animals spend at least part of their lives in the plankton, research will have to focus on zooplanktonic organisms, both holoplankton and meroplankton. The holoplankton, such as numerous copepods and other macro- and microzooplankters, complete their entire life cycle in the plankton. Most nektonic animals—the large swimming forms such as fish and squid —also have at least one planktonic life stage during which they too are at the mercy of the fluid motions of the sea. Accordingly, success of recruitment into the adult population for marine animals does not depend on biological processes alone. The transport of organisms into regions that may be either favorable or inhospitable plays a major role in dictating the success of recruitment and thus the abundance of marine animals. Moreover, the characteristics of a region that determine its suitability for any given organism include not only the availability of food and abundance of predators but also the dynamic physical features of the local environment that influence the success of recruitment, efficiency of feeding, and susceptibility to predation.
Several issues arise in investigating the relationship between organism-level functional diversity and ecosystem function. First, what is the relationship between biological diversity (number of species) and functional diversity (number of species groups that differ with respect to a given aspect of function—for example, trophic characteristics, drought resistance, or nutrient use efficiency)?
-
Do systems contain more functionally different species when they contain more species overall?
-
If so, how do the two characteristics scale?
Second, what impact does functional diversity have on long-term ecosystem dynamics? Analysis has shown that increased diversity increases stability but also that diversity may contribute to vulnerability to perturbation. 96
-
What effects does the functional diversity of species have on ecosystem function within biomes and at the global level?
-
How different is global NPP, carbon storage, and trace gas production in a world with highly diverse biota, as compared to simplified biota?
-
Could potential changes to global biodiversity affect global NPP, trace gas exchange, or other critical aspects of ecosystem function?
The above questions should be used to guide a careful program of hypothesis development and testing. Both models themselves and the use of model systems for experimental studies in natural and managed landscapes are crucial. Aspects of functional diversity must be included explicitly in dynamic global vegetation models and biogeochemical models. Potential future biodiversity losses could affect global ecosystem function. The issue of functional diversity links the interdisciplinary science of global change to some of the core questions of ecology and should motivate a strong research program. 97
Summary: Key genotype to community measurements
-
Measurements of ecosystem function in experiments in which diversity is manipulated.
-
Measurements of ecosystem function in “control” and inadvertently modified systems where diversity has been altered by extinction or invasion.
-
Coordinated measurements of diversity and function in regional-scale studies.
LESSONS LEARNED
Need for Long-Term Studies and Time Series Measurements
The analysis of long observational time series has been an important feature of global change research. While many of the USGCRP planning documents for ecology emphasize long-term experimental manipulations, several do not even mention long-term observations. This may be historically consistent with ecology's past as a largely experimental field and the structure developed for global change research, but much has been gained from minimally intrusive long-term observations. Ecologists, who have always used such data opportunistically, have derived from statisticians and earth scientists rigorous analytical methods of observational records (using time series analysis and inverse modeling). Large-scale ecology includes problems that, for logistical or ethical reasons, cannot be addressed by direct manipulation (like some problems in geology and medicine). Despite favoring experimental manipulation as a means of hypothesis testing,
ecology must increasingly rely on rigorous methods using observational data to test hypotheses or in conjunction with process studies (experiments).
Long time series are critical for detecting and measuring rates of system change. Many phenomena in marine and terrestrial ecosystems occur over long periods of time, including soil carbon accumulation and turnover, nutrient accumulation, forest succession and plant community responses to climate, ocean circulation changes and consequent physical and nutrient influences on marine ecosystems, and population and evolutionary responses to environmental change.98
Long time series are also critical for model validation. Global change encompasses changes to many environmental controls over ecosystem processes. Potential consequences of global change include changes to climatic means and extremes, large-scale changes in ocean circulation, changes in ultraviolet radiation, atmospheric deposition of toxins and nutrients, trophic changes, species invasions or extinctions, and changes in landscape pattern and fragmentation. Experimental studies cannot consider all of these influences in factorial combination for logistical and technical reasons, nor can we feasibly imagine all of the potential combinations of variables that could occur. As ecosystems are monitored over time, they experience an increasingly wide range of combinations of environmental variables. By observing ecosystem responses to increasingly many combinations of variables, models (whether quantitative or conceptual) can be tested under a wide range of conditions.
Observational and experimental studies, sustained for years to decades, have played a crucial role in conclusions about ecosystems at large scales. The global observations of CO2, beginning with Keeling et al.'s Mauna Loa record, have provided an increasingly rich resource for ecosystem studies 99 (see Figure 2.8). The observed increases in atmospheric CO2 provide a key constraint, together with estimates of anthropogenic emissions (from fossil fuels, cement production, and land-use change) on estimated changes to terrestrial metabolism (hence the so-called missing sink). 100 The seasonal CO2 cycle, evident as an annual time-scale oscillation in atmospheric concentrations, has also provided a crucial tool for understanding ecosystem behavior.101 More recently, the spatial distributions of CO2 and 13CO2 have provided critical information identifying the existence and location of a region of terrestrial uptake. In the 1990s measurements of atmospheric oxygen (O2) were initiated and have likewise proved to be complementary to CO2 and 13CO2 measurements and of great value in confirming the role of terrestrial processes relative to oceanic processes102 (see Figure 2.9).
Terrestrial time series observations on plot scales (less than 10 m 2) to ecosystem scales (tens to several thousands square meters) have likewise been crucial. Time series observations of plant production (NPP) from grasslands around the world formed the centerpiece of an international collaboration organized by SCOPE.103 These datasets were used to understand the common principles relating grassland processes to climate and to validate a model of those processes and in a global-scale assessment of the potential responses of grasslands to climate
FIGURE 2.8 (a) Monthly average atmospheric carbon dioxide concentration at Mauna Loa Observatory, Hawaii. The smooth oscillating curve is a fit of the data to a trend plus the annual cycle, which increases linearly with time and does not try to connect all of the monthly data points, shown as dots. SOURCE: Adapted from Keeling et al. (1989). Courtesy of the American Geophysical Union. (b) Annual rate of change of carbon dioxide concentration at Mauna Loa. SOURCE: IPCC (1996), as updated by Keeling et al. Courtesy of the Intergovernmental Panel on Climate Change (IPCC).
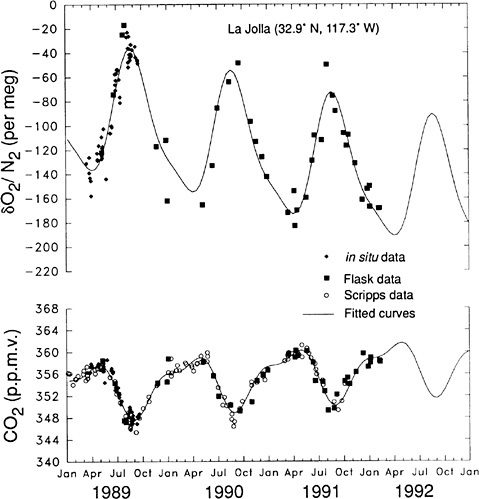
FIGURE 2.9 Measurements of L(O2/N 2) and CO2 mole fraction at La Jolla, California. The axes are scaled (5 per meg = 1ppm) so that changes in L(O2/N2) and CO2 are directly comparable on a mol O2 to mol CO2 basis. ▪, Averages of flasks taken on a given day. ◆, Daily averages for background conditions using the continuous in situ measurement method. ![]() , Supplementary CO2 data from programs at the NOAA Climate Monitoring and Diagnostics Laboratory and the Scripps Institution of Oceanography. SOURCE: Keeling and Shertz (1992). Courtesy of Macmillan Magazines Ltd.
, Supplementary CO2 data from programs at the NOAA Climate Monitoring and Diagnostics Laboratory and the Scripps Institution of Oceanography. SOURCE: Keeling and Shertz (1992). Courtesy of Macmillan Magazines Ltd.
change. Experimental plots established and sustained for over a decade by the NSF's LTER program in temperate grasslands have led to critical understanding of the role of species diversity in sustaining ecosystem processes through drought cycles and of the impact of cumulative nitrogen stress on ecosystem carbon storage and nitrogen cycling. In all this work, observations spanning substantial interannual variability (e.g., Kursk, Russia, 29 years) were critical because the responses of ecosystems to changes in temperature and precipitation were actually observed.
interannual variability (e.g., Kursk, Russia, 29 years) were critical because the responses of ecosystems to changes in temperature and precipitation were actually observed.
Long-term observations of net CO2 exchange (NEE) using micrometeorological techniques are also providing a new window on ecosystem sensitivity to climate. Eddy covariance measurements of CO2 exchange display the observed correlation at high frequency (~10 Hz) between vertical air motion and CO2 concentration to compute a flux. Prior to the 1990s, this method had only been used in a “campaign” mode to study the short-term processes governing exchange as a function of light intensity, temperature, and turbulence characteristics. Beginning in the early 1990s, a long-term effort was mounted at the Harvard Forest in Massachusetts. That time series now spans unusual freeze conditions, a major drought, and other weather fluctuations and provides information on the response of NEE to climate104 (see Figure 2.10). Continuing and expanding the network of global observations of concentrations and fluxes (CO2, 13CO2, C18O2, O2, CH4, etc.) is mandatory. Measuring fluxes in a range of ecosystems must be one goal of a network.105 Another goal is to test the quantitative relationships between climate and other drivers and ecosystem responses, a goal that appears to require measurements replicated within vegetation types and along environmental gradients.
Several key lessons can be distilled from the scientific community 's experience with time series measurements.
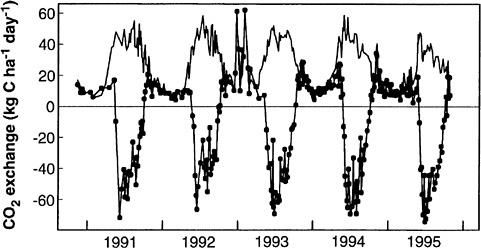
FIGURE 2.10 Daily net CO2 exchange (NEE) (filled symbols connected by lines) and daily respiration (R) (solid line) during five years at Harvard Forest. Observations are means for four days. SOURCE: Goulden et al. (1996). Courtesy of the American Association for the Advancement of Science.
-
First, the measurement should be designed around a question. Even if serendipity eventually intervenes, so that a program addresses initially unforeseen questions, more focused and successful programs emerge when they are based on initial hypotheses.
-
Second, the time series should include both the response of interest (e.g., atmospheric CO2 or net primary productivity) and the hypothesized controls over the response variable's dynamics. Without observations of both cause and effect, rejecting incorrect explanations of observed temporal dynamics is often impossible.
-
Third, data management and quality control, including metadata, must be excellent. Changes in methodology, calibration, location, and external influences (adjacency effects) must be documented over time. If not, the data will be compromised for time series analyses or needless effort will be spent on data archeology. Support for the assembly, maintenance, and distribution of datasets for synthetic ecological research must increase.
-
Fourth, ecosystem scientists have learned much from both long-term experiments and long-term observations. Long-term observational studies, designed to address specific questions, are fundamental to understanding the interaction of ecosystem and climate system processes. Rigorous methods for the analysis of observational data must be used, adapted, and implemented. The requirements of rigorous data analytical techniques must be considered when designing long-term studies. Overall, a balance between manipulative and observational studies must be preserved.
Multiscale Investigations
Ecological processes inherently occur at multiple scales in space and time. The existence of controls at multiple scales is effectively illustrated by transpiration, the loss of water from leaf surfaces. At the leaf level, transpiration occurs through tiny pores on leaf surfaces called stomata and is controlled by the stomatal aperture, itself under physiological control. At the level of an entire plant, transpiration is controlled by the aggregate of leaf-level water losses.106 At this level the amount of leaf area and light, temperature, and physiological gradients within the plant's canopy add regulation by canopy architecture, allocation of carbon and nitrogen, and leaf optical properties (which influence light extinction). 107 At yet larger scales, transpiration is influenced by boundary layer humidity and is thus controlled by the aggregate of water loss over many plants, by advection and mixing in the atmosphere, and by the effect of latent heat (water vapor) exchange on atmospheric mixing and turbulence. 108
On longer timescales, transpiration is influenced by climatic regulation of vegetation, by physiological adjustment to prevailing environmental conditions, and by long-term influences of climate on vegetation types. There is abundant
evidence for an adjustment of leaf area to an equilibrium with prevailing soil moisture conditions. This occurs both within vegetation types and through the distribution of species and ecotypes along moisture gradients.
Terrestrial nitrogen cycling exhibits similar scaling. On short timescales, nitrogen availability is determined by the biophysical regulation of the activity of soil microbes, with nitrogen mineralization increasing as conditions become more microclimatically favorable for soil biological activity. On somewhat longer timescales, nitrogen cycling is greatly influenced by changes in litter organic chemistry, which may change with the growth stages of a plant or over successional changes, as species with different litter chemistries change in abundance over time.109 At longer timescales yet (centuries), nitrogen cycling may be dominated by changes in the amount of nitrogen contained in the ecosystem, controlled by the balance between inputs from the atmosphere and outputs to the atmosphere and hydrological cycle. Thus, variations in atmospheric deposition at mesoscales or larger, and in hydrology at the watershed or larger scale, are key controls on N cycling over very long timescales.110
Nitrogen cycling in marine ecosystems is similarly dominated by a fast cycle on short timescales, governed by local trophic interactions and microbial recycling. But on seasonal to millennia timescales, transport of nitrogen to the euphotic (sunlit) zone is controlled by the coupling of large-scale ocean circulation to ocean biogeochemistry. 111 Variations in nitrogen supply occur spatially on length scales of hundreds to thousands of kilometers and on timescales associated with interannual variability in coastal upwelling (e.g., N availability varies with the ENSO cycle) to longer timescales associated with variations in global ocean circulation (decadal to glacial-interglacial timescales). 112
Because of these multiple scales of regulation, issues of scale are critical in ecology. This fact has long been recognized: the first science workshop explicitly charged with addressing the science agenda of the IGBP produced the volume Scales and Global Change.113 Over the past decade a number of projects have addressed ecological problems and ecosystem-atmosphere exchange at multiple scales. This approach has been extraordinarily fruitful and has also catalyzed successful interdisciplinary interactions (e.g., regarding the intersection of physiological and boundary layer meteorological controls over transpiration).
It has proved resolutely difficult to extrapolate from basic enzymatic or physiological controls to large scales and long time intervals because of the hierarchy of control across scales. For example, in the early 1980s the problems of calculating whole-plant or ecosystem photosynthesis from first principles of leaf physiology seemed intractable and empirical models dominated.114 Two developments changed this situation. Work on leaf physiology along light extinction gradients within plant canopies demonstrated regularities in the relationship of canopy environmental variables to leaf physiology. The development of eddy covariance measurement techniques also allowed canopy flux measurements to be made so that the “answer” to the problem of scaling leaf gas ex-
change to the canopy was known. Today there are many successful models of ecosystem CO2 exchange based on first principles of photosynthesis and stomatal regulation. Two lessons can be learned from this research example and from experience with multiscaled measurements in general:
-
First, measurements of phenomena and their controls on multiple scales are crucial to knowledge at large scales. Fine-scale knowledge allowed studies of the environmental patterns of controlling variables to be appropriately designed. Large-scale measurements then provided a scaling theory for extrapolation.
-
Second, extrapolation to ecosystems and the globe from just the fine-scaled or “fundamental” body of knowledge can lead to confusion, stasis (as large-scale model predictions and observations persistently fail to agree), or fundamental error. For example, in the absence of a scaling theory for leaf physiological characteristics within the canopy, model error propagation dominated simulations of canopy photosynthesis.
Need for Technical Infrastructure in Ecology
Major advances in understanding terrestrial ecosystems have resulted from the use of new measurement and data analytical techniques. Such measurement techniques have included flux measurements by eddy covariance, isotopic measurements, remote sensing of vegetation canopies, and measurements of atmospheric O2. New inference techniques, especially the application of inverse modeling techniques, also have been important. The infrastructure for the development of new technologies and their adoption and implementation is weak in ecology. There are few institutions providing engineering, instrumentation, or electronics support for ecologists, and relatively few ecologists receive training in advanced instrumental techniques. Only in the past few years have disciplinary ecologists received rigorous training in remote sensing, and the number being trained with the technical know-how to effectively participate in the design of new sensors is still small. Ecology has no equivalent of the major facilities and engineering staffs supported for the atmospheric sciences by the National Center for Atmospheric Research or for oceanography by the major oceanographic research centers and the University National Oceanographic Laboratory System. In ecology, long-term funding commitments analogous to those for research vessels, advanced instruments, and aircraft tend to be made to field stations, which for the most part do not support the development of new observing technology.
Thus, the suite of advanced techniques that have transformed our understanding of ecosystems (flux measurements, isotopic techniques, remote sensing, inverse modeling, atmospheric oxygen) have largely originated outside ecology. Ecologists, however, have effectively adopted techniques from other fields: for
example, flux measurements from atmospheric chemistry and micrometeorology and isotopic techniques from geochemistry. The interdisciplinary projects sponsored by the USGCRP (e.g., Arctic Systems Science [ARCSS], FIFE, BOREAS, Oregon Transect Ecological Research) have resulted in many more ecologists becoming exposed to flux measurements, advanced instruments, aircraft platforms, and remote sensing techniques. The ability of ecologists to adapt and, where necessary, to develop new measurement techniques must be enhanced. As the opportunities for space missions initiated by principal investigators increase, there should be a growing cadre of ecologists with the technical expertise to design and implement remote sensing missions, from instrument and spacecraft design through algorithms and data analysis. At a minimum, training and apprenticeship opportunities for students and practitioners of ecology in instrument principles and development can be expanded. Beyond that, partnerships with national labs, with disciplines containing more expertise in instrumentation and instrument development, and with the private sector could all be encouraged. Precedent exists for both of these approaches, and concern over ecology 's technological weakness is not new. 115
Increased adoption of advanced inference techniques should also be fostered. The situation in this case differs somewhat from that for instrumentation. Many ecologists are familiar with advanced statistical and simulation modeling techniques. Many ecologists possess strong mathematical and computational skills,116 and biologists have appreciably contributed to the development of mathematics. Encouraging the application of advanced techniques for interpreting observational data and for model-data fusion will build on a strong foundation. There are clear needs for the infusion of quantitative inferential methods such as time series analysis, Kalman filters, data assimilation and adjoint techniques, inverse modeling, and many other approaches in ecology, as well as novel adaptation of these methods to important ecological problems. A series of workshops, tutorial publications, and apprenticeship opportunities could accelerate the adoption of new inference techniques in ecology. Partnerships among agencies supporting ecology and applied mathematics and statistics could provide a valuable funding vehicle for this process.
Several lessons may be drawn from the experience of the past decade.
-
The capability to adopt and develop advanced technology requires an investment in supporting infrastructure; such an investment is made in many disciplines.
-
Training of ecological scientists, postdoctoral fellows, and students in advanced technologies to produce a cadre that can develop new technologies and be more effective partners with technologists would be of great value to the field.
-
Advanced inference techniques have been a critical force in advancing the physical global change disciplines. Many of these techniques (e.g.,
-
data assimilation, inverse modeling) are foreign to the training of most ecologists, who often have strong quantitative backgrounds. Joint projects and workshops with applied mathematicians and statisticians could quickly transfer such advanced techniques to ecologists.
Organizing Interdisciplinary Ecosystem Science
As global change research has evolved over the past decade or so, considerable work has been done at the interface of ecology, traditionally thought of as a life science, and the earth sciences, especially atmospheric science and geochemistry. Some of the major advances have come in studying the carbon cycle. The carbon cycle exemplifies a science area where rigorous interdisciplinary research is needed, and lessons from this research area are informative about how interdisciplinary science can function with excellence, as well as its potential weaknesses. Similar lessons could be drawn from studies of vegetation and hydro-meteorology and of biological sources of trace gases. Study of the carbon cycle required an interdisciplinary approach for several reasons:
-
Both terrestrial and marine biological processes are central to its regulation.
-
Inference from measurements of carbon in the atmosphere and oceans requires a profound knowledge of transport processes in fluid media.
-
The carbon system is perturbed by human land use and energy production, requiring the linkage of natural science inquiry to the social and economic sciences.
An analysis of carbon cycle research allows the strengths and weaknesses of the USGCRP's support for interdisciplinary research to be assessed. Many seminal papers on the carbon cycle emerged as the authors found they had common problems and joined forces, rather than being written by groups of people specifically funded to work together. d Many of the collaborations were in fact international. For example, by the mid-1990s, papers utilizing CO2 concentrations and isotopes were typically authored by interdisciplinary teams collaborating informally. This work was motivated by excitement about what could be accomplished by pooling knowledge, by the self-evident importance of the problem scientifically and societally, and by a sense of healthy competition with other informal teams. The arena for much of this work was found in meetings for which ecologists and a broad range of earth scientists were funded to examine interdisciplinary linkages, such as the carbon cycle symposia of the late 1970s
|
d |
Papers cited in the 1990, 1992, 1995, and 1996 IPCC assessments of the carbon cycle served as a database. |
and early 1980s and more recently such forums as the Office of Interdisciplinary Earth Sciences (OIES) Global Change Institutes and the German Dahlem Conferences as well as at IGBP, European Space Agency, and American Geographical Union meetings.
This working style was also sustained by a funding environment that, although relatively healthy, provided few opportunities for atmospheric scientists, oceanographers, and ecologists to seek funding together through a single venue. Regardless of the success of symposiums such as those just mentioned, joint research programs have been much more difficult to initiate. An NRCNews Report article described the Tropical Oceans and Global Atmosphere (TOGA) program in its early days as follows: “Research efforts were strongly divided between meteorologists and oceanographers, and the disciplinary boundaries were significant” but as a result of TOGA, “meteorologists and oceanographers worked together, crossing disciplinary and institutional barriers in universities and government.”117 This could well be a description of the current state of carbon cycle research and how it might change—especially with respect to institutional barriers in government. While barriers exist between atmospheric, ecological, and ocean scientists, the interagency Terrestrial Ecosystems initiative may be a model for future carbon science programs as it allows several agencies to jointly manage a combined competition for ecologically oriented proposals. A joint review of all proposals is conducted on the basis of scientific merit without great attention to which agency will ultimately fund the proposal. Although great progress has been made in understanding the carbon cycle—accompanied by a liveliness reflecting the informal and collegial nature of the program—the fragmentation of the problem has produced one demonstrable weakness. Because there is no “U.S. National Carbon Cycle Research ” agenda, it is very difficult to evaluate the state of knowledge of the field as a whole and or to establish research priorities. Whereas this can be done within disciplinary areas (e.g., atmospheric science, oceanography) or occasionally within agencies, integration cannot be achieved for the field. Perhaps as a consequence, while great progress has been made in some areas, some questions have persisted for decades. Even within the IGBP the carbon cycle is just emerging as a coherent program-level activity, spanning the terrestrial Global Change and Terrestrial Ecosystems Core Project, oceanic (JGOFS, Land-Ocean Interactions in the Coastal Zone, IGAC), and integrative (Global Analysis, Interpretation, and Modeling) programs.
Several key lessons for interdisciplinary research can be extracted from this history.
-
First, the excitement and rewards of working in important areas are sufficient to motivate a significant amount of research, major commitments to collegial collaborations, and progress. This occurs even in the absence of a formal program to address a problem. The effort on the carbon cycle, which is not funded in a coordinated manner or attacked as a USGCRP
-
priority, is an apt comparison to the programmatic approach taken to El Ninõ-Southern Oscillation or stratospheric O3.
-
Second, the absence of either a formal program for carbon cycle research (along the lines of TOGA) or the rigidity of the standing programs (e.g., the disciplinary programs at NSF) restricts the full participation of the scientific community. Programs intended to promote interdisciplinary team building can be effective in developing “institutes without walls,” as alternatives to center- or institute-style programs. Supporting this conjecture, university investigators in collaborative programs such as NSF's JGOFS, LTER, and ARCSS programs and the National Aeronautics and Space Administration's Mission to Planet Earth (MTPE) program seem disproportionately important contributors to the interdisciplinary literature.
-
The development of effective interdisciplinary teams, especially those that cross major disciplinary boundaries (e.g., life and earth sciences), requires the existence of stable support aimed specifically at interdisciplinary questions. Such awards need not be large per principal investigator, but they must meet at least three criteria: (1) they need to be in place long enough to allow the development of effective interdisciplinary communication; (2) the group must be explicitly reviewed on its ability to function as a team; and (3) the project must be judged high-quality science on the basis of initial and, for long-term projects, ongoing and rigorous peer review. The last criterion is crucial both for recruitment of top scientists into such partnerships and for community acceptance of nontraditional funding vehicles. Long-term, problem-oriented, team-oriented, peer-reviewed research should be a part of the large-scale ecosystem research funding portfolio. Such investigations should be available for a wide range of science problems as the scientific agenda continues to evolve. When research areas, like the carbon cycle, require the partnership of disciplines whose funding mechanisms are not traditionally coordinated, mechanisms must be developed to allow the establishment of problem-oriented rather than disciplinary priorities within and across agencies.
RESEARCH IMPERATIVES
Again, critical Research Imperatives for ecosystem studies in the USGCRP for the next decade include the following:
-
Land surface and climate: Understand the relationships between land surface processes and weather prediction and changing land cover and climate change.
-
Biogeochemistry. Understand the changing global biogeochemical cycles of carbon and nitrogen.
-
Multiple stresses. Understand the responses of ecosystems to multiple stresses.
-
Biodiversity. Understand the relationship between changing biological diversity and ecosystem function.
Observations and Experiments
Research on global ecosystem processes motivates four broad classes of observations and experimental studies. As noted in the section “A Research Agenda for the Next Decade,” large-scale measurements in ecology tend to support all of the Research Imperatives above in a crosscutting fashion, with any one measurement set helping to test a variety of hypotheses. The four key measurement areas are the following:
-
Time series observations of ecosystem state.
-
Land-use and land cover change.
-
Site-based networks.
-
Measurements of diversity, functional diversity, and ecosystem function.
Time Series Observations of Ecosystem State
Global time series of vegetation and phytoplankton state, derived from NOAA's AVHRR and Coastal Zone Color Scanner sensors, for land and ocean, respectively, have proven their value in understanding the seasonal and spatial characteristics, interannual variability, and trends of large-scale biogeochemistry and biophysical processes. 118 Space-based measurements of ecosystem state are fundamental in determining the link of terrestrial ecosystems to climate, the biogeochemistry of the land and oceans, and the impacts of climate and other disturbances. While the measurements of “greenness” and ocean color are not direct ecological properties, they have proven to be highly correlated with the spatiotemporal dynamics of ecosystems. Recent work highlights both the utility of these records and the dependence of the science on long consistent records.119 Stable calibration and removal of the atmospheric signals of ozone, water vapor, and aerosols are critical to detecting ecological signals. While there is ample room for innovation in land surface remote sensing, stable calibration and correction impose stringent requirements on the sensor or sensors deployed. New instruments, while adding new capabilities, must also be “backwards compatible” to preserve time series. Atmospheric correction requires that coincident observations to quantify water, ozone, and aerosols be available for use in land surface retrieval algorithms. Spatial resolution and temporal resolution for time series instruments are typically a compromise (0.25 to 1 km 2) between suffi-
ciently high spatial resolution to resolve ecosystem structure and swath width and data rate limitations associated with near-daily coverage. High temporal coverage is needed to ensure adequate sampling of seasonality, especially in cloudy environments. These requirements apply generally for both terrestrial and marine ecosystems: marine ecosystems add additional instrument requirements to avoid saturation by sun glint or high reflection. Note that data for land cover change require higher spatial resolution but lower temporal resolution.
Land Cover, Land Use, and Disturbance
Changing land use and land cover are fundamental drivers of global change and are direct reflections of human activity and impacts. Land-use changes have profound effects on the biogeochemistry of carbon, infrared-active gases, photochemically active gases, and aerosol production (via dust and biomass burning). Land-use changes also affect hydrology and erosion and by changing surface albedo and energy exchange can have direct effects on climate. People often create highly heterogeneous landscapes—mosaics that may encompass activities with highly divergent effects on ecological processes. The spatial arrangement of landscapes can affect exchanges of water and associated solutes and particulates in freshwater and coastal margin areas, with land cover at the land-water margins having substantial effects on water chemistry. The arrangement of landscapes also affects biological diversity, pests and pathogens, invasiveness and extinctions. Data on land cover and its change over time must thus capture the spatial scales of natural and human patterns. Space-borne sensors with resolutions from a few square meters to tens of square meters have proven to meet these needs. 120 Sensors with two to seven spectral bands are adequate for land-cover mapping, although new technology using spectrometers,121 radar, or lidar has great potential to provide new information. As in measuring ecosystem state, backwards compatibility must be preserved to continue existing time series when new technologies and capabilities are introduced.
Site-Based Networks
In situ measurements of ecological processes tend to be highly multivariate. In terrestrial systems, understanding a measurement of CO2 flux and determining NPP require sampling multiple plant parts (leaves, wood, roots), often of several life forms (e.g., co-occurring grasses, shrubs, and trees). The plant parts are then analyzed for carbon and nitrogen. Understanding spatial variations in gradients of atmospheric CO2, 13CO2, CO, 18O, or O 2 requires a network of such sites over large areas. To understand process, leaf physiology, soil microbial processes, water fluxes, and other variables must be determined. Parallel issues arise in marine ecosystems regarding trophic dynamics and transport. At many sites these measurements are made as part of an experimental design including con-
trols and various manipulations (e.g., of nutrients, species composition, disturbance frequency). These measurements are the essence of ecological data: the satellite and other geographic data serve to knit together disparate process studies in space and time. While there is much interest in a common set of quantities in ecological site studies (quantities such as CO2, trace gas and water fluxes, NPP, nutrient availability, and species composition and diversity), achieving consensus on a core set of measurements, standard methods, and data formats is just beginning. No global-scale experimental design implementing such sites is in place to sample the marine and terrestrial ecosystems (such a design is proposed by the IGBP, using long baseline transects across ecological gradients). A high priority of global ecosystem science is to develop a network of appropriately sited atmospheric concentration and isotope, flux, and ecological process sites. Both the overall experimental design and the suite of measurements and methods must be decided. Minimally intrusive measurements (e.g., flux measurements) and manipulations (e.g., of CO2 concentration) must be components of such a network design.
Measurements of Diversity, Functional Diversity, and Ecosystem Function
The issue of diversity and species composition changes has emerged as a critical topic of global change in recent years. It is clear that the functional diversity of the Earth's biota is a first-order control over global ecosystem function, but how changes to the biota will affect global ecosystem function still is a young research topic. 122 Designing a global observing system and network of experimental studies, analogous to those described above for biogeochemical fluxes, is premature; the necessary monitoring and manipulations at global scales are currently far from obvious. But a major exploratory effort involving manipulations, studies of ecosystem function in the face of ongoing invasions, extinctions, and species range shifts, along with global monitoring of species diversity, invasion, and extinction rates, are all needed. These exploratory studies will lay the groundwork for a more systematic attack. The foundation for systematic study and monitoring of changing diversity and functional diversity must be laid quickly and a global research program put in place.
Modeling and Theory for Integration in Time and Space and Across Ecological Levels of Organization
Models of ecosystem processes have become fundamental tools in Earth system science. The development of models over the past five years has been extremely rapid, and ecosystem models have become important parts of coupled climate system models, analyses of contemporary global data, and assessments of global change.123 Currently, global terrestrial ecosystem models fall into three broad categories: (1) biophysical models that address the exchange of water,
energy, and carbon on minute to seasonal timescales, such as the “fast water-carbon-energy system”; (2) biogeochemical models that address the dynamics of carbon, nitrogen, water, and energy on monthly to centennial timescales and that focus on carbon storage; and (3) biogeographic models that simulate the response of vegetation (“biome”) distribution to climate and other factors124 (see Plate 4, Table 2.2). The next generation of models, the so-called dynamic global vegetation models, couple the above three types of models because the time-dependent change of vegetation results from interactive biophysical, biogeochemical, and population-community processes. 125
Marine ecosystem models often simulate the coupled dynamics of carbon and nutrients (usually nitrogen) but differ from terrestrial ecosystem models in that the focus, even in biogeochemical models, is much more on trophic interactions. The understanding of processes that determine abundance, fluctuation in abundance, and production of marine animals must necessarily involve coupled physical-biological models, linking performance of the individual organism to local physical processes, and linking both the biology and local physics to basinscale changes in global climate. Modeling is expected to play an increasingly significant role at several levels. The explicit incorporation of physical variables and processes in biological population models holds promise for great originality and progress. Appropriately constructed models of both physical and biological processes should guide the choice of field experiments and observations, while results of those field exercises should feed back interactively into the models.
Clearly, ecosystem models are an important aspect of data analysis, experimental design, and prediction in ecology. For terrestrial and marine ecosystem models to become powerful and credible tools for disciplinary and interdisciplinary science and more useful for assessment, rapid progress must continue. There are a number of imperatives for the development of improved models.
-
Global models and key datasets should be continually related, with models being compared to observations and experiments and new observations and experiments arising from model uncertainties and failures. Ecosystem modelers and observationalists must work together to define data requirements for operating and evaluating ecosystem models. 126
-
Models are becoming essential to understanding complex datasets in ecology. 127 Advanced data-model fusion techniques, such as data assimilation, adjoint modeling, and inverse modeling, must join the ecological tool kit.
-
Currently, few ecosystem models include both intensively managed and wildland ecosystems. Many global and regional experiments are done using “potential natural” vegetation. Global models must incorporate the impacts of human land use along with other environmental factors. In addition, although the role of disturbance is central in site- and landscape-level ecosystem models, many global models have simplified the
-
modeling of disturbance effects.128 Future models must adequately include the effects of disturbance on ecosystem heterogeneity and on water, carbon, and nutrient fluxes. 129
-
Most global ecosystem models are run using climate as an external driver. Ecosystem models are required that can be run as components of Earth system models for a range of timescales to simulate the coupled effects of changing ecosystems and changing physical and chemical climate. These models must simulate biophysical processes, biotic uptake and storage of carbon, and ecosystem effects on atmospheric CO2 and other trace gases.
-
Changes to the distribution of species and/or functional types will have profound effects on ecosystem functioning. Current models represent the population processes underlying vegetation change in the form of calibrated empirical models. A stronger theoretical basis and scaling theory for large-scale biotic change must be developed.
Long-Term Questions for Ecology and Ecologists
Taking a longer perspective, two issues must eventually be confronted. First, although we have at least the beginnings of a theory for biophysical and biogeochemical processes at larger scales, we lack an integrative understanding of the biological processes that regulate the interactions of ecosystems with global change. Simply put, there is not yet enough biology in our paradigm for global ecology. Currently, global ecosystem models use reasonably general formulations for processes such as photosynthesis and decomposition, formulations that are based in theory and supported by strong empirical evidence. Processes that vary at the organism level, such as energy or nutrient allocation within plants, biochemical plasticity of microorganisms, and trophic interactions, are typically neglected or represented so as to agree with limited empirical data. Responses of processes such as allocation, herbivory, or microbial decomposition to environmental change reflect the genetically constrained adaptability of organisms, genetic variations in relevant traits, and rates of population processes.
On large scales and in the long term these evolutionary constraints may substantially influence the trajectory of ecosystems, but to date we do not have theories or empirical data of sufficient breadth or robustness to predict any biological limitations to adaptation under global change.130 Because of these unknown limitations, considerable uncertainty exists in quantitative projections of future ecosystem states. Current dynamic global vegetation models use empirical formulations for large-scale controls on plant community processes, rather than making explicit reference to the actual underlying population biology. Developing a more integrated and general theory for the population and biogeographic bases for ecosystem change is a high priority for the long term. Current efforts should be evaluated against this ambitious long-term goal.
Second, with the expectation that human populations and economies will continue to grow, the direct human impact on ecosystems is almost certain to increase. Increased management of the biosphere will be a crucial part of any response to global change to ensure the sustainability of key natural resources in the face of climate change and direct human impacts.131 In addition, management of ecosystems is an important aspect of mitigation for CO2 and other trace gases, the emissions of which to the atmosphere are dependent on land cover, fertilization, and other human impacts. 132 Quantitative knowledge of the roles of ocean and terrestrial ecosystems is indispensable for any future agreements to stabilize CO2, N 2O, and methane. It is essential to assess proposed mitigation measures for greenhouse gas emissions and experimental manipulations with ecosystem models prior to implementing such measures. These efforts may depend on the response of long-lived species such that experimental evaluation would require decades. The economic and nonmarket impacts of ecosystem change are just beginning to be recognized, and these impacts will likely become increasingly important drivers of policy. 133
As the needs and potential for ecosystem management in environmental mitigation increase, the scientific potential of mitigation must be continually assessed. Attempts at ecological mitigation and restoration have a mixed record of success, and scientific progress does not guarantee that the knowledge required for successful management has been developed. As the global change program advances, the feasibility and value of mitigation through ecosystem management must be assessed continually. Although many proposals for mitigation such as afforestation, as proposed by the IPCC,134 have great appeal, many of the realized projects are either more difficult to accomplish than originally envisioned135 or are affected by the “law of unintended consequences”—that is, the mitigation practice has some unexpected environmental consequences. While it is likely that ecologically based mitigation will play an increasing role in environmental management, better understanding at the system level is required for new management strategies to have a high probability of success.
In conclusion, as we look five or ten years into the future of global change research, integration of empirical knowledge and theory of specific systems must be used to produce more general and generally applicable theories, in some cases for phenomena for which we currently have no quantitative understanding. As the human impact on ecosystems becomes ever greater and the motivation to involve ecosystem management in mitigation strategies increases, there is evergreater need for an improved scientific capability to assess the likely success of proposed manipulations at the system level. Ecology must develop a robust predictive capability for environmental change and management consequences.
NOTES
1. NRC (1986).
2. NRC (1994).
3. Andreae and Schimel (1989); Matson and Ojima (1990).
4. Breymeyer et al. (1996).
5. For example, Keeling et al. (1989) and Tans et al. (1990).
6. For example, Ciais et al. (1995a, b); and Denning et al. (1995).
7. For lab studies, see, e.g., Holland et al. (1995); for field studies on soil warming, see Harte and Shaw (1995); for observational programs, see Oechel et al. (1993); for data synthesis, see Raich and Schlesinger (1992).
8. Terrestrial ecosystems, Jenkinson et al. (1991), Townsend et al. (1992), Dai and Fung (1993), VEMAP Participants (1995), Schimel et al. (1996); marine ecosystems, Kleypas (1997).
9. Ecosystem response to climate, IPCC (1990); temperature and moisture effects on biota, e.g., Raich and Schlesinger (1992); climate and succession, Pastor and Post (1993); climate and nitrogen cycle, Schimel et al. (1994).
10. Oechel et al. (1994).
11. Eddy covariance techniques, Lenschow (1995); NEE, Wofsy et al. (1993).
12. Goulden et al. (1996).
13. Woodwell (1989); Woodwell and Mackenzie (1995).
14. Braswell et al. (1997).
15. Mortality, Smith and Shugart (1993); soils, Jenkinson et al. (1991); nitrogen cycle, Melillo et al. (1993); Schimel et al. (1994); interactions, VEMAP Participants (1995).
16. E.g., Ciais et al. (1995a, b), Schimel et al. (1996), Thompson et al. (1996), Bruno and Joos (1997), Randerson et al. (1997).
17. E.g., Pastor and Post (1986), Schimel et al. (1990), Melillo et al. (1993)
18. Hunt et al. (1996), Schimel et al. (1996), Waelbroeck et al. (1997).
19. Land cover and climate, Anthes (1984), Dickinson and Henderson-Sellers (1988), Pielke and Avissar (1990); land surface processes, Henderson-Sellers (1993), Sellers et al. (1997); international field campaigns, Sellers et al. (1995, 1996c); LBA Concise Experimental Plan, http://www.cptec.inpe.br/lba/conciseplan/cpi06.html.
20. Verma et al. (1993).
21. Canopies, Field and Mooney (1986); canopy modeling, Sellers et al. (1992).
22. Bonan et al. (1992).
23. Farquhar et al. (1993), Flanagan et al. (1997).
24. Paleoclimate, Kutzbach et al. (1996); contemporary weather, Pielke et al. (1992); climate, Shukla et al. (1990); weather forecasting, Pielke et al. (1997); climate projections, Sellers et al. (1996b, 1997).
25. Entekhabi et al. (1992), Sellers and Schimel (1993), Braswell (1996), Hunt et al. (1996), Sellers et al. (1997).
26. FIFE, Sellers et al. (1996c); BOREAS, Sellers et al. (1995); HADEX, André et al. (1989).
27. Potter et al. (1993), Sellers and Schimel (1993).
28. Luizão et al. (1989), Crutzen and Andreae (1990), Mosier et al. (1991, 1997), Keller et al. (1993).
29. Vitousek et al. (1986).
30. E.g., Skole and Tucker (1993); Defries and Townshend (1994).
31. Houghton et al. (1983).
32. Skole and Tucker (1993).
33. Groffman and Likens (1994); Woodward and Steffen (1997).
34. Wedin and Tilman (1996).
35. Lodge (1993), Rejmanek and Randall (1994), Carleton (1996).
36. Vitousek et al. (1996).
37. Niemela and Mattson (1996).
38. D'Antonio and Vitousek (1992).
39. Carpenter and Kitchell (1993), Jones and Lawton (1995).
40. Tilman and Downing (1994), Lawton (1995), Tilman et al. (1996), Wedin and Tilman (1996).
41. E.g., Sellers et al. (1996a).
42. Carpenter et al. (1995).
43. Oechel et al. (1994).
44. Drake (1992).
45. Idso and Idso (1994).
46. Chapin et al. (1995).
47. Harte and Shaw (1995).
48. Wedin and Tilman (1996).
49. Carpenter et al. (1995).
50. Martin (1990).
51. Coale et al. (1996).
52. Chameides et al. (1994).
53. Galloway et al. (1995), Howarth et al. (1996).
54. Peterson and Melillo (1985).
55. Schindler and Bayley (1993), Townsend et al. (1995), Bruno and Joos (1997), Holland et al. (1997).
56. Holland et al. (1997).
57. Ibid.
58. Aber et al. (1989), Schulze et al. (1989), Wedin and Tilman (1996).
59. Schimel et al. (1995, 1996, 1997a, b).
60. Siegenthaler and Sarmiento (1993).
61. Schimel et al. (1995).
62. Krankina and Dixon (1994), Oechel et al. (1994), Archer et al. (1995), Sellers et al. (1995), Brown et al. (1996).
63. Comins and McMurtrie (1993).
64. VEMAP Participants (1995).
65. Friedlingstein et al. (1995), Randerson et al. (1997).
66. Post et al. (1997), Randerson et al. (1997).
67. Schindler and Bayley (1993), Townsend et al. (1996).
68. Pastor and Post (1993), Bolker et al. (1995), VEMAP Participants, (1995), Schimel et al. (1996).
69. Smith and Shugart (1993).
70. VEMAP Participants (1995), Schimel et al. (1997a, b), Kittel et al. (1996).
71. Sponsored by SCOPE, Breymeyer et al. (1996), and the IGBP's GAIM Task Force; Cramer et al. (in preparation) and by Electric Power Research Institute's Carbon Cycle Model Linkage Project.
72. VEMAP Participants (1995).
73. Field, Stanford, personal communication.
74. Schimel et al. (1997a, b).
75. Ibid.
76. Ibid.
77. Schimel et al. (1996).
78. Andreae and Schimel (1989), Ojima and Svensson (1992).
79. Direct effects of climate, Gifford et al. (1997) interactions with nitrogen cycle, VEMAP Participants (1995), Schimel et al. (1996), Randerson et al. (1997); interactions with other stresses, Schindler and Bayley (1993), Townsend et al. (1996), Holland et al. (1997).
80. Chameides et al. (1994).
81. Howarth et al. (1996).
82. Carbon storage, Goulden et al. (1996); trace species, Bakwin et al. (1995).
83. Mosier et al. (1997).
84. Responses to trends, Oechel et al. (1993), climatic events, Schimel et al. (1996), Wedin and Tilman (1996).
85. Cumulative impacts, e.g., Walker et al. (1987); Holland et al. (1997); analyses of land-use effects, e.g., Houghton et al. (1983).
86. Ambio vol. 23 (1994), Turner et al. (1995).
87. Asner et al. (1997), Holland et al. (1997).
88. Schimel et al. (1997a).
89. E.g., Dlugokencky et al. (1994), Hein et al. (1997).
90. Bakwin et al. (1995), Hurst et al. (1997).
91. Myneni et al. (1996, 1997); Braswell et al. (1997); Randerson et al. (1997).
92. Sellers et al. (1997), Braswell et al. (1997).
93. Harriss et al. (1994).
94. Groffman and Likens (1994).
95. Bolker et al. (1995), Schimel et al. (1996), Wedin and Tilman (1996).
96. Stability, Lawton (1995); vulnerability, Wedin and Tilman (1996); in their experiment the presence of low C-to-N ratio C3 plants reduced system-level nitrogen retention.
97. http:\\www.unesco.org/mab/collab/diver1.htm.
98. Pastor and Post (1986), Harden et al. (1992).
99. Keeling et al. (1996).
100. Schimel (1995).
101. Randerson et al. (1997).
102. Keeling and Shertz (1992).
103. Breymeyer et al. (1996).
104. Goulden et al. (1996).
105. Global Change Biology, vol. 2 (1996).
106. McNaughton and Jarvis (1991).
107. Schimel et al. (1991).
108. McNaughton and Jarvis (1991).
109. Aber and Melillo (1982).
110. Asner et al. (1997).
111. Najjar (1992).
112. Carpenter and Capone (1983).
113. Rosswall et al. (1988).
114. E.g., Shugart et al. (1988).
115. Hibbard and Shaulis (1997).
116. Levin (1992).
117. NRC (1997).
118. Sellers et al. (1997), Fung et al. (1987), Myneni et al. (1997), Potter et al. (1993), Randerson et al. (1997), Braswell et al. (1997).
119. Braswell et al. (1997) and Myneni et al. (1997).
120. Skole and Tucker (1993).
121. Wessman (1988).
122. Vitousek et al. (1996), Braswell et al. (1997), Schimel et al. (submitted).
123. Models, Sarmiento and Le Quéré (1996), Sellers et al. (1996a); data analyses, Denning et al. (1995), Sarmiento et al. (1995), Schimel et al. (1996); assessments, Melillo et al. (1993), VEMAP Participants (1995), Sarmiento and Le Quéré (1996).
124. (1) Sellers and Schimel (1993), (2) VEMAP Participants (1995); (3) Ibid.
125. Foley et al. (1996).
126. Sellers and Schimel (1993).
127. Ciais et al. (1995a, b), McKane et al. (1997a, b).
128. Ecosystem models, Pastor and Post (1986), Shugart et al. (1988); global models, VEMAP Participants (1995).
129. VEMAP Participants (1995), Schimel et al. (in press).
130. Karieva et al. (1993).
131. IPCC (1995), Wisnewski and Sampson (1993).
132. Schimel et al. (1997b).
133. Watson et al. (1996), Schimel et al. (1997b).
134. IPCC (1990).
135. Joos et al. (1991).
REFERENCES AND BIBLIOGRAPHY
Aber, J.D., and J.M. Melillo. 1982. Nitrogen immobilization in decaying hardwood leaf litter as a function of initial nitrogen and lignin content. Canadian Journal of Botany 60:2263-2269.
Aber, J.D., J.K. Nadlehoffer, P.A. Steudler, and J.M. Melillo. 1989. Nitrogen saturation in northern forest ecosystems—hypotheses and implications. BioScience 39:378-386.
André, J.-C., J.P. Goutorbe, T. Schmugge, and A. Perrier. 1989. HAPEX-MOBILHY: Results from a large-scale field experiment. Pp. 13-20 in Remote Sensing and Large-Scale Global Processes, A. Rangos, ed. International Association of Hydrological Sciences, Wallingford, U.K.
Andreae, M.O., and D.S. Schimel, eds. 1989. Exchange of Trace Gases Between Terrestrial Ecosystems and the Atmosphere. John Wiley & Sons, New York.
Anthes, R.A. 1984. Enhancement of convective precipitation by mesoscale variations in vegetative covering in semiarid regions. Journal of Climate and Applied Meteorology23:541-554.
Archer, S., D.S. Schimel, and E.A. Holland. 1995. Mechanisms of shrubland expansion: Land use, climate or CO2? Climatic Change 29:91-100.
Asner, G.P., T.R. Seastedt, and A.R. Townsend. 1997. The decoupling of terrestrial carbon and nitrogen cycles. BioScience 47:226-234.
Bakwin, P.S., C. Zhao, W. Ussler III, P.P. Tans, and E. Quesnell. 1995. Measurements of carbon dioxide on a very tall tower. Tellus 47B:535-549.
Bolker, B.M., S.W. Pacala, F.A. Bazzaz, C.D. Canham, and S.A. Levin. 1995. Species diversity and ecosystem response to carbon dioxide fertilization: Conclusions from a temperate forest model. Global Change Biology 1:373-381.
Bonan, G.B., D. Pollard, and S.L. Thompson. 1992. Effects of boreal forest vegetation on global climate. Nature 359:716-718.
Braswell, B.H. 1996. Global terrestrial biogeochemistry: Perturbations, interactions, and time scales. Ph.D. dissertation, Department of Earth Sciences, University of New Hampshire.
Braswell, B.H., D.S. Schimel, E. Linder, and B. Moore III. 1997. Global terrestrial of ecosystems respond to interannual temperature variability over multiple time scales. Science 278(5339):870.
Breymeyer, A.I., D.O. Hall, J.M. Melillo, and G.I. Agren, eds. 1996. Global Change: Effects on Coniferous Forests and Grasslands. John Wiley & Sons, New York.
Broecker, H.C., J. Petermann, and W. Siems. 1978. The influence of wind on carbon dioxide exchange in a windwave tunnel including the effects of mono-layers. Journal of Marine Research 36(4):595-616.
Broecker, W.S.J.R. Ledwell, R. Takahashi, R. Weiss, L. Merlivst, L. Memery, P. Tsung-Hung, B. Jahne, and K.O. Munnich. 1986. Isotopic versus micrometerologic ocean CO2 fluxes: A serious conflict. Journal of Geophysical Research 91:10,517-10,527.
Brown, S., J. Sathaye, M. Cannell, and P. Kauppi. 1996. Management of forests for mitigation of greenhouse gas emissions. Pp. 773-797 in Climate Change 1995. Impacts, Adaptations and Mitigation of Climate Change: Scientific-Technical Analysis. Contribution of Working Group II to the Second Assessment Report of the Intergovernmental Panel on Climate Change, R.T. Watson, M.C. Zinyowera, and R.H. Moss, eds. Cambridge University Press, Cambridge, U.K.
Bruno, M., and F. Joos. 1997. Terrestrial carbon storage during the past 200 years: A Monte Carlo analysis of CO2 data from ice core and atmospheric measurements. Global Biogeochemical Cycles11:111-124.
Carleton, J.T. 1996. Pattern, process, and prediction in marine invasion ecology. Biological Conservation 78:97-106.
Carpenter, E.J., and D.G. Capone, eds. 1983. Nitrogen in the Environment. Academic Press, New York.
Carpenter, S.R., and J.F. Kitchell. 1993. The Trophic Cascade in Lakes. Cambridge University Press, Cambridge, U.K.
Carpenter, S.R., S.W. Chisholm, C.J. Krebs, D.W. Schindler, and R.F. Wright. 1995. Ecosystem experiments. Science 269:324-327.
Chameides, W.L., P.S. Kasibhatla, J. Yienger, and H. Levy II. 1994. Growth of continental-scale metro-agro-plexes, regional ozone pollution, and world food production. Science 264:74-77.
Chapin, F.S., III, G.R. Shaver, A.E. Giblin, K.G. Nadelhoffer, and J.A. Laundre 1995. Responses of Arctic tundra to experimental and observed changes in climate. Ecology 76:694-7116.
Ciais, P., P.P. Tans, M. Trolier, J.W.C. White, and R.J. Francey. 1995a. A large northern hemisphere terrestrial CO2 sink indicated by 13C/12C of atmospheric CO2. Science 269:1098-1102.
Ciais, P., P. Tans, J.W. White, M. Trolier, R. Francey, J. Berry, D. Randall, P. Sellers, J.G. Collatz, and D. Schimel. 1995b. Partitioning of ocean and land uptake of CO2 as inferred by d13C measurements from the NOAA/Climate Monitoring and Diagnostics Laboratory Global Air Sampling Network. Journal of Geophysical Research 100D:5051-5070.
Coale, K.H., K.S. Johnson, S.E. Fitzwater, R.M. Gordon, S. Tanner, F.P. Chavez, L. Feriolli, C. Sakamoto, P. Rogers, F. Millero, P. Steinberg, P. Nightingale, D. Cooper, W.P. Cochlan, M.R. Landry, J. Constaninou, G. Rollwagen, A. Trasvina, and R. Kudela. 1996. A massive phytoplankton bloom induced by an ecosystem-scale fertilization experiment in the equatorial Pacific Ocean. Nature 383:495-501.
Comins, H.N., and R.E. McMurtrie. 1993. Long-term response of nutrient-limited forests to CO2 enrichment: Equilibrium behavior of plant-soil models. Ecological Applications 3:666-681.
Crutzen, P.J., and M.O. Andreae. 1990. Biomass burning in the tropics: Impact on atmospheric chemistry and biogeochemical cycles. Science 250:1669-1678.
Dai, A., and I.Y. Fung. 1993. Can climate variability contribute to the “missing” CO2 sink? Global Biogeochemical Cycles 7:599-609.
D'Antonio, C.M., and P.M. Vitousek. 1992. Biological invasions by exotic grass, the grass-fire cycle, and global change. Annual Review of Ecology and Systematics 23:63-87.
Defries, R.S., and J.R.G. Townshend. 1994. NDVI-derived land-cover classifications at a global scale. International Journal of Remote Sensing 15:3567-3586.
Denning, A.S., I.Y. Fung, and D. Randall. 1995. Latitudinal gradient of atmospheric CO2 due to seasonal exchange with land biota. Nature 376:240-243.
Dickinson, R.E., and A. Henderson-Sellers. 1988. Modeling tropical deforestation: A study of GCM land-surface parameterizations Quarterly Journal of the Royal Meteorological Society 114:439-462.
Dixon, J.D.Kinsman, R.N. Sampson, and A.E. Lugo. 1993. Carbon dioxide sequestration in terrestrial ecosystems. Climate Research 3:1-5.
Dlugokencky, E.J., L.P. Steele, P.M. Lang, and K.A. Masarie. 1994. The growth rate and distribution of atmospheric methane. Journal of Geophysical Research 99:17,021-17,043.
Drake, B.G. 1992. A field study of the effects of elevated CO2 on ecosystem processes in a Chesapeake Bay wetland. Australian Journal of Biological Sciences 19:579-595.
Entekhabi, D., I. Rodriguez-Iturbe, and R.L. Bras. 1992. Variability in large-scale water balance with land surface-atmosphere interaction. Journal of Climate 5:798-813.
Everett, J.T., A. Krovnin, D. Lluch-Belda, E. Okemwa, H.A. Regier, and J.-P. Troadec. 1996. Fisheries. Pp. 511-537 in Climate Change 1995. Impacts, Adaptations and Mitigation of Climate Change: Scientific-Technical Analysis. Contribution of Working Group II to the Second Assessment Report of the Intergovernmental Panel on Climate Change, R.T. Watson, M.C. Zinyowera, and R.H. Moss, eds. Cambridge University Press, Cambridge, U.K.
Farquhar, G.D., J. Lloyd, J.A. Taylor, L.B. Flanagan, J.P. Syvertsen, K.T. Hubick, S.C. Wong, and J.R. Ehleringer. 1993. Vegetation effects on the isotope composition of oxygen in atmospheric CO2. Nature 363:439-442.
Field, C.B., and H.A. Mooney. 1986. The photosynthesis-nitrogen relationship in wild plants. Pp. 25-55 in On the Economy of Plant Form and Function, T.J. Givnish, ed. Cambridge University Press, Cambridge, U.K.
Flanagan, L.B., J.R. Brooks, G.T. Varney, and J.R. Ehleringer. 1997. Discrimination against C 18O16O during photosynthesis and the oxygen isotope ratio of respired CO2 in boreal forest ecosystems. Global Biogeochemical Cycles 11:83-98.
Foley, J.A., I.C. Prentice, N. Ramankutty, S. Levis, D. Pollard, S. Sitch, and A. Haxeltine. 1996. An integrated biosphere model of land surface processes, terrestrial carbon balance, and vegetation dynamics. Global Biogeochemical Cycles 10:603-628.
Friedlingstein, P., I. Fung, E. Holland, J. John, G. Brasseur, D. Erickson, and D. Schimel. 1995. On the contribution of CO2 fertilization to the missing biospheric sink. Global Biogeochemical Cycles9:541-556.
Fung, I.Y., C.J. Tucker, and K.C. Prentice. 1987. Application of advanced very high resolution radiometer vegetation index to study atmosphere-biosphere exchange of CO2. Journal of Geophysical Research 92(3):2999-3015.
Galloway, J.N., W.H. Schlesinger, H. Levy II, A. Michaels, and J.L. Schnoor. 1995. Nitrogen fixation: Anthropogenic enhancement-environmental response Global Biogeochemical Cycles 9:235-252.
Gifford, R.M., D.J. Barrett, J.L. Lutze, and A.B. Samarakoon. 1997. The CO2 fertilizing effect: Relevance to the global carbon cycle. In The Carbon Cycle, T.M.L. Wigley and D.S. Schimel, eds. Cambridge University Press, Stanford, Calif.
Global Change Biology Special Issue. 1996. Coral reefs and global change. Global Change Biology 2(6) (ISSN 1354-1013). Blackwell Science Ltd., Oxford, U.K.
Goulden, M.L., J.W. Munger, S.-M. Fan, B.C. Daube, and S.C. Wofsy. 1996. Exchange of carbon dioxide by a deciduous forest: Response to interannual climate variability. Science 271:1576-1578.
Groffman, P.M., and G.E. Likens, eds. 1994. Integrated Regional Models: Interactions Between Humans and the Environment. Chapman & Hall, New York.
Harden, J.W., E.T. Sundquist, R.F. Stallard, and R.K. Mark. 1992. Dynamics of soil carbon during deglaciation of the Laurentide ice sheet. Science 258:1921-1924.
Harriss, R.C., S.C. Wofsy, J.M. Hoell, Jr., R.J. Bendura, J.W. Drewry, R.J. McNeal, D. Pierce, V. Rabine, and R.L. Snell. 1994. The Arctic Boundary Layer Expedition (ABLE 3B): July-August 1990. Journal of Geophysical Research 99D:1635-1643.
Harte, J., and R. Shaw. 1995. Shifting dominance within a montane vegetation community: Results of a climate-warming experiment. Science 267:876-880.
Hein, R., P.J. Crutzen, and M. Heimann. 1997. An inverse modeling approach to investigate the global atmospheric methane cycle. Global Biogeochemical Cycles 11:43-76.
Henderson-Sellers, A. 1993. Continental vegetation as a dynamic component of a global climate model: A preliminary assessment. Climatic Change 23:337-377.
Hibbard, K.A., and L.K. Shaulis. 1997. UCAR coloquium on advances in ecosystem-atmosphere interactions. Bulletin of the Ecological Society of America 78:164.
Holland, E.A., A.R. Townsend, and P.M. Vitousek. 1995. Variability in temperature regulation of CO2 fluxes and N mineralization from five Hawaiian soils: Implication for a changing climate. Global Change Biology 1:115-123.
Holland, E.A., B.H. Braswell, J.-F. Lamarque, A. Townsend, J.M. Sulzman, J.-F. Müller, F. Dentener, G. Brasseur, H. Levy II, J.E. Penner, and G. Roelofs. 1997. Variations in the predicted spatial distribution of atmospheric nitrogen deposition and their impact on carbon uptake by terrestrial ecosystems Journal of Geophysical Research 102:15,849-15,866.
Houghton, R.A., J.E. Hobbie, J.M. Melillo, B. Moore III, B.J. Peterson, G.R. Shaver, and G.M. Woodwell. 1983. Changes in the carbon content of terrestrial biota and soils between 1860 and 1980: A net release of CO2 to the atmosphere. Ecological Monographs 53:235-262.
Howarth, R.W., G. Billen, D. Swaney, A. Townsend, N. Jaeorski, K. Lajtha, J.A. Downing, R. Elmgren, N. Caraco, T. Jordan, F. Berendse, J. Freney, V. Kudeyarov, P. Murdoch, and Z. Zhao-Liang. 1996. Regional nitrogen budgets and riverine N&P fluxes for the drainages to the North Atlantic Ocean: Natural and human influences. Biogeochemistry 35:75-139.
Hunt, E.R.J., S.C. Piper, R. Nemani, C.D. Keeling, R. Otto, and S.W. Running. 1996. Global net carbon exchange and intra-annual CO2 concentrations predicted by an ecosystem process model and a three-dimensional atmospheric transport model. Global Biogeochemical Cycles 10:431-456.
Hurst, D.F., P.S. Bakwin, R.C. Myers, and J.W. Elkins. 1997. Behavior of trace gas mixing ratios at a very tall tower site in North Carolina. Journal of Geophysical Research 102:8825-8835.
Idso, K.E., and S.B. Idos. 1994. Plant response to atmospheric CO2 enrichment in the face of environmental constriants: A review of the past 10 years of research. Agricultural Forestry and Meteorology 69(3-4):153-203.
International Geosphere-Biosphere Programme (IGBP) Scientific Advisory Council. 1994. Integrating Earth System Science: Climate, biogeochemistry, social science, policy. Ambio 23(1), Royal Swedish Academy of Sciences, Stokholm, Sweden.
IPCC. 1990. Climate Change: The IPCC Scientific Assessment. Cambridge University Press, Cambridge, U.K.
IPCC. 1992. Climate Change 1992: The Supplementary Report to the IPCC Scientific Assessment, J.T. Houghton, B.A. Callander and S.K. Varney (eds.), WMO/UNEP, Cambridge Unviersity Press, Cambridge, U.K. 200 pp.
IPCC. 1995. Climate Change 1994. Radiative Forcing of Climate Change and an Evaluation of the IPCC IS92 Emission Scenarios, Reports of Working Groups I and III of the Intergovernmental Panel on Climate Change, forming part of the IPCC Special Report to the first session of the Conference of Parties to the UN Framework Convention on Climate Change. Cambridge University Press, Cambridge, U.K.
IPCC. 1996. Climate Change 1995—The Science of Climate Change. Contribution of WGI to the Second Assessment Report of the Intergovernmental Panel on Climate Change. Cambridge University Press, Cambridge, U.K.
Jenkinson, D.S., D.E. Adams, and A. Wild. 1991. Model estimates of CO2 emissions from soil in response to global warming. Nature 351:304-306.
Jenkinson, D.S., J.M. Ports, J.N. Perry, V. Barnett, K. Coleman, and A.E. Johnston. 1994. Trends in herbage yields over the last century on the Rothamsted long-term continuous hay experiment. Journal of Agricultural Science 122:365-374.
Jones, C., and J. Lawton, eds. 1995. Linking Species and Ecosystems. Chapman and Hall, London.
Joos, F., J.L. Sarmiento, and U. Siegenthaler. 1991. Estimates of the effect of Southern Ocean iron fertilization on atmospheric CO2 concentrations. Nature 349:772-775.
Kareiva, P.M., J.G. Kingsolver, and R.B. Huey, eds. 1993. Biotic Interactions and Global Change. Sinauer Associates, Inc., Sunderland, Mass.
Keeling, R.F., and S.R. Shertz. 1992. Seasonal and interannual variations in atmospheric oxygen and implications for the global carbon cycle. Nature 358:723-727.
Keeling, C.D., S.C. Piper, and M. Heimann. 1989. A three-dimensional model of atmospheric CO2 transport based on observed winds: Mean annual gradients and interannual variations. Pp. 305-363 in Aspects of Climate Variability in the Pacific and Western Americas, D.H. Peterson, ed. American Geophysical Union, Washington, D.C.
Keeling, C.D., J.F.S. Chin, and T.P. Whorf. 1996. Increased activity of northern vegetation inferred from atmospheric CO2 measurements. Nature 382:146-149.
Keller, M., E. Veldkamp, A.M. Weltz, and W.A. Reiners. 1993. Effect of pasture age on soil tracegas emissions from a deforested area of Costa Rica. Nature 365:244-246.
Kittel T., N.A. Rosenbloom, T.H. Painter, D.S. Schimel, and VEMAP modeling participants. 1996. The VEMAP integrated database of modeling United States ecosystem/vegetation sensitivity to climate change. Journal of Biogeography 22:857-862.
Kleypas, J.A. 1997. Modeled estimates of global reef habitat and carbonate production since the last glacial maximum. Paleoceanography 12:533-545.
Krankina, O.N., and R.K. Dixon. 1994. Forest management options to conserve and sequester terrestrial carbon in the Russian Federation. World Resources Review 6:88-101.
Kutzbach, J., G. Bonan, and S.P. Harrison. 1996. Vegetation and soil feedbacks on the response of the African monsoon to orbital forcing in the early to middle Holocene. Nature 384:623-626.
Lawton, J.H. 1995. Ecological experiments with model systems. Science 269:328-331.
Lenschow, D.H. 1995. Micrometeorological techniques for measuring biosphere-atmosphere trace gas exchange. Pp. 126-163 in Biogenic Trace Gases: Measuring Emissions from Soil and Water, P.A. Matson and R.C. Harriss, eds. Blackwell Science, Cambridge, Mass.
Levin, S.A., ed. 1992. Mathematics and Biology: The Interface. Challenges and Opportunities. Cornell University Press, Ithaca, N.Y.
Liss, P., and L. Merlivat. 1986. 113-127 in The Role of Air-Sea Exchange in Geochemical Cycling, P. Buat-Menard, ed. Advanced Science Institute Series, Reidel, Hingham.
Lodge, D.M. 1993. Biological invasions: Lessons for ecology. Trends in Ecology and Evolution 8:133-137.
Luizão, F., P. Matson, G. Livingston, R. Luizão, and P. Vitousek. 1989. Nitrous oxide flux following tropical land clearing. Global Biogeochemical Cycles 3:281-285.
Martin, J.H. 1990. Glacial-interglacial 2 change: The iron hypothesis. Paleoceanography5:1-13.
Matson, P.A., and D.S. Ojima, eds. 1990. The Terrestrial Biosphere Exchange with Global Atmospheric Chemistry —Companion to the Dookie Report. IGBP Report 13. IGBP, Stockholm.
McKane, R.B., E.B. Rastetter, G.R. Shaver, K.J. Nadelhoffer, A.E. Giblin, J.A. Laundre, and F.S. Chapin III. 1997a. Climatic effects on tundra carbon storage inferred from experimental data and a model. Ecology 78:1170-1187.
McKane, R.B., E.B. Rastetter, G.R. Shaver, K.J. Nadelhoffer, A.E. Giblin, J.A. Laundre, and F.S. Chapin III. 1997b. Reconstruction and analysis of historical changes in Arctic tundra Ecology 78:1188-1198.
McNaughton, K.G., and P.G. Jarvis. 1991. Effects of spatial scale on stomatal control of transpiration. Agricultural and Forest Meteorology54:279-301.
Melillo, J.M., A.D. McGuire, D.W. Kicklighter, B. Moore III, C.J. Vorosmarty, and A.L. Schloss. 1993. Global climate change and terrestrial net primary production. Nature363:234-240.
Mooney, H.A., B.G. Drake, and R.J. Luxmoore. 1991. Predicting ecosystem responses to elevated CO2 concentrations. BioScience41:96-104.
Mosier, A., D. Schimel, D. Valentine, K. Bronson, and W. Parton. 1991. Methane and nitrous oxide fluxes in native, fertilized and cultivated grasslands. Nature 350:330-332.
Mosier, A.R., W.J. Parton, D.W. Valentine, D.S. Ojima, D.S. Schimel, and J.A. Delgado. 1996. CH4 and N 2O fluxes in the Colorado shortgrass steppe: 1. Impact of landscape and nitrogen addition. Global Biogeochemical Cycles 10:387-399.
Mosier, A.R., W.J. Parton, D.W. Valentine, D.S. Ojima, D.S. Schimel, and O. Heinemeyer. 1997. CH4 and N 2O fluxes in the Colorado shortgrass steppe 2. Long-term impact of land-use change. Global Biogeochemical Cycles 11:29-42.
Myneni, R.B., S.O. Los, and C.J. Tucker. 1996. Satellite-based identification of linked vegetation index and sea surface temperature anomaly areas from 1982-1990 for Africa, Australia and South America. Geophysical Research Letters 23:729-732.
Myneni, R.B., R.R. Nemani, and S.W. Running. 1997. Algorithm for the estimation of global land cover, LAI and FAPAR based on radiative transfer models. IEEE Transactions in Geoscience. Remote Sensing (submitted).
Najjar, R.G. 1992. Marine biogeochemistry. Pp. 241-280 in Climate System Modeling, K.E. Trenberth, ed. Cambridge University Press, New York.
National Research Council (NRC). 1986. Global Change in the Geosphere-Biosphere: Initial Priorities for an IGBP. National Academy Press, Washington, D.C.
National Research Council (NRC). 1994. The Role of Terrestrial Ecosystems in Global Change: A Plan for Action. National Academy Press, Washington, D.C.
National Research Council (NRC). 1997. News Report XLVII. Winter.
Niemela, P., and W.J. Mattson. 1996. Invasion of North American forests by European phytophagus insects —legacy of the European crucible. BioScience 46:741-753.
Oechel, W.C., S.J. Hastings, G. Vourlitis, M. Jenkins, G. Riechers, and N. Grulke. 1993. Recent change of Arctic tundra ecosystems from a net carbon dioxide sink to a source. Nature 361:520-523.
Oechel, W.C., S. Cowies, N. Grulke, S.J. Hastings, B. Lawrence, T. Prudhomme, G. Riechers, B. Strain, D. Tissue, and G. Vourlitis. 1994. Transient nature of CO2 fertilization in Arctic tundra. Nature 371:500-503.
Ojima, D.S., and B.H. Svensson, eds. 1992. Trace Gas Exchange in a Global Perspective. Ecological Bulletins 42, Munksgaard International Booksellers and Publishers, Copenhagen, Denmark.
Pastor, J., and W.M. Post. 1986. Influence of climate, soil moisture and succession on forest carbon and nitrogen cycles. Biogeochemistry 2:3-27.
Pastor, J., and W.M. Post. 1993. Linear regressions do not predict the transient responses of eastern North American forests to CO2-induced climate change. Climatic Change 23:111-119.
Penner J.E., C.S. Atherton, J. Dingnon, S.J. Ghan, J.J. Walton, and S. Hameed. 1991. Tropospheric nitrogen: A three-dimensional study of sources, distributions, and deposition. Journal of Geophysical Research 96:959-990.
Peterson, B.J., and J.M. Melillo. 1985. The potential storage of carbon caused by eutrophication of the biosphere Tellus 37B:117-127.
Pielke, R.A., and R. Avissar. 1990. Influence of landscape structure on local and regional climate. Landscape Ecology 4:133-135.
Pielke, R.A., W.R. Cotton, R.L. Walko, C.J. Tremback, W.A. Lyons, L.D. Grasso, M.E. Nicholls, M.D. Moran, D.A. Wesley, T.J. Lee, and J.H. Copeland. 1992. A comprehensive meteorological modeling system—RAMS. Meteorology and Atmospheric Physics 49:69-91.
Pielke, R.A., T.J. Lee, J.H. Copeland, J.L. Eastman, C.L. Ziegler, and C.A. Finley. 1997. Use of USGS-provided data to improve weather and climate simulations Ecological Applications 7:3-21.
Post, W.M., A.W. King, and S.D. Wullschleger. 1997. Historical variations in terrestrial biospheric carbon storage. Global Biogeochemical Cycles 11:99-109.
Potter, C.S., J.T. Randerson, C.B. Field, P.A. Matson, P.M. Vitousek, H.A. Mooney, and S.A. Klooster. 1993. Terrestrial ecosystem production: A process model based on global satellite and surface data. Global Biogeochemical Cycles 7:811-841.
Raich, J.W., and W.H. Schlesinger. 1992. The global carbon dioxide flux in soil respiration and its relationship to vegetation and climate. Tellus 44:81-99.
Randerson, J.T., M.V. Thompson, I.Y. Fung, and C.B. Field. 1997. The contribution of terrestrial sources and sinks to trends in the seasonal cycle of atmospheric carbon dioxide. Global Biogeochemical Cycles 11(4):535-560.
Rejmanek, M., and J. Randall. 1994. Invasive alien plants in California: 1993 Summary and comparison with other areas in North America. Madrono 41:161-177.
Rosenzweig, C., and M.L. Parry. 1994. Potential impact of climate change on world food supply. Nature 367:133-138.
Rosswall, T., R.G. Woodmansee, and P.G. Risser, eds. 1988. Scales and Global Change. John Wiley & Sons, New York.
Sarmiento, J.L. 1991. Slowing the buildup of fossil CO2 in the atmosphere by iron fertilization: A comment. Global Biogeochemical Cycles 5:1-2.
Sarmiento, J.L., and C. Le Quéré. 1996. Oceanic carbon dioxide uptake in a model of century-scale global warming. Science 274:1346-1350.
Sarmiento, J.L., C. Le Quéré, and S.W. Pacala. 1995. Limiting future atmospheric carbon dioxide. Global Biogeochemical Cycles 9:121-137.
Schimel, D.S. 1995. Terrestrial ecosystems and the carbon cycle. Global Change Biology 1:77-91.
Schimel, D.S., W.J. Parton, T.G.F. Kittel, D.S. Ojima, and C.V. Cole. 1990. Grassland biogeochemistry: Links to atmospheric processes. Climatic Change 17:13-25.
Schimel, D.S., T.G.F. Kittel, A.K. Knapp, T.R. Seastedt, W.J. Parton, and V.B. Brown. 1991. Physiological interactions along resource gradients in a tallgrass prairie. Ecology 72:672-684.
Schimel, D.S., B.H. Braswell, Jr., E.A. Holland, R. McKeown, D.S. Ojima, T.H. Painter, W.J. Parton, and A.R. Townsend. 1994. Climatic, edaphic, and biotic controls over carbon and turnover of carbon in soils. Global Biogeochemical Cycles 8:279-293.
Schimel, D., I.G. Enting, M. Heimann, T.M.L. Wigley, D. Raynaud, D. Alves, and U. Siegenthaler. 1995. CO2 and the carbon cycle. Pp. 35-71 in Climate Change 1994: Radiative Forcing of Climate Change and an Evaluation of the IPCC IS92 Emission Scenarios, J.T. Houghton, L.G. Meira Filho, J. Bruce, H. Lee, B.A. Callander, E. Haites, N. Harris, and K. Maskel, eds. Cambridge University Press, Cambridge, U.K.
Schimel, D.S., B.H. Braswell, R. McKeown, D.S. Ojima, W.J. Parton, and W. Pulliam. 1996. Climate and nitrogen controls on the geography and time scales of terrestrial biogeochemical cycling. Global Biogeochemical Cycles 10:677-692.
Schimel, D.S., VEMAP Participants, and B.H. Braswell. 1997a. Spatial variability in ecosystem processes at the continental scale: Models, data, and the role of disturbance. Ecological Monographs 67:251-271.
Schimel, D.S., M. Grubb, F. Joos, R. Kaufmann, R.H. Moss, W. Ogana, R. Richels, and T.M.L. Wigley. 1997b. Stabilisation of Atmospheric Greenhouse Gases: Physical, Biological and Socioeconomic Implications. IPCC Technical Paper 3. WMO/UNEP, Geneva.
Schindler, D.W., and S.E. Bayley. 1993. The biosphere as an increasing sink for atmospheric carbon: Estimates from increased nitrogen deposition. Global Biogeochemical Cycles 7:717-734.
Schulze, E.D., W. De Vries, M. Hauhs, K. Rosén, L. Rasmussen, O.-C. Tann, and J. Nilsson. 1989. Critical loads for nitrogen deposition in forest ecosystems. Water, Air, and Soil Pollution 48:451-456.
Sellers, P., and D. Schimel. 1993. Remote sensing of the land biosphere and biogeochemistry in the EOS era: Science priorities, methods and implementation—EOS land biosphere and biogeochemical cycles panels. Global and Planetary Change 7:279-297.
Sellers, P.J., J.A. Berry, G.J. Collatz, C.B. Field, and F.G. Hall. 1992. Canopy reflectance, photosynthesis, and transpiration. III. A reanalysis using improved leaf models and a new canopy integration scheme. Remote Sensing of the Environment 42:187-216.
Sellers, P., F. Hall, H. Margolis, B. Kelly, D. Baldocchi, G. den Hartog, J. Cihlar, M.G. Ryan, B. Goodison, P. Crill, K.J. Ranson, D. Lettenmaier, and D.E. Wickland. 1995. The Boreal Ecosystem-Atmosphere Study (BOREAS): An overview and early results from the 1994 field year. Bulletin of the American Meteorological Society 76:1549-1577.
Sellers, P.J., D.A. Randall, G.J. Collatz, J.A. Berry, C.B. Field, D.A. Dazlich, C. Zhang, G.D. Collelo, and L. Bounoua. 1996a. A revised land surface parameterization (SiB2) for atmospheric GCMs. Part I: Model formulation. Journal of Climate 9:676-705.
Sellers, P.J., S.O. Los, C.J. Tucker, C.O. Justice, D.A. Dazlich, G.J. Collatz, and D.A. Randall. 1996b. A revised land surface parameterization (SiB2) for atmospheric GCMs. Part II: The generation of global fields of terrestrial biophysical parameters from satellite data. Journal of Climate 9:706-737.
Sellers, P.J., B.W. Meeson, J. Closs, J. Collatz, F. Corprew, D. Dazlich, F.G. Hall, Y. Kerr, R. Koster, S. Los, K. Mitchell, J. McManus, D. Myers, K.-J. Sun, and P. Try. 1996c. The ISLSCP initiative I global datasets: Surface boundary conditions and atmospheric forcings for land-atmosphere studies. Bulletin of the American Meteorological Society 77:1987-2005.
Sellers, P.J., R.E. Dickinson, D.A. Randall, A.K. Betts, F.G. Hall, J.A. Berry, G.J. Collatz, A.S. Denning, H.A. Mooney, C.A. Nobre, N. Sato, C.B. Field, and A. Henderson-Sellers. 1997. Modeling the exchanges of energy, water, and carbon between continents and the atmosphere. Science 275:502-509.
Shugart, H.H., P.J. Michaels, T.M. Smith, D.A. Weinstein, and E.B. Rastetter. 1988. Simulation models of forest succession. Pp. 125-151 in Scales and Global Change T. Rosswall, R.G. Woodmansee, and P.G. Risser, eds. John Wiley & Sons, New York.
Shukla, J., C. Nobre, and P. Sellers. 1990. Amazon deforestation and climate change. Science 247:1322-1325.
Siegenthaler, U., and J.L. Sarmiento. 1993. Atmospheric carbon dioxide and the ocean. Nature 365:119-125.
Skole, D., and C. Tucker. 1993. Tropical deforestation and habitat fragmentation in the Amazon: Satellite data from 1978 to 1988. Science 260:1905-1910.
Smith, T.M., and H.H. Shugart. 1993. The transient response of terrestrial carbon storage to a perturbed climate. Nature 361:523-526.
Tans, P.P., I.Y. Fung, and T. Takahashi. 1990. Observational constraints on the global atmospheric CO2 budget. Science 247:1431-1438.
Takahashi, T., et al. 1986. Seasonal and Geographic Variability of Carbon Dioxide Sink/Source in the Oceanic Areas. Technical Report for Contract MRETTA 19X-89675C. Lamont-Doherty Goelogical Observatory, Palisades, N.Y.
Thompson, M.V., J.T. Randerson, C.M. Malmström, and C.B. Field. 1996. Change in net primary production and heterotrophic respiration: How much is necessary to sustain the terrestrial carbon sink? Global Biogeochemical Cycles 10:711-726.
Tilman, D., and J.A. Downing. 1994. Biodiversity and stability in grasslands. Nature 367:363-365.
Tilman, D., D. Wedin, and J. Knops. 1996. Productivity and sustainability influenced by biodiversity in grassland ecosystems. Nature 379:718-720.
Towne, G., and C.E. Owensby. 1984. Long-term effects of annual burning at different dates in ungrazed Kansas tallgrass prairie. Journal of Range Management 37:392-397.
Townsend, A.R., P.M. Vitousek, and E.A. Holland. 1992. Tropical soils could dominate the shortterm carbon cycle feedbacks to increased global temperatures. Climatic Change 22:293-303.
Townsend, A.R., P.M. Vitousek, and S.E. Trumbore. 1995. Soil organic matter dynamics along gradients in temperature and land use on the island of Hawaii. Ecology 76:721-733.
Townsend, A.R., B.H. Braswell, E.A. Holland, and J.E. Penner. 1996. Spatial and temporal patterns in terrestrial carbon storage due to deposition of anthropogenic nitrogen. Ecological Applications 6:806-814.
Turner II, B.L., D. Skole, S. Sanderson, G. Fischer, L. Fresco, and R. Leemans. 1995. Land-Use and Land-Cover Change (LUCC): Science/Research Plan. IGBP, Stockholm.
VEMAP Participants. 1995. Vegetation/ecosystem modeling and analysis project (VEMAP): Comparing biogeography and biogeochemistry models in a continental-scale study of terrestrial ecosystem responses to climate change and CO2 doubling. Global Biogeochemical Cycles 9:407-438.
Verma, S.B., P.J. Sellers, C.L. Walthall, F.G. Hall, J. Kim, and S.J. Goetz. 1993. Photosynthesis and stomatal conductance related to reflectance on the canopy scale. Remote Sensing of the Environment 44:103-116.
Vitousek, P.M., P.R. Ehrlich, A.H. Ehrlich, and P.A. Matson. 1986. Human appropriation of the products of photosynthesis. BioScience 36:368-373.
Vitousek, P.M., C.M. D'Antonio, L. Loope, and R. Westbrooks. 1996. Biological invasions as global environmental change. American Scientist 84:468-478.
Waelbroeck, C., P. Monfray, W.C. Oechel, S. Hastings, and G. Vourlitis. 1997. The impact of permafrost thawing on the carbon dynamics of tundra Geophysical Research Letters 24:229-232.
Walker, D.A., P.J. Webber, E.F. Binnian, K.R. Everett, N.D. Lederer, E.A. Nordstrand, and M.D. Walker. 1987. Cumulative impacts of oil fields on northern Alaska's landscapes. Science 238:757-761.
Watson, R.T., M.C. Zinyowera, and R.H. Moss, eds. 1996. Technologies, Policies and Measures for Mitigating Climate Change. WMO/UNEP, Geneva.
Wedin, D.A., and D. Tilman. 1996. Influence of nitrogen loading and species composition on the carbon balance of grasslands. Science 274:1720-1723.
Wessman, C.S., J.D. Aber, D.L. Peterson, and J.M. Melillo. 1988. Remote sensing of canopy chemistry and nitrogen cycling in tenperate forest ecosystems. Nature 335(6186):154-157.
Wisniewski, J., and R.N. Sampson, eds. 1993. Terrestrial biospheric carbon fluxes: Quantification of sinks and sources of CO2. Proceedings of workshop held in Harzburg, Germany, March 1993. Kluwer Academic, Dordrecht; Boston.
Wofsy, S.C., M.L. Goulden, J.W. Munger, S.-M. Fan, P.S. Bakwin, B.C. Danbe, S.L. Bassow, and F.A. Bazzaz. 1993. Net exchange of CO2 in a mid-latitude forest. Science 260:1314-1317.
Woodward, F.I., and W.L. Steffen eds. 1997. Natural Disturbances and Human Land Use in Dynamic Global Vegetation Models. Report of a workshop co-convened by the GAIM, GCTE, LUCC, and IGBP-DIS Programme Elements of the IGBP. IGBP, Stockholm.
Woodwell, G.M. 1989. The warming of the industrialized middle latitudes 1985-2050: Causes and consequences. Climatic Change 15:31-50.
Woodwell, G.M., and F.T. Mackenzie, eds. 1995. Biotic Feedbacks in the Global Climatic System: Will the Warming Feed the Warming? Oxford University Press, New York.




































































