
This paper was presented at a colloquium entitled “Neuroimaging of Human Brain Function,” organized by Michael Posner and Marcus E.Raichle, held May 29–31, 1997, sponsored by the National Academy of Sciences at the Arnold and Mabel Beckman Center in Irvine, CA.
Neuroimaging studies of word reading
JULIE A.FIEZ*† AND STEVEN E.PETERSEN‡
*Department of Psychology, University of Pittsburgh and the Center for the Neural Basis of Cognition, University of Pittsburgh/Carnegie Mellon University, Pittsburgh, PA 15260; and ‡Departments of Neurology and Neurological Surgery, Anatomy and Neurobiology, Radiology, Washington University School of Medicine, St. Louis, MO 63110
ABSTRACT This review discusses how neuroimaging can contribute to our understanding of a fundamental aspect of skilled reading: the ability to pronounce a visually presented word. One contribution of neuroimaging is that it provides a tool for localizing brain regions that are active during word reading. To assess the extent to which similar results are obtained across studies, a quantitative review of nine neuroimaging investigations of word reading was conducted. Across these studies, the results converge to reveal a set of areas active during word reading, including left-lateralized regions in occipital and occipitotemporal cortex, the left frontal operculum, bilateral regions within the cerebellum, primary motor cortex, and the superior and middle temporal cortex, and medial regions in the supplementary motor area and anterior cingulate. Beyond localization, the challenge is to use neuroimaging as a tool for understanding how reading is accomplished. Central to this challenge will be the integration of neuroimaging results with information from other methodologies. To illustrate this point, this review will highlight the importance of spelling-to-sound consistency in the transformation from orthographic (word form) to phonological (word sound) representations, and then explore results from three neuroimaging studies in which the spelling-to-sound consistency of the stimuli was deliberately varied. Emphasis is placed on the pattern of activation observed within the left frontal cortex, because the results provide an example of the issues and benefits involved in relating neuroimaging results to behavioral results in normal and brain damaged subjects, and to theoretical models of reading.
Reading is one of the most important skills we learn, and not surprisingly it has been the focus of many investigations across a range of methodologies, including behavioral studies in normal subjects, computational modeling, and neuropsychological assessment of patients with acquired brain damage. This review discusses the contributions that neuroimaging can make to this interdisciplinary effort, with a focus on one aspect of skilled reading: the ability to pronounce a visually presented word.
In the most general sense, neuroimaging provides a tool for localizing and measuring the activity of brain regions that are recruited during the performance of a cognitive task in normal subjects. The value of this contribution will be explored in the context of two questions. The first is whether the method is reliable. In other words, if the same task is studied by two different investigators, do they localize the same brain regions? Clearly, the utility of a method in which the results fail to converge across studies must be seriously questioned. The first section addresses this issue by evaluating the regions of activation reported across nine neuroimaging studies of word reading.
However, just being reliable does not make a method worthwhile. A second question is whether the information gained from neuroimaging yields important insights into cognition and the brain. In terms of reading, if the goal is limited to localizing brain areas, then most would agree that neuroimaging offers the opportunity to refine the basic answer to this question that has been gained through studies of brain-damaged subjects (1), but the value of this objective might be questioned. A far more interesting challenge is to use neuroimaging to provide novel insights about how reading is accomplished. It is possible that the neuronal processing that supports reading is diffusely and equivalently distributed across multiple areas (e.g., throughout the perisylvian cortex). If this turns out to be the case, neuroimaging will be of limited value in understanding the cognitive processes involved in reading. Another possibility is that the areas active during reading are not all equivalent—i.e., not only is each area topographically specific, it has a unique and specific set of information inputs, it does specific types of transformations on this information, and it provides unique and specific types of output to other areas. Consequently, by understanding the effects of various experimental manipulations on areas of functional brain activation, we might hope to ultimately understand not only which brain regions contribute to reading, but how they do so.
In the broadest sense, reading presumably entails basic sensory and motor components, as well as more central components, such the analysis of visual word forms (orthography), the analysis of word sounds (phonology), and the analysis of word meaning (semantics). As a first step toward understanding whether the brain regions active during reading make specific contributions to these different processes, the results obtained by using baseline vs. sensorimotor control conditions, and reading aloud vs. reading silently as task conditions, will be reviewed. Although important, such analyses permit only rough fractionations of the regions involved in reading. To go beyond using neuroimaging for merely localizing broadly defined functions to broadly defined regions, other experimental approaches are necessary. In the latter portion of this review, the utility of one such approach, manipulations of stimulus type, will be evaluated. Specifically, the behavioral importance of spelling-to-sound consistency will be discussed, and results from three neuroimaging studies that manipulated the spelling-to-sound consistency of the stimuli will be presented. In interpreting the results from these studies, the focus will be on the pattern of activation observed
© 1998 by The National Academy of Sciences 0027–8424/98/95914–8$2.00/0
PNAS is available online at http://www.pnas.org.
|
|
Abbreviations: BA, Brodmann area; SMA, supplementary motor area; PET, positron emission tomography. |
|
† |
To whom reprint requests should be addressed at: 605 LRDC, 3939 O’Hara Street, Department of Psychology, University of Pittsburgh, Pittsburgh, PA 15260. e-mail: fiez+@pitt.edu. |
in a left frontal region. Rather than providing a broad review of the suspected functions of each area active during word reading, the goal will be to use the left frontal region as a “case study” to illustrate the challenges and benefits that may arise from attempts to relate neuroimaging results to those obtained by using other methodologies.
Do Results Converge Across Studies?
As noted above, one of the most obvious uses of neuroimaging is that it provides tools for localizing brain regions that are active during the performance of a cognitive task. But how good are these tools, and what has been learned so far about the location of brain regions that are active during reading? To address this question, results were reviewed from nine studies in which subjects read aloud single words.§ In six of the studies, word reading was compared with a passive control condition in which subjects either rested with their eyes closed (2, 3), maintained fixation on a crosshair (refs. 4 and 5; J.A.F., D.A. Balota, M.E.Raichle, and S.E.P., unpublished data), or viewed meaningless line drawings (6). In the remaining three studies, word reading was compared with a sensorimotor control condition in which subjects produced an utterance (e.g., “hiya”) in response to letter-like “false font” strings (7, 8) or consonant strings (9).
In evaluating these nine studies, the first issue to be confronted is that the results vary. For instance, the number of significant changes found in any given study ranges from 2 (8) to 32 (6), and within an anatomical region the locations of activation vary (e.g., peak changes that localize to primary motor cortex are separated by as much as 14 mm). This variability arises from many sources, including: (i) limitations of neuroimaging technology, especially the fact that in both positron emission tomography (PET) and functional magnetic resonance imaging (fMRI) the ratio of the signal to the noise is low. (ii) differences in the procedures used to analyze neuroimaging data, and (iii) anatomical and cognitive differences between subjects. An additional and potentially avoidable source of variability across studies is task-related. For instance, in all nine of the studies described above subjects read words, but many features of this task varied (presentation rate, stimuli, etc.), as did the control task to which reading was compared (as described above). Because the task comparisons thus are only approximately the same, there is no reason to expect the results to be more than approximately the same. In fact, as will be discussed below, such variations can be exploited to understand the specific contributions of each region involved in reading.
Although a full account of the functional neuroanatomy of reading will require an understanding of how the results are affected by each source of variability, it is also useful to understand what, if anything, is common across studies. To get at this question, a quantitative review of the nine studies of word reading was conducted. On average, in each study 16 significant activations were reported as coordinate locations (foci) in the Talairach and Tournoux brain atlas (10). yielding a total of 147 foci.¶ Of these 147 foci, 104 were determined to represent a commonly found activation.∥ These foci fell into clusters,** as summarized in Table 1 and Fig. 1. Left-lateralized groups of foci were found in the ventral occipitotemporal cortex in the fusiform and lingual gyri (near BA 18 and 37), and in left frontal operculum (near BAs 44, 45, and the anterior insula). Given that language functions generally are viewed as strongly left-lateralized, it is interesting to note that bilateral activation was observed in many areas, including anterior and posterior portions of the superior temporal gyrus (near BA 22), dorsal and more ventral portions of the postcentral gyrus (near BA 4). the region near the basal ganglia and insular cortex, and the cerebellar hemispheres. Medially, activation was found in the medial frontal gyrus [at or near the supplementary motor area (SMA)], the anterior cingulate gyrus (near BA 32), and the cerebellum. These results are broadly consistent with neuroanatomical expectations derived from the neurological literature (e.g., see ref. 1).
Overall, the outcome of this quantitative review is encouraging, because it demonstrates that neuroimaging results can and do converge across studies to identify brain areas that are active during reading. However, several cautionary points should be noted. First, the foci within each cluster were in some cases separated by as much as 20 mm. This raises serious questions about whether each identified region should be treated as a single functional area. Second, many foci did not meet the criteria for common activation, but this does not mean they should be dismissed as noise. By emphasizing regions of common activation, other important areas that are detected less consistently, or that are very susceptible to task-related differences, may have been overlooked.
Fractionating Regions and Processes: Making Broad Cuts
As discussed in the Introduction, the goal of neuroimaging extends beyond localization. A more interesting challenge is to use neuroimaging to understand how reading is accomplished by using a set of localized areas distributed throughout the brain. In this section, the focus will be on task and control comparisons that can help illuminate which of the regions active during reading are involved in more basic sensory and motor functions, and which may be specialized for more central components of reading, such as the orthographic, phonological, and semantic processing.
Two different general classes of control conditions were used in the nine studies reviewed above: (i) baseline conditions in which reading was compared with a simple visual task (e.g., fixate on a crosshair) or to a rest task (e.g., “close your eyes and empty your mind”), and (ii) sensorimotor control conditions in which subjects viewed nonlinguistic but comparably complex visual stimulus (e.g., strings of “false fonts”) and also produced an articulatory output for each trial. Evaluation of Table 1 reveals several regions of activation that were almost always found when a baseline control task was used, but not when a
|
§ |
All neuroimaging studies of reading that met the following criteria were included in this analysis: (i) subjects read aloud a series of individually displayed words, (ii) data were compared with a relatively passive or sensorimotor control condition (see text for further description), (iii) the majority of the brain was imaged, and (iv) regions of activation were reported as coordinate locations (foci) in terms of the Talairach and Tournoux atlas space (10). In all nine studies that met these criteria, the data were acquired by using PET. |
|
¶ |
In several of the studies word reading was compared with different control conditions, or different task variations were compared with the same control condition. In such studies, activation of the same region often was found in multiple comparisons. As an alternative to dealing with each focus separately, or arbitrarily selecting only one comparison to evaluate, similarly located foci within an individual study were averaged together. |
|
∥ |
A focus from any given study was judged to represent commonly found activation if it was near (within 20 mm) foci from a majority of studies. In other words, each focus had to be near a focus from at least four of the other eight studies, or near a focus from at least three of the six studies that used a passive control condition (to avoid excluding areas involved in visual processing and speech production). The 20-mm distance criterion was chosen because it is near the resolution of most analyzed PET images, and it is beyond the range of the typical response variability found between subjects and studies. |
|
** |
Foci were assigned to clusters based on the Brodmann area (BA)/gyral location to which each focus plotted. In some cases, more than one focus from a given study fell within the same Brodmann region. When this tendency appeared across studies, the foci were divided into two clusters (e.g., in many of the studies both a dorsal and a more ventral focus were identified within BA 4). Finally, the mean location of the foci falling within each cluster was computed. |
Table 1. Locations of positive foci across studies
|
Brodmann area |
Mean location |
Petersen (5) |
Bookheimer (7) |
Price (2) |
Price (3) |
Fiez (unpublished data) |
Rumsey (4) |
Price (7) |
Howard (8) |
Herbster (9) |
|
L 44/45/ins |
–41±9, 8±5, 6±4 |
|
X |
XX |
X |
XX |
X |
|
|
X |
|
M 6(SMA) |
1±3, 0±4, 57±4 |
X |
X |
X |
X |
X |
|
|||
|
L 32/24 |
–8±6, 2±4, 40±3 |
|
X |
|
X |
X |
X |
X |
||
|
L4 |
–46±2, –12±1, 37±5 |
X |
X |
X |
X |
X |
|
|||
|
L 4/43 |
–50±5, –6±4, 22±2 |
X |
X |
X |
X |
|
X |
|
||
|
R4 |
44±4, –10±3, 35±3 |
|
X |
|
X |
X |
|
|||
|
R 4/43 |
46±5, –8±9, 21±6 |
X |
X |
X |
X |
X |
|
|||
|
L In/thal/ins |
–24±5, –15±6, 10±7 |
X |
X |
XX |
X |
X |
X |
X |
|
|
|
R put/thal/ins |
7±7, –28±5, 7±7 |
|
XX |
X |
X |
|
X |
|
||
|
L 22/42/41 |
–55±4, –14±9, 4±4 |
|
X |
|
X |
|
X |
|
XX |
|
|
L 22/21 |
–54±5, –41±6, 12±7 |
|
XX |
X |
XX |
XX |
|
X |
X |
|
|
R 22/42/41 |
54±3, –14±4, 8±3 |
X |
X |
|
X |
|
XX |
|
||
|
R 22/21 |
51±4, –33±5, 4±4 |
|
X |
X |
X |
X |
X |
|
||
|
L 37/19 |
–36±6, –57±9, –5±3 |
X |
X |
|
X |
XX |
|
|||
|
L 18/19 |
–16±10, –82±8, –6±6 |
|
X |
|
XX |
|
X |
X |
|
|
|
L cerebellum |
–17±10, –63±5, –17±6 |
|
XX |
|
X |
XX |
|
X |
||
|
R cerebellum |
19±4, –66±11, –17±6 |
X |
X |
|
X |
X |
X |
|
X |
|
|
cblm, midline |
5±4, –58±11, –14±6 |
X |
X |
X |
X |
XX |
X |
|
||
|
The mean location (x±SD, y±SD, z±SD) for each foci cluster is given in terms of the Talairach and Tournoux (10) atlas. Each “X” indicates a focus that was found in the given cluster in the study indicated by the appropriate column heading. The studies by Price et al. (7), Howard et al. (8), and Herbster et al. (9) used a sensorimotor control condition. |
||||||||||
sensorimotor control task was used: the supplementary motor area (BA 6), regions in primary motor cortex (BA 4) bilaterally, the medial cerebellum, and the right superior temporal gyrus (BA 22/21). It is likely that these regions were “subtracted out,” because they are related to the auditory stimulation provided by the subject’s own voice (right BA 22) or motoric aspects of speech production (BA 4, SMA, and the medial cerebellum). One region, the anterior cingulate (BA 32), showed the converse pattern; given the proximity of the cingulate and SMA foci, it seems likely that both are commonly active during reading, but they are poorly resolved as separate foci unless the cingulate activation is “unmasked” by subtracting out the SMA activation. For the remaining regions, the story is less clear because significant activation was found in only one or two of the studies that employed a sensorimotor control condition (left thalamus/putamen, left BA 22/42, left BA 22/21, left BA 18, lateral cerebellum bilaterally), or the lack of significant activation with a sensorimotor control condition is complicated by the more sporadic activation of the region with a baseline control task (left BA 37/19, right thalamus/putamen).
Some clarification of the superior temporal activation comes from three studies that included both reading silently and reading aloud tasks, relative to a common baseline control condition (2, 3, 6). If the activation of these regions is solely related to auditory stimulation provided by the subject’s own voice, then activation should only be found in the reading aloud conditions. In support of this conclusion, significant activation in the right superior temporal cortex was not found during the silent reading conditions, with the exception of one focus near right BA 22/21. In contrast, in all three studies significant activation was found in the left superior temporal cortex during silent reading, though the location varied. In all cases, the z-scores for activations identified during reading silently were less than those identified during reading aloud. It is possible that one or more regions in the left superior/middle temporal cortex support acoustic/phonological representations that are necessary for silent reading, or that are activated
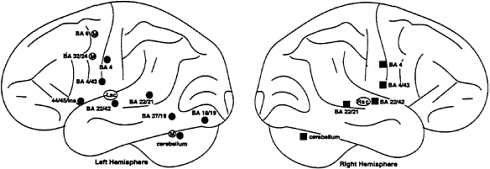
FIG. 1. Schematic diagram showing mean location of positive foci across studies of word reading (see Table 1). Foci in the right hemisphere are shown with filled squares, foci in the left hemisphere are shown with filled circles, three foci near the midline are shown in the left hemisphere with an “M,” and two subcortical foci near the thalamus, basal ganglia, and insular cortex are shown with “Lsc” and “Rsc.” It should be noted that these are only mean locations, that the distribution of locations in some cases covered a fairly wide range (as indicated by the standard deviations listed in Table 1). and that each mean location does not necessarily signify the center of a single functional area.
as a more general consequence of covert speech (e.g., in models of working memory, silent articulation is thought to generate acoustically based phonological representations of verbal material; refs. 11 and 12). Other interpretations can also be formed, such as the possibility that the superior/middle temporal cortex contributes to semantic analysis associated with both reading silently and reading aloud. And of course, as noted above, there is no reason to believe that the broad regions identified in this review represent single functional areas, and thus multiple interpretations of the results may be correct.
One point illustrated by these comparisons is that even at basic levels of analysis, interesting and unexpected questions for further research can be identified. For instance, the cerebellum is commonly thought to contribute only to motoric aspects of speech production, but only the medial portion of the cerebellum consistently showed the pattern of results expected on the basis of this assumption. Also, even though bilateral activation was observed in superior and middle temporal regions, hemispheric differences were observed in these regions for both the baseline vs. sensorimotor control comparisons, and for the reading silently vs. reading aloud comparison. These findings are at odds with the assumption that similarly located right and left hemisphere regions are functionally homologous, and suggests several hypotheses for further investigation (e.g., the assumption of homologous regions is invalid, the larger spatial extent of the left BA 22/21 region encompasses a functional subregion not activated during reading in the right hemisphere, etc.).
However, it can also be argued that much of what is learned through comparisons such as those discussed above is “old news.” For instance, there is a wealth of other evidence to indicate that primary motor cortex (BA 4), SMA, and the medial cerebellum contribute to motoric aspects of speech production, whereas regions such as Broca’s area (BA 44/45) and Wernicke’s area (21) contribute to more central aspects of linguistic analysis. What is needed are experimental manipulations that can provide insight into how the information that supports reading is represented and what types of transformations occur between representations. There are a variety of experimental approaches that can be used to investigate the cognitive processes involved in reading (e.g., reading can be compared with other related tasks, such as picture naming), and converging evidence from a range of language-related neuroimaging studies can be marshaled to support the following possible functional specializations: (i) primary motor cortex (BA 4), supplementary motor cortex (SMA/BA 6), and the medial cerebellum are involved in motoric aspects of speech production, (ii) anterior superior temporal regions (BA 22/41) are activated bilaterally by the auditory stimulation provided by one’s own vocalizations, (iii) left posterior temporal regions (BA 22) contribute to acoustically based phonological analysis, (iv) left inferior frontal and anterior insular regions contribute to articulatorily based phonological analysis, (v) regions near the border between the superior and middle temporal gyrus (BA 22/21) are involved in semantic analysis, and (vi) left occipital and occipitotemporal regions (BA 18 and 37/19) are involved in visual analysis specific to word-like stimuli (for further review of these claims, see refs. 13–18). Though such an account provides further confirmation that neuroimaging can be used to fractionate the regions and processes involved in reading, it does little to move beyond broad functional-anatomic associations between brain regions historically associated with language processing (e.g., Broca’s area), and broad processing domains (e.g., reading involves orthographic, phonological, and semantic analysis).
As an alternative to a more general review of neuroimaging results, the remainder of this review will be deliberately narrow in focus with respect to discussing a specific experimental approach and a set of related neuroimaging results, but deliberately broad in terms of discussing the approach and results in the context of information from other methodologies. Such in-depth and interdisciplinary analyses will be critical as neuroimaging moves beyond merely localizing brain regions involved in reading, and toward using neuroimaging as a tool for understanding the cognitive processes involved in reading and their neural substrates. Specifically, the remainder of this review first will discuss the importance of spelling-to-sound consistency in the transformation from orthographic (word form) to phonological (word sound) representations. Then, results from three neuroimaging studies in which the spelling-to-sound consistency of the stimuli was deliberately varied will be evaluated, with an emphasis on the pattern of activation observed in the left frontal cortex.
Orthographic to Phonological Transformation
One component of reading a word aloud is the generation of appropriate word sound (phonological) and articulatory representations on the basis of what are initially orthographic (visual word form) representations. A critical feature of orthographic to phonological transformation in English is the fact that the correspondence between how words look and how they sound is high, but it is not absolute: i.e., although orthographically similar words are usually pronounced similarly (e.g., hint, mint, lint), exceptions occur (e.g., pint) (for review, see ref. 19). Evidence that normal subjects develop processes for orthographic-to-phonological transformation that take advantage of this spelling-to-sound consistency comes in part from two observations: first, subjects can read low-frequency consistent words faster than low-frequency exception words, and second, they can readily pronounce nonwords without training.
The impact of spelling-to-sound consistency on the functional anatomy of word reading has already been explored in three studies. Herbster et al. (9) examined the reading of low-frequency consistent and exception words, and pronounceable nonwords. Fiez et al. (unpublished data) examined the reading of low-frequency consistent, low-frequency exception, high-frequency consistent, and high-frequency exception words, and nonwords. Rumsey et al. (4) examined the reading of low-frequency exception words and nonwords, and additionally investigated two other tasks: a lexical decision task (judge whether the presented items are words or nonwords), and a pseudohomophone decision task (judge whether the presented nonwords sound like a real word—e.g., “brane”).
Each of these studies identified several regions in which the activity differed across stimulus conditions. Furthermore, the patterns of these stimulus-related differences were different for different areas—in other words, there was both specificity in which areas showed an effect and what type of effect they showed. For example, whereas some areas (e.g., the left frontal cortex) were more active when subjects read low-frequency exception words than when they read low-frequency consistent words, other areas [e.g., primary motor cortex (BA 4)] showed the opposite pattern, and in some areas (e.g., the fusiform gyrus), no difference between these conditions was found. Such specificity is hard to reconcile with the hypothesis that the processes involved in reading are diffusely and equivalently distributed throughout many areas. Rather, the results provide support for the claim that there are discrete functional areas that perform specific types of information transformations, and that the activation of these areas can be influenced by different types of experimental manipulations. Discovering that areas are sensitive to an experimental manipulation of interest thus is important, but it is only a first step. To illustrate this point, stimulus-related differences in the activation of the left frontal cortex will be described below, and then potential interpretations of these differences will be explored within the
context of results from behavioral studies in patients with acquired brain damage and theoretical models of reading.
In both the study by Herbster et al. (9) and the study by Fiez et al. (unpublished data), significant left frontal opercular activation (activation located near the borders between BA 44, 45, and the anterior insula) was found for low-frequency exception words but not for low-frequency consistent words; furthermore, in both cases significant activation was found for pronounceable nonwords. Significant left frontal opercular activation for low-frequency exception words was also reported by Rumsey et al. (4), but in contrast to the results of Herbster et al. (9) and Fiez et al. (unpublished data), significant activation for nonwords was not found. However, other observations reported by Rumsey et al. (4) make this failure less problematic: (i) the left frontal opercular activation for low-frequency exception words was not significantly different from the activation for nonwords, and (ii) in another task that presumably involves covert pronunciation of nonwords (the pseudohomophone decision task described above), significant left frontal opercular activation was found. The results across these three studies are summarized in Table 2. (As a cautionary note, it should again be stressed that even though the results will be discussed in terms of a single region in the left frontal operculum, the available data suggest that there are different functional subregions within the neuroanatomical area that encompasses BA 47, 45, 44, and the anterior insula; refs. 20 and 21).
Effects of spelling-to-sound consistency are related to the transformation from orthographic to phonological representations (19). An area such as the frontal operculum, which shows an effect of consistency (e.g., exception words produce more activation than consistent words), is a candidate participant in this transformation process. Thus, the claim is that neuroimaging can successfully identify an area of interest (left frontal opercular cortex) and a candidate process (orthographic-to-phonological transformation) to which this region contributes (either directly, by computing the transformations, or indirectly, as a recipient of the transformations). The next section explores this finding within the context of results drawn from the neurological and neuropsychological literatures.
The Left Frontal Cortex and Phonological Dyslexia
If the left frontal operculum contributes to orthographic to phonological transformation, then damage to the left frontal cortex should lead to impairments in this ability, unless alternate strategies are available or significant recovery of function occurs. Is this the case? Answering this question is not as straightforward as it might seem. This is because the study of brain-damaged subjects often tends to focus either on a cognitive process (a typical approach in the neuropsychological literature) or on an anatomical region (a typical approach in the neurological literature). Thus, to evaluate whether damage to left frontal cortex is associated with impairments in orthographic-to-phonological transformation, the results from two different literatures must be pieced together.
Within the neurological literature, damage to the left frontal cortex and surrounding tissue typically is associated with
Table 2. Left frontal opercular (BA 44/45/insular cortex) activation across studies and conditions
|
|
Low-frequency words |
|
|
|
Study |
Consistent |
Exception |
Nonwords |
|
Herbster et al. (9) |
NS |
–40, 12, –4 |
–48, 6, 0 |
|
Rumsey et al. (4) |
Not tested |
–32, 8, 4 |
–32, 16, 4* |
|
Fiez et al. (unpublished data) |
NS |
–35, 15, 6 |
–39, 11, 12 |
|
NS, not significant. *From pseudohomophone condition. |
|||
Broca’s aphasia, which includes such characteristics as nonfluent speech production, agrammatism, impaired sentence repetition, and relatively preserved speech comprehension (22–24). Deficits in word and/or nonword reading are not commonly considered to be a feature of Broca’s aphasia, and case descriptions of Broca’s aphasics do not usually include reports of word or nonword reading performance (e.g., see refs. 25 and 26). The lack of robust inferior frontal activation found in neuroimaging studies when subjects read aloud high-frequency words (ref. 5; Fiez et al., unpublished data) may explain this apparent discrepancy. If high-frequency words can be read without left frontal involvement, or they are more robust to damage, specific impairments in reading low-frequency words and nonwords could easily be missed by standard diagnostic measures of aphasia, which are composed mainly of high-frequency words (e.g., see ref. 24). Interestingly, there have been suggestions that at least some Broca’s aphasics do exhibit a reading impairment, with one distinguishing feature being the relative inability to read single letters as compared with words (27–29). Unfortunately, neurological case descriptions of such Broca’s aphasics have not systematically included measures of both word and nonword reading accuracy, and in some cases reading skills were assessed by using a measure of comprehension rather than pronunciation (e.g., point to the picture that matches a written word; ref. 29). Furthermore, in many of the Broca’s aphasics with reading impairments, the location of damage extended well beyond or did not even include the frontal cortex (28, 29).
Within the neuropsychological literature, different types of reading impairments resulting from brain damage (acquired dyslexia) have been described. The syndrome of phonological dyslexia is particularly relevant for this discussion, because within the domain of reading it has been viewed as a relatively pure impairment in orthographic-to-phonological transformation (though some reports have emphasized that phonological dyslexics have more general phonological deficits; for review, see ref. 30). The behavioral hallmark of phonological dyslexia is an impairment in nonword reading, with normal to relatively preserved word reading (31–33). Explanations for why these subjects are able to read words, even though an impairment in orthographic-to-phonological transformation is thought to exist, will be discussed and related to the neuroimaging results in the next section. For the moment, the question of interest is whether damage to the left frontal cortex results in phonological dyslexia. To address this question, a review of the neuropsychological literature was conducted. A total of 32 cases of phonological dyslexia (as described within the original publications) were identified in 18 reports. Of these cases, a neuroanatomical description of the damaged region was provided in 14 instances (32–43), as summarized in Table 3. In three cases the damage was limited to the left frontal cortex, and in another four cases the damage extended beyond the frontal cortex to the parietal and/or temporal cortices. These results provide tentative support for a connection between left frontal lobe damage and phonological dyslexia. However, damage to the frontal cortex is not necessarily associated with phonological dyslexia, as demonstrated by the fact that seven subjects with phonological dyslexia had damage limited to the parietal, temporal, and/or occipital cortices. Interestingly, six of the seven patients with left frontal damage tended to produce real-word responses when attempting to read nonwords, whereas four of five patients with damage limited to left posterior regions tended to produce incorrect nonwords. It is possible that there are different subtypes of phonological dyslexia, and the location of anatomical damage is one factor that accounts for differences between patients.
Taken as a whole, findings in both the neurological and neuropsychological literatures provide some evidence that the left frontal operculum contributes to the process of orthographic-to-phonological transformation, but further work
Table 3. Phonological dyslexics with neuroanatomically described lesions
|
|
Words |
Nonwords |
Letters |
|||||||
|
Original source |
Sbj |
Lesion |
Prod |
rep |
read |
rep |
read |
err |
Read |
snd |
|
Bradley and Thompson (34) |
PM |
F |
Fluent |
|
91% |
OK |
13% |
W |
77% |
0% |
|
Caramazza et al. (35) |
LB |
F |
Mild imp |
OK |
100% |
93% |
71% |
N |
|
|
|
Sasanuma et al. (36) |
TY |
F |
Mild imp |
100% |
93% |
86% |
40% |
W |
|
82% |
|
Denes et al. (37) |
ML |
F+P |
Imp |
80% |
94% |
50% |
30% |
W |
100% |
8% |
|
Bub et al. (38) |
MV |
F+P |
Mild imp |
|
92% |
|
23% |
W |
|
93% |
|
Patterson (32) |
AM |
F+P+T |
Mild imp |
OK |
≈95% |
83% |
≈15% |
W |
|
|
|
Friedman (39) |
MS |
F+T |
Fluent |
OK |
85% |
|
15% |
W |
100% |
|
|
Friedman and Kohn (40) |
HR |
P+T |
Fluent |
68% |
87% |
87% |
7% |
N |
|
|
|
Friedman (39) |
BR |
P+T |
Fluent |
100% |
98% |
87% |
61% |
W? |
100% |
|
|
Bisiacchi et al. (41) |
RR |
P+T |
Fluent |
98% |
97% |
100% |
63% |
N |
100% |
100% |
|
Beauvois and Derousense (33) |
RG |
P+T+O |
Fluent |
100% |
100% |
83% |
26% |
|
87% |
|
|
Cuetos et al. (42) |
AD |
P+O |
Fluent |
100% |
93% |
100% |
35% |
N |
|
95% |
|
Derouesne and Beauvois (43) |
JA |
T |
Fluent |
100% |
100% |
|
53% |
|
||
|
Caramazza et al. (35) |
AG |
T |
Fluent |
|
98% |
|
69% |
N |
|
|
|
For each case report (listed by original source and subject initials), the location of the lesion is described (F, frontal; P, parietal; T, temporal; and O, occipital; with the exception of left-handed subject AM, all lesions were in the left hemisphere), along with the speech production (Prod) characteristics of the subject [fluent, mildly impaired (mild imp), or impaired (imp)]. Word repetition (rep) and reading (read) accuracy is listed, followed by nonword repetition and reading accuracy, along with a notation of whether the subject’s nonword reading errors (err) were predominantly real words (W) or nonwords (N). Finally, accuracy at reading (read) and sounding out (sound) individual letters is listed. Blank cells indicate that the relevant data were not available in the original |
||||||||||
clearly is necessary to provide solid support for this conclusion. Rather than being problematic, the need for additional research demonstrates an important benefit of neuroimaging: by focusing attention on both a particular region and a hypothesized function, gaps in previous work can be revealed and directions for future work can be established. For instance, further evaluation of word and nonword reading performance in patients with damage limited to the left frontal cortex could help establish the relationship of this region to the syndrome of phonological dyslexia.
The Left Frontal Operculum and Models of Reading
Up to this point, the transformation from orthography to phonology has been discussed as if it were a single, indivisible process. But consideration of the behavioral data reveals a capacity that most of us take for granted: our reading ability generalizes to the pronunciation of nonwords, but at the same time we can accurately pronounce exception words, which violate these same generalizations. Whether humans can and do accomplish these two tasks by using the same computational process to transform orthography to phonology has been a matter of significant debate.
The dual-route model of reading asserts that two different processes exist for translating print to sound (reviewed in refs. 44–46). In this model, a direct route translates entire word form representations into phonological representations by using an associative process, whereas an assembled route allows the word to be “sounded out” by translating letter units (graphemes) into corresponding sound units (phonemes) by using a rule-based process. Words that violate the “rules” of English (exception/inconsistent words, such as “pint”) can only be pronounced correctly by using the direct route, although an incorrect representation of the phonology may also be created via the assembled route (e.g., the assembled route should generate a pronunciation that rhymes with “lint”). In contrast, the “rules” contained within the assembled route provide the generalizations that support nonword reading, although the direct route may be utilized in the search for an existing lexical representation. Dual-route models can account for phonological dyslexia by assuming that the assembled route is damaged, but the direct route is still available to support the reading of words.
The development of connectionist models for word reading (19, 47) called into question many of the basic assumptions of the dual-route model. Even though the computations were performed by using a parallel distributed process without any explicit coding of the spelling-to-sound “rules” of English, the model could successfully pronounce both consistent words and exception words, as well as nonwords (19). These computational results formed the basis for an alternative model of reading in which regularity effects are thought to arise not from the operation of two, distinct computational processes, but instead through a single process that is sensitive to statistical relationships between orthographic and phonological patterns that develop through repeated word exposure. It is these statistical relationships, encoded in terms of distributed representations across phonological units with different connection strengths to orthographic units, that support generalizations to nonwords (19, 47). Connectionist models can account for phonological dyslexia by assuming that the connections between orthography and phonology, or to the phonology itself, are damaged; as a result, nonword reading is impaired, but word reading receives additional support via connections from orthography to semantics to phonology (19).
Because the two classes of models posit that different types of computational processes are involved in reading, one might expect that the models would predict different patterns of activation data, and that neuroimaging thus could be used to distinguish which class of models best captures the processes involved in reading. But relating the two methodologies turns out to be quite tricky. One point of uncertainty concerns the type of equivalence that should be sought. Should the models predict patterns of activation across a whole set of brain areas, or should they inform interpretations of a single region? Implementing either answer is difficult, because even though both the dual-route and connectionist models are composed of separate layers or subcomponents, it is the overall performance of the models that mimics human reading behavior. As a result, measures that allow an individual layer or subcomponent in either class of models to be mapped onto the activation of a brain region have not been explored. As discussed below, inferences can be made on the basis of the known computational functions of the layers and subcomponents of the models. However, these inferences can become problematic when interactions with other layers and subcomponents are taken into account. This raises a third issue: just because two layers or components are computationally distinct, there is no guarantee that some measure of their “use” or “activation” will be distinct. Finally, there are issues in
neuroimaging that also make computational interpretations difficult—for instance, the changes measured with neuroimaging do not distinguish between distant and local synaptic connections, or between excitatory and inhibitory connections.
The potential ambiguity in relating the models to neuroimaging results is well illustrated by considering the activation pattern observed in the left frontal operculum. This pattern of activation can be summarized as nonsignificant activation for low-frequency consistent words, and robust activation for low-frequency exception words and nonwords (see Table 2). On the one hand, this pattern of activation agrees nicely with inferences about the “activation” of the phonological layer in connectionist models (or possibly the hidden layer between orthography and phonology). Representations sufficient to produce a correct response should be formed rapidly (producing less “activation” per stimulus) for low-frequency consistent words, because the representations benefit not only from prior exposure to the exact word, but also from prior exposure to other words with similar spelling-to-sound patterns. Representations for low-frequency exception words and nonwords should be formed more slowly (producing more “activation”). For exception words, this is because the representations must depend largely on prior exposure to the exact word, because only limited exposure to similar exception words will have occurred. For nonwords, the converse is true—there is no benefit from prior exposure to the stimulus, and instead a representation must be based on similarity to previously presented words.
Dual-route models (44–46) can also account for the left frontal opercular activation pattern, but for very different reasons. In this framework, the left frontal operculum could be involved in the assembled route, in which a rule-based process is used to “sound out” a pronunciation on the basis of correspondences between individual graphemes and phonemes, or the operculum could be a recipient of information from both the assembled and the direct routes (such as the phonological buffer proposed in ref. 46). “Activation” of the assembled route in the nonword condition makes sense, because the direct route (which is based on associations between existing orthographic and phonological representations of whole words) cannot support generalizations to nonwords. Low “activation” of this route for consistent words is also interpret able, because the pronunciation of low-frequency consistent words can be driven beneficially by output from both the direct and the assembled route. For low-frequency exception words, this is not the case: the output from the assembled route will actually produce incorrect information (e.g., that “pint” should be pronounced as if it rhymes with “hint”). Until the competition with the output from the direct route can be resolved and a single pronunciation established, continued “activation” of the assembled route, or a recipient buffer, might be necessary. It is this competition that is thought to cause subjects to read low-frequency exception words more slowly than low-frequency consistent words (44–46).
It is thus possible to form interpretations of the neuroimaging data that can account for the results by using either a dual-route or a connectionist framework, though as noted this involves a set of inferences that have yet to be evaluated empirically. At some level, it is hardly surprising that multiple interpretations can be formed, because the activation pattern in the left frontal operculum parallels the behavioral effects of consistency on reaction time, and it is these behavioral effects that the models seek to explain. But the existence of multiple interpretations does not negate the value of having additional constraints for consideration in the further development of theoretical models. For instance, one intriguing aspect of the proposed interpretations is that they lead to different predictions about the effects of left frontal brain damage. Specifically, in the connectionist framework, the greater activation of this region when subjects read aloud exception as opposed to consistent words can be thought of in some sense as a reflection of the fact that the transformation is slower. Hence, the prediction would be that patients with left frontal damage should have the greatest difficulty reading low-frequency exception and nonwords, and that they should still read low-frequency exception words more slowly than low-frequency consistent words (although in severe cases, phonology may be accessed almost entirely via semantics, and hence the magnitude of the consistency effect would be reduced; ref. 19). But in the dual-route framework, the activation of the left frontal operculum for exception words is not only unnecessary, it is actually counterproductive. Hence, the prediction would be that the patients should read low-frequency exception words just as rapidly as low-frequency consistent words, because in dual-route models the elevated reaction time seen in normal subjects emerges from competition between the direct and assembled routes.
Another important point to keep in mind is that although this review focused on the pattern of activation observed in the left frontal operculum, other regions showed different patterns of stimulus-related differences that also place constraints on theoretical accounts of reading. For instance, both the left frontal operculum and the left posterior superior temporal cortex (BA 22) have been implicated in phonological processes, but of these two regions, only the left frontal operculum showed any effects of consistency. This suggests that the nature of the phonological processes supported by these two regions differs. On the other hand, effects of consistency observed in primary motor cortex (greater activation for low-frequency consistent words and nonwords than low-frequency exception words) are difficult to reconcile with existing models of reading (Fiez et al., unpublished data). This result suggests that the transformation from orthography to phonology involves processing that is not accounted for within existing models, or that motor cortex is involved in a process outside the scope of the implemented models, such as the transformation from phonological to articulatory representations (Fiez, et al., unpublished data).
Summary
In the first section of this review, the results from nine neuroimaging studies were reviewed to give a “big picture” of the brain regions that are active during reading. Though such localization is important, it is only the beginning. Neuroimaging can also be used to reveal differences in the activation of brain regions, and this in turn opens up the possibility that neuroimaging can provide new insights into both the cognitive processes involved in reading and the location of these cognitive processes. Although there is much work to be done, results from neuroimaging can already be used to provide new perspectives on the results obtained in using other methodologies, and there is every reason to believe that neuroimaging will continue to be a value component of an interdisciplinary effort to understanding reading.
This work was supported by in part by NIH grants NS06833, EY08775, HL13851, NS32979.
1. Geschwind, N. (1972) Sci. Am. 226, 76–83.
2. Price, C.J., Moore, C.J. & Frackowiak, R.S.J. (1996) Neuroimage 3, 40–52.
3. Price, C.J., Wise, R.J.S., Warburton, E.A., Moore, C.J., Howard, D., Patterson, K., Frackowiak, R.S.J. & Friston, K.J. (1996) Brain 119, 919–931.
4. Rumsey, J.M., Horwitz, B., Donohue, C., Nace, K., Maisog, J.M. & Andreason, P. (1997) Brain 120, 739–759.
5. Petersen, S.E., Fox, P.T., Posner, M.I., Mintun, M. & Raichle, M.E. (1989) J Cognit. Neurosci. 1, 153–170.
6. Bookheimer, S.Y., Zeffiro, T.A., Blaxton, T., Gaillard, W. & Theodore, W. (1995) Human Brain Map 3, 93–106.
7. Price, C.J., Wise, R.J.S., Watson, J.D.G., Petterson, K., Howard, D. & Frackowiak, R.S.J. (1994) Brain 117, 1255–1269.
8. Howard, D., Patterson, K., Wise, R., Brown, D., Friston, K., Weiller, C. & Frackowiak, R. (1992) Brain 115, 1769–1782.
9. Herbster, A.N., Mintun, M.A., Nebes, R.D. & Becker, J.T. (1997) Human Brain Map 5, 84–92.
10. Talairach, J. & Tournoux, P. (1988) Co-Planar Stereotaxic Atlas of the Human Brain (Thieme, New York).
11. Baddeley, A. (1986) Working Memory (Clarendon, Oxford).
12. Paulesu, E., Frith, C.D. & Frackowiak, R.S.J. (1993) Nature (London) 362, 342–345.
13. Demb, J.B. & Gabrieli, J.D.E. in Converging Methods for Understanding Reading and Dyslexia, eds. Klein, R. & McMullen, P. (MIT Press, Cambridge, MA).
14. Demonet, J.F., Wise, R. & Frackowiak, R.S.J. (1993) Human Brain Map 1, 39–47.
15. Zatorre, R.J., Meyer, E., Gjedde, A. & Evans, A.C. (1996) Cereb. Cortex 6, 21–30.
16. Fiez, J.A., Tallal, P., Raichle, M.E., Miezin, F.M., Katz, W.F. & Petersen, S.E. (1995) J. Cognit. Neurosci. 7, 357–375.
17. Price, C., Wise, R. & Frackowiak, R. (1996) Cereb. Cortex 6, 62–70.
18. Chertkow, H. & Murtha, S. (1997) Clin. Neurosci. 4, 78–86.
19. Plaut, D.C., McClelland, J.F., Seidenberg, M.S. & Patterson, K. (1996) Psychol. Rev. 103, 56–115.
20. Fiez, J.A. (1997) Human Brain Map 5, 79–83.
21. Buckner, R.L., Raichle, M.E. & Petersen, S.E. (1995) J. Neurophysiol. 74, 2163–2173.
22. Damasio, A.R. (1992) N. Engl. J. Med. 326, 531–539.
23. Benson, D.F. (1979) Aphasia, Alexia, and Agraphia (Churchill Livingstone, New York).
24. Goodglass, H. & Kaplan, E. (1983) The Assessment of Aphasia and Related Disorders (Lea & Febiger, Philadelphia).
25. Mohr, J.P., Pessin, M.S., Finkelstein, S., Funkenstein, H.H., Duncan, G.W. & Davis, K.R. (1978) Neurol 28, 311–324.
26. Alexander, M.P., Naeser, M.A. & Palumbo, C. (1990) Neurol 40, 353–362.
27. Benson, D.F. (1977) Arch. Neurol. 34, 327–331.
28. Kirshner, H.S. & Webb, W.G. (1982) Arch. Neurol. 39, 84–87.
29. Boccardi, E., Bruzzone, M.G. & Vignolo, L.A. (1984) Neuropsychology 22, 745–754.
30. Coltheart, M. (1996) Cognit. Neuropsychol. 13, 749–762.
31. Funnell, E. (1983) Br. J. Psychol. 74, 159–180.
32. Patterson, K.E. (1982) in Normality and Pathology in Cognitive Functions, ed. Ellis, A.W. (Academic, London).
33. Beauvois, M.F. & Derouesne, J. (1979) J. Neurol. Neurosurg. Psychiatry 42, 1115–1124.
34. Bradley, V.A. & Thomson, M.E. (1984) Brain Lang. 22, 292–302.
35. Caramazza, A., Miceli, G., Silveri, G.M. & Laudanna, A. (1985) Cognit. Neuropsychol. 2, 81–114.
36. Sasanuma, S., Ito, H., Patterson, K. & Ito, T. (1996) Cognit. Neuropsychol. 13, 823–848.
37. Denes, G., Cipolotti, L. & Semenza, C. (1987) Cognit. Neuropsychol. 4, 11–31.
38. Bub, D., Black, S., Howell, J. & Kertesz, A. (1987) in The Cognitive Neuropsychology of Language, eds. Coltheart, M., Sartoir, G. & Job, R. (Erlbaum, London).
39. Friedman, R.B. (1996) Cognit. Neuropsychol. 13, 869–885.
40. Friedman, R.B. & Kohn, S.E. (1990) Brain Lang. 38, 278–297.
41. Bisiacchi, P.S., Cipolotti, L. & Denes, G. (1989) Q. J. Exp. Psychol. 41A, 293–319.
42. Cuetos, F., Valle-Arroyo, F. & Suarez, M.P. (1996) Cognit. Neuropsychol. 13, 1–24.
43. Derouesne, J. & Beauvois, M.F. (1979) J. Neurol. Neurosurg. Psychiatry 42, 1125–1132.
44. Humphreys, G.W. & Evett, E.J. (1985) Behav. Brain Sci. 8, 689–740.
45. Coltheart, M., Curtis, B., Atkins, P. & Haller, M. (1993) Psychol. Rev. 100, 589–608.
46. Besner, D. (1997) in Converging Methods for Understanding Reading and Dyslexia, eds. McMullen, P. & Klein, M. (MIT Press, Cambridge, MA).
47. Seidenberg, M.S. & McClelland, J.L. (1989) Psychol. Rev. 96, 523–568.








