
Proc. Natl. Acad. Sci. USA
Vol. 94, pp. 8354–8361, August 1997
Colloquium Paper
This paper was presented at a colloquium entitled “Carbon Dioxide and Climate Change,” organized by Charles D. Keeling, held November 13–15, 1995, at the National Academy of Sciences, Irvine, CA.
Geochemistry of corals: Proxies of past ocean chemistry, ocean circulation, and climate
ELLEN R. M. DRUFFEL
Department of Earth System Science, University of California, Irvine, CA 92697
ABSTRACT This paper presents a discussion of the status of the field of coral geochemistry as it relates to the recovery of past records of ocean chemistry, ocean circulation, and climate. The first part is a brief review of coral biology, density banding, and other important factors involved in understanding corals as proxies of environmental variables. The second part is a synthesis of the information available to date on extracting records of the carbon cycle and climate change. It is clear from these proxy records that decade time-scale variability of mixing processes in the oceans is a dominant signal. That Western and Eastern tropical Pacific El Niño-Southern Oscillation (ENSO) records differ is an important piece of the puzzle for understanding regional and global climate change. Input of anthropogenic CO2to the oceans as observed by 13C and 14C isotopes in corals is partially obscured by natural variability. Nonetheless, the general trend over time toward lower d18O values at numerous sites in the world’s tropical oceans suggests a gradual warming and/or freshening of the surface ocean over the past century.
The biogeochemical cycling of carbon on Earth has undergone marked changes over the past glacial-to-interglacial period, and nearly all of these changes have occurred prior to scientific observation. The key to understanding present and future changes in climate and the cycling of biogeochemically important elements (i.e., C, N, S, P, O, Cd, Ba, Si, etc.) is to adequately account for the past changes in their cycling. To enable retrospective studies, a number of substrates on Earth have acted as integrators of these changes. Within the bands of tree rings lie records of relative seasonal rainfall and 14C/12C ratios ( 1) of the atmospheric CO2 used during photosynthesis. Another recorder of climate is oceanic sediment, which contains a layered time history of faunal shells that lived in overlying surface and deep waters. Ice cores have been used to reveal past air temperatures ( 2 ) and atmospheric CO2 concentrations ( 3 ) during the past 100,000 years.
The skeletons of corals provide an unaltered record of the chemical and physical conditions that existed in the surrounding seawater at the time of accretion of its calcium carbonate skeleton. The advantages of corals as an oceanic recorder are the enhanced time resolution (biweekly to seasonal) available from the high growth rate, and the absence of mixing processes that are present in all oxic sediments (i.e., bioturbation).
There are two aspects of this synthesis paper. The first is a brief review of the field of coral geochemistry. A brief discussion of coral biology, taxonomy, and microstructure is presented. Factors affecting coral skeletal growth and isotopic and chemical records within the coralline aragonite are reviewed, and a discussion of the annual and subannual density banding within the skeletal matrices is included.
Second, a synthesis is presented of the available proxy records from corals as they relate to past climate, ocean circulation changes, and anthropogenic input of excess CO2. For some databases, the anthropogenic CO2 appears clear and globally distributed. For other databases it appears that variability on interannual to decadal time scales complicates a straightforward attempt to separate natural perturbations from the anthropogenic CO2 signal on the Earth’s environment. Shen ( 4 ) presents a review of the types of coral archives available from the perspective of historical El Niño-Southern Oscillation (ENSO) influences on the tropical Pacific Ocean. Also, Druffel et al. ( 5 ) present a review of coral growth and the factors that affect it. The present paper attempts to compliment the information contained in these two publications.
Most of the records available to date are from surface corals that are restricted to the upper 50–75 m of the ocean’s surface. However, there is a growing effort devoted to the study of deep-sea corals and gorgonians found at all depths of the world’s oceans (=10,000 m). Most of the discussion in this review centers on surface corals, though new and exciting research involving the use of deep species as paleoclimatologic tools are mentioned.
Review of Corals as Geochemical Proxies
Biology and Microstructure. Corals are from the order Scleractinia, a group in the subclass Zoantharia. Scleractinians include solitary and colonial species of corals, many of which secrete external skeletons of aragonite. The oldest known scleractinians are shallow water corals from the Middle Triassic ( 6).
The polyp portion of the coral secretes calcium carbonate (CaCO3) as the mineral aragonite. The cylindrical outer portion of the polyp is called the polyp wall and is terminated above by the horizontal oral disc and below by the basal disc (see Fig. 1 ). The oral disc is bound by one or more ringlets of tentacles that are covered with stinging nematocysts, which aid in the collection of food and in defense. The mouth is located in the center of the oral disc and leads to the interior gastrovascular cavity where the organs of digestion, absorption, and excretion lie. Within the polyp are three layers of tissue: ectoderm, mesogloea, and endoderm. Parts of the ectoderm that are in direct contact with the skeleton are known as the calcioblast and are responsible for secretion of the aragonite. Within the endoderm of most hermatypic (reef-building) corals are found symbiotic dinoflagellatealgae, called zooxanthellae.
Scleractinians reproduce both sexually and asexually. In most species, ova and sperm are located in the same polyp. Fertilization takes place inside the gastrovascular cavity, and the larvae are ejected through the mouth. The young larvae or planulae settle on a hard, solid surface to form a coral colony. A few days after fixation, the first complete set of mesenteries
|
|
© 1997 by The National Academy of Sciences 0027-8424/97/948354-8$2.00/0 PNAS is available online at http://www.pnas.org. |
and septa are formed. Also, asexual reproduction takes place in an existing coral colony, thereby rendering the distance between coral polyps roughly constant as the surface area of the colony increases.
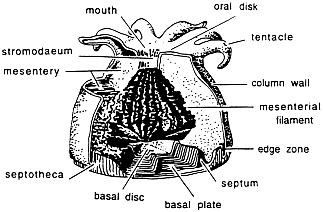
FIG. 1. Anatomical structure of a coral polyp and its association with the skeleton (redrawn from ref.7).
The microstructure of a coral skeleton reveals an intricate matrix that is reflective of the coral’s habits. The skeleton consists of minute, orthorhombic crystalline needles of aragonite, about 2 µ in diameter. When a planulae first settles to form a coral colony, it accretes a horizontal layer of aragonite called the basal disk (Fig. 1). As the polyp grows upward, the margin of the base turns upward, forming a cup, called the epitheca. The epitheca is believed to contain daily growth bands (8) that are formed as a result of daily changes in theshape of the calcioblastic layer (9). Also extending from this basal disk are vertical sheets of septa that support and separatethe mesenteries in the polyps (Fig. 1). Each septum consists of numerous crystal clusters called sclerodermites. A sclerodermite is made from fibrous aragonitic needles radiating from a single calcification center believed to be an amino acid. Dissepiments are horizontal layers of aragonite that form both inside and outside the corallite. They form skeletal boundaries found at the bottom of the coral polyp and are left behind as the result of upward growth of the polyp.
Massive hermatypic corals are more desirable than the branching varieties as the morphology to use for paleoreconstructions. First, massive corals form round, wave-resistant structures that can include hundreds of years of uninterrupted growth. Second, the accretion rate of calcium carbonate is much higher for hermatypic corals that contain symbiotic zooxanthellea than for deep species. There is a considerable amount of evidence that links zooxanthellar activity with calcification rate (10). The fundamental significance of the association between zooxanthellea and coral host has long been controversial as there are several mechanisms through which scleractinians might benefit from the presence of the algae, including translocation of photosynthate-derived carbon to the coral polyp (11,12).
It is important to note that some authors have observed high-Mg calcite in coral skeletons. MacIntyre and Towe (13) examined Porites lobata from an Oahu reef and concluded that the calcite was contained within the tubular organic network formed by microborers (i.e., fungus or algae). Thus, it appears that the presence of calcite in coral skeletons indicates accretion from noncoral sources and reflects contamination. Methods available to detect calcite are x-ray diffractometry (=1% calcite can be detected) and visual inspection under sunlight (calcite twinkles, whereas coralline aragonite is a dull off-white).
Density Banding in Corals. Ma (14) suggested that groupings of constricted epithecae reflected seasonal variations in the growth rate of coral skeletons. Later, Wells (8) studied middle Devonian fossil corals and cautiously interpreted fine ridges on the surface of the coral epitheca (see Fig. 1) to be daily growth bands. He found about 400 ridges (days) per annum, which agreed with astronomical expectations of the deceleration of the Earth’s period of rotation since the Devonian.
The most significant record contained in most surface coral skeletons are annual density bands. They are primary skeletal characteristics that consist of a high and low density portion per year discernible by x-ray of a thin slab cut along the axis of upward corallite growth (15,16). Annual variations in density represent changes in both the rate of linear skeletal extension and calcification. These growth bands were first conclusively demonstrated as annual by Knutson et al. (17). They found excellent agreement between autoradiographs of Enewetak coral, which depict nuclear bomb products incorporated during nuclear blasts (mostly 90Sr), and x-radiographs, which reveal density structure. At a few locations, multiple banding within an annual growth period has been identified, though this is not common (18,19). Also, Buddemeier and Kinzie (18) have identified lunar bands in P. lobata from Kahe Point, Oahu, Hawaii. Using x-radiography and scanning densitometry of Montastrea annularis from St. Croix, U.S. Virgin Islands, Dodge and Brass (20) found that skeletal extension (linear growth) is negatively correlated with density and positively correlated with calcification (mass addition).
Hudson et al. (21) used alizarin-staining techniques and x-radiography to observe annual density bands in M. annularis from the Florida Straits and the Gulf of Honduras. They noticed that dense aragonitic skeleton accreted during the warm summer months of July through September and that thicker, less dense bands accreted during the cooler months of October through June. At some locations, such as the Gulf of Eilat in the Red Sea, corals (Porites lobata) accrete high density bands during the season of cold sea surface temperatures (SST). In Florida, thicker, dense layers, known as stress bands, also formed during the unusually severe winters of late 1969, 1963, 1957, 1941, 1898, 1894, 1885, and 1856. These cold winters or “cold fronts” are the result of weather phenomena that originate in the Northwest and pass over the reef in a southeastern direction. This correlation between stress bands and recorded cold fronts is proof that the density bands in these corals are indeed annual in nature.
Glynn and Wellington (22) studied Pavona clavus colonies from the Galapagos Islands and revealed that dense bands form during the warm water months of January through March and thicker, less dense bands form during the cold, upwelling season, usually April through December. These workers attributed increased growth rates during certain years as the result of ENSO events. These events are linked to the intensity of the Southern Oscillation Index, which is the difference in air pressure between Easter Island and Darwin, Australia (23). When this index is low, the Peru Current decreases its flow and upwelling along the western coast of South America is restricted to below the sea surface for a period of 4 to 10 months. Simultaneously, a thin skin of warm, low-nutrient tropical surface water moves into the Galapagos region from the West and North. Just as stress bands in M. annularis from the Florida Straits depict recorded cold fronts, unusually high growth rates during El Niño years provide markers of known years for the Galapagos corals. Severe ENSO events, such as that during 1982–83, caused interruption of normal growth of Galapagos corals due to the partial or complete loss of their zooxanthellae or bleaching (22). In areas with smaller temperature changes during this ENSO event, such as Costa Rica, the corals were not bleached and hence contained a complete record of the 1982–83 ENSO (24).
Just as dendrochronologists cross date wood samples to extend tree-ring chronologies (25), so can sclerochronology be used to obtain lengthened records of coral growth. Stress bands, like those observed in the Florida Straits coral, could be used as markers of known years in dead colonies. Though annual growth rates of some massive hermatypic corals show
no systematic change for a given colony during its lifetime (26, 27 ), intercolony variations in annual growth rate have been observed (22, 27). Therefore, stress band matching would prove more useful for sclerochronologists than direct growth rate comparisons among coral skeletons. A very fruitful method of long time-series reconstruction of coral records over the past glacial cycle involves measurement of the ingrowth of 230Th (28) (see section titled “Dating of Corals”).
Fac tors Affecting Growth. Optimal coral growth occurs at SST that range from 20° to 26°C (29, 30). The limits of coral tolerance range from a low of about 16°–18°C to a high of 30°–32°C, depending on the species under consideration. Thus, coral growth is limited to the temperate and tropical latitudes between about 35°N and 32°S. For massive corals, skeletal growth rate is dependent on SST (31, 32).
The influence of light as an environmental factor has been extensively studied but is still not well understood. The rate of growth of M. annularis was positively correlated with the number of sun hours per day (33) and the water depth of the corals (31). Calcification rates during daylight periods are 3- to 15-times higher than those during dark periods (34). Goreau and Goreau (35) showed that coral with zooxanthellae calcified about 19-times faster than those that had lost their algae. These zooxanthellae-free corals calcified even less in the light than did the corals with zooxanthellae in the dark. This illustrates that photosynthesis is not the only role that the zooxanthellae play as a symbiont. Buddemeier (27) suggested a negative correlation between skeletal density and available light.
Many other environmental factors affect the growth of corals. For instance, salinity must be between 25‰ and 40‰ to support healthy corals (36). The depth in which corals live also affects their growth. For example, M. annularis shows an optimal growth rate at 10 m depth, but is only 10% of this rate at 30 m depth (37). Sedimentation has long been known as a detriment to the well-being of corals (38). Large amounts of sediment covering the colony may interfere with feeding processes and zooxanthellar photosynthesis. There is also an energy loss by the polyps due to increased mucous production used for shedding the sediment.
In summary, optimal growth conditions for hermatypic corals include temperatures within the range of 20°–26°C, low sediment load, growth at 5–15 m depth, normal ocean salinities, and relatively protected water. These conditions yield an optimum vertical growth rate of 10–15 mm/yr for massive species (39).
Dating of Corals. Dating of fossil corals has been done by measuring concentrations of radioisotopes incorporated within the skeletal matrix at the time of accretion (i.e., 14C, 228Ra, 210Pb) and those that have grown in since accretion (i.e., 230Th, 231Pa). For example, 14C dating of coralline aragonite from Enewetak and Oahu has revealed that subtle changes in eustatic sea level have occurred during the last 6,000 years in the North Pacific Ocean (18,40). Moore and Krishnaswami (41) revealed that 228Ra (t1/2 = 6.7 yr), which is incorporated in corals at about the same ratio to calcium as that in seawater, can be used to date corals less than 30 years of age. Lead-210 (t1/2 = 22.7 yr) dating in corals has been shown to agree within 10% with annual banding chronologies of corals from the North Atlantic over the past 100 years of growth (26,41,42). Several bomb-produced radionuclides have also aided in the determination of coral growth rates. Bomb 14C is detected above natural levels in all surface coral bands younger than about 1957 (19, 43, 44 ,45 and 46). Other products of thermonuclear weapons testing (e.g., 90Sr and 239,240Pu) are found in post-1950 corals and reveal past levels of these isotopes in the surrounding seawater (47, 48,49 and 50).
Measurements of 234U and its in-growth daughter, 230Th, in corals from the Florida Keys and the Bahamas revealed that sea level was close to its present interglacial level at about 85,000, 130,000, and 190,000 years ago (51). A second in-growth method was presented by Ku (52), who measured 231Pa in corals from Barbados; he found good agreement between the ages of corals from the last interglacial determined from the in-growth of 231Pa and those determined using the 230Th in-growth method.
A most exciting discovery in the field of coral geochemistry was made by Edwards et al. (28). They developed techniques for measuring 230Th in corals by using isotope dilution thermal ionization mass spectrometry (TIMS). This new method greatly reduced both the sample size (from tens of grams to 250 mg of coral) and the uncertainty in the age determination (by a factor of 10–20) compared with the previously used alphaspectrometry methods. It is now possible to date corals with ages of 30, 12,000, and 123,000 years with uncertainties of ±3, ±60, and ±1,000 years [two sigma errors, respectively (28,53)]. This discovery made it possible to more accurately date the last interglacial period (28), the Younger Dryas, and the levels of 14C in the surface ocean dissolved inorganic carbon during the last glacial maximum (53,54).
Carbon and Oxygen Isotopes in Coral Skeletons. Most of the records obtained from corals have been of stable oxygen (δ18O) and carbon (δ13C) isotopes in annual and seasonal bands. The majority of the carbon found in coral skeletons originates as dissolved inorganic carbon (DIC) in sea water. Goreau ( 34 ) proposed the following as the dominant reaction that occurs during the accretion of calcium carbonate by corals:
Ca2+ + 2 HCO−3 → H2O + CaCO3 + CO2.
McConnaughey ( 55 ) refined this by suggesting intermediate reactions involving dehydration of the HCO−3 (2HCO−3 → 2H2O + 2CO2), then the subsequent rehydration and disassociation of the hydrogen ions to form CO=3, which is used in the precipitation of CaCO3. It is believed that zooxanthellae increase CaCO3 accretion by using the CO2 produced by this reaction during photosynthesis. This is one of the reasons corals with symbiotic algae accrete calcium carbonate at such rapid rates.
Weber (56) found that the δ13C (per thousand deviation of 13C/12C ratio with respect to that of PDB-1 standard) of skeletal carbonate ranged from 1‰ to −1‰, whereas the δ13C of the DIC in the surrounding seawater was 3‰ to 1‰. Earlier, the lower δ13C values were attributed in part to the incorporation of a small amount of metabolic carbon from the coral’s diet of zooplankton (δ13C = −18‰; ref.56 ) to the carbonate matrix (58 ). Recent interpretation shows that the reasons for carbon and oxygen isotope disequilibria in corals with respect to those in seawater are twofold (55,59). First, kinetic isotopic effects during the hydration and hydroxylation of CO2 cause the simultaneous depletions of 18O and 13C with respect to their lighter isotopes (16O and 12C) by as much as 4–6‰ and 10–15‰, respectively. There is an approximately linear correlation between δ18O and δ13C signals in corals (59,60 and 61) and in a pure calcite gorgonian (62), due to the degree of kinetic disequilibria that exists in the calcioblastic layer. Second, alteration of the skeletal δ13C signal is caused mainly by changes of the δ13C in the internal DIC pool within the calcioblastic layer, which are caused by varying rates of photosynthesis and respiration by the coral polyp and its symbiotic algae. During photosynthesis, selective removal of 12CO2 by the zooxanthellae during photosynthesis causes an increase of δ13C in the DIC left behind to form the accreted aragonite. Likewise, during respiration of organic matter, addition of 12C-enriched CO2 decreases the δ13C of the skeletal carbonate (63). These metabolic effects are responsible for large shifts of skeletal δ13C resulting primarily from the seasonally variant ambient light incident on the coral surface (64). McConnaughey (59) showed that nonseasonal changes of δ13C can be minimized by sampling along major growth axes (i.e., tops of coral heads). The periodicity of subannual δ13C and high density bands in three Atlantic corals varied as a function of seasonal cloud cover (65). The δ18O patterns in the same
corals, however, were a function of both SST and salinity in the seawater measured at each site. δ13C of carbonate is only slightly affected by ambient temperature (0.035‰/°C).
Epstein and Mayeda ( 66) first showed that δ18O in mollusk aragonite varied inversely with SST. It was subsequently demonstrated that the δ18O of coralline aragonite also varied inversely with SST though different genera were offset by constant amounts from this δ18O vs. SST relationship (67, 68). Offsets demonstrate that precipitation occurs faster than the establishment of oxygen isotope equilibrium between CaCO3 and H2O in the calcioblastic layer. Several authors have successfully used δ18O as a monitor of past SST (5,19,65, 69,70). The slope of SST vs. δ18O is similar to the mollusk results (66), a 0.22‰ rise in δ18O per 1°C decrease (Fig. 2).
The other important influence in coral δ18O is the isotopic signature of seawater, which is related to the salinity in the surface water through evaporation and precipitation cycles. Several investigators have demonstrated that subannual δ18O measurements from surface corals are a predictable function of SST and salinity ( 65 , 70 , 71 and 72 ).
Radiocarbon in modern corals was measured first by Knutson and Buddemeier ( 73 ), who detected bomb levels of 14C in the outer 10 cm of a massive coral from Fanning Island. They hypothesized that corals hold records of the input of bomb-produced radionuclides to the surface waters of the ocean. Independently, two groups reconstructed time histories of ∆14C in two Atlantic corals. Nozaki et al. ( 46 ) reported variations in ∆14C and in stable isotopes (δ13C and δ18O) from a 200-year Dipoloria strigosa from North Rock, Bermuda. Druffel and Linick ( 43 ) observed changes of ∆14C from a 360-year core of M. annularis from the Florida Keys. These and other papers have shown that ∆14C of corals closely matches the ∆14C of DIC in surrounding seawater ( 44 , 45 , 74 , 75 ). This isotope record is not affected by isotope fractionation, as all ∆14C measurements are normalized to a δ13C value of −25‰ by first measuring the δ13C value ( 76 ) in the coral. Radiocarbon in corals has been used to reconstruct time histories of climate and circulation (see first two sections of part 2 of this paper).
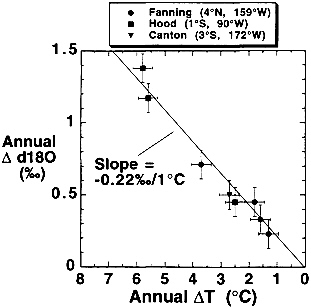
FIG. 2. Correlation between annual range in historical sea surface temperature (SST) records (∆T°C) and annual range in δ18O values (∆δ18O) in coral bands from the Galapagos Islands, Fanning Island, and Canton Island. The line shown represents a slope of 0.22‰ per 1°C decrease in SST predicted by the temperature-dependent aragonite-water fractionation factor. This slope is the same as that defined for the equilibrium precipitation of calcite (66). (Data are from ref. 69.)
Minor and Trace Element Composition of Corals. Divalent cations that are abundant in seawater (i.e., Mg, Sr, and Ba) appear as minor elements in corals (>1 µmol/mol Ca), and other less-abundant divalent cations (M) are in trace abundances (<1 nmol/mol Ca–1 µmol/mol Ca). Most minor and trace element concentrations in corals are incorporated independent of bulk tissue concentrations ( 77 ) and are proportional to ambient levels in the surrounding seawater. Thus the M/Ca ratio in the skeleton can be used to reconstruct past concentrations of M in seawater, assuming Ca levels have remained unchanged in seawater. Shen ( 4 ) reports that the distribution coefficients based on the measured divalent M/Ca ratios in corals and estimated seawater concentrations of the respective metals vary only within an order of magnitude. This indicates kinetic control of the M2+ incorporation, as a wider range is expected from a thermodynamic control for this process ( 78 ).
Using flame atomic absorption spectrophotometry (AAS), Schneider and Smith ( 79 ) measured Sr/Ca ratios in Hawaiian corals and found that this ratio varied as a function of SST, though there were problems with corals that had undergone diagenesis. Subsequent work using TIMS methods has shown that there are corals whose Sr/Ca records track SST with an apparent accuracy of better than ±0.5°C (80). DeVilliars et al. ( 81 ) report that both variable extension rate and biogenic cycling of Sr vs. Ca can complicate some Sr/Ca records in serving as simple sclerothermometers.
The U/Ca ratio in corals is not a simple function of the U/Ca ratio in seawater. Flor and Moore ( 82 ) suggested that decaying organic matter in skeletons of dead corals may release U that exchanges with the carbonate matrix. Shen and Dunbar ( 83 ) report a correlation between U/Ca and δ18O in corals from the Galapagos Islands, although they cannot rule out additional control of U/Ca by salinity and carbonate ion content of surface waters.
Work by Shen et al. ( 84 ) using graphite furnace AAS revealed that Cd/Ca measurements in Bermudian corals uncovered records of historical upwelling and industrial fallout. Only after exhaustive oxidative and reductive cleaning procedures was it possible to obtain the lattice-bound Cd concentrations in the corals, separated from the organic and detrital phases that were much more enriched. These phases contributed to the much higher concentrations of trace metals reported in corals by previous investigators ( 85 , 86 ). Likewise, Ba/Ca ratios in a Galapagos coral revealed historical changes of the nutrient content in surface waters surrounding the Galapagos Islands ( 87 ). Other investigators have reported records in corals for a variety of other minor and trace elements, including Pb, V, Zn ( 42 ), Cu ( 88 ), Mn ( 88 , 89 and 90 ), Mg ( 91 , 92 ), and rare earth elements ( 93 ). An in-depth review of minor and trace element records from corals is presented by Shen ( 4 ). The use of these records as climate recorders and pollution monitors appears later in the paper (see first and third sections of part 2).
Phosphate has been shown to act as a crystal poison of calcification ( 94 ). Dodge et al. ( 95) showed that concentrations of P in corals from three sites in the North Atlantic were consistent with the history of sewage and other pollution in the surrounding reef. Rasmussen (96) demonstrated that high levels of phosphate in seawater were inversely proportional to concentrations of Sr in Porites from the Great Barrier Reef. She further showed that morphological alteration of the coral skeleton is associated with high phosphate levels in the surrounding seawater that came from agricultural fertilizers.
Corals as Proxies of the Carbon Cycle and Climate Change
Synthesis of the information obtained from coral proxy records is done in the second part of this paper. Three questions are
addressed about the cycling of excess carbon in the oceans and changes in climate over the past several hundred years. (i) What are the regional changes in climate within the Atlantic and Pacific Ocean basins? (ii) How have coral parameters revealed variability of climate and ventilation in the North Atlantic and South Pacific? and (iii) What evidence exists of anthropogenic CO2 (fossil fuel and biogenic CO2) in the ocean?
Regional and Global Changes in Climate. Changes of climate on Earth have been recorded in corals for millions of years, though relatively few of the clues have been decoded. These climate records are revealed through physical, chemical, and isotopic imprints in corals. There are several examples in coral records of how changes in regional wind patterns, rainfall, and occurrences of ENSO demonstrate the existence of variability in climate within a given ocean basin.
Early on, Dodge and Vaisnys ( 97 ) established correlations between the linear growth rate of Bermuda corals and recorded air temperature and air pressure variations, demonstrating that paleoclimatic records were recoverable from corals. Also, from growth band analysis of recent and fossil corals from Barbados, Dodge et al. ( 98 ) observed both average band width and its variability were lower in corals from 105,000 years ago (kya) than those from recent times or from 85 kya and 125 kya. Despite this link, they were unable to identify a specific climatic variable as the dominant cause of the differences.
Later, Shen et al. ( 99 ) used multiple tracers for extracting climate records from corals. In a 47-year record (quarter-annual sampled) of Pavona clavus from the Galapagos Islands, highly significant linear correlations were found between four of the tracers examined (δ18O, δ13C, Ba/Ca, Cd/Ca) and Peruvian coast SST. These correlations result from seasonal upwelling cycles and periodic interruptions of these cycles caused by changes in climate (i.e., ENSO).
In a striking example of past climate reconstruction in the tropical western Pacific, Cole et al. ( 71 ) measured δ18O in monthly samples from a 96-year Tarawa Atoll coral and found that δ18O reflected seawater salinity changes induced by regional variations in rainfall. They determined that ENSO extremes in the central and western tropical Pacific were occasionally decoupled from those observed in coastal South America. From spectral analysis of the δ18O data, the distribution of variance had shifted among annual and interannual periods between about 1930 and 1950, in concert with an observed shift in the strength of the Southern Oscillation. This is an example of regional changes in climate within an ocean basin that can be reconstructed from coral skeletons.
Periodic wind bursts associated with ENSO are also recorded in western Pacific coral. Historic Mn/Ca records from a Tarawa Atoll coral revealed high ratios during the ENSO years of 1965, 1972, and 1976 ( 100 ). They concluded that these high Mn periods were due to remobilized Mn from lagoon sediments caused by vigorous westerly wind bursts that happened along the equator during ENSO events. A new tracer of past ENSO events, controlled by the climatic forcing of the tropical winds, was uncovered by this novel study.
Large decadal changes in ventilation rate that were induced by changes in climatic parameters were uncovered by geochemical tracer records ( 101 , 102 ). From post-bomb records of high precision ∆14C in Bermuda and Florida corals, Druffel ( 102 ) used an inverse model to calculate the water mass renewal rate (WMRR) with respect to CO2 ventilation in the Sargasso Sea. Results showed that the WMRR in the Sargasso Sea was high in the early 1960s, decreased in the late 1960s, and remained low throughout the 1970s. This factor-of-three change in the ventilation of the upper North Atlantic reflects the long-term change of climate within an ocean basin.
Climate change has been demonstrated at other locations of the world’s oceans. Chakraborty and Ramesh ( 103 ) demonstrated that Indian summer monsoon rainfall records can be reconstructed from δ18O measured in corals from the Lakshadweep archipelago in the Arabian Sea. Isdale and others ( 104 , 105 ) reported that fluorescent bands in coral skeletons are coincident with periods of high runoff from local rivers feeding into the Great Barrier Reef. Humic materials from terrestrial plants are a major contributor to this fluorescence. This demonstrates a significant correlation between ancient rainfall in Australia and a tracer easily quantified in coastal reefs.
Dunbar et al. ( 106 ) reported the variability of δ18O measured for the past four centuries from Pavona clavus from Urvina Bay in the Galapagos Islands. They found cooler temperatures during the early 1600s and early 1800s, which is in agreement with many North American tree ring records. They suggest that major shifts in the dominant ENSO modes within their data throughout time indicate reorganizations of the tropical Pacific climate system.
A compilation of the δ18O data from several coral series in the tropical Pacific region reveals a striking trend ( 107 ). Although it is not clear-cut, there is a general decrease of δ18O values from the latter part of the last century to the present in nearly all of the coral records available. This appears to be evidence of large-scale warming in the upper layers of the ocean, or a general freshening of these waters, or a combination of these. Is this evidence of global warming as related to anthropogenic inputs of CO2? It is a tantalizing set of records, nonetheless, and decoupling of the SST and water mass components of this trend has been set as a high priority for future research.
Temporal Changes in Ocean Circulation. How have coral parameters revealed variability of climate and ventilation in the North Atlantic and South Pacific? First, there are several modes of circulation in the ocean. Vertical mixing processes include upwelling and winter mixing, and horizontal processes encompass upper ocean currents (i.e., Gulf Stream) and eddies. Some of the best tracers of vertical and horizontal mixing processes are those that record nutrient levels (i.e., Cd/Ca ratios), SST (i.e., δ18O and Sr/Ca), and relative degrees of upwelling (i.e., ∆14C). The available data sets show that temporal variability within the oceans on interannual-to-decade time scales is prevalent during the past few hundred years.
A useful paleocirculation indicator is δ18O in corals, as it records past SST changes in regions that have little or no changes in salinity. From seasonal δ18O measurements in numerous tropical Pacific corals, records of past ENSO events were reconstructed and compared with historical records from the South American coast ( 69 , 106 ). Century time-scale changes in mixing have been observed in the North Atlantic. From an 800-year-old Bermuda coral, Patzold and Wefer ( 108 ) revealed a decrease in the summer SST values during the Little Ice Age. They also found lower overall growth rate during this period, suggesting a relation between coral growth rate and nutrient availability during periods of cooler climate in the North Atlantic.
Radiocarbon in oceanic DIC is controlled mainly by changes in ocean circulation rather than by atmospheric exchange of CO2. There is a large reservoir of DIC in the surface ocean, and it takes a long time (≈10 years) for the 14CO2 in the atmosphere to turn over with respect to exchange with the surface ocean. An excellent example of the use of 14C as a circulation tracer is the study by Toggweiler et al. ( 45 ). They reconstructed the pre-bomb ∆14C values at numerous surface locations in the tropical Pacific using corals and concluded that low ∆14C waters off Peru had their origin as 8°C water in the subantarctic region of the southwestern Pacific. Using comparisons between a model simulation of radiocarbon ( 109 ) and coral ∆14C distributions, they suggested that diapycnal alteration of successively less dense features of the South Pacific
thermocline is responsible for upper ocean ventilation in the southern and tropical Pacific. Thus, Toggweiler et al. ( 45 ) showed that the circulation of subsurface waters in a major ocean basin was not obvious when using traditional conservative properties (temperature and salinity) and that the lagrangian tracer 14C was needed for the patterns and sources to become clear.
Temporal variations in circulation were revealed using high precision (±2–3‰) ∆14C measurements in a 356-year-old coral core from the southern Great Barrier Reef ( 110 ). This study revealed increased decade time-scale variability of ∆14C between A.D. 1680 and 1730 ( Fig. 3 ), coincident with the Maunder minimum in solar activity. Changes in vertical mixing and changes in the transport of the low-14C, South Equatorial Current source waters to the western Coral Sea region are likely processes that could account for this variability. These data also suggest century time-scale changes in the periodicity of ENSO as it is manifest in the southwestern Pacific, and this change correlates with an apparent reduction in ventilation of upper waters by fossil fuel CO2 during the first half of the 20th century ( 110 ).
The main conclusion from these records is that the amplitude of interannual and decadal variability complicates the task of assessing long-term anthropogenic trends. This makes it a challenge, though still a possibility, to decouple the natural variability from the anthropogenic CO2 signal in the oceans.
Evidence of Anthropogenic CO2in the Global Ocean. Corals are diaries that record within their pages many types of environmental information, including nuclear fallout products and excess CO2 input to the ocean. What evidence exists of anthropogenic CO2 (excess CO2 from fossil fuel and biomass burning) in the ocean today?
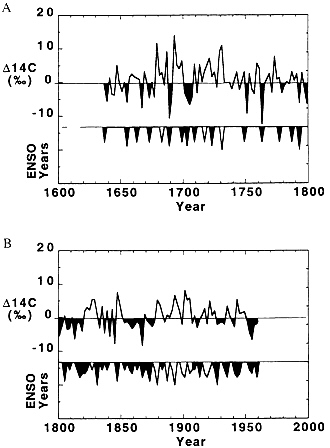
FIG. 3. Biannual Australian coral n∆14C values (in ‰, detrended with respect to a third-order polynomial fit) plotted with incidences of strong and very strong ENSO events before A.D. 1800 (a) and including medium ENSO events after A.D. 1800 (b) as reported by Quinn et al. ( 111 ). Solid peaks represent correlation between ENSO events and coincident (±2–4 years) n∆14C lows, and open areas indicate the absence of this correlation. (Data are from ref. 110 ).
Records of aeolian input of heavy metal pollution to the oceans was presented by Shen et al. ( 84 ). They measured Cd/Ca in a Bermuda coral and found that it revealed a record of aeolian fluxes of industrial Cd to the western North Atlantic from the continental United States via the westerlies. Peaks of high Pb/Ca were also observed in the same coral and resulted from the use of tetraethyl lead in gasoline for cars ( 42 ). The initial rise in the Cd from about 1900 to 1925 was coincident with a rise in zinc production (from which Cd is a byproduct) that occurred at a time when flue dust recovery of Cd was negligible. The second rise of Cd from 1942 to the present was due both to Cd flue dust that still escaped during Zn ore production, as well as to other smelting exhausts ( 84 ).
Records of both global and localized inputs of nuclear fallout to the oceans are available from corals. Toggweiler and Trumbore ( 49 ) measured 90Sr in corals from the Pacific and Indian Oceans and found that localized inputs of 90Sr to the North Pacific atolls between 1952 and 1958 were higher than previously had been believed. They also concluded that major surface currents through the Indonesian archipelago were responsible for the transport of large amounts of 90Sr, and thus water, from the Pacific to the Indian. Finally, this study revealed that there was a major transport of water from temperate to tropical regions of the Pacific on an interannual time scale. Records of 239,240Pu have also been extracted from Caribbean island corals to show a close correlation with the fallout history of 90Sr ( 50 ).
Fossil fuel CO2 has been produced since the middle of the 19th century, and its invasion from the atmosphere into the ocean has been recorded by corals and sclerosponges. This excess CO2 is unique in its carbon isotopic signatures, owing to the absence of 14C (∆14C = −1,000‰) and low δ13C signature (−28‰), in comparison with those in atmospheric CO2 (>0‰ and <−7‰, respectively). Efforts to extract the fossil fuel CO2 signal from the δ13C signature in corals has yielded mixed results owing to the alteration of the DIC δ13C signature by kinetic isotopic and metabolic effects. This δ13C record is preserved without fractionation in sclerosponge skeletons, as these animals accrete aragonite in equilibrium with the surrounding seawater DIC. Using the sclerosponge Ceratoporella nicholsoni from Jamaica, Druffel and Benavides ( 112 ) measured a δ13C decrease of 0.5‰ from 1850 to 1972 and attributed this to fossil fuel CO2 invasion into surface waters. They deconvolved this time history to reveal that the input of an additional source of excess CO2, the reduction in the size of the terrestrial biosphere due to biomass burning, peaked at the turn of the 20th century.
Reduction of the 14C/12C ratio in atmospheric CO2 since 1850, known as the Suess effect, is due mainly to the input of 14C-free CO2 from fossil fuel burning into the atmosphere and ocean ( 113 ). The Suess effect in the surface ocean has been measured in corals from the Atlantic and Pacific Oceans and varies regionally from −3‰ to −12‰ ( 43 , 74 , 110 , 114 ). However, decade time-scale changes in mixing of the upper oceans have caused the Suess effect to vary temporally and spatially, much more so than if ocean mixing had remained in steady state. Large changes in ∆14C of 10–20‰ on decade time scales mask the Suess effect at a number of locations in the southwest Pacific Ocean and Sargasso Sea ( 110 , 114 ). More work needs to be done at other locations in the surface world’s oceans to decouple the Suess effect from decade time-scale changes in circulation.
At present, the magnitude of the anthropogenic CO2 signal in surface waters is difficult to ascertain in surface corals. Changes in mixing parameters that occur at some sites on interannual-to-decade time scales compete with the anthropogenic signal, making it difficult to deconvolve the two. This
problem also arises in studies of the concentration of DIC in the upper ocean. The d13C signal in seawater DIC ( 115 ) and as recorded in sclerosponges remains as a promising method for reconstructing the time history of the anthropogenic CO2 signal into the surface ocean.
To decouple the natural and anthropogenic effects on the carbon cycle, a more intensive effort has to be made to define natural variability in the upper ocean. We must also identify other accurate integrators of the anthropogenic CO2 signal.
Future Research Directions. Reconstruction of past SST changes in the world’s oceans during the period of rising atmospheric pCO2 is a top priority. A lowering of d18O in numerous coral records over the past 150 years may be evidence of a general rise of SST throughout the temperate and tropical surface oceans ( 107 ). Measurements of high precision Sr/Ca ratios will help resolve this important issue. Also, intense sampling and analyses of coral along a transect known for its sensitive response to climate change will add significantly to our knowledge of the global ocean’s response to climate change. Such an area is a meridional section through the warm pool in the western tropical Pacific.
The reconstruction of multiple tracer records from the same corals are essential to fully glean all of the available information regarding past climate and circulation change in the oceans. Measurements of nutrient-like elements (i.e., Ba/Ca), 14C, and stable isotopes on monthly time scales will enhance pursuits of the timing of past changes in water transport and climate. This approach can be extended to the last deglaciation, where the question of the timing of glacial melting and the onset of deep water formation is still not well known. A recorder of salinity changes in ocean water contained within the skeletons of corals should be pursued with vigor.
One of the most exciting and potentially fruitful areas of coral geochemical research is the study of deep-sea species. The primary carbon source to several species of deep calcareous corals is from the DIC in the surrounding seawater ( 116 ). The life spans of single specimens have been shown to be several hundred years ( 62 , 117 ) to more than 1,000 years ( 118 ). It is now possible to reconstruct time histories of tracers in deep water masses, and thus extend our view to three dimensions within the world’s oceans. Records of deep water stable isotopes (d18O and d13C) ( 117 ), trace metals ( 119 ), and bomb fallout products ( 62 ) are starting to emerge in the literature. We need to understand the processes controlling isotopic fractionation of the CaCO3, as well as high-resolution age dating of the fine structure within the skeleton of deep species using the highly precise chronometer 230Th. Subsequently, we can reconstruct time histories of nutrient distributions and ventilation of the deep ocean basins ( 120 ).
Reconstruction of past dissolved organic carbon (DOC) concentrations in seawater is very important for understanding the changes in the carbon cycle over the past 100 years, as well as the past glacial–interglacial cycle. Whether DOC concentrations are recorded within corals remains to be demonstrated. Tritium, a bomb product used as a tracer of ocean circulation, could be reconstructed from the organic matter within corals. Accelerator mass spectrometry measurements of a variety of radionuclides in corals (e.g., 10Be and 7Be) could help to understand particle cycling in seawater. Individual organic compounds would be of interest as indicators of primary production or pollution in coastal locales. It is likely that these records, and many more, are hidden within the pages of the coral diary.
I dedicate this article to Roger Revelle. I fondly remember his comments and encouragement in that melodic, deep voice. I thank Jess Adkins and an anonymous reviewer for their helpful reviews of the manuscript, Sheila Griffin for assisting with drawings and preparation, Andrea Grottoli-Everett for advice, and Ed Urban and Karl Turekian for editorial treatment. This work was funded by the Chemical Oceanography Division of the National Science Foundation through Grant OCE-9314691.
1. Suess, H. E. ( 1980 ) Radiocarbon 22(2) , 200–209 .
2. Dansgaard, W. , Clausen, H. B. , Gundestrup, N. , Hammer, C. U. , Johnsen, S. F. , Kristinsdottir, P. M. & Reeh, N. ( 1982 ) Science 218 , 1273–1277 .
3. Neftel, A. , Moor, E. & Stauffer, B. ( 1985 ) Nature (London) 315 , 45–47 .
4. Shen, G. T. ( 1993 ) Bull. Inst. Fr. Etud. Andines 22(1) , 125–158 .
5. Druffel, E. R. M. , Dunbar, R. , Wellington, G. & Minnis, S. ( 1990 ) in Global Ecological Consequences of the 1982/83 El Niño Southern-Oscillation , ed. Glynn, P. ( Elsevier , New York ), pp. 233–253 .
6. Stanley, G. ( 1981 ) Geology 9 , 507–511 .
7. Wells, J. ( 1956 ) in Treatise on Invertebrate Paleontology , ed. Moore, R. ( Geological Society of America and Univ. of Kansas Press , Lawrence ), pp. 328–444 .
8. Wells, J. W. ( 1963 ) Nature (London) 197 , 948–950 .
9. Barnes, D. ( 1972 ) Proc. R. Soc. Lond. B 182 , 331–351 .
10. Goreau, T. F. ( 1959 ) Ecology 40 , 67–90 .
11. Muscatine, L. & Cernichiari, E. ( 1969 ) Biol. Bull. (Woods Hole, Mass.) 137 , 506–523 .
12. Smith, D. , Muscatine, L. & Lewis, D. ( 1969 ) Biol. Rev. 44 , 17–90 .
13. MacIntyre, I. & Towe, K. ( 1976 ) Science 193 , 701–702 .
14. Ma, T. ( 1934 ) Proc. Imp. Acad. (Tokyo) 10 , 353–356 .
15. Buddemeier, R. , Maragos, J. & Knutson, D. ( 1974 ) J. Exp. Mar. Biol. Ecol. 14 , 179–200 .
16. MacIntyre, I. & Smith, S. ( 1974 ) Proc. Int. Symp. Coral Reefs, 2nd. ( Great Barrier Reef Committee , Brisbane, Australia ), pp. 277–287 .
17. Knutson, D. W. , Buddemeier, R. W. & Smith, S. V. ( 1972 ) Science 177 , 270–272 .
18. Buddemeier, R. & Kinzie, R. ( 1975 ) in Growth Rhythms and the History of the Earth’s Rotation , eds. Rosenberg, G. D. & Runcorn, S. K. ( Wiley , London ), pp. 135–147 .
19. Druffel, E. R. M. ( 1987 ) J. Mar. Chem. 45 , 667–698 .
20. Dodge, R. & Brass, G. ( 1984 ) Bull. Mar. Sci. 34(2) , 288–307 .
21. Hudson, J. , Shinn, E. A. , Halley, R. B. & Lidz, B. ( 1976 ) Geology 4 , 361–364 .
22. Glynn, P. & Wellington, J. ( 1990 ) in Global Ecological Consequences of the 1982/83 El Niño Southern-Oscillation , ed. Glynn, P. ( Elsevier , New York ).
23. Wyrtki, K. , Stroup, E. , Patzert, W. , Williams, R. & Quinn, W. ( 1976 ) Science 191 , 343–346 .
24. Carriquiry, J. , Risk, M. & Schwarcz, H. ( 1988 ) Palaios 3 , 359–364 .
25. Fergusson, C. ( 1968 ) Science 159 , 839–846 .
26. Dodge, R. & Thomson, J. ( 1974 ) Earth Planet. Sci. Lett. 23 , 313–322 .
27. Buddemeier, R. ( 1974 ) Proc. Int. Symp. Coral Reef 2nd. ( Great Barrier Reef Committee , Brisbane, Australia ), pp. 259–267 .
28. Edwards, R. L. , Chen, J. H. & Wasserburg, G. J. ( 1986/7 ) Earth Planet. Sci. Lett. 81 , 175–192 .
29. Stoddart, D. ( 1969 ) Biol. Rev. 44 , 433–498 .
30. Jokiel, P. L. & Coles, S. L. ( 1977 ) Mar. Biol. 43 , 201–208 .
31. Highsmith, R. ( 1979 ) J. Exp. Mar. Biol. Ecol. 37 , 105–125 .
32. Glynn, P. & Wellington, G. ( 1983 ) Corals and Coral Reefs of the Galaagos Islands ( Univ. of California Press , Berkeley ).
33. Bak, R. P. M. ( 1974 ) Proc. Int. Symp. Coral Reefs 2nd. ( Great Barrier Reef Committee , Brisbane, Australia ), pp. 229–233 .
34. Goreau, T. F. ( 1963 ) Ann. N.Y. Acad. Sci. 109 , 127–167 .
35. Goreau, T. & Goreau, N. ( 1959 ) Biol. Bull. Mar. Biol. Lab. Woods Hole, Mass. 117 , 239–250 .
36. Coles, S. L. & Jokiel, P. L. ( 1978 ) Mar. Biol. 49 , 187–195 .
37. Baker, P. & Weber, J. ( 1975 ) Earth Planet. Sci. Lett. 27 , 57–61 .
38. Dodge, R. , Aller, R. & Thompson, J. ( 1974 ) Nature (London) 247 , 574–577 .
39. Buddemeier, R. & Kinzie, R. ( 1976 ) Oceanogr. Mar. Biol. 14 , 183–225 .
40. Easton, W. & Olson, E. ( 1976 ) Geol. Soc. Am. Bull. 87 , 711–719 .
41. Moore, W. & Krishnaswami, S. ( 1972 ) Earth Planet. Sci. Lett. 15 , 187–190 .
42. Shen, G. T. & Boyle, E. A. ( 1988 ) Chem. Geol. 67 , 47–62 .
43. Druffel, E. M. & Linick, T. W. ( 1978 ) Geophys. Res. Lett. 5 , 913–916 .
44. Konishi, K. , Tanaka, T. & Sakanoue, M. ( 1982 ) Proc. Int. Symp. Coral Reef 4th , ed. Gomez, E. D. ( Marine Science Center, Univ. of the Philippines , Manila ).
45. Toggweiler, J. R. , Dixon, K. & Broecker, W. S. ( 1991 ) J. Geophys. Res. 96 , 20 , 467–20 , 497 .
46. Nozaki, Y. , Rye, D. M. , Turekian, K. K. & Dodge, R. E. ( 1978 ) Geophys. Res. Lett. 5 , 825–828 .
47. Purdy, C. , Druffel, E. R. M. & Livingston, H. ( 1989 ) Geochim. Cosmochim. Acta 53 , 1401–1410 .
48. Moore, W. , Krishnaswami, S. & Bhat, S. ( 1973 ) Bull. Mar. Sci. 23(2) , 157–176 .
49. Toggweiler, J. R. & Trumbore, S. E. ( 1985 ) Earth Planet. Sci. Lett. 74 , 306–314 .
50. Benninger, L. & Dodge, R. ( 1986 ) Geochim. Cosmochim. Acta 50 , 2785–2797 .
51. Broecker, W. & Thurber, D. ( 1965 ) Science 149 , 59–61 .
52. Ku, T.-L. ( 1968 ) J. Geophys. Res. 73(6) , 2271–2276 .
53. Edwards, R. L. , Beck, J. W. , Burr, G. , Donahue, D. , Chappell, J. M. A. , Bloom, A. L. , Druffel, E. R. M. & Taylor, F. M. ( 1993 ) Science 260 , 962–967 .
54. Bard, E. , Hamelin, B. , Fairbanks, R. G. , Zindler, A. & Mathieu, G. ( 1990 ) Nucl. Instrum. Methods Phys. Res. Sect. B 52 , 461–468 .
55. McConnaughey, T. ( 1989 ) Geochim. Cosmochem. Acta 53 , 163–171 .
56. Weber, J. ( 1974 ) Proc. Int. Symp. Coral Reef 2nd. ( Great Barrier Reef Committee , Brisbane, Australia ), pp. 289–298 .
57. Land, L. S. , Lang, J. C. & Smith, B. N. ( 1975 ) Limnol. Oceanogr. 20 , 283–287 .
58. Weil, S. , Buddemeier, R. W. , Smith, S. V. & Kroopnick, P. M. ( 1981 ) Geochim. Cosmochim. Acta 45 , 1147–1153 .
59. McConnaughey, T. ( 1989 ) Geochim. Cosmochem. Acta 53 , 151–162 .
60. Emiliani, C. , Hudson, J. H. , Shinn, E. A. & George, R. Y. ( 1978 ) Science 202 , 627–629 .
61. Swart, P. K. ( 1983 ) Earth Sci. Rev. 19 , 51–80 .
62. Druffel, E. R. M. , King, L. L. , Belastock, B. A. & Buesseler, K. O. ( 1990 ) Geochim. Cosmochim. Acta 54 , 1493–1500 .
63. Smith, S. & Kroopnick, P. ( 1981 ) Nature (London) 294 , 252–253 .
64. Cole, J. ( 1992 ) Interannual-Decadal Variability in Tropical Climate Systems: Stable Isotope Records and General Circulation Model Experiments ( Columbia University , New York ).
65. Fairbanks, R. G. & Dodge, R. E. ( 1979 ) Geochim. Cosmochim. Acta 43 , 1009–1020 .
66. Epstein, S. & Mayeda, I. ( 1953 ) Geochim. Cosmochim. Acta 4 , 213–224 .
67. Weber, J. & Woodhead, P. ( 1970 ) Chem. Geol. 6 , 93–117 .
68. Weber, J. & Woodhead, P. ( 1972 ) J. Geophys. Res. 77 , 463–473 .
69. Druffel, E. R. M. ( 1985 ) Geophys. Monogr. 32 , 111–122 .
70. Dunbar, R. B. & Wellington, G. M. ( 1981 ) Nature (London) 293 , 453–455 .
71. Cole, J. , R. Fairbanks, & Shen, G. ( 1993 ) Science 260 , 1790–1793 .
72. Swart, P. & Coleman, M. ( 1980 ) Nature (London) 283 , 557–559 .
73. Knutson, D. & Buddemeier, R. ( 1973 ) in Radiocarbon Contamination of the Marine Environment ( International Atomic Energy Agency , Vienna ), p. 735 .
74. Druffel, E. R. M. & Suess, H. E. ( 1983 ) J. Geophys. Res. 88(C2) , 1271–1280 .
75. Druffel, E. R. M. & Griffin, S. ( 1995 ) Radiocarbon 37(2) , 517–524 .
76. Stuiver, M. & Polach, H. A. ( 1977 ) Radiocarbon 19(3) , 355–363 .
77. Buddemeier, R. , Schneider, R. & Smith, S. ( 1981 ) Proc. Int. Symp. Coral Reef 4th 2 , 81–85 .
78. Shen, G. & Stanford, C. ( 1990 ) in Global Ecological Consequences of the 1982–83 El Niño-Southern Oscillation , ed. Glynn, P. ( Elsevier , New York ), pp. 255–283 .
79. Schneider, R. & Smith, S. ( 1982 ) Mar. Biol. 66 , 121–131 .
80. Beck, J. , Edwards, R. L. , Ho, E. , Taylor, F. W. , Recy, J. , Rougeri, F. , Joannot, P. & Henin, C. ( 1992 ) Science 257 , 644–647 .
81. DeVilliars, S. , Shen, G. T. & Nelson, B. T. ( 1995 ) Geochim. Cosmochim. Acta 58 , 197–208 .
82. Flor, T. & Moore, W. ( 1977 ) in Proc. Int. Symp. Coral Reef 3rd ( Rosenstiel School of Marine and Atmospheric Sciences, University of Miami , Miami ), pp. 555–561 .
83. Shen, G. & Dunbar, R. ( 1994 ) Geochim. Cosmochim. Acta 59 , 2009–2024 .
84. Shen, G. , Boyle, E. & Lea, D. ( 1987 ) Nature (London) 328 , 794–796 .
85. St. John, B. ( 1974 ) Proc. Int. Symp. Coral Reef 2nd. 2 , 461–469 .
86. Livingston, H. & Thompson, G. ( 1971 ) Limnol. Oceanogr. 16 , 786–796 .
87. Lea, D. , Shen, G. & Bole, E. ( 1989 ) Nature (London) 340 , 373–376 .
88. Linn, L. J. , Delaney, M. L. & Druffel, E. R. M. ( 1990 ) Geochim. Cosmochim. Acta 54 , 387–394 .
89. Shen, G. , Campbell, T. M. , Dunbar, R. B. , Wellington, G. M. , Colgan, M. W. & Glynn, P. W. ( 1991 ) Coral Reefs 10 , 91–101 .
90. Delaney, M. L. , Druffel, E. R. M. & Linn, L. ( 1993 ) Geochim. Cosmochim. Acta 57 , 347–354 .
90. Swart, P. ( 1981 ) Palaeogeogr. Palaeoclimatol. Palaeoecol. 34 , 115–136 .
92. Oomori, T. , Kaneshima, T. K. , Nakamura, Y. & Kitano, Y. ( 1983 ) Galaxea 1 , 77–86 .
93. Sholkovitz, E. & Shen, G. ( 1995 ) Geochim. Cosmochim. Acta 59(1) , 2749–2756 .
94. Simkiss, K. ( 1964 ) Biol. Rev. 39 , 487–505 .
95. Dodge, R. , Jickells, T. D. , Knap, A. H. , Boyd, S. & Bak, R. P. M. ( 1984 ) Mar. Poll. Bull. 15(5) , 178–187 .
96. Rasmussen, C. Proc. Int. Symp. Coral Reef 6th ( 1988 ) ( Coral Reef Symposium Executive Committee , Townsville, Queensland, Australia ), pp. 325–330 .
97. Dodge, R. & Vaisnys, J. ( 1975 ) Nature (London) 258 , 706–708 .
98. Dodge, R. , Turekian, K. & Vainys, J. ( 1977 ) Proc. Int. Symp. Coral Reef 3rd ( Rosenstiel School of Marine Atmospheric Sciences, University of Miami , Miami ), pp. 361–365 .
99. Shen, G. , Cole, J. E. , Lea, D. W. , Linn, L. J. , McConnaughey, T. A. & Fairbanks, R. G. ( 1992 ) Paleoceanography 7(5) , 563– 588 .
100. Shen, G. , Linn, L. J. , Campbell, T. M. , Cole, J. E. & Fairbanks, R. G. ( 1992 ) J. Geophys. Res. 97(C8) , 12689–12697 .
101. Jenkins, W. J. ( 1982 ) J. Mar. Res. 40, (Suppl.) 265–290 .
102. Druffel, E. R. M. ( 1989 ) J. Geophys. Res. 94 , 3271–3285 .
103. Chakraborty, S. & Ramesh, R. ( 1993 ) Terra Nova 5 , 545–551 .
104. Isdale, P. ( 1984 ) Nature (London) 310 , 578–579 .
105. Susic, M. , Boto, K. & Isdale, P. ( 1991 ) Mar. Chem. 33 , 91–104 .
106. Dunbar, R. B. , Wellington, G. M. , Colgan, M. W. & Glynn, P. W. in Proceedings of Seventh Annual Pacific Climate (PACLIM) Workshop, Int. Ecol. Stud. Prid. Technol. , eds. Betancourt, J. L. & Tharp, V. L. ( California Dept. of Water Resources Press , Los Angeles ), pp. 183–198 .
107. Cole, J. E. ( 1996 ) in Climatic Fluctuations and Forcing: Mechanisms of the Last 2000 Years , eds. Jones, P. D. , Bradley, R. S. & Jouzel, J. ( Springer , Berlin ), pp. 333–355 .
108. Patzold, J. & Wefer, G. ( 1992 ) Fourth International Conference on Paleoceanography ( Kiel , Germany ), pp. 224–225 .
109. Toggweiler, J. , Dixon, K. & Bryan, K. ( 1989 ) J. Geophys. Res. 94(C6) , 8217–8242 .
110. Druffel, E. R. M. & Griffin, S. ( 1993 ) J. Geophys. Res. 98 , 20249–20259 .
111. Quinn, W. H. , Neal, V. T. & DeMayolo, S. E. A. ( 1987 ) J. Geophys. Res. 92 , 14449–14461 .
112. Druffel, E. R. M. & Benavides, L. M. ( 1986 ) Nature (London)321, 58–61.
113. Suess, H. E. ( 1953 ) in Proceedings Conference Nuclear Processes in Geological Settings , ( University of Chicago Press , Chicago ), pp. 52–56 .
114. Druffel, E. ( 1997 ) Science 275 , 1454–1457 .
115. Quay, P. D. , Tilbrook, B. & Wong, C. S. ( 1992 ) Science 256 , 74–79 .
116. Griffin, S. & Druffel, E. R. M. ( 1989 ) Radiocarbon 31 , 533–542 .
117. Smith, J. ( 1993 ) Isotopes in a Deep-Sea Coral from Orphan Knoll ( McMaster University, Hamilton , Ontario, Canada ). Dissertation .
118. Druffel, E. , Griffin, S. , Witter, A. , Nelson, E. , Southon, J. , Kashgarian, M. & Vogel, J. ( 1995 ) Geochim. Cosmochim. Acta 59 , 5031–5036 .
119. Adkins, J. & Boyle, E. ( 1994 ) Eos 75(44) , 347 .
120. Adkins, J. , Boyle, E. A. , Cheng, H. , Edwards, R. L. & Druffel, E. R. M. ( 1995 ) Eos 76(46) , F290 .








