
Protein and Amino Acids, 1999
Pp. 169-216. Washington, D.C.
National Academy Press
9
Inherent Difficulties in Defining Amino Acid Requirements
D. Joe Millward1
INTRODUCTION
To address the role of protein and amino acids in performance, this chapter is based on the premise that it is an inherently difficult problem to define the dietary requirements of human adults for indispensable amino acids and to assess the nutritional value (protein quality) of different food protein sources to provide for those needs. There are three major reasons for this difficulty. The first is adaptation, that is, a variable metabolic demand for amino acids set by the habitual intake. Thus, the extent to which any intake appears to be adequate will depend on the completeness of adaptation to that intake. The second is methodology, with no entirely satisfactory practical nitrogen or amino acid balance method or other measure of dietary adequacy. The third is lack of quantifiable, unequivocal metabolic indicators of adequacy that can validate
balance measurements. The questions posed in this review cannot currently be answered because of the absence of studies of outcome in terms of physical performance in long-term, controlled feeding trials.
BACKGROUND TO THE CURRENT CONTROVERSY
In 1985, the Food and Agriculture Organization (FAO) report on protein and energy requirements was published (FAO/WHO/UNU, 1985), a feature of which was the recommendation that protein quality should be evaluated by the PDCAAS method (protein-digestibility corrected amine acid score), making use of age-specific amine acid scoring patterns. Because the indispensable amine acid (IAA) requirement values used to calculate the scoring patterns fell markedly with age, from over 50 percent of total protein requirement in infants to only 16 percent in adults (see Table 9-1), the quality of any protein would now be assessed as higher when used for adults than for children. Furthermore the low requirement level of IAA in adults meant that all natural diets and food proteins would be adequate. Thus, apart from digestibility, protein quality ceased to be an issue in the nutrition of adults.
Considerable disquiet arose about the 1985 report. Young (1986) argued that the adult IAA requirement values were seriously flawed because of the way Rose (1957) conducted his nitrogen (N) balance studies (mainly excess energy and no account for miscellaneous N losses). Millward and Rivers (1988) reviewed the subject, paying particular attention to the adaptive changes in amine acid oxidation that can occur and that will influence requirement values. They argued that the marked fall with age in the requirement values was mainly a reflection of the methodologies used in their assessment. Thus, the infant values were largely patterned on the composition of breast milk, while the adult values, measured in balance studies with excess nonessential nitrogen and low levels of indispensable amine acid, would have identified minimum requirement values. They concluded that IAA requirements are complex, include an adaptive component, and can only be defined under specific artificial conditions that would allow definition of a minimum value and that ''current estimates of adult requirements may be close to this level." To identify which IAA might be rate limiting for the obligatory N losses (ONL), they calculated the obligatory oxidative amine acid losses (OOL) as estimates of the losses of tissue IAAs that would give rise to the ONL, as discussed in detail below.
Young et al. (1989) then published a paper entitled "A Theoretical Basis for Increasing Current Estimates of the Amine Acid Requirements in Adult Man with Experimental Support." This paper reproduced the table of eel values from Millward and Rivers (1988). After making some small adjustments in lysine, threenine, and valine values derived from their stable isotope studies and increasing all values assuming a 70 percent efficiency of utilization, Young and colleagues proposed that this pattern, the "MIT" (Massachusetts Institute of
TABLE 9-1
Protein and Indispensable Amino Acid Requirements (mg/kg/d) and Obligatory Indispensable Amino Acid Oxidative Losses*
|
|
|
|
|
|
Obligatory oxidative losses† |
|
|
|
Infants (3-4 too) |
Children (2 y) |
Schoolboys (10-12y) |
Adults |
Value |
Requirement multiple |
|
Protein |
|
|
|
|
|
|
|
Growth |
625 |
187 |
1 06 |
— |
|
|
|
Maintenance |
750 |
738 |
681 |
600 |
|
|
|
Total |
1,375 |
925 |
787 |
600 |
338 |
0.56 |
|
Amino acids |
|
|
|
|
|
|
|
Histidine |
28 |
(20) |
(20) |
8-12 |
11.5 |
1.05 |
|
Isoleucine |
70 |
31 |
30 |
10 |
16.2 |
1.62 |
|
Leucine |
161 |
73 |
45 |
14 |
27.4 |
1.96 |
|
Lysine |
103 |
64 |
60 |
12 |
30.1 |
2.51 |
|
TSA |
58 |
27 |
27 |
13 |
13.5 |
1.01 |
|
TAA |
125 |
69 |
27 |
14 |
27 |
1.93 |
|
Threonine |
87 |
37 |
35 |
7 |
15.5 |
2.21 |
|
Tryptophan |
17 |
12.5 |
4 |
3.5 |
4.0 |
1.14 |
|
Valine |
93 |
38 |
33 |
10 |
16.9 |
1.69 |
|
Total |
742 |
372 |
281 |
94 |
162 |
1.72 |
|
% Protein requirement |
54 |
40 |
36 |
16 |
48 |
|
|
NOTE: TSA, total sulfur amino acids; TAA, total aromatic amino acids. * Protein requirements arc mean values † These are the rates of oxidative loss of IAAs predicted to occur based on the assumption that the ONL, (54 mg N/kg/d) derive from the oxidation of amino acids liberated from body protein (amino acid composition as beef), and the composition of the free amino acid pool does not change. SOURCE: Adapted from FAO/WWHO/UNU, 1985. |
||||||
Technology) scoring pattern, should be used as the basis for protein quality evaluation in adults and in children given the similarity between their pattern and that of the FAO preschool child pattern.
In 1989, FAO/WHO convened a meeting to consider protein quality evaluation and to endorse the PDCAAS method recommended by FAO in 1985. However the report (FAO/WHO, 1991) rejected both the 1985 adult and older school child IAA requirement values as flawed, was unable to identify any other appropriate adult scoring pattern, and proposed that the scoring pattern for the preschool child be utilized for older children and for adults as a strict interim measure. It was argued that (a) the preschool child data were reliable, (b) in the absence of any other data, some pattern was needed for older children and adults, and (c) the slow growth of children compared with adults means that a major change in the requirement pattern with age was unlikely.
Although Young and colleagues broadly agreed with this conclusion in that the MIT and preschool patterns were similar, Millward (1994) argued that the report was flawed. In fact, the data that formed the basis of the preschool child pattern had never been published and were not available for scrutiny except for some "typical" data for lysine published in a book review (Pineda et al., 1981). The data, which were derived from study of preschool children who had recovered from protein energy malnutrition (PEM), show that the N balances were so large in the children studied that they would have been exhibiting catchup growth as far as lean tissue was concerned (growth rates and N retentions of 3 times the expected values). This growth would markedly increase the need for indispensable amine acids compared with that of normal preschool children, older children, and especially adults. Millward (1994) also argued against acceptance of the MIT scoring pattern on the grounds that amine acid requirements for maintenance cannot be predicted from the amine acid composition of body proteins.
Fuller and Garlick (1994) have reviewed the controversy, and while they did not endorse the MIT pattern, they did conclude that the FAO values were likely to be underestimates, having failed to include miscellaneous N losses in the original balance studies. They reported adjusted higher values, taking into account estimated miscellaneous N losses. They also raised concern that the tracer studies may also suffer from an underestimate of losses and that neither N nor tracer studies are inherently better than the other. These issues are considered in more detail below. Waterlow (1996) made a detailed analysis of the tracer studies and came to conclusions similar to Fuller and Garlick, that is, they did not accept the theoretical basis of the MIT pattern but recognized that the 13C leucine studies do point to a higher leucine requirement than does the FAO value. The issue was considered at an international meeting of an expert group in London in 1994. However, contrary to what was published (Clugston et al., 1996), the MIT pattern was not endorsed at this meeting since, as subsequently reported by Millward and Waterlow (1996), the published statement "a large majority of the group accepted as an interim operational
pattern that [that was] proposed by Young et al.," had in fact emerged during postmeeting editing.
The current views of Young and colleagues are described in the chapter following this one. What follows is an account of this author's current understanding of the debate. Much of the argument made here has been reported previously in publications by the author and in correspondence relating to publications by Young and colleagues (Millward and Rivers, 1988, 1989; Millward, 1990, 1991, 1992, 1993, 1994; Millward et al., 1989, 1990; Millward and Pacy, 1995).
METABOLIC BACKGROUND
One cause of potential confusion within this debate is lack of consistency in terminology. To avoid that here, protein and amino acid requirements will be discussed in terms of metabolic demand, dietary requirement, and dietary allowances. Metabolic demand (MD) is determined by the nature and extent of those metabolic pathways that consume amino acids. The dietary requirement is the amount of protein and/or its constituent amino acids that must be supplied in the diet to satisfy the metabolic demand, usually greater than the MD because of less-than-perfect protein utilization. Dietary Reference Values (U.K. terminology) or Recommended Dietary Allowances (U.S. terminology) are a range of intakes derived from estimates of individual requirements that are designed to meet the dietary requirements of the population and that take into account the variability among individuals in that dietary requirement. This chapter focuses on the MD and dietary requirement.
Obligatory Metabolic Demand
The MD for dietary protein is to provide amino acid precursors for the synthesis of tissue proteins and a range of nonprotein products. Although most proteins are in a dynamic state of constant turnover, little metabolic demand for amino acids is generated by this avenue because of amino acid recycling. "Wear and tear" as a driver of MD is not an appropriate biological analogy. Only net protein synthesis contributes to MD. This growth aspect of MD is straightforward in that the qualitative nature of MD is determined by the amino acid pattern of tissue protein deposited. This pattern is usually assumed to be influenced only by the changes in body composition occurring during growth and is not generally assumed to vary with the diet within cells and tissues. However, as discussed by Fuller and Garlick (1994), some evidence exists for changes in amino acid content of tissues during growth on amino acid-limiting diets.
Nonprotein products of amino acid precursors include nucleic acids, and a range of smaller molecules such as creatine, taurine, glutathione, hormones
(e.g., catecholamines and thyroxine), neurotransmitters (serotonin, dopamine), and nitric oxide, a key regulator of blood flow and other physiological processes.
In human nutrition, growth occurs very slowly after the first few months of life. Net protein synthesis contributes a small and decreasing component of MD during pre- and immediate post-adolescence. In the adult, it comprises only that associated with continuing growth of skin and hair and the synthesis of those gastric secretions (e.g., threonine-rich mucus glycoproteins) that pass into the colon to be utilized for bacterial metabolism. Thus, apart from these small components, humans are normally at nitrogen or amine acid equilibrium, with MD reflecting mainly nonprotein pathways of amine acid metabolism and catabolism associated with maintenance of normal function and composition. In the traditional nutritional terminology of human growth and maintenance, growth needs are low at all ages after early infancy with maintenance dominating the MD. The task, then, is to define the amounts and amine acid pattern of the maintenance requirement.
Obligatory Metabolic Demands and Obligatory Oxidative Losses
The diverse obligatory maintenance MDs for amine acids represent an important, but small, intrinsic part of MD, the magnitude of which is the main subject of current debate. Table 9-1 shows the requirement values for IAAs and for protein as reported by the FAO (FAO/WHO/UNU, 1985). The feature of these values that has been at the heart of the controversy is that the IAA requirement as a proportion of the protein requirement falls markedly with age, from 54 percent in infants to 16 percent in adults. Millward and Rivers (1988) argued that some information could be obtained from the magnitude of the ONL. In subjects fed a protein-free but otherwise nutritionally adequate diet, N losses fall to a stable and reproducible low level after 7 to 14 days. Subjects lose body protein at a constant daily rate, about 54 mg/kg/day, which is equivalent to 0.34 g of protein/kg/day (FAO/WHO/UNU, 1985). These ONLs are assumed to represent nitrogen end products of amine acids derived from body protein and utilized for the obligatory metabolic demand (OMD) that is tacitly assumed to be the same in subjects consuming a protein-free diet as in subjects consuming a normal diet. The ONL is a function of body weight and, when normalized to "metabolic body size" (kg0.75), varies little with age (FAO/WHO/UNU, 1985). Millward and Rivers (1988) reported a simple calculation of the eels. These are the oxidation rates of amine acids that give rise to the ONL and they are equal to the amounts of amine acids in the protein equivalent of the ONL. It was assumed that tissue protein composition could be approximated by that of muscle and the values for beef muscle listed by FAO (FAO/WHO/UNU, 1985) were used. This was only a first approximation calculation, since some of the ONL occurs as protein per se (skin, hair, some fecal nitrogen, and secretions), and protein is lost from several tissues in addition to skeletal muscle. They made
the assumption that of the individual amino acids that comprise the obligatory MD (which, on a protein-free diet, is in effect fueled by tissue protein), one amino acid would be rate limiting, with the highest ratio of obligatory MD to OOL. All other amino acids with a lower ratio would be present in excess in the OOL but would be nevertheless oxidized because they could not be returned on their own to the tissue protein pool. They argued that if protein turnover is tightly regulated, allowing just enough of the rate-limiting amino acid to be withdrawn from tissue protein to provide for its MD, the OOL of this amino acid should be a reasonable guide to its requirement. For all others, the values for the OOL should be greater than the maintenance requirements.
Use of the Obligative Oxidative Loss Pattern to Predict a Requirement Pattern
The actual listed values of the OOL were adopted by Young et al. (1989) as the basis of the MIT scoring pattern, something that was contrary to what Millward and Rivers (1988) intended. In effect, this defined a maintenance pattern with the same composition as tissue protein, a novel assumption given the widespread assumption of different amino acid patterns for maintenance and growth.
Millward and Rivers (1988) assumed that the pattern of the obligatory MD is different from that of tissue protein, so that there would be a rate-limiting amino acid that "drives" the ONL. The identification of this driver can be done by reference to an actual requirement pattern. Thus, they compared the values of OOL with the 1985 FAO/WHO/UNU requirement values such that if the FAO values were accurate, the values would be similar for one amino acid. In fact, while the OOL for most amino acids was greater than the FAO requirement, with lysine, threonine, leucine, and the aromatic amino acids being particularly in excess (2-2.5 times the requirement), the OOL of the total sulfur amino acids (TSAs) was quite close to the FAO requirement values; this latter observation showed that the TSAs are rate determining for the mobilization of tissue protein to provide for obligatory MD. The possibility that the TSAs are the rate-limiting amino acids that drive the ONL in humans was attractive on the basis of animal studies. Providing the rate-limiting amino acid to the protein-free diet fed during measurement of ONL should result in a fall in N excretion to the rate determined by the demand for the second limiting amino acid. Studies in dogs (Allison et al., 1947) and rats (Yoshida, 1983) have shown that supplementation by S amino acids reduces N excretion.
Animal Data for the Pattern of the Obligatory Metabolic Demand
Animal data clearly indicate that the amino acid pattern of the obligatory maintenance MD differs from that of tissue protein, which necessarily repre-
sents the pattern for the growth requirement. However, this view is not accepted by Young (e.g., Young and El-Khoury, 1995), who, having used the OOL pattern to derive the MIT pattern, assumed maintenance requirements to be broadly similar to tissue protein.
Before reviewing the animal data, their relevance to human nutrition needs to be addressed. Young and El-Khoury (1995) have discussed the relevance of the high-quality nitrogen balance data obtained in the young pig (Fuller et al., 1989). They argued that any comparison of human requirement with that of the young pig is invalid because (a) relative amounts of maintenance and growth vary too much between the species (maintenance = < 10 percent total [growth + maintenance] requirement in the young pig and < 5 percent total in the rat), and (b) the efficiency of dietary protein utilization in the young pig at maintenance is much higher (100 percent) compared with human values (assumed to be 70 percent). In fact, the first point is irrelevant when human-animal comparisons are limited to discussions of either maintenance or growth needs specifically, and the second point is irrelevant when what is considered is MD rather than dietary requirement.
Young also argues that data from adult pigs (boars) support the proposition that the pattern of tissue protein is similar to the maintenance pattern. Yet he acknowledges the data to be poor and at variance with most other animal data.
Finally, Young and El-Khoury (1995) argue that maintenance patterns derived from rapidly growing animals held at maintenance by food restriction are unphysiological and, consequently, may be an unreliable guide to the human obligatory IAA MD. To date there has yet to be a claim that human IAA bid can be accurately predicted from animal values, only that consideration of the animal data as a whole, including both growing and adult data, may provide useful general information. There appear to be few major differences between mammalian species with regard to the fundamentals of amine acid and protein metabolism. With obvious exceptions (e.g., arginine requirements for growing cats and growing and adult dogs, a taurine requirement for the kittens, and a high-maintenance amine acid requirement in avian species for feather growth), interspecies comparisons appear to be legitimate since robust animal data should provide general principles about the nature of human needs.
Two kinds of studies are pertinent. The first is deletion studies, in which individual amine acids are removed from the diet and the extent of the negative balance is monitored. If the maintenance requirement patterns corresponded exactly to the patterns of tissue protein, then there should be a similar negative balance on removal of each IAA. If not, then negative balance will occur in proportion to the ratio of obligatory MD to tissue content of each amine acid.
Only one report exists for the adult rat (Said and Hegsted, 1970), a high-quality study based on measured changes in body water. Gahl et al. (1991) reported N balance data for young rats, while Fuller et al. (1989) studied 41 kg pigs, with N balance; data from the latter study are widely recognized as the most robust data for the pig.
Table 9-2 shows the relative losses normalized for the response to a protein-free diet. The first, second, and third limiting amino acids are threonine, TSA, and isoleucine for the growing rat; threonine, isoleucine, and tryptophan for the adult rat; and TSA, threonine, and tryptophan for the pig. The most highly conserved, least-limiting amino acids are lysine and leucine in the rat and all three branched-chain amino acids (BCAAs) and lysine in the pig.
Supplementation with each limiting amino acid allows the slope of the balance curve to be established and the consequent requirement values to be determined; these are shown in Table 9-3 compared with carcass protein content. For ease of comparison, these patterns have been normalized for threonine. Leucine and lysine are the two most abundant amino acids in carcass proteins and in the growth requirement patterns for both rat and pig; in the maintenance requirement patterns, the most abundant amino acids are threonine and TSA in the pig; threonine, isoleucine, valine, and TSA in both adult and growing rats.
TABLE 9-2 Responses (Negative Balance) to Deletion of Individual Indispensable Amino Acids or a Protein-Free Diet
|
Amino acid |
Growing pig* |
Growing rat† |
Adult rat‡‡ |
|
Histidine |
nd |
0.31 |
0.55 |
|
Isoleucine |
0.24 |
0.94 |
0.90 |
|
Leucine |
0.21 |
0.38 |
0.36 |
|
Lysine |
0.33 |
0.25 |
0.30 |
|
TSA |
0.93 |
1.09 |
0.51 |
|
TAA |
0.34 |
0.06 |
0.47 |
|
Threonine |
0.68 |
1.19 |
0.92 |
|
Tryptophan |
0.53 |
0.75 |
0.80 |
|
Valine |
0.23 |
0.88 |
0.71 |
|
All (protein free) |
1.00 |
1.00 |
1.00 |
|
NOTE: TAA, total aromatic amino acids; TSA, total sulfur amino acid; nd, not determined. * Fuller et hi. (1989). † Gahl et al. (1991). ‡ Said and Hegsted (1970). |
|||
TABLE 9-3 Amino Acid Composition and Requirement Patterns
|
|
Growing pig* |
|
Growing rat† |
|
Adult rats‡ |
|||
|
|
Body |
G |
M |
Body |
G |
M |
G |
M |
|
Histidine |
0.74 |
— |
0.65 |
0.34 |
0.14 |
0.41 |
0.48 |
— |
|
Isoleucine |
0.92 |
0.92 |
0.30 |
0.81 |
1.07 |
0.56 |
1.08 |
1.03 |
|
Leucine |
1.88 |
1.66 |
0.43 |
1.67 |
1.61 |
0.38 |
1.35 |
0.94 |
|
Lysine |
1.87 |
1.45 |
0.68 |
1.58 |
1.23 |
0.33 |
1.76 |
0.74 |
|
TSA |
0.74 |
0.76 |
0.92 |
0.87 |
1 .40 |
0.63 |
0.98 |
0.97 |
|
TAA |
1.89 |
1.80 |
0.70 |
1.67 |
1.25 |
0.20 |
1.41 |
1.15 |
|
Threonine |
1.00 |
1.00 |
1.00 |
1.00 |
1.00 |
1.00 |
1.00 |
1.00 |
|
Tryptophan |
nd |
0.26 |
0.21 |
0.26 |
0.16 |
0.06 |
0.22 |
0.22 |
|
Valine |
1.25 |
1.12 |
0.38 |
1.03 |
1.27 |
0.66 |
1.10 |
1.03 |
|
NOTE: G, growth; M, maintenance; TSA, total sulfur amino acids; TAA, total aromatic amino acids; nd, not determined. * Fuller et al. (1969). † Bencvenga et al. (1994). ‡ Said and Hegsted (1970). |
||||||||
The major implication of these animal data is that there are marked differences between the MD for maintenance and for growth. It is clear that in the growing rat and pig and the adult rat, leucine and lysine exhibit the biggest difference between growth and maintenance patterns, these two amine acids being most abundant for growth and among the least abundant for maintenance. The practical consequence of this, as pointed out by Hegsted (1973), is that the balance-intake curve is extremely shallow for leucine and lysine both in the sub-maintenance and growth range. This means that small differences in balance result in large differences in maintenance intakes so that measurement of a requirement value for maintenance is very difficult and depends on the exact criterion for adequacy. The several early reports of rats maintaining body weight for 6-month periods on very low lysine diets (e.g., zein [Osborne and Mendel, 1916] or even lysine-free diets [Bender, 1961]) are probably explained by coprophagy, given the clear evidence of a metabolic need for lysine in terms of the rapid onset of symptoms on a lysine-free diet in humans (Rose, 1957). However, no evidence exists for anything other than a low metabolic need for this amine acid.
A second type of study which is pertinant is the work of Yoshida (1983) who has done most to explore the concept that rate-limiting amine acids at maintenance differ from those that rate-limit growth. Having established that in
adult rats fed a protein-free diet, the most rate-limiting amino acids were threonine and TSA, he also showed that in adult rats fed limiting amounts of rice or wheat diets, the limiting amino acids were threonine and the sulfur amino acids. When these two amino acids were added to the cereal diets, they restored nitrogen balance and transformed body weight loss to growth (See Figure 9-1). This may explain why attempts to show in human adult supplementation trials that lysine is the limiting amino acid in wheat were so disappointing (Scrimshaw et al., 1973).
Although the nature of the relative metabolic need for individual amino acids is by no means clear, Fuller's work with the pig points to ileal amino acid losses as a partial explanation, accounting for some 60 percent of pig amino acid maintenance requirements (Wang and Fuller, 1989). Table 9-4 compares ileal losses of the pig and humans. These data show that in each case, threonine is the largest component, and while the patterns differ somewhat, the absolute values are much lower in humans than in the pig. Thus, despite discussion of the pig as an inappropriate model for humans, to the extent that ileal losses comprise a component of obligatory ME), these data point to a lower MD in humans than in the pig.
To summarize research on obligatory maintenance MD, a consistent and extensive body of animal data shows the maintenance pattern to differ from the growth pattern, with lower levels of lysine and leucine in the maintenance pattern. As a result, the rate-limiting amino acids in dietary proteins for maintenance may differ from those for growth, the example being that lysine limits wheat for growth but not for maintenance.
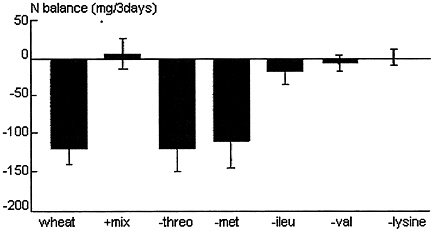
FIGURE 9-1 Adult rats were fed limiting amounts of a wheat diet that did not allow balance and were supplemented with a mixture of indispensable amino acids that did allow balance. By removing individual amino acids, the limiting amino acids were identified as threonine and the sulfur amino acids. Lysine was not needed to improve balance. Note: threo, threonine; met, methionine; lieu, isoleucine; val, valine; N, nitrogen; IAA, indispensable amino acids. Source: Adapted from Yoshida (1983).
TABLE 9-4 Ileal Indispensable Amine Acid Losses of the Pig and Human
|
|
Pig* 40 kg mg/kg/d |
Human adult† 66 kg mg/kg/d |
|
Isoleucine |
11.3 |
1.7 |
|
Leucine |
25 |
3.2 |
|
Lysino |
15.3 |
3.9 |
|
TSA |
18.3 |
1.8 |
|
TAA |
28.5 |
3.9 |
|
Threonine |
32.3 |
4.2 |
|
Valine |
18.3 |
2.9 |
|
NOTE: TSA, total sulfur amine acid; TAA, total aromatic amine acid. * Wang and Fuller (1989). † Fuller et al. (1994). |
||
The Adaptive Component of the Metabolic Demand
It may appear a simple task to identify and quantify the maintenance MD for each amine acid in terms of the various metabolic pathways involved. However, difficulty arises in distinguishing between obligatory intrinsic, functionally important demands and those that serve a purpose, but can and do vary according to circumstances, that is, an adaptive component It is this adaptive component that brings complexity to the maintenance MD.
The ONL at 54 mg N or 0.34 g protein/kg/day, is only 50 percent of current estimates of the protein requirement (0.6 g/kg/d), and the nature of this additional need (the difference between 0.34 and 0.6 g protein) has in the past been difficult to account for. Usually, it has been attributed to an inefficiency of utilization, although why proteins such as those in milk, eggs, or meat were not utilized more efficiently was always puzzling. It is much easier to understand the inefficiency of utilization as representing an adaptive component of MD.
When subjects are fed a protein-free diet, their urinary N losses initially reflect their normal dietary protein intake and then falls over 7 to 14 days to reach a low stable output level (see FAO/WHO, 1973). That is, an additional loss of body N occurs on a daily basis for some time before equilibrium is reached at the lower level. This additional daily N loss demonstrates the existence of this adaptive component of the MD. Traditionally, this has been defined as ''the labile protein reserves," which imply that metabolically it was a pool of protein that varied in size with the dietary protein intake. In the rat, liver and visceral protein content does vary directly with dietary protein intake (see Munro, 1964) in support of the labile protein reserve concept. However, no such
protein pool has ever been identified in humans, and more importantly, recent detailed metabolic studies of balance regulation have pointed to an alternative explanation.
Thus, studies of the distribution of urinary N losses between the postabsorptive and postprandial periods, in subjects adapted to a wide range of protein intakes (Price et al., 1994), showed that not only did postprandial losses of N increase with increasing intakes as would be expected, but so did postabsorptive losses. Price and colleagues utilized both N and 13C leucine balance methods to assess the magnitude of the total losses in the postabsorptive and postprandial state in normal adults with protein intakes ranging from a very low to a high level (0.36 g/kg-2.3 g/kg) for 2 weeks. Comparisons of the rate of 13C leucine oxidation and total N excretion (corrected for acute changes in the body urea pool) indicated the two measures were proportional over the entire range of intakes, although leucine oxidation underestimated N excretion by about 25 percent (see below). The results calculated from postabsorptive 13C-1 leucine oxidation are shown in Figure 9-2. The N equivalent of postabsorptive leucine loss is shown calculated as a daily rate and divided between the estimated obligatory MD (54 mg N/kg), with the remainder representing the adaptive MD. Even for the lowest intake, total losses (assumed to be tissue protein and association amine acids) were in excess of the obligatory MD, which indicates that subjects may not have fully adapted to this low intake in the relatively short (2-week) period of the study. However, it is clear that with higher intakes, the adaptive component of the MD varied up to more than twice the obligatory MD.
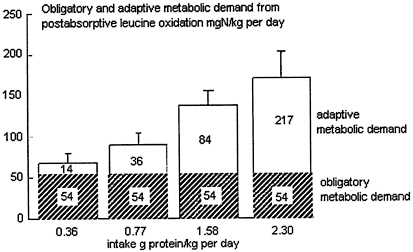
FIGURE 9-2 The magnitude of the obligatory and adaptive metabolic demand in normal adults. Values calculated from postabsorptive leucine oxidation rates measured in subjects who were f ed the indicated protein concentrations for 2 weeks. Source: Adapted from Price et al. (1994).
In other studies of the transition from a high-to-lower protein diet (Quevedo et al., 1994), it was shown that the expected period of negative balance involved a similar lag in reduction of these N losses in both postabsorptive and postprandial states (see Figure 9-3). As a result, there was an overall marked negative balance, and even at the end of 2 weeks, equilibrium had not been reached. Thus, these studies are inconsistent with the concept of labile protein reserves. Instead, they imply that the transitional losses during adaptation to a lower intake reflect the time taken for catabolic pathways of amine acid metabolism to adapt from a level set to deal with one level of protein intake to that required for a lower level.
The implications of these adaptive responses are that the increasing catabolic losses with increasing protein intake represent an actual MD generated in response to the presence of protein in the diet at levels in excess of minimal metabolic needs.
Since amine acids can only be stored as protein, the capacity for which is limited in the body, excess dietary amine acids are oxidized and converted to glucose or fat. As argued by Millward and Rivers (1988), many amine acids, especially the branched-chain, aromatic, and sulfur amine acids, represent a potentially toxic challenge to the organism and are maintained at very low levels in the tissue-free amine acid pools. Thus, after a meal, these amine acids are rapidly removed by oxidative catabolism if not deposited in protein by high
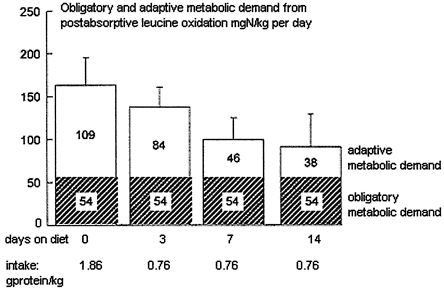
FIGURE 9-3 Rate of change in the adaptive metabolic demand during transition from a high- to lower-protein diet. Because of the slow change in the adaptive metabolic demand after the reduction in protein intake, there was a negative nitrogen balance throughout the 14 days after the diet change. Source: Adapted from Quevedo et at. (1994).
capacity, sensitively regulated pathways. Furthermore, the capacity and activity of these pathways adapt to match the protein levels in the diet to ensure rapid postprandial disposal. Most importantly, because this adaptation of oxidative catabolism to a change of protein intake is relatively slow, the extent of postprandial oxidative catabolism reflects mainly habitual rather than actual protein intake in meals. In effect, the habitual level of protein in the diet creates a level of oxidative amino acid catabolism sufficient to avoid accumulation of toxic concentrations of certain free amino acids, and this becomes part of the MD. Apparently, during slow growth or at weight maintenance, in order to be able to rapidly dispose of dietary protein in excess of minimal needs, pathways of oxidative amino acid catabolism and particular catabolic enzymes are in effect primed to operate at the appropriate rate set by habitual protein intakes. This rate continues regardless of the actual acute intake, utilizing tissue protein if the dietary level falls or during the postabsorptive state, for as long as it takes to adapt to the lower level of intake.
The overall metabolic scheme describing these various aspects of the MD is shown in Figure 9-4. The metabolic fate of dietary protein is shown as providing amino acids into the tissue free amino acid pool, which is in a state of dynamic
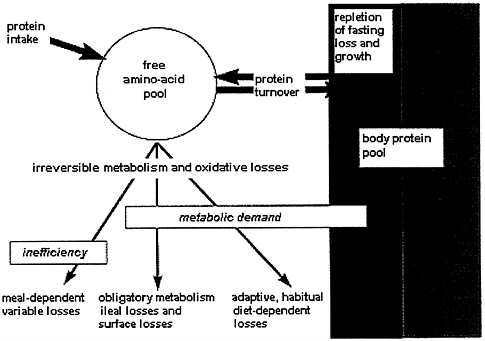
FIGURE 9-4 Scheme to describe the metebolic fate of amino acids in relation to the metabolic demand.
exchange with the protein pool through protein turnover. The MD includes both net protein synthesis (i.e., the repletion of postabsorptive losses and any growth) and irreversible amine acid metabolism and oxidative metabolism. The latter component is shown divided into three parts: (l) the metabolic demands for nonprotein products that eventually give rise to an obligatory N loss, (2) adaptive oxidative amino acid catabolism, which is also part of the MD, and (3) any additional oxidative amine acid catabolism due to an inefficiency of protein utilization. Within this metabolic framework the determination of protein requirements is a problem of assessing both intrinsic, fixed MDs and the adaptive oxidative catabolism, which together define the MD. Assessment of this latter component accounts for the major practical and conceptual difficulties in applying N balance techniques to the study of protein requirements.
Diurnal Cycling: A Qualitative Influence on Metabolic Demand?
There is an important but complex implication of diurnal feeding and fasting for the IAA pattern of the MD. Clearly, from the above discussion, with increasing dietary protein intake, there will be an increasing MD generated by the adaptive oxidative losses. With both adaptive and obligatory MD occurring continuously, overall daily balance is achieved within a complex diurnal cycle of postprandial repletion of tissue proteins mobilized to provide for the postabsorptive demand. Furthermore, the amplitude of this cycle increases with increasing habitual level of protein intake (Price et al., 1994). The key question is: To what extent does this diurnal cycle of body protein influence the IAA composition of the adaptive MD? Does it mean that regardless of the amine acid composition of the intake that induces the adaptive MD, the adaptive MD will have a tissue protein pattern of amine acids to enable postprandial protein deposition? Young and El-Khoury (1995) have assumed this to be the case and a partial justification for their assertion that the maintenance requirement pattern is like that of tissue protein.
In fact, the actual amount of amine acids needed in the diet to provide for this adaptive requirement cannot be predicted for three reasons. First, the amount of actual postabsorptive tissue protein loss depends on the extent of the true postabsorptive state. Price et al. (1994) utilized 12 + 12-hour diurnal cycles in their studies, but the actual amount will vary with the pattern of meal feeding, and individuals consuming both an early breakfast and late supper may spend less than 12 hours in a true postabsorptive state and mobilize less tissue protein. In subjects in the postprandial state, the diet will directly provide for the losses without inducing tissue protein loss.
Second, when true postabsorptive losses of tissue protein do occur it does not follow that all IAAs liberated from the net tissue proteolysis are oxidized. Although increases in the concentrations of the BC, aromatic, and sulfur amine acids liberated from tissue protein are minimized by increasing their oxidation,
this is unlikely to be the case for lysine and threonine. These two amino acids differ from most other IAAs with higher Km2 values for their main oxidative catabolic enzymes, (18 and 52 mM, respectively), larger pool sizes, (about 1 mM) and with less evidence of free control of their catabolic pathways. The difference in the handling of lysine and leucine may well be important in allowing conservation of these amino acids and recycling from postabsorptive losses for postprandial gains.
Evidence for conservation of lysine and threonine comes from muscle biopsy studies after feeding subjects protein (albumin) or a protein-free diet (Bergstrom et al., 1990). These investigators biopsied human muscle at 3 and 7 hours after a meal of 50 g of albumin. They showed that for leucine and lysine, although the intakes were the same and removal of these two amino acids into protein will be at the same rate (since their concentrations in protein are similar), the increase in the concentration of lysine was twice that of leucine, and the same was true for threonine in comparison with valine. By 7 hours, the concentration of all the BCAAs, methionine, and the aromatic amino acids had fallen below the baseline, but for threonine and lysine, there was still an excess of amino acid over the baseline value.
Some indication of how much of these free pools of lysine and threonine might be available to supplement dietary protein that was inadequate in lysine and threonine was indicated by biopsy studies after feeding subjects a protein-free meal. Subsequently, protein deposition would have been entirely dependent on the free amino acid pools (Bergstrom et al., 1990). The fall in the free lysine and threonine pools in muscle were sufficient to have enabled about 250 mg protein deposition per kg body weight. Given that for an adult consuming 0.75 g/kg/d this author's studies indicate that about 50 percent of this is deposited during feeding, (i.e., 380 mg), the free pool of lysine and threonine could in theory contribute two-thirds of the subjects' needs if the dietary source was inadequate. If this is the case, then the organism in overall balance may be less sensitive to poor dietary quality than would be expected in terms of lysine and threonine needs for transient protein deposition.
Evidence for this lack of sensitivity to protein quality is the very small difference in the efficiency of postprandial protein utilization of wheat protein in normal adults compared with that of milk (see Millward et al., 1996). Studies with either frequent small meals (Fereday et al., 1994) or a larger single meal (Fereday et al., 1997) indicate that about 80 percent of dietary wheat protein is deposited in the tissues even though the lysine and threonine content of the wheat is insufficient to allow this. Recycling of the free lysine and threonine liberated in the postabsorptive state allows efficient postprandial protein utilization.
Third, evidence supports that the amplitude of diurnal cycling and the consequent need for postprandial protein deposition is adaptive according to the IAA composition of the diet. Thus, Marchini et al. (1993) showed that postabsorptive leucine balance was 7.8, 9.3, and 12.8 μmol/kg/h for subjects on isonitrogenous diets fed for 3 weeks with FAO patterns (very low IAAs), MIT patterns (intermediate IAAs), or egg patterns (high IAAs) of amine acids, respectively. That is, postabsorptive losses with the FAO pattern fell to < 60 percent of those induced by the egg diet even though the overall level of total amine acid intake was the same. This is an important adaptive response. More recent 24-hour 13C-leucine balances in subjects fed purified amine acid diets that varied only in terms of leucine content (14, 38, and 89 mg/kg/d; El-Khoury et al., 1994a, b) also indicated adaptive reduction in postabsorptive losses, with postabsorptive leucine oxidation rates equivalent to 34.7, 20.9, and 15 mg leucine/kg/12 h. Thus, these results point to substantial adaptive reductions in the amplitude of diurnal cycling and in the consequent metabolic demand for IAAs in response to reductions in dietary protein quality. Therefore, the qualitative influence of diurnal cycling on the metabolic demand cannot be predicted.
Summary of the Metabolic Demand for Indispensable Amine Acids
In normal adults, the ME) for IAAs includes the following:
-
an obligatory component, which in most cases is less than that contained in 0.33 g tissue protein and has a pattern that cannot be predicted from first principles. However, on the basis of animal data, it is likely to contain considerably less lysine and leucine than in the tissue protein pattern.
-
an adaptive component, the amount and composition of which is variable according to the nature and feeding pattern of the habitual dietary protein intake, with a particular adaptive mechanism allowing conservation of lysine and threenine when their intakes are low.
A scheme showing these components of the MD throughout the diurnal cycle is shown in Figure 9-5.
FAO REQUIREMENT VALUES AND N BALANCE STUDIES
The FAO requirement values were derived from N balance studies so that any consideration of these values requires consideration of the N balance technique.
Nitrogen balance studies were initiated in the mid-nineteenth century by Carl Voit, and they have been central to the definition of protein and amine acid
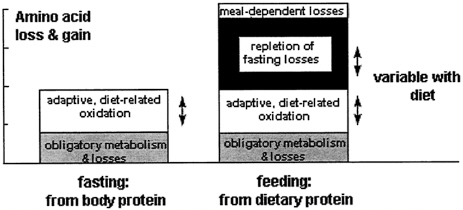
FIGURE 9-5 During fasting, body protein provides for both obligatory and adaptive demands. During feeding, the diet provides for obligatory and adaptive demands, for repletion of tissue protein lost during fasting, and for losses due to inefficiency of utilization. The adaptive components vary markedly with dietary intake, as shown in Figure 9-2, and change only slowly with intake (see Figure 9-3).
requirements ever since. Notwithstanding the simple aim of nitrogen balance studies (that is, definition of the relationship between intake and all losses—urinary, fecal, and surface), the methodology has sustained considerable criticism. For example, Young and colleagues have argued that because of "conceptual limitations" and poor reproducibility, N balance studies in adults in relation to assessment of amino acid requirements are of questionable value (Marchini et al., 1993; Young, 1986). Some of the difficulties reflect the practical problems that arise in performing N balance studies.
Measurements of N balance (i.e., N intake minus all N losses) are relatively imprecise, with balance being a small value compared with the much larger values of N intake and N excretion. This results in considerable error in the prediction of balance (see Forbes, 1973; Wallace, 1959). Hegsted (1976) commented that intake tends to be overestimated because of spillage and incomplete recovery of uneaten food, while excreta are often less than completely recovered. This has variously been attributed to denitrification by bacteria in the colon with loss of N gas (Costa et al., 1968); to an underestimate of urea losses from skin and expired ammonia (Calloway et al., 1971) with losses of nitrogen as gaseous ammonia rarely measured; to nitrate in food and urine, which is not measured by the Kjeldahl technique (Kurzer and Calloway, 1981); and possibly to endogenous NO production via NO synthase (Anggard, 1994). NO is oxidized to nitrate, which is excreted in urine, or diffuses into the colon, where it is reduced to ammonia and nitrite (MacFarlane and Cummings,
1991). Nitrate excretion and fecal N-nitroso compound levels increase in response to increased protein, nitrate, and meat intakes (Bingham et al., 1996; Rowland et al., 1991). Furthermore, although fecai losses might be assumed to be predictable according to knowledge of dietary protein digestibility, the major determinant of fecal N output is biomass arising from bacterial fermentation in the colon (Stephen and Cummings, 1979). It is dependent on dietary nonstarch polysaccharide (NSP) and other fermentable carbohydrate intake (Cummings et al., 1992). These factors result in typical errors estimated in the range of -11 to +11 mg/kg/d (see Millward and Roberts, 1996), more than the entire miscellaneous losses (8 mg N/kg).
A problem in very short-term studies is how to account for the changing size of the body urea pool. In subjects fed a high-protein diet, losses from the body urea N pool in a 12-hour postabsorptive period accounted for on average 34 percent of the observed N balance with individual values as high as 66 percent, with gains of similar magnitude in the postprandial state (Price et al., 1994). Without correcting for these changes the distribution of N losses between the postabsorptive and postprandial phase of the diurnal cycle would be markedly in error, with an underestimate of postprandial and an overestimate of postabsorbtive losses.
Another difficulty stems from the analysis of the balance data and estimation of a requirement value, given the nonlinearity of the balance curve. In multi-level feeding trials in adults which start at low intakes, losses rise to match intakes when body protein reaches its maximum regulated level. Thus, the intake balance curve is, in theory, curvilinear. Balance increases with intake asymptotically toward zero or (as is often observed) some positive value, as losses increase to eventually match intake. This means that there is no simple term to define the overall shape of the balance curve and allow prediction of the requirement (as the zero balance intake). In fact, as is well documented in protein requirement balance studies, prediction of a zero balance intake intercept from a few balance points by linear regression will result in requirement values that vary according to where the intake values lie on the balance curve (see Millward and Roberts, 1996). Thus, studies conducted at low intakes will underestimate requirements, while studies conducted with supramaintenance intakes will overestimate requirements. The logic of this is that (a) reliable balance studies are those conducted with intakes very close to the actual requirement and (b) studies with intakes based on preconceived requirement values will tend to confirm such preconceptions.
Finally, the conceptual limitations raised by Young relate to the problem of evaluating whether zero balance, when obtained, signifies an adequate metabolic state (i.e., appropriate rates of protein turnover and lack of any cellular pathology). Young discussed attainment of balance in terms of adaptation (metabolic changes with no adverse consequences) and accommodation, metabolic changes which include some adverse response such as a loss of body tissue or reduction of protein turnover rates. Clearly, such a response would indicate a
lack of adequacy. In this author's view, minimum requirements need to be defined both in terms of balance and body nitrogen, the latter being a level of body protein judged to be appropriate for the subject's height and frame size. This raises a major design difficulty in balance studies. Losses of body N and achievement of balance at a lower body N level must be differentiated from any transient losses during an adaptation period that are gradually replaced, thus restoring appropriate body protein stores. Many of the balance studies reported have involved this initial low-protein period and the restoration stage has generally been ignored, with the test diets fed to subjects who would be depleted. This will result in very high efficiency of utilization and a probable overestimation of the requirement. Experimentally, however, the problem becomes more difficult since studies need to be long enough not only for balance to be achieved at the lowered level, but also for repletion of losses induced during the adaptation to occur. This will be most difficult to measure.
A final issue is raised by Fuller and Garlick (1994), who pointed to animal data and indicated that nitrogen equilibrium may not necessarily mean amino acid equilibrium. They cited the depletion of the tissue peptides, camosine (β-alanyl histidine) and anserine (β-alanyl-l-methyl histidine), and the depletion of hemoglobin, a protein very rich in histidine, when histidine-free diets are fed as was shown in the original balance studies of Rose and others. They also pointed to animal experiments that showed changes in the amino acid composition of the body induced by feeding amino acid-deficient diets; for example, pigs fed low-lysine diets have less lysine, phenylalanine, and tyrosine, and more glycine and arginine in whole body protein than those fed an adequate diet.
Are There Robust N Balance Data that Support the FAO Requirement Values?
Whether the FAO requirement levels maintain N balance in normal adults has been examined in N balance studies. Weller et al. (1971) tested in one group the pattern that Rose fed or in another group the pattern with a 30 percent addition of all essential amino acids together with nonessential N to give a total level of 7 g N. These diets did not maintain N balance, but an egg pattern with at least 2 times the Rose values did maintain balance. Note that in these studies, no histidine was fed, and the overall level of N at 7 g (0.6 g protein/kg) is low compared with usual intakes.
In contrast, Young and colleagues (Marchini et al., 1993) tested the 1985 FAO pattern (with histidine), fed for 3 weeks at a higher total N level (9.8 g N, 0.9 g protein/kg), against the MIT and an egg pattern and showed that all three diets maintained N balance; as shown in Figure 9-6, there were no significant differences among FAO, MIT, or egg diets. Young rejected these data as evidence for the validity of the FAO requirement values. Since the balance studies were carefully done, with due consideration given to miscellaneous losses, re-
jection implies nonacceptance of N balance in adults under any circumstances, which in this author's view is an extreme and unjustifiable position.
Biological Value of Plant Proteins in Human N Balance Trials
The extent to which a very low IAA requirement for maintenance is valid can be examined in terms of whether differences in the apparent quality of plant proteins are observed in N balance trials. Such trials have been reported in children and in adults. In children, responses need to be considered in relation to the expected rates of growth and the relative magnitude of growth and maintenance demands. In fact, tissue growth represents the major part of human requirements only in the first few months of life, 60 percent of metabolic demand at I month, 20 percent at 12 months, 10 percent at 2 years, and lower subsequently (Dewey et al., 1996). When rapid growth is occurring, differences in dietary protein quality are observed. Graham and colleagues studied the nutritive value of various unsupplemented plant protein sources (wheat, maize, potato, rice, beans, and sorghum) in terms of both digestibility and biological value relative to casein in young children recovering from malnutrition (Graham et al.; 1979; Maclean et al., 1981). They observed that sorghum and wheat were poorly utilized, which is consistent with a lysine limitation. However, the relatively small increases in lysine and other IAAs provided by rice, maize, and improved maize strains increased the biological value (BV) to between 80 and
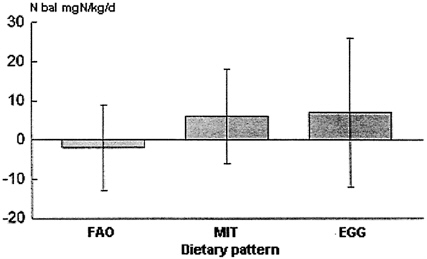
FIGURE 9-6 Nitrogen balances observed in young adults fed diets containing the FAO pattern, the MIT pattern, and an egg pattern. Diets contained 9.8 g/N, 0.9 g protein/kg, and subjects were fed for 3 weeks. Note: FAO, Food and Agricultural Organization; MIT, Massachusetts Institute of Technology. Source: Adapted from Marchini et al. (1993).
95 percent that of casein, with potato protein actually higher than casein. Furthermore, in a more recent study (Graham et al., 1990), they showed that in children growing at 2 to 3 g/kg/d after the most rapid phase of catch-up growth has passed, the genetically improved maize was as effective as casein in supporting both weight and height growth. This suggests that in these relatively rapidly growing children, their growth needs do influence their amino acid requirement, and the maintenance component of their needs, which is still substantial, includes a lower level of IAAs compared with their growth needs. This is supported by the observation that lysine supplementation of Indian children fed a wheat- based diet had no effect on weight gain or N balance (Reddy, 1971). In other preschool children fed either wheat-or rice-based diets, there was lower height growth with wheat compared with rice, which improved with lysine supplementation. However, the rice-based diet, which would have had a low BV in rat growth trials, allowed both weight and height growth at the fiftieth centile of the National Center for Health Statistics (NCHS) standards (Begum et al., 1970).
A large number of adult trials of protein quality have been conducted with a variable range of findings. Although differences among protein sources have been reported in N balance studies in young adults (e.g., relative biological values of 0.66 for wheat compared with beef [Young et al., 1975]), when the calculated BVs of a range of plant and animal proteins measured in separate studies are examined together (see Millward et al., 1989), the differences between wheat and other proteins and mixed diets are much less apparent. This is because of the lack of reproducibility between studies with the same protein. Within individual studies, interindividual variability is very marked, with biological values often associated with coefficients of variation (CVs) of 15 to 20 percent (e.g., Young et al., 1973), and even 50 percent (Young et al., 1984). In a study of the effect of lysine supplementation of a wheat gluten-based diet, (Scrimshaw et al., 1973), which is widely quoted as indicating protein quality effects in adults, the magnitude of the response (a 2.9-7.7 percent fall in urea excretion) was such that with the CV of mean urea excretions ranging from 11 to 36 percent, the response was not statistically significant. The combination of within-study variability and poor reproducibility between trials means that statistical analysis of the data is nearly impossible.
However, there have been several long-term trials examining the effects of a wheat-based diet on both weight and maintenance of N balance. In the Minnesota bread study (Bolourchi et al., 1968), young adults were fed a ''bread" diet for 50 days. The bread contained 90 to 95 percent protein from wheat flour, which supplied 67 g protein/d, 0.95 g/kg, and 18 mg/kg lysine. The result was a negative balance during the first 10 days, followed by a positive balance of 1 g N/d, sufficient to cover miscellaneous losses. Body weight was stable, and fitness, measured in terms of pulse rate with exercise, improved.
In the North Carolina wheat study (Edwards et al., 1971), responses were measured over 60 days in young men who were fed bread-based diets supplying
46 g protein [35 g from wheat (76%) and 11 g from other plant sources]. Body weights were maintained (± 4 pounds) as was N balance (at +1 g N/d), and this was not influenced by replacing 20 percent wheat by beans, rice, or peanuts.
Clark et al. (1962) fed 56 g protein (9 g N), of which 45 percent was from wheat flour and corn, together with amino acids, with lysine supplying 600, 700 or 950 mg/d (14 mg/kg). There was a progressive increase in N retention for 30 days, and at 700 and 950 mg/d balance of lysine, was maintained when 0.5 g N is included for miscellaneous losses.
Taken together, these N balance data do point to a relatively low MD for IAAs in the human maintenance requirement. Regardless of one's view about the validity of N balance, weight and fitness maintenance data from the Minnesota bread study strongly support the possibility that the MD for IAAs is very low.
Adjustment of the Food and Agriculture Organization Requirement Values
Although many of these potential sources of error in the N balance methodology preclude correction of published data, the error induced by not including surface losses, as in the N balance experiments of Rose, Leverton, and others, can be corrected. Hegsted (1963) performed a meta-analysis on published data, excluding the Rose data, which he considered poor (see Hegsted, 1973), and performing a regression analysis to calculate the requirement for zero balance and suggested that an adjustment could be made for unmeasured losses. Clearly, the magnitude of the additional losses requires careful consideration. Calloway et al. (1971) made comprehensive measurements of dermal and other N losses in adult subjects, reporting greatly increased dermal losses with exercise-induced sweating and with protein intake. They estimated a potential total loss of about 0.5 g/d; a value essentially adopted by FAO/UNU/WHO (1985) (8 mg/kg/d). Fuller and Garlick argued that since the loss is variable with protein intake and exercise, such an adjustment may be excessive in the balance experiments reviewed by Hegsted. They proposed a lower value estimated by Calloway et al. (1971) for sedentary subjects on moderate nitrogen intakes: 0.30 g N/d, 4 to 5 mg N/kg. The impact of this adjustment is shown in Table 9-5; the total requirement values were doubled for most amino acids. Nevertheless, when these values are compared with the pattern obtained using OOL, the former (values recalculated for sedentary individuals on moderate N intakes) are lower than the latter, with the exception of the TSAs, which are similar to the OOL as would be expected from the calculations. Lysine in particular is half the concentration in the OOL pattern. Given the care with which Hegsted selected and analyzed the published data to arrive at his values in Table 9-5, and given that none of the studies involved excessive energy intakes, since the Rose values were excluded, the recalculated
values in Table 9-5 are probably the best estimates of the minimum requirement for indispensable amino acids available.
Stable Isotope Estimates of Requirement Values
Isotopic studies of amino acid oxidation particularly based on 13C were proposed as an alternative to N balance by Young (1986). Initially, such isotopic studies involved an examination of the relationship between intake and some response, such as protein synthesis or amino acid oxidation in order to define an endpoint (e.g., Meredith et al., 1982). Subsequently, daily balances were calculated from measured rates of amino acid oxidation over a range of intakes. Balances were reported for leucine (Meguid et al., 1986a), lysine (Meredith et al., 1986), threonine (Zhao et al., 1986), and valine (Meguid et al., 1986b). Subsequently, a large number of 13C leucine and some valine balances have
TABLE 9-5
FAO Requirement Values Recalculated to Allow for Miscellaneous Losses
|
|
FAO* mg/d |
mg/kg (70)§ |
Recalculated† mg/d |
mg/kg(70) |
OOL‡ |
|
Histidine |
— |
— |
— |
— |
11.5 |
|
Ileucine |
550 |
7.9 |
1,083 |
15.5 |
16.2 |
|
Leucine |
730 |
10.4 |
1,580 |
22.6 |
27.4 |
|
Lysine |
545 |
7.8 |
1,118 |
16.0 |
30.1 |
|
TSA |
350 |
5.0 |
960 |
13.7 |
13.4 |
|
TAA |
350 |
5.0 |
1,184 |
16.9 |
27.0 |
|
Threonine |
375 |
5.4 |
942 |
13.5 |
15.5 |
|
Tryptophan |
168 |
2.4 |
222 |
3.2 |
4.0 |
|
Valine |
622 |
8.9 |
811 |
11.6 |
16.9 |
|
Total |
3,690 |
|
7,898 |
|
162 |
|
% protein |
10.5 |
|
22 |
|
48 |
|
NOTE: FAO, Food and Agricultural Organization; OOL, Obligatory oxidative loss; TAA, total aromatic amino acid; TSA, total sulfur amino acid. * Values from Hegsted (1963) calculated for zero balance. † With the exception of lysine, TSA and TAA, all values computed from regression equations reported by Hegsted (1963) with addition of 0.3gN losses. Lysine is recalculated from Jones et al. (1958) with original balance data expressed per kg bodyweight, adjusted for 5 mgN/kg per day additional losses and with zero balance calculated from regression of balance (mgN/kg per d) on logo intake (mgN/kg per d) with all reported balance periods included in the regression. ‡ Pattern in an amount of tissue protein equal to the obligatory nitrogen loss (54 rng N/kg). § Based on n 70 kg man. TSA is the OOL adjusted for utilization as described by Young et al. (1989). TAA is from Tolbert and Watts (1963). |
|||||
been reported, culminating with the most recent reports in which leucine oxidation and balance have been monitored for entire 24-hour periods (El-Khoury et al., 1994a,b, 1995). Taken together, this represents a large and complex body of experimental work. Reviewing the work is difficult since the experimental design and calculation methods have changed over time as potential problems have emerged. For example, the way in which leucine oxidation has been calculated has changed from an approach that by attempting to compensate for the tracer would have underestimated the true oxidation by 5 to 15 percent (see Millward, 1993) to calculation of actual rate in current reports (El-Khoury et al., 1994b). The current 24-hour studies are state of the art although, as discussed below, some problems persist. However, as discussed elsewhere (Millward and Rivers, 1988), the initial leucine, lysine, valine, and threonine studies, which have been used as experimental support for the MIT scoring pattern, are deeply flawed. Oxidation rates are underestimated due to use of the plasma amine acid labeling as opposed to intracellular amine acid labeling as a precursor, with no account taken for losses of 13C from threonine by routes other than CO2, and with balance calculated from a single fed value and an assumed (nonmeasured) postabsorptive value. In essence, 13C tracer balances can be applied in one of two ways: as measures of the intake-balance relationship for the particular tracer under examination (as with the leucine, lysine, valine, and threonine studies) or, alternatively, the balance of one amine acid such as leucine can be used as an alternative to N balance. Thus, for an indispensable amine acid like leucine, which has a small, highly regulated free pool, if the intake and oxidative loss are measured accurately, then the net accretion or loss of that amine acid can be assumed to be protein. This was the approach used by Marchini et al. (1993) to compare the FAO, MIT, and egg IAA patterns.
Many of the kinetic assumptions and isotope-related problems associated with this approach are well known (Bier et al., 1985) and were reviewed by Millward and Rivers (1988) and by Millward et al. (1991). The main problems are listed in Table 9-6. This list of problems is by no means exhaustive; it has been separated into problems that are ultimately soluble and those that are currently less tractable. Problem 1 is unavoidable with current methodology. Although the 24-hour studies (El-Khoury et al., 1994a, b) represent heroic experiments, they represent a short period in the context of balance studies, in general, with subjects necessarily restricted resulting in unknown consequences for protein balance. Day-to-day variation is large for N balance studies, some of which may reflect the operation of the regulator of body composition. The extent of day-to-day variation in leucine balance is currently unknown. El-Khoury et al. (1994a, b) reported daily balances in subjects on a high leucine intake (89 mg/d) ranging from -3.58 to +5.83 mg leucine/kg (0.76 ± 2.99 mg/kg/d) and falling to -13.4 ± 5.2 mg/kg/d on an intake of 14 mg/kg/d. This would indicate less inherent variability than N balance.
TABLE 9-6
Potential Problems Relating to 13C Isotopic Measurements of Amino Acid Oxidation Rates and Amino Acid Balance
|
A. Practical problems with potential solutions? |
|
1. Need for subject restriction and limitation to short periods of study. |
|
2. The extent of isotope retention as bicarbonate. |
|
3. Measurement of CO2 production rates. |
|
4. Variation in background tracer enrichment. |
|
5. Quantitative excretion of labeled carbon as CO2. |
|
6. Oxidation rates as a tracer for overall N excretion influenced by mismatch of food and body tissue amino acid composition. |
|
7. Underestimation of losses due to non-CO2 routes. |
|
8. Excessive positive balances in some studies. |
|
B. Model problems: Less tractable and possibly insoluble. |
|
9. Amount of tracer excessive and may influence balance. |
|
10. True precursor amino acid enrichment of measured value. |
Problems 2 to 4 are solvable by appropriate methodologies. Problem 5 can probably be ignored for leucine and most carboxyl-labeled amino acids, but possibly not for threonine, which exhibits unrealistic positive balances (Zhao et al., 1986), most likely because of substantial fixation as glycine (Ballevre et al., 1990). Problem 6 is important when oxidation rates are measured to calculate overall amino acid oxidation and N excretion. Thus, during the fed state with milk-based food, for example, the higher leucine content in milk (10%) compared with tissue protein (8%) means that excess leucine oxidation during feeding will occur, and unless corrected for, will result in an underestimate of overall amino acid balance during feeding (see Price et al., 1994). Problem 7 is important but usually ignored. Any route of leucine loss as the entire amino acid (free or protein bound in skin, hair, urinary and fecal peptides, or fecal bacterial protein) will not be measured. Although the magnitude of this loss is unknown, it could represent the equivalent of several mgs of N/kg. Problem 8 is observed in several reported studies. Indeed, in the studies of Marchini et al. (1993), the leucine balances ranged from -100 μmol/kg/d on the FAO diet to +180 μmol/kg/d on the egg diet. These values were equivalent to changes in body weight over the 3 weeks of the study of-1.5 kg to +2.5 kg. Whether such changes in body weight occurred is not mentioned by the authors.
As for problem 9, the fact that in stable isotope studies, the tracer is not massless but may be infused at significant rates compared with either dietary intakes or measured oxidation rates is a serious problem. In some circumstances, the tracer infusion rate can be of the same magnitude as the oxidation rate that it is purporting to trace. For example in a valine infusion (Pelletier et al., 1991a), the tracer infusion rate was 80 percent of the oxidation rate being studied. In the lysine studies reported by Meredith et al. (1986) on the lowest intakes of lysine, the infusion rate was 72 percent of the reported oxidation rate, equal to 3 times the rate of lysine intake from the diet at the lowest dietary intake studied. In other leucine infusion studies (Pelletier et al., 1991b), 8.2 μmol/kg/h of leucine was infused, equivalent over 12 hours to an intake of 13 mg/kg leucine in individuals fed 13 mg/d of leucine. Clearly, the way in which the infusion intake is assumed to influence the system under study and the consequent way it is treated in calculating oxidation rates and balance can markedly influence the results.
This problem was discussed by Fuller and Garlick (1994), who considered several issues. One was treatment of the tracer in 24-hour balances extrapolated from measurements made during a few hours in subjects in the fed and fasted states. In the initial studies, only the labeled amine acid intake given during the actual period of infusion was included (e.g., 6, 42-44), which would have underestimated balance by 9 to 13 mg. A second issue was that the tracer represents an additional level of intake above the dietary level to which the subjects have adapted, so that what is studied is not what has been previously fed. In the 24-hour studies (El-Khoury et al., 1994a,b), the diet was adjusted on the infusion day by removing an amount of leucine equal to that infused (about 10 mg). Although this means an overall (24-hour) leucine intake that is the same as during the adaptation period, the diet supplied less leucine than the nominal intake during the feeding period because of the tracer infused in the postabsorptive period. In fact at the lowest dietary level of 14 mg/kg/d, this reduced the actual intake during the feeding period (10 hours) to about 8 mg (food + tracer), the other 6 mg (tracer) being supplied during the postabsorptive 14-hour period. The likely effect of this would be to reduce the capacity for net postprandial protein synthesis through leucine limitation and to increase oxidation and negative balance in the postabsorptive state, with an overall leucine balance that is more negative than otherwise. El-Khoury et al. (1994) did show a gradual increase in leucine oxidation during the fasting period, as the excess was oxidized, and a fall during feeding, because of the high efficiency of utilization on the very low-leucine diet. Attempting to study the limits of adaptation of leucine oxidation and balance with a nutritionally significant amount of the tracer amine acid is highly unsatisfactory. Fuller and Garlick (1994) concluded that this problem is likely to give rise to an appreciable overestimate of the amine acid requirement.
Finally, Problem 10 represents a "model" problem resulting from an inadequate understanding of the kinetic model under analysis and especially the
uncertainty about the true precursor isotopic abundance compared with what is measured. Currently, only leucine and the other branched-chain amino acids allow any solution in terms of their keto acids in plasma, which derive from actual intracellular precursor pools. Even in these cases, the extent of any error remains unknown.
Price et al. (1994) attempted to test the assumptions empirically by comparing rates of leucine oxidation with measured N excretion rates in subjects in the fed and fasted states who were fed protein in amounts ranging from 0.33 to 2.2 g/kg/day. Leucine oxidation rates varied in proportion to N excretion over the entire range of N excretion rates, although the amounts of leucine oxidation were lower than expected. The ratio of predicted versus measured N excretion for the entire group was 0.79 (standard deviation 0.23, n = 38). In other words, leucine oxidation resulted in a 27 percent overestimate of balance compared with N.
In summary, 13C leucine oxidation studies, while not entirely free from potential problems, do appear to allow reasonable estimates of whole body protein balance when applied with care, particularly when intakes of leucine are not especially low. The 24-hour 13C leucine balances currently employed by Young and colleagues involve far fewer assumptions and uncertainties compared with the original tracer studies used to validate the MIT scoring pattern, which can, in this author's view, be generally discounted. Nevertheless at low leucine intakes, the tracer problem remains a serious one. Uncertainty still remains as to whether the apparent inability of 14 mg/kg/d to maintain balance is due to excessive postabsorptive oxidation of the tracer.
Toronto "Break Point" Studies
The Toronto Break Point studies are important because they represent the only stable isotope data on phenylalanine (Zello et al., 1990) and the only lysine study (Zello et al., 1992) apart from the MIT data. Furthermore, the lysine study involved a method that does not depend on balance, that is, the indicator amino acid oxidation method.
As shown in studies of animals fed amino acid mixtures, when the test amino acid intake falls below the requirement for adequate postprandial net protein synthesis, oxidation of a labeled indicator amino acid increases. The lysine study (Zello et al., 1992), measured 13C phenylalanine oxidation in subjects fed diets with fixed phenylalanine intakes but varying lysine intakes. The break point or increase in phenylalanine oxidation indicated the lysine intake that is inadequate. The phenylalanine study (Zello et al., 1990) involved measurement of postprandial phenylalanine oxidation on diets with decreasing amounts of phenylalanine.
The important problem with these studies is their design and interpretation. Phenylalanine oxidation is calculated from the isotopic enrichment of plasma phenylalanine, not tyrosine, and in fact, the investigators give excess tyrosine in
the diets. The authors argued that phenylalanine is oxidized from a pool that does not equilibrate with tyrosine so that plasma tyrosine enrichment, which was very low compared with that of phenylalanine, was not the relevant precursor enrichment. If their assumption is not correct, then their data are uninterpretable because (a) the 13CO2 will reflect the oxidation rates of both phenylalanine and tyrosine and (b) the excess tyrosine will dominate the oxidation of phenylalanine and blunt any usefulness the protocol might have in assessing phenylalanine oxidation.
It is difficult to accept the assumptions of Zello's studies in light of previous studies by Thompson et al. (1989), who developed Clark and Bier's original study (1982). Tyrosine labeling would be expected to be low since the labeled tyrosine formed by phenylalanine hydroxylation would be diluted by the tyrosine flux and would not be expected to achieve plateau enrichment within the 4 hour infusion without priming. Since these studies were measuring phenylalanine oxidation, the true shape of the oxidation curve in relation to the lysine intake is not known. In this author's view, neither study can be interpreted in a meaningful way in relation to either lysine or phenylalanine requirements. Therefore, there are no reliable kinetic data for a minimum lysine requirement.
Functional Indicators of Adequacy of Intakes Allowing Zero Balance
Protein Turnover
One proposed advantage of tracer studies is that they allow validation of balance in terms of measurement of protein turnover. In this context, Young and Marchini (1990) reported marked reductions in whole body protein synthesis in subjects fed low levels of specific IAAs. In fact, it is likely that these data are artifactual.
Most would agree that if reductions in protein turnover occurred in response to reduced protein or amine acid intakes, it would be a cause for concern. In fact, although the animal data for reductions in protein turnover with low-protein diets are quite clear (e.g., Jepson et al., 1988; Waterlow et al., 1978), evidence for this in humans is generally scant even in chronically undernourished Indian laborers (Soares et al., 1991). Considerations of the significance of changes in protein synthesis need to be made in an appropriate framework. In the context of the diurnal cycling model of balance, the increasing amplitude of gains and losses with increasing protein intake, does involve changes in protein synthesis (although changes in proteolysis are the main mediator of cycling). As a result, postprandial synthesis is higher for someone on a high- rather than low-protein intake, although postabsorptive synthesis rates are not changed (Pacy et al., 1994). Thus the changes in protein synthesis are only related to the increasing amplitude of diurnal cycling. Since
there is no obvious physiological benefit from any particular amplitude of diurnal cycling as long as overall balance is achieved, there is no obvious reason to be concerned if dietary protein or indispensable amino acid levels result in lower amplitudes of diurnal cycling and lower rates of postprandial protein synthesis. In fact, Pacy et al. (1994) reported that the mean daily rate of protein synthesis, S, was higher on very high protein intakes, (S=4.55 ± 0.5 g/kg/d), compared with 2 weeks on a very low protein diet of 0.36 g/kg/d (S=4.09 ± 0.63 g/kg/d), although P > 0.05, but even this was explainable by the fact that overall daily leucine balances became positive at 0.82 g protein/kg/d. Thus turnover per se (i.e., replacement) was not increased, and it was concluded that any influence of protein intake on protein turnover was below the detection limits of current methods. In general, previous results provided little support for a nutritional sensitivity of whole body protein turnover within the range of intakes likely to be consumed.
Marchini et al. (1993) reported no significant changes in protein synthesis in the postabsorptive or postprandial state on diets with the FAO, MIT, or egg amino acid patterns. The only kinetic responses commented upon were a reduced rate of postabsorptive proteolysis after 3 weeks on the FAO diet and a lack of a feeding-induced inhibition. Although this explains the lack of postprandial gain and overall negative balance, it appears that no changes in turnover occurred in these studies.
The one exceptional body of data is the large reduction in protein synthesis observed in subjects fed very low levels of individual amino acids in the MIT studies (Young and Marchini, 1990). However, Pacy and coworkers (1994) have suggested that since such responses are only observed when the tracer is the same as the limiting amino acid, and not observed in response to an identical dietary protocol when turnover is measured by a nonlimiting labeled amino acid (e.g., Zello et al., 1992), a methodological problem associated with either compartmentation or the nonsteady state may exist.
On this basis, it appears that the data suggesting that protein synthesis falls at inadequate intakes of individual amino acids are artifactual. Thus, with balanced amino acid mixtures and whole proteins, whole body protein synthesis changes only minimally at intakes below those capable of supporting overall balance. Thus, protein turnover is not a useful indicator of nutritional adequacy that can be used in the debate.
Other Metabolic Influences of Amino Acids
A minimal requirement for protein and amino acids may be identifiable with sufficient time allowed for adaptation. This value is likely to be much lower than the amounts provided by natural diets that also provide sufficient energy and other nutrients. Thus, the magnitude of this minimal requirement becomes to some extent only an issue of scientific curiosity. It might be considered that the requirement for balance is less important than the functional
consequences of particular levels of protein in the diet. In other words, the key issue is that of an optimal requirement, which not only allows balance but also supports optimal body function such as optimal immune function, minimal long-term deterioration of renal function, and in the present context, physical performance, especially under stress.
Although the issue of an optimal protein requirement is generally recognized as important, there is little qualitative or quantitative information. In growing animals, dietary protein intakes in excess of those associated with maximum efficiency of protein utilization have been shown to increase rates of bone growth through hormonal mechanisms involving insulin, thyroid hormones, and insulin-like growth factor-I (IGF-I). This has led to the concept of an "anabolic drive" of dietary protein exerted by protein intakes in excess of minimal needs (see Millward and Rivers, 1989). However, the extent to which this is important in human growth has yet to be explored.
The importance of dietary protein for the maintenance of the immune system has also been suggested, with dietary protein influencing the human response to infection through several mechanisms (see Reeds and Becket, 1996). One mechanism is the maintenance of gut barrier function, through provision of threonine, cysteine, and other amine acids involved in synthesis of mucus glycoproteins. Another mechanism is maintenance of general immune-competence through provision of specific amine acids for synthesis of cellular proteins of the immune system and support of the hepatic acute-phase protein response. In particular, glutathione, a key free radical scavenger synthesized from glutamate (glutamine), glycine, and cysteine is depleted in hepatic and intestinal mucosa in protein-restricted animals, in the erythrocytes of infants suffering from kwashiorkor. Also infants with low birth weight have low circulating levels of glutathione. The reduced levels of glutathione in rats are restored by providing cysteine in the diet. The influence of protein intake on glutamine levels is also a possible factor influencing immune function.
Glutamine is strongly concentrated in skeletal muscle, appears to play a specific role in maintaining function of rapidly proliferating cells such as lymphocytes and mucosal enterocytes, and regulates muscle protein turnover. Under conditions of infection and trauma, muscle concentrations of glutamine fall, and this may limit the provision of glutamine for the immune system or to the splanchnic bed, where it appears to play a specific role in maintaining glutathione synthesis during trauma. Taurine, a beta-amine sulfonic acid derived from cysteine, appears to be an effective scavenger of peroxidation products (particularly those containing the oxychloride groups) and acts as a neuromodulatory agent. Taurine is specifically concentrated in skeletal musculature and in the central nervous system. Creatine is a compound that is crucial for energy flow within skeletal muscle and is synthesized from glycine, arginine, and a suitable source of methyl groups. Creatine is concentrated in skeletal muscle and brain and is lost from muscle during infection and trauma, with a deterioration of muscle contractile function. Because glutamine, creatine,
and taurine are maintained at substantial concentrations in the free amino acid fraction of milk, provision of all three compounds is believed to be important in supporting postnatal development. Finally, provision of arginine is important in relation to the immune system. This may relate to its role as precursor for synthesis of nitric oxide, a key regulator of a variety of physiological processes that include the regulation of hepatic protein metabolism, macrophages' killer function, and interactions between macrophages and lymphocyte adhesion and activation. However, as yet the quantitative relationship between intakes of any of these individual amino acids and function has not been quantified.
As for physical activity, it is often assumed that increasing protein intakes is of benefit (e.g., Lemon, 1996), but much of the data indicating apparent increased protein requirements of athletes are based on arguably misleading N balance studies. In this author's view, there is no unequivocal evidence and some data suggesting potential disadvantages (see Millward et al., 1994). However, as is often the case with the nutrition literature, the key experiments are described in the classic texts written decades ago. Chittenden took a group of soldiers and elite Yale University athletes and persuaded them to reduce their protein intake by 50 percent over 5 months, mainly by switching to a vegetarian diet of about 0.75 g protein/kg/d. Extensive measures of strength were made. Over the 5 months, soldiers' strength increased by > g5 percent (largely due to a training effect), but even the athletes' strength increased on average by 35 percent (see Table 9-7), coupled with a fall in perceived fatigue. Chittenden
TABLE 9-7 Strength of Male Athletes Before and After a S-Month Period of a Reduced-Protein Diet*
|
|
Total Strength† |
||
|
Athlete |
Initial |
Final |
Increase (%) |
|
1 |
4,913 |
5,722 |
16 |
|
2 |
6,016 |
9,472 |
57 |
|
3 |
5,993 |
8,165 |
36 |
|
4 |
2,154 |
3,993 |
85 |
|
5 |
4,554 |
5,917 |
29 |
|
6 |
4,548 |
5,917 |
30 |
|
7 |
5,728 |
7,135 |
23 |
|
8 |
5,351 |
6,833 |
28 |
|
|
|
Mean |
38 |
|
* Exclusion of most dietary meat and reduction of protein intake by ''more than 50%" to about 55 g protein/d over 5 months. † Sum of 15 distinct tests of measured strength and work performed in various exercises (Chittenden, 1907). |
|||
concluded that his experiments "afford reasonable proof of the beneficial effects of a lowered protein intake upon the muscular strength of man," and that "man can profitably maintain nitrogen equilibrium and body weight upon a much smaller amount of protein food than he is accustomed to consume (Chittenden, 1907, p. 207)." In the context of minimal and optimal protein requirements, Chittenden defined an optimal requirement of about 0.75 g/kg, the current safe allowance, with most of it coming from plant protein sources. This is significantly less than average intakes. Chittenden's views were controversial at the time and have been given less prominence than they deserve in the intervening years of continued controversy. However, the data speak for themselves.
AUTHOR'S CONCLUSIONS AND RECOMMENDATIONS
Defining amine acid requirements is inherently difficult even in the context of "normal" needs, let alone in the specific context of this workshop. Figure 9-7 summarizes what can and cannot be defined in relation to amine acid requirements and, especially, the problem of assessing nutritional adequacy of proteins by scoring. The scheme is drawn for lysine, but the principle applies generally. A requirement for rapid growth can be defined, and it may well be close to the FAO preschool values. A minimum obligatory metabolic demand
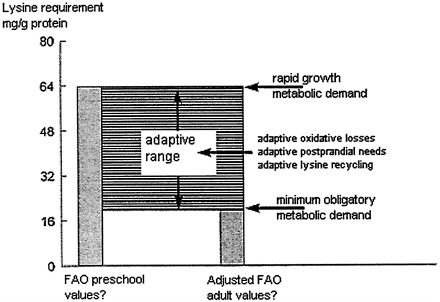
Figure 9-7 Amino acid requirements can be defined for rapid growth, which may be close to the FAO preschool values, and for a minimum obligatory metabolic demand, possibly represented by the adjusted values in Table 9-5. Assessment of the nutritional value of intakes in the adaptive range between these two extremes is problematic. Note: FAO, Food and Agricultural Organization.
can be deemed, and this is low, possibly represented by the adjusted values in Table 9-5. The problem arises in assessing the nutritional value of intakes in the adaptive range between these two extremes. There are mechanisms allowing considerable adaptation of MD to match intakes in this adaptive range, but because it is not easy to predict whether complete adaptation will occur; one cannot be confident of the nutritional value of the protein. This means that the protein cannot be scored. Protein scoring as a means of predicting net protein utilization (NPU) in animals was adopted after demonstrating that it worked (more or less). That is, the score was closely correlated with experimentally determined NPU within a relatively simple animal model in which tissue growth dominated needs. Humans have a much more complex MD that cannot be defined as a fixed quantity. In the absence of suitable indicators of adequacy that allow judgments to be made about optimal requirements, which are independent of balance, a scoring pattern for assessment of nutritional value in the adaptive range cannot be deemed and is unlikely to be useful. Although a scoring pattern based on the adjusted FAO values in Table 9-5 may represent an ultimate minimum nutritional value, actual adequacy could not be judged without long-term studies of the extent to which diets with intakes as low as this allowed balance, health, and fitness.
Thus, the key test of adequacy of either protein or amino acid intake must be the long-term response in terms of the specific function of interest. A specific question that this workshop addressed related to amino acid requirements is the following: Is there evidence that a different mix of amino acids would optimize military performance (cognitive function, long-term muscle mass preservation) during high workload, psychological stress, and/or energy deficit? Given the uncertainty about amino acid requirements for normal health, it is not surprising that this question cannot currently be answered. As far as gender issues are concerned, none of the investigations that provided the FAO values identified gender differences in normal requirements.
Regarding protein needs, what would be of interest is a controlled trial repeating Chittenden's measurements, with both protein level and quality as independent variables. Although N balance and stable isotope studies would be of great interest in such studies, the main outcome test of adequacy would be, simply, performance. There is no reason why such a trial should not be undertaken.
REFERENCES
Allison, J.B., J.A. Anderson, and R.D. Seeley. 1947. Some effects of methionine on the utilization of nitrogen in the adult dog J. Nutr. 33:361-370.
Anggard, E. 1994. Nitric oxide: mediator, murderer and medicine. Lancet 343:1199-1206.
Ballevre, O., A. Cadenhead, A.G. Calder, W.D. Rees, G.E. Lobley, M.F. Fuller, and P.J. Garlick. 1990. Quantitative partition of threonine oxidation in pigs: Effect of dietary threonine. Am. J. Physiol. 259:E483-E491.
Begum, A., A.N. Radhakrishnan, and S.M. Pereira. 1970. Effect of amine acid composition of cereal-based diets on growth of preschool children. Am. J. Clin. Nutr. 23:1175-1183.
Bender, A.E. 1961. Determination of the nutritive value of proteins by chemical analysis. Pp. 407-415 in Progress in Meeting Protein Needs of Infants and Preschool Children. Pub. 843. Washington, D.C.: National Academy of Sciences and National Research Council.
Benevenga, N.J., M.J. Gahl, T.D. Crenshaw, and M.D. Fink. 1994. Protein and amine acid requirements for maintenance and growth of laboratory rats. J. Nutr. 124:451-453.
Bergstrom, J, P. Furst, and E. Vinnars. 1990. Effect of a test meal with and without protein on muscle and plasma free amine acids. Clin. Sci. 79:331-337.
Bier, D.M., D.E. Matthews, and V.R. Young. 1995. Interpretation of amine acid kinetic studies in the context of whole body protein metabolism. Pp. 27-36 in Substrate and Energy Metabolism in Man, J.S. Garrow and D. Halliday, eds. London and Paris: John Libbey.
Bingham, S., B. Pignatelli, J. Pollock, A. Ellul, C. Mallaveille, G. Gross, S. Runswick, J.H. Cummings, and I.K. O'Neill. 1996. Does increased formation of endogenous N nitroso compounds in the human colon explain the association between red meat and colon cancer? Carcinogenesis 17:515-523.
Bolourchi, S., C.M. Friedeman, and O. Mickelson. 1968. Wheat Flour as a source of protein for adult human subjects. Am. J. Clin. Nutr 21:827-835.
Calloway, D.H., A.C.F. Odell, and S. Margen. 1971. Sweat and miscellaneous nitrogen losses in human balance studies. J. Nutr. 101:775-796.
Chittenden, R.H. 1907. The Nutrition of Man. London: Heinemann.
Clugston, G., K.G. Dewey, C. Field, D.J. Millward, P. Reeds, N.S. Scrimshaw, K. Tontisirin, J.C. Waterlow, and V.R. Young. 1996. Report of the working party on protein and amine acid requirements. Eur. J. Clin. Nutr. Suppl. 1:S193-S195.
Clark, H.E., L.L. Reitz, T.S. Vacharotayan, and E.T. Mertz. 1962. Effect of certain factors on nitrogen retention and lysine requirements of adult human subjects. J. Nutr. 78:173-178.
Clarke, J.T.R., and D.M. Bier. 1982. The conversion of phenylalanine to tyrosine in man. Direct measurement by continuous intravenous tracer infusions of L-[ring-2H5] phenylalanine and L-[l-13C]tyrosine in the postabsorptive state. Metabol. 31:999-1005.
Costa, G., L. Ullrich, F. Kantor, and J.F. Holland. 1968. Production of elemental nitrogen by certain mammals including man. Nature 281:546-551.
Cummings, J.H., S.A. Bingham, K.W. Heaton, and M.A. Eastwood. 1992. Quantitative estimates of bowel habit, bowel disease risk and the role of diet. Gastroenterol. 103:1783-1789.
Dewey, K.G., G. Benton, C. Fjeld, B. Lonnerdal, and P. Reeds. 1996. Protein requirements of infants and children. Eur. J. Clin. Nutr. 50:(suppl. I):S119-S147.
Edwards, C.H., L.K. Booker, C.H. Rumph, W.G. Wright, M.S. Ganapathy, and Ganapathy Seetha. 1971. Utilization of wheat by adult man: nitrogen metabolism, plasma amine acids and lipids. Am. J. Clin. Nutr. 24:181-193.
El-Khoury, A.E., N.K. Fukagawa, M. Sanchez, R.H. Tsay, R.E. Gleason, T.E. Chapman, and V.R. Young. 1994a. Validation of the tracer-balance concept with reference to leucine: 24-h intravenous tracer studies with L-[l-13C]leucine and [15N-15N]urea. Am. J. Clin. Nutr. 59(5):1000-1011.
El-Khoury, A.E., N.K. Fukagawa, M. Sanchez, R.H. Tsay, R.E. Gleason, T.E. Chapman, and V.R. Young. 1994b. The 24-h pattern and rate of leucine oxidation, with particular reference to tracer estimates of leucine requirements in healthy adults. Am. J. Clin. Nutr. 59(5):1012-1020.
El-Khoury, A.E., M. Sanchez, N.K. Fukagawa, R.E. Gleason, R.H. Tsay, and V.R. Young. 1995a. The 24-h kinetics of leucine oxidation in healthy adults receiving a generous leucine intake via three discrete meals. Am. J. Clin. Nutr. 62(3):579-590.
FAO/WHO. 1973. Energy and protein requirements. Report of a joint FAO/WHO Ad Hoc expert committee. WHO Technical Report Series No. 522. Geneva: World Health Organization.
FAG/WHO. 1991. Protein quality evaluation. Report of a joint FAG/WHO expert consultation . Food and Agricultural Organization Food and Nutrition Paper 51. Geneva: Food and Agricultural Organization.
FAO/WHO/UNU. 1985. Energy and protein requirements. Report of a joint expert consultation. World Health Organization Technical Report Series No. 724. Geneva: World Health Organization.
Feteday, A., N. Gibson, M. Cox, D. Halliday, P.J. Pacy, and D.J. Millward. 1994. Postprandial utilization of wheat protein in normal adults. Proc. Nutr. Soc. 53:201a.
Fereday, A., N. Gibson, M. Cox, D. Halliday, P.J. Pacy, and D.J. Millward. 1997a. Postprandial protein utilization of what protein from a single meal in normal adults. Proc. Nutr. Soc. 56:80a.
Forbes, G.B. 1973. Another source of error in the metabolic balance method. Nutr. Rev. 31:297-300.
Fuller, M.F., and P.J. Garlick. 1994. Human amino acid requirements: Can the controversy be resolved? Annu. Rev. Nutr. 14:217-241.
Fuller, M.F., R. McWilliam, T.C. Wang, and L.R. Giles. 1999. The optimum dietary amino acid pattern for growing pigs; requirements for maintenance and for tissue protein accretion. Brit. J. Nutr. 62:255-267.
Fuller, M.F., A. Milne, C.I. Harris, T.M. Reid, and R. Keenan. 1994. Amino acid losses in ileostomy fluid on a protein-free diet. Am. J. Clin. Nutr. 59(1):70-73.
Gahl, M.J., M.D. Finke, T.D. Crenshaw, and N.J. Benevenga. 1991. Use of a four-parameter logistic equation to evaluate the response of growing rats to ten levels of each indispensable amino acid. J. Nutr. 121(11):1720-1729
Graham, G.G., E. Morales R.P. Placko, and W.C. Maclean. 1979. Nutritive value of brown and black beans for infants and small children. Am. J. Clin. Nutr 32:2362-2366.
Graham, G.G., J. Lembcke, and E. Morales. 1991. Quality-protein maize as the sole source of dietary protein and fat for rapidly growing young children. Pediatrics 85(1):85-91.
Hegsted, D.M. 1973. The amino acid requirements of rats and human beings. Pp. 275-292 in Proteins in Human Nutrition, J.W.G. Porter and B.A. Rolls, eds. London: Academic Press.
Hegsted, D.M. 1963. Variation in requirements of nutrients: Amino acids. Fed. Proc. 22:1424-1430.
Hegsted, D.M. 1976. Balance studies. J. Nutr. 106:307-311.
Jepson, M.M., P.C. Bates, and D.J. Millward. 198g. The role of insulin and thyroid hormones in the regulation of muscle growth and protein turnover in response to dietary protein. Brit. J. Nutr. 59:397-415.
Jones E.M., C.A. Bauman and M.S. Reynolds. 1956. Nitrogen balances in women maintained on various levels of Lysine. J. Nut. 60:549-559.
Kurzer, M.S., and D.H. Calloway. 1981. Nitrate and nitrogen metabolism in men. Am. J. Clin. Nutr. 34:1305-1313.
Lemon, P.W. 1996. Is increased dietary protein necessary or beneficial for individuals with a physically active lifestyle? Nutr. Rev. 54:S169-S 175.
MacFarlane, G., and J.H. Cummings. 1991. The colonic flora, fermentation, and large bowel digestive function. Pp. 51-92 in The Large Intestine: Physiology, Pathophysiology and Disease, S. Phillips, J.H. Peniberton, and R.G. Shorter, eds. New York: Raven.
Maclean, W.C., G.L. De Romana, R.P., Placko, and G.G. Graham. 1981. Protein quality and digestibility of sorghum in pre school children: Balance studies and plasma free amine acids. J. Nutr. 111:1928-1936.
Marchini, J.S., J. Cortiella, T. Hiramatsu, T.E. Chapman, and V.R. Young. 1993. Requirements for indispensable amine acids in adult humans: Longer-term amine acid kinetic study with support for the adequacy of the Massachusetts Institute of Technology amine acid requirement pattern. Am. J. Clin. Nutr. 58:670-683.
Meguid, M.M., D.E. Matthews, D.M. Bier, C.N. Meredith, J.S. Soeldner, and V.R. Young 1986a. Leucine kinetics at graded leucine intakes in young men. Am. J. Clin. Nutr. 43:770-780.
Meguid, M.M., D.E. Matthews, D.M. Bier, C.N. Meredith, and V.R. Young. 1986b. Valine kinetics at graded valine intakes in young men. Am. J. Clin. Nutr. 43:781-786.
Meredith, C., D.M. Bier, M.M. Meguid, D.E. Matthews, Z. Wen, and V.R. Young. 1982. Whole body amine acid turnover with 13C tracers: A new approach for estimation of human amine acid requirements. Pp. 42-59 in Clinical Nutrition '81, R.I.C. Wesdorp and P.B. Soeters, eds. Edinburgh and London: Churchill Livingstone.
Meredith, C.N., Z-M. Wen, D.M. Bier, D.E. Matthews, and V.R. Young. 1986. Lysine kinetics at graded lysine levels in young men. Am. J. Clin. Nutr. 43:787-794.
Millward, D.J. 1990. Amine acid requirements in adult man. Am. J. Clin. Nutr. 51:492-493.
Millward, D.J. 1992. The metabolic basis of the amine acid requirement. Pp. 31-57 in Protein-Energy Interactions I/D/E/C/G, N.W. Scrimshaw and B. Schurch, eds. Lausanne, Switzerland: Nestle Foundation.
Millward, D.J. 1993. Stable-isotope-tracer studies of amine acid balance and human indispensable amine acid requirements Am. J. Clin. Nutr. 57(1):81-86.
Millward, D.J. 1994. Can we define indispensable amine acid requirements and assess protein quality in adults? J. Nutr. 124:1509-1516.
Millward, D.J., and J.P.W. Rivers. 1988. The nutritional role of indispensible amine acids and the metabolic basis for their requirements. Eur. J. Clin. Nutr. 42:367-393.
Millward, D.J., and J.P. Rivers. 1989. The need for indispensable amine acids: The concept of the anabolic drive. Diab. Metab. Rev. 5(2):191-211.
Millward, D.J., and P.J. Pacy. 1995. Postprandial protein utilization and protein quality assessment in man. Clin. Sci. 88:597-606.
Millward, D.J., and J.C. Waterlow. 1996. Letter to the editor. Eur. J. Clin. Nutr. 50:832-833.
Millward, D.J., and S.R. Roberts. 1996. Protein requirement of older individuals . Nutr. Res. Rev. 9:67-88.
Millward, D.J., A.A. Jackson, G. Price, and J.P.W. Rivers. 1989. Human amine acid and protein requirements: Current dilemmas and uncertainties Nutr. Res. Revs. 2:109-132.
Millward, D.J., G.M. Price, P.J.H. Pacy, and D. Halliday. 1990. Maintenance protein requirements: the need for conceptual revaluation. Proc. Nutr. Sec. 49:473-487.
Millward, D.J., G. Price, P.J.H. Pacy, and D. Halliday. 1991. Whole body protein and amino acid turnover in man: What can we measure with confidence? Proc. Nutr. See. 50:197-216.
Millward, D.J., J.L. Bowtell, P. Pacy, and M.J. Rennie. 1994. Physical activity, protein metabolism and protein requirements. Proc. Nutr. Sec. 53(1):223-240.
Millward, D.J., A. Fereday, N. Gibson, and P.J. Pacy. 1996. Postprandial protein metabolism. Ballieres Clin. Endocrinol. Metabol. 10(4):533-549.
Munro, H.N. 1964a. General aspects of the regulation of protein metabolism by diet and hormones. Pp. 381-481 in Mammalian Protein Metabolism, Vol. I., Munro H.N. and J.B. Allison, eds. New York: Academic Press.
Osborne, T.B., and L.B. Mendel. 1916. The amino-acid minimum for maintenance and growth, as exemplified by further experiments with lysine and tryptophan. J. Biol. Chem. 25:1-8.
Pacy, P.J., G.M. Price, D. Halliday, M.R. Quevedo, and D.J. Millward. 1994. Nitrogen homeostasis in man: The diurnal responses of protein synthesis and degradation and amino acid oxidation to diets with increasing protein intakes. Clin. Sci. 86(1):103-116.
Pelletier, V., L. Marks, D.A. Wagner, P.A. Hoerr, and V.R. Young. 1991a. Branched-chain amino acid interactions with reference to amino acid requirements in adult men: Valine metabolism at different leucine intakes. Am. J. Clin. Nutr. 54:395-401.
Pelletier, V., L. Marks, D.A. Wagner, P.A. Hoerr, and V.P. Young. 1991b. Branched-chain amino acid interactions with reference to amino acid requirements in adult men: Leucine metabolism at different valine and isoleucine intakes. Am. J. Clin. Nutr. 54:402-407.
Pineda, O., B. Torun, F.E. Viteri, and G. Arroyave. 1981. Protein quality in relation to estimates of essential amino acids requirements. Pp. 131-139 in Protein Quality in Humans: Assessment and In Vitro Estimation, C.E. Bodwell, ed. Westport, Conn.: AVI Publishing Company Inc.
Price, G.M., D. Halliday, P.J. Pacy, M.R. Quevedo, and D.J. Millward. 1994. Nitrogen homeostasis in man: 1. Influence of protein intake on the amplitude of diurnal cycling of body nitrogen. Clin. Sci. 86:91-102.
Quevedo, M.R., G.M. Price, D. Halliday, P.J. Pacy, and D.J. Millward. 1994. Nitrogen homeostasis in man: 3. Diurnal changes in nitrogen excretion, leucine oxidation and whole body leucine kinetics during a reduction from a high to a moderate protein intake. Clin. Sci. 86:155-193.
Reddy, V. 1971. Lysine supplementation of wheat and nitrogen retention in children. Am. J. Clin. Nutr. 24:1246-1249.
Reeds, P.J, and P.R. Becket. 1996. Protein and amino acids. Pp. 67-86 in Present Knowledge in Nutrition, 7th ed., E.E. Ziegler and L.J. Filer, eds. Washington, D.C.: ILSI Press.
Rose, W.C. 1957. The amino acid requirements of adult man. Nutr. Abstr. Rev. 27(3):631-647.
Rowland, I.R, T. Granli, O.C. Bockman, P.E. Key, and P.C. Massey. 1991. Endogenous N nitrosation in man assessed by measurement of apparent total N nitroso compounds in faeces. Carcinogenesis 12:1359-1401.
Said, A.K., and D.M. Hegsted. 1970. Response of adult rats to low dietary levels of essential amino acids. J. Nutr. 100:1363-1376.
Scrimshaw, N.S, Y. Taylor, and V.R. Young. 1973. Lysine supplementation of wheat gluten at adequate and restricted energy intakes in young men. Am. J. Clin. Nutr. 26:965-972.
Soares, M.J., L.S. Piers, P.S. Shetty, S. Robinson, A.A. Jackson, and J.C. Waterlow. 1991. Basal metabolic rate, body composition and whole body protein turnover in Indian men with differing nutritional status . Clin. Sci. Colch. 81(3):419-425.
Stephen, A.M., and J.H. Cummings. 1979. The influence of dietary fibre on fecal nitrogen excretion in man. Proc. Nutr. Soc. 38:141A.
Thompson, G.N., P.J.H. Pacy, H. Merritt, G.C. Ford, M.A. Read, K.N. Cheng, and D. Halliday. 1989. Rapid measurement of whole body and forearm protein turnover using a [2H5]phenylalanine model. Am. t. Physiol. 256:E631-E639.
Tolbert, B, and J.H. Watts. 1963. Phenylalanine requirements of women consuming a minimal tyrosine diet and the sparing effect of tyrosine on the phenylalanine requirement. J. Nut. 80, 111-117.
Wallace, W.M. 1959. Nitrogen content of the body and its relation to retention and loss of nitrogen. Fed. Proc. 18:1125-1130.
Wang, T.C, and M.F. Fuller. 1989. The optimum dietary amino acid pattern for growing pigs. 1. Experiments by amino acid deletion. Br. J. Nutr. 62(1):77-89.
Waterlow, J.C. 1996. The requirements of adult man for indispensable amino acids . Eur. J. Clin. Nutr. 50 (suppl. 1):S151-176.
Waterlow, J.C., P.J. Garlick, and D.J. Millward. 1978. Protein Turnover in Mammalian Tissues and the Whole Body. Amsterdam: North Holland Elsevier.
Weller, L.A., D.H. Calloway, S. Margen. 1971. Nitrogen balance of men fed amine acid mixtures based on Rose's requirements, egg white protein, and serum free amine acid patterns. J. Nutr. 101(11):1499-1507.
Yoshida, A. 1983. Specificity of amine acids for the nutritional evaluation of proteins. Pp. 163-182 in Proceedings of the International Association of Cereal Chemists Symposium on Amine Acid Composition and Biological Value of Cereal Proteins, R. Lasztity and M. Hidvegi, eds. Budapest: Akademiai Kiado.
Young, V.R. 1986. Nutritional balance studies: Indicators of human requirements or adaptive mechanisms. J. Nutr. 116:700-703.
Young, V.R., and A. E. El-Khoury. 1995a. Can amine acid requirements for nutritional maintenance in adult humans be approximated from the amine acid composition of body mixed proteins? Proc. Natl. Acad. Sci. USA 921:300-304.
Young, V.R., and J.S. Marchini. 1990. Mechanisms and nutritional significance of metabolic responses to altered intakes of protein and amine acids with reference to nutritional adaptation in humans Am. J. Clin. Nutr. 51:270-289.
Young, V.R., Y.S.M. Taylor, W.R. Rand, and N.S. Scrimshaw. 1973. Protein requirements of man: efficiency of egg protein utilization at maintenance and sub-maintenance levels in young men. J. Nutr. 103:1164-1174.
Young, V.R., L. Fajardo, E. Murray, W.M. Rand, and N.S. Scrimshaw. 1975. Protein requirements of man: Comparative nitrogen balance response within the submaintenance-to-maintenance range of intakes of wheat and beef proteins. J. Nutr. 105:534-544.
Young, V.R., M. Puig, E. Queiroz, N.S. Scrimshaw, and W.M. Rand. 1984. Evaluation of the protein quality of an isolated soy protein in young men: Relative nitrogen requirements and effect of methionine supplementation. Am. J. Clin. Nutr. 39:16-24.
Young, V.R., D.M. Bier, and P.L. Pellet. 1989. A theoretical basis for increasing current estimates of the amine acid requirements in adult man with experimental support Am. J. Clin. Nutr. 50:80-92.
Zello, G.A., P.B. Pencharz, and R.O. Ball. 1990. Phenylalanine flux, oxidation and conversion to tyrosine in humans studied with 13C phenylalanine. Am. L Physiol. 259:E835-E843.
Zello, G.A., P.B. Pencharz, and R.O. Ball. 1992. Lysine requirement in young adult males. Am. J. Physiol. 264:E677-E685.
Zhao, X-H., Z-M. Wen, C.N. Meredith, D.E. Matthews, D.M. Bier, and V.R. Young. 1986. Threonine kinetics at graded threonine intakes in young men. Am. J. Clin. Nutr. 43:795-802.
DISCUSSION
ROBERT NESHEIM: Thank you. Are there questions? Yes, sir, John.
JOHN VANDERVEEN: I am interested in the reality of the current situation. At the present time, our troops are getting plenty of protein in amounts far beyond this whole discussion. But the issue of adaptation raises the issue of survival. If, indeed, higher levels of protein intake change our metabolism to the point where additional adaptation, if you will, occurs in our needs, are we
putting soldiers at risk to survive starvation or low protein intake for a period of time on rations?
D. JOE MILLWARD: I think it is a very complicated question. I think that it is quite possible that we are putting soldiers at risk. Certainly, the view that Chittenden expressed was that high protein intakes represent a metabolic load that is hard to deal with.
Now, Mike Rennie will present data on physical activity and intakes and deal with this problem to a certain extent. But, in my view, the real point about the human response to protein intake is that we adapt our metabolic demand according to how much we eat. The point about any individuals who stuff themselves with very high-protein diets are two things. Firstly, as soon as they stop taking in that high protein diet, they lose lean tissue very, very rapidly. So the soldier who suddenly finds himself without food for a few days on a background of a high protein diet will lose more lean body mass than somebody who has been on a lower intake.
Secondly, the response to exercise, I think, can be shown to induce a bigger loss when one's background intake is higher. Now, there are things about infection and all of that which complicate the issue and that I do not think we have an answer to.
ROBERT NESHEIM: Yes?
G. RICHARD JANSEN: Joe, I guess, having been recruited to work on the ill-fated lysine program at Dupont 40 years ago, this is like dèja vu all over again.
I remember a 1960 conference in Chicago at the Federation Meetings discussing protein reserves. If you forget the turnover, it was the flux data. It is the same argument basically. But it seems to me that your position as you have published it is more agnostic than atheistic to Vernon's dogma here. And it seems to me you have not really addressed his argument that there is an advantage in terms of metabolic control to that increased oxidation. And, in fact, in your slide there you said, number one, that we basically cannot determine the indispensable requirements because we do not have enough data. However, you say protein scoring is invalid. It seems to me that those two statements cannot be made together. If we cannot define it, we cannot determine what is valid or invalid yet.
D. JOE MILLWARD: Okay. The point about protein scoring is that it is a procedure that was adopted for quality assessment in the rat because it was shown to work. In other words, scoring correlated with net protein utilization in the growing rat. The reason that it correlates is simply because for the growing
rat the requirement is quite simple and straight-forward. It has to do with the requirement for tissue deposition.
What I am saying is that for adult humans, our requirement is more complicated because it is variable. We have this fixed requirement that may well be very low. And, in fact, FAO took the view that you use the minimum requirement value and you score protein with that. If you do that, then all proteins in the world are nutritionally adequate. Now, they are only adequate if you can actually adapt to them successfully. So for a population with a habitual protein intake that is relatively high, the question is what is adequate for that population. That is what is very difficult to define, because ultimately one has to test it against that group and against the extent to which they are adapted to their intakes.
If you add the problem that there may well be other components of intake that are more important than balance, like performance, like the immune response, like any of these things, and we talked about the anabolic drive on growth and development, then it becomes even more difficult to define nutritional adequacy against this single pattern of individual amine acids. That is the point that I am trying to make.
G. RICHARD JANSEN: I assumed that. But I think, at the same time, I do not think that you addressed at all Young's argument that there is an advantage in terms of metabolic flux and control to that increased protein turnover, which then would make the argument that he has made, that the pattern should be what it is and, therefore, it would be valid. I understand the rat gross data. I have run a lot of those experiments; but you are not addressing the argument he is making.
D. JOE MILLWARD: Well, I did address it partly. That argument is made basically in the Young and Marchini paper, which he quotes in his abstract, where they show that whole body protein synthesis rates plummet dramatically as the intake of essential amine acids plummet. I think that data are wrong, to be quite honest. I think they are artifactual.
There are other data where the same diet is fed, for example, with decreasing lysine intake, but the protein turnover is measured with another amine acid, namely phenylalanine, and there is no change in the phenylalanine flux over the whole range of intake.
So I think there is a technical problem with the observation that higher intakes are associated with higher rates of protein turnover. Because I do not think there are higher rates of protein turnover with higher intakes. I do not think they actually change.
SUSAN HUDSON: I have a question. We have argued about this for years. But is there any way to make the steady isotope labeling data more precise, that is, those you say are flawed? Is there any kind of information that could be gathered that would, say for leucine, make the data better?
D. JOE MILLWARD: I think that if Vernon were here, what he would say is that over the years, the studies have gotten better. There has been a better understanding of what was going on. The way the calculations have been done has been improved. The studies that Vernon is doing at the moment in India where he looking at lysine intakes and he is using leucine balances are probably as good as you can actually get. So we do await the outcome of those studies. The difficulty is that many of the data that have been used in the past to support the MIT scoring pattern, in my view, simply are unusable because they are seriously flawed. So we are still waiting for hard data that support a better requirement pattern. Now, Denny [Bier] was a co-author of those papers, so I am sure he will want to respond.
ROBERT NESHEIM: Defray?
DENNIS BIER: Well, first, I think that Joe has probably presented the clearest and the fairest view of this subject that I have heard in the last 10 years. I would like to see it come out as clearly as it was presented here today. I would agree with you on this whole business of requirements, that is, the adaptive component and how it affects those requirements. I think it is precisely in that slide you showed us of the obligatory losses. I think that, unfortunately, that is really what we would like to know. That is what the military would like to know in the circumstances that they are addressing. How do we get at that?
I think we are trying to address a problem about intake and requirements in the face of a body that is filled with amino acids, as you pointed out in your remark about the utilization and recycling. It becomes very difficult, I think, to solve the problems in that context when really what we are looking for is some change from the baseline.
In addition, the balance studies really do not tell us in any way, of course, what happened to what was in the body at that time and whether or not there has been an advantage or disadvantage to what it took to get the balance in the context of these sort of intakes.
So adding functional measurements, adding things like whole body nitrogens or whole body potassium, something that really lets us know over the long term what happened to the body proteins, would be very useful.
Now, to go back to the models themselves, my position on this, and I have stated it many times, is that the models are not very good. But the original data that went into some of these calculations (I am not going to talk about scoring
patterns because I have no idea what they even mean), but, the original data that went into the calculations were based not on any complicated model but simply on the amount of C13 recovered outside of the body during the course of the tracer study. Admittedly, that has its own set of problems. But, if anything, most of those problems entail underestimation of loss rather than overestimation.
If you just take those numbers, they were those net losses that generated the initial part of this. So, admittedly, they were done at MIT with young, healthy adult men, adapted longterm, and using a different protein technique for a maximum of three weeks. I have no idea, frankly, whether that applies to Northern Thais who eat rice or adapt to a different type of protein for the rest of their lives.
K. SREEKUMARAN NAIR: I want to echo what was said. This is a complex problem. My concern is we are going to try to solve this problem by a very simplistic approach.
What we need is a long-term study on the effects of different proteins on performance changes. They are not easy to perform.
D. JOE MILLWARD: I think we have tried many times to erect very complicated models to understand amine acid and protein metabolism. I have always taken the view that, if you cannot actually measure the total amount of body protein that is there and whether it is going up or whether it is going down, any other information is basically irrelevant. You have got to start with good data about what is the state of the overall system, i.e. the lean body mass. And, while that is a simplistic approach, it is a necessary precondition to answering any of the other questions.
DENNIS BIER: But the balance method does not allow that—the only way to do that is with the total body nitrogen.
D. JOE MILLWARD: Exactly. I think that we need much more data on whole body nitrogen.
NANCY BUTTE: Joe, the committee has been asked to address a very straightforward question. Is the military RDA for protein, which is one hundred grams per men and 80 for women, appropriate? Given the current state of knowledge with the techniques we have, would you recommend that they even attempt to revise that right now?
D. JOE MILLWARD: Well, there is certainly, in my mind, no evidence to suggest that it is inadequate. The only issue is whether that level is too much.
NANCY BUTTE: And what would you advise them if you were asked to advise them at this time? Do you think the recommendations are too high?
D. JOE MILLWARD: Well, I think it is. To be quite honest, I do not think we have yet done the right studies. I mean, if you work on the basis of this very significant series of studies that were done over three or four years on Army recruits and young athletes, then you would have to conclude that the current requirements are too high and that soldiers may well perform better on a lower intake. Mike [Rennie] is going to present data this afternoon that suggest that for physical activity, there may well be benefits of lower protein intakes.
ROBERT WOLFE: I would just like to follow up on Dr. Jansen's point. I do not know that it can be so readily dismissed that a higher protein intake affects protein turnover, because even your own anecdotal comment, that if you eat a higher protein intake and then stop the protein intake, you will lose protein faster. The only way that could occur would be if you have a higher protein turnover; the higher protein intake caused the higher protein turnover. The only way you would lose it more rapidly would be if it were turning over more rapidly before you stopped. But, furthermore, there are some studies that have done what you have suggested and avoided the problem of using the same amino acid tracer. Because we performed a study a number of years ago, albeit not in normal subjects, but in severely burned patients in a crossover study in which they were given different levels of protein, and three different essential amino acids were used, and all three tracers showed parallel increases in whole body protein turnover when the protein intake was increased, although none indicated an improvement in balance.
So I think the fact that the same amino acid tracer as the test compound has been used with different levels of protein intake may not be valid, and that it is a jump to go from that observation to the conclusion that there is no effect of protein intake on protein turnover.
D. JOE MILLWARD: Let me say that it was not an anecdotal statement. We have published a series of studies where we used a multiple tracer approach. We used N15, we used C13 leucine, we used deuterated phenylalanine to address the question of whether the level of protein intake affects the overall rate of protein turnover. The outcome of that experiment was that from an intake of .3 grams per kilogram, up to an intake of 2.5 grams per kilogram over three weeks, there was no effect on the overall replacement rate. However, those subjects who
were in positive balance obviously had a higher rate of synthesis than proteolysis and those on the lower intake had a lower rate of synthesis than proteolysis. But, if you calculated rate of replacement as the lower of the two processes, that rate did not change over the entire range.
DENNIS BIER: Two comments, first on the last statement. If you change the protein intake, one way or another you at least have to change components of the protein turnover. That is, you have to deal with the protein that comes in. So the total protein turnover may not change, but how it is distributed has to be changed, because you have a new term in the equation that is introducing protein into the body. So the fact is that something has to happen, not to the total number [turnover] necessarily, but to how it is distributed. Our models do not really allow us to analyze that.
The second comment goes back to Chittenden's study. How did he control for the training effect on this?
D. JOE MILLWARD: Well, that was the basis of the problem with his initial study, because he did that study on army recruits. There was this enormous increase in strength. He concluded immediately that it was the weekly measurement over five months that trained them and that was responsible for the increase in strength. So he only controlled for it by recruiting elite athletes. He had the U.S. National Decathlon equivalent champion among his subjects. They were as highly trained as they could get. That was how he got the results that he did. But it is not a perfect study. But it is the only study that has ever seriously addressed the problem. Just because it was published ninety years ago, it is completely ignored.
DENNIS BIER: Well, you know, it is not on Medline!
JEFF ZACHWIEJA: There are some recent data that are germane to that issue. Others may help me out with this. I think that Bill Evans' group (when he was still at Tufts), recently published data that suggested that exercise training in older individuals, whether they had a dietary protein intake of .8 grams per kg or 1.6 grams per kg, essentially achieved the same level of muscle strength and I think those individuals who were on the lower protein intake were still in positive nitrogen balance, but not as great as those on the higher protein intake. But, in terms of performance, the increments in performance were very similar on those two types of diets. These results spoke to the issue of the body being more efficient or becoming more efficient on the lower protein intake to achieve essentially the same functional outcome.
ROBERT NESHEIM: I think we could have more discussion on this, and we will have more time later.
HARRIS LIEBERMAN: I just have one more comment in regard to that issue of one hundred g protein per day. There is a lot of confusion about that. The really critical issue for us is not the MRDA level of protein that is recommended but the standard for operational rations. Right now, the standard operational ration must include at least a hundred grams of protein. That typically is not fully consumed. If you lower that ration content, a hundred grams or 90 grams will not be consumed; substantially less than that will be consumed. So when we think about it and talk about it, please keep in mind that what really is critical for us is what we recommend to the ration developers at Natick, the minimum level of protein they can include in a combat ration.
JOHN VANDERVEEN: I have one last comment. The concern about ration design depends on how you calculate that 100 grams of protein as well. What protein quality factor do you use? Do you have a cut-off in what you consider protein? If the protein has an amino acid score that is such that you have a cutoff at 20 percent or 30 percent, or whatever, is that counted in? I think that those are issues.
I guess maybe the last question I would raise is whether there is any thinking as to how long the amino acids are available in the pool so that you can take advantage of a lower-quality protein at some later time in the day or at a different time?
D. JOE MILLWARD: Well, there are good data on that. There are good data on muscle biopsy studies, which I referred to in my talk, of the rate at which amino acids disappeared from the intracellular muscle pool after a meal. It is quite clear that, whereas leucine, all three BCAAs, the aromatic amino acids, and the sulfur amino acids disappear quite quickly, lysine and threonine stick around for a long time. Seven hours after an albumin meal, lysine and threonine levels are still elevated, whereas all of the branch chains have fallen below baseline levels. I think that what we know about the Kms of the catabolic pathways for lysine and threonine would indicate that the body does not turn up their oxidation quickly after a meal in the same way that it does for the BCAAs.
ROBERT NESHEIM: One more question.
PATRICK DUNNE: It is more of a request for background information. It is really our group that, over the years, has been doing the nutritional assessment of the ration contents. What we do not know is the amino acid composition of
the diets. It is way too expensive to do that. We can make some guestimates like anyone else can from the databases. We would say that the majority of our protein intake is coming from meat. But that is switching as we look at newer concepts in our rations, where we have more vegetarian items. We are getting better at providing bread to people in the field and developing shelf-staple breads. That is a changing phenomenon and may be an area where we need to gather more data.
JOHN VANDERVEEN: So you are telling us that you take N values and multiply times the conversion factor and that is how you estimate ration protein content?
PATRICK DUNNE. I would say that is correct, with some minor corrections using different factors for dairy versus muscle proteins. You typically perform kjeldahl nitrogen assays and that is what you get. You get approximate values for N, and that forms the database. We are trying to reconcile that database with the nutrient content of the meal, ready to eat (MRE)-15 as best we can. That is the level of knowledge.
D. JOE MILLWARD: In fact, I think it is highly unlikely that protein quality is going to be an issue in the sort of rations that are provided unless they are really quite unusual. In other words, unless they are basically unsupplemented wheat. There have been a number of papers recently that have used rats to do protein quality assays of basically vegetarian diets, and these have shown that vegetable-type mixtures can have higher NPU values than those for meat and milk. So, unless these military diets were really very peculiar, then I do not think that the amine acid composition would become an issue.
PATRICK DUNNE: Right. However, in the field setting, we probably have the normal level of intake of protein, lower in dairy proteins than you would have in the garrisons just because of what is shelf stable. We do look at cross-linking effects and do not think it is a problem to really reduce the quality of protein during the normal storage period, which could be three-plus years.
We also have some data relative to the antioxidant issue. Until recently, when more commercial items were added to our rations, we were not feeding our troops a lot of BHA and BHT. We were feeding them lots of vitamin C and vitamin E. As we add more commercial items, you are going to see changes in macro- and micronutrients, something for the good.
ROBERT NESHEIM: Thank you very much. I think it has been a very interesting morning. It is time to break for lunch.
















































