
Protein and Amino Acids, 1999
Pp. 309-329. Washington, D.C.
National Academy Press
15
Supplementation with Branched-Chain Amino Acids, Glutamine, and Protein Hydrolysates: Rationale for Effects on Metabolism and Performance
Anton J. M. Wagenmakers1
INTRODUCTION
Endurance athletes in periods of intense training with a high daily workload tend to believe that they need more protein and amino acids in the diet than sedentary subjects. According to Munro (1964b), this belief originates from the 1840s when the German physiologist Von Liebig hypothesized that muscle protein was the main fuel used to achieve muscular contraction. After Von Liebig's hypothesis had been invalidated around 1870 by the first experimental data on urinary nitrogen excretion during and following prolonged demanding exercise, many exercise physiologists took the opposite stand and for the next 100 years disregarded the amino acid pool in muscle as playing any role of significance in exercise and energy metabolism. Despite this 100 percent change in scientific opinion, the general public continued to believe that a high
workload leads to an increase in dietary protein needs. Industrial workers were and are served bigger steaks the more demanding their daily workload is. Milk is promoted as the "white motor" in advertisements. For these reasons, tons of protein are sold each year to strength athletes, as they believe they can only make their muscles grow when they ingest protein supplemental to their usual diet, and to endurance athletes, because they believe they break down muscle protein during training and competition.
The high-purity crystalline amine acids now available have created an easy new health food and supplementation market among the athletes who already believed in the beneficial effect of protein. Advertisements claim that free amine acids and small peptides are a better source of amine acids to support high net rates of protein deposition because they are more readily absorbed than whole dietary protein. The rapid absorption of amine acids leads to high plasma and tissue amine acid concentrations, In fact, it seems to drive the amine acids primarily into the oxidation pool, since the Michaelis constant (Km) of the oxidative enzymes is much higher than the Km of the enzymes involved in protein synthesis. By definition, increased oxidation means a reduced net rate of protein deposition, at least in the first hours after ingestion. Apart from serving as building blocks for protein synthesis, several amine acids have unique functions in human physiology, and they are marketed as supplements to support these functions. However, the scientific evidence to support the claim that they are needed in addition to the habitual ingestion of dietary proteins is usually lacking.
This chapter focuses on several aspects of protein and amine acid metabolism in endurance exercise and on the effect of amine acid and protein supplementation on performance-related aspects of metabolism. First presented will be the author's opinion on whether endurance exercise leads to net protein catabolism, increased amine acid oxidation, and therefore an increased protein requirement. A detailed overview is given elsewhere in this volume (see also Rennie, 1996). Subsequently, the rationale for supplementation with branched-chain amine acids (BCAAs) and glutamine and the effects on metabolism and performance will be evaluated. Finally, suggestions will be presented that the glycogen resynthesis rates in muscle following exercise can be accelerated by the combined ingestion of carbohydrate and protein (Zawadzki et al., 1992) and by glutamine infusion (Varnier et al., 1995).
PROTEIN AND AMINE ACID METABOLISM IN ENDURANCE EXERCISE
Recent stable isotope tracer studies have reopened the discussion on whether net protein breakdown (protein synthesis < protein degradation) and increased amino acid oxidation occur during prolonged endurance exercise at the whole body level. If the answer is yes, the next question is whether net protein breakdown occurs in muscle or in the gut and liver area. Stable isotope
tracer methods, although valuable in other areas of nutrition and medicine, do not clarify the issue. Some amino acid tracers and some studies report a net protein breakdown during exercise at 40 to 70 percent maximal oxygen consumption (VO2max) for 1 or 2 or maximally 4 hours, whereas others do not (for a review see Rennie, 1996).
When considering whether net protein break-down occurs during prolonged high workload exercise in the field, it is important to know whether or not carbohydrates are ingested and whether energy balance can be maintained around the clock. Many laboratory studies using tracers were conducted in subjects after overnight fasting without carbohydrate ingestion, a condition that favors net protein breakdown during exercise but bears little relevance for athletes. A triathlete or a Tour de France cyclist with an energy expenditure of over 30 MJ/(7143 kcal/d) in a stage with five to six mountain passes at altitudes of over 2,000 m in the mountains (Saris, 1997) would never begin exercise without a proper breakfast and without carbohydrate ingestion before, during, and after competition (see glycogen resynthesis section of this chapter). Carbohydrate ingestion during exercise prevents activation of the branched-chain α-keto acid dehydrogenase in muscle (the rate-determining step in the oxidation of the branched-chain amino acids, Wagenmakers et al, 1991) and leucine oxidation in several tracer studies (see Rennie in this volume for references). Recently this laboratory conducted the most demanding exercise study ever performed in a laboratory setting. It was observed that 6 hours of exercise (2.5 h of cycling at 50% Wmax followed by I h of running at 11 km/hr and then again by 2.5 h of cycling at 50% Wmax; Wmax = maximum power output) by elite cyclists while ingesting carbohydrate led to an imbalance between protein synthesis and degradation (synthesis < degradation) and an increase in amino acid oxidation when a L-[l-13C]leucine tracer was used (Pannemans et al., 1997). In contrast to expectations, net protein degradation and leucine oxidation did not increase in duration, possibly because carbohydrate was ingested. Without carbohydrate ingestion, subjects could not have exercised for 6 hours. Other tracers, [2H5]phenylalanine and [15N2]urea, did not show an increase of amino acid oxidation and net protein breakdown during exercise. Therefore, it is unclear whether proteins are broken down in highly demanding endurance and ultra-endurance exercise under laboratory conditions and in the field. It is difficult to believe that during a stage in the Tour de France with a cyclist's energy expenditure of > 20 MJ within 8 h of cycling, that there will be no increase in protein breakdown and amino acid oxidation. During such events, the rate of carbohydrate oxidation far exceeds the maximal amount of carbohydrate ingestion, and these athletes, at least during the last hours of the race, are in a net negative energy balance. The combination of a negative energy balance and highly demanding muscle contractions leads to net protein breakdown in the muscle. Evidence of this has been obtained during one-leg knee-extensor exercise with normal and low muscle glycogen content (Van Hall et al., 1995a; Wagenmakers et al., 1996; Van Hall, 1996). In one-leg knee-
extensor exercise, approximately 3 kg of skeletal muscle contracts. Maximal oxygen consumption and the metabolic rate of the active muscle was two- to three-fold higher under these conditions than for whole body dynamic exercise (Andersen and Saltin, 1985). The net muscle production rate of threonine, lysine, and tyrosine (amine acids that are not transaminated and metabolized in muscle) and of the sum of the non-metabolized amine acids was 9- to 20-fold higher during one-leg exercise than at rest, implying that the one-leg exercise led to a massive increase in the net muscle protein degradation rate (protein degradation minus protein synthesis). During exercise with the low-glycogen-content leg, the release of these amine acids was again 1.5- to 2.5-fold higher than during exercise with the normal-content-leg. This indicates that net muscle protein degradation increased further when the active muscle was glycogen depleted (Van Hall, 1996).
Another observation of a high release of non-metabolized amine acids from the leg muscles, reflecting net protein catabolism, occurred during two-legged cycling in patients with McArdle's disease (Wagenmakers et al., 1990). These patients have no or no active glycogen phosphorylase in muscle and, therefore, cannot use muscle glycogen as an energy source during exercise. During incremental exercise they reached a maximal workload of only 40 to 100 W. Due to the energy deficiency in their muscles these subjects rapidly broke down their muscle proteins, even at relatively low-intensity exercises. These data suggest that during demanding endurance or ultra-endurance exercise in the field, where muscle glycogen depletion cannot be prevented despite oral carbohydrate ingestion, net protein breakdown will occur in skeletal muscle once the glycogen stores have been emptied. Demanding endurance exercise may also lead to net protein breakdown in gut and liver, but direct evidence in humans has not been reported.
BRANCHED-CHAIN AMINE ACIDS
Central Fatigue Hypothesis
Prolonged exercise that leads to glycogen depletion inevitably causes fatigue. Fatigue is defined physiologically as the inability to maintain power output. To the endurance athlete, this means a gradually increasing sense of effort, sometimes even discomfort and pain, and the need to reduce the speed. Both peripheral (i.e., muscle [Fitts, 1994]) and central (brain) mechanisms have been suggested to play a role in the development of fatigue in endurance exercise. In 1987, Newsholme and colleagues launched the "central fatigue hypothesis" as an important mechanism contributing to the development of fatigue during prolonged moderate-intensity exercise. During exercise, free fatty acids (FFA) are mobilized from adipose tissue and via the blood transported to muscle to serve as fuel. As a consequence, the blood FFA concentration will increase. FFA and the amine acid tryptophan both bind to albumin and compete
for the same binding sites. Tryptophan will be displaced from binding to albumin by the increasing FFA concentration, and therefore, the free tryptophan concentration in the blood will rise. Simultaneously, the increased oxidation of BCAA in muscle (Wagenmakers et al., 1991) leads to a decrease of the sum concentration of the BCAA in the blood, and the free tryptophan:BCAA ratio, therefore, will increase substantially. The increase in this ratio leads to increased tryptophan transport across the blood-brain barrier, since BCAA and tryptophan compete for carrier-mediated entry into the central nervous system by the large neutral amino acid (LNAA) transporter (Knott and Curzon, 1972; Chaouloff et al., 1985). Once taken up, conversion of tryptophan to serotonin (5-hydroxytryptamine) occurs and leads to a local increase of this neurotransmitter (Knott and Curzon, 1972; Chaouloff et al., 1985). This increase has been found in certain brain areas in the rat, but it has not been established in humans. According to the central fatigue hypothesis, the increase in serotoninergic activity subsequently leads to central fatigue, forcing athletes to stop exercise or reduce running or cycling speed. Neurobiologists have established that serotonin plays a role in the onset of sleep and is a determinant of mood and aggression. It is uncertain, however, whether it also plays a role in fatigue experienced during prolonged exercise as suggested in the central fatigue hypothesis. One of the implications of the central fatigue hypothesis is that ingestion of BCAA could reduce the exercise-induced increase in brain tryptophan uptake and thus delay fatigue and give athletes the ability to push on for a more prolonged period even when the peripheral fatigue mechanism has come into operation. BCAA, in other words, would be ergogenic. Another implication is that ingestion of tryptophan prior to exercise would reduce time to exhaustion. Tryptophan, in other words, would be ergolytic.
Effects of Ingestion of Branched-Chain Amino Acids and Tryptophan on Endurance Performance
The effect of BCAA ingestion on physical performance was investigated for the first time in a field test by Blomstrand et al., 1991. Male subjects (n = 193) were studied during a marathon in Stockholm. Subjects were randomly (without matching) divided into an experimental group that received 16 g of BCAA in plain water during the race and a placebo group that received flavored water. Subjects also had ad libitum access to carbohydrate (CHO)-containing drinks. No difference was observed in the marathon time of the two groups. However, when the original subject group. was divided into fast and slower runners, a small significant reduction in marathon time was observed in the slower runners only. In retrospect, this first study has been the only one to claim a positive effect of BCAA ingestion during exercise. However, three criticisms can be raised against its design, and each of these points may have biased the data obtained: 1. In a performance test investigating a potentially ergogenic effect, subjects in the two groups should have been matched for previous
performance; 2. CHO intake and nutritional status should have been controlled and matched in the two groups; and 3. division of subjects in a group of fast and slower runners, taking an arbitrary marathon time as selection criterion is not in accordance with accepted statistical methods.
Varnier et al. (1994) investigated six moderately trained subjects after glycogen-depleting exercise, followed by overnight fasting. Subjects were investigated the morning after the fast during graded incremental exercise to exhaustion and received an intravenous infusion of BCAA (260 mg/kg/h for 70 min) or saline only. No significant differences were observed between the tests in total work performed.
Blomstrand and colleagues (1995) also investigated performance in the laboratory in five male endurance-trained subjects during exhaustive exercise on a cycle ergometer at a work rate corresponding to 75 percent of VO2max after reduction of their muscle glycogen stores. Subjects were given, during exercise and in random order, a 6 percent CHO solution containing 7 g/liter of BCAA, a 6 percent CHO solution, and flavored water. The positive effect of the field test was not confirmed in this controlled laboratory study, because no difference in performance was seen when the subjects were given CHO + BCAA or only CHO.
Madsen and colleagues (1996) recently investigated performance in nine trained cyclists in a 100-km timed trial in the laboratory. Subjects used their own bikes at a freely chosen power output, simulating field conditions, and were studied while ingesting flavored water only (placebo), a 5 percent CHO solution (66 gram/h), and CHO (66 gram/h) plus BCAA (6.8 gram/h). There was no difference between treatments in the time needed to finish the 100-km trial.
Based on the two implications of the central fatigue hypothesis, (BCAA ingestion improves performance and tryptophan ingestion reduces time to exhaustion), Van Hall and colleagues (1995b) designed an experiment in which both aspects of the central fatigue hypothesis were investigated. Ten endurance-trained male athletes were studied during cycle exercise at 70 to 75 percent of Wmax, while ingesting, in random order and double blinded, drinks that contained 6 percent sucrose (control) or 6 percent sucrose supplemented with (1) tryptophan (3 g/liter), (2) a low dose of BCAA (6g/liter, comparable to doses used by Blomstrand et al., [1991]), and (3) a high dose of BCAA (18 g/liter). These treatments greatly increased the plasma concentration of the respective amine acids to values well outside the normal physiological range. By measuring the concentration of all amine acids competing for transport by the LNAA carrier, Van Hall and colleagues (1995b) were able to calculate the rate of unidirectional influx of circulating plasma tryptophan into the brain using kinetic parameters of transport of human brain capillaries reported by Hargreaves and Pardridge (1988). These calculations showed that the administered BCAAs only reduced tryptophan transport at exhaustion by 8 to 12 percent, while tryptophan ingestion caused an increase of 600 to 1,900 percent (depending on the use of free or total tryptophan concentration in the
calculations). Despite these massive differences in tryptophan transport, time to exhaustion was not different between the four treatments. Van Hall and colleagues, therefore, concluded that manipulation of tryptophan supply to the brain by ingestion of BCAA and tryptophan in CHO containing drinks either did not change the serotonin concentration in relevant local areas in the brain or a change in serotoninergic activity during prolonged exercise contributed little to mechanisms of fatigue. Two earlier studies (Segura and Ventura, 1988; Stensrud et al., 1992) investigated the effect of lower doses of tryptophan. Segura and Ventura (1988) reported that 1.2 g of L-tryptophan supplementation taken in 300-mg doses over a 24-hour period before exercise increased total exercise time by 49 percent in 12 subjects who were running at 80 percent of VO2max (which fully contradicts to the central fatigue hypothesis). The results of this study were questioned by Stensrud et al., (1992), who studied 49 well-trained males in a randomized, double-blind, placebo experiment. Subjects in the tryptophan group (n = 24) and placebo group (n = 25) were matched for performance (maximal oxygen uptake, anaerobic threshold, and speed during an all-out run). Tryptophan ingestion (again 1.2 g over a 24-h period prior to the run) had no effect on running performance, when subjects ran until exhaustion at a speed corresponding to 100 percent of their VO2max.
Summary of Performance Studies with BCAA and Tryptophan
Neither BCAA nor tryptophan ingestion has an effect on endurance performance in healthy subjects. Thus, performance studies have not provided experimental support for the central fatigue hypothesis as playing a dominant role in fatigue mechanisms during prolonged exercise.
Interaction of the BCAA-Aminotransferase Reaction with the Tricarboxylic Acid (TCA) Cycle in Muscle
The carbon flux in the tricarboxylic acid (TCA) cycle is one of the processes which determines the rate of ATP turnover in skeletal muscle. One possible way to achieve an increase in TCA cycle activity when going from rest to exercise (to meet the increased energy demand of exercise) is to increase the concentration of the TCA cycle intermediates in muscle such that more substrate is available for the individual enzymatic reactions. This increase in concentration has been observed for the most abundant TCA cycle intermediates during early exercise (Essen and Kaijser, 1978; Sahlin et al., 1990). It is achieved by rapid conversion of the muscle glutamate pool into α-ketoglutarate (Van Hall et al., 1995a). The reaction used to achieve that increase is the alanine aminotransferase reaction: glutamate + pyruvate ↔ α-ketoglutarate + alanine (Van Hall et al., 1995a). The alanine aminotransferase reaction is a near-equilibrium reaction, which implies that the increase in muscle pyruvate
concentration that occurs at the start of exercise due to an acceleration of glycolysis will automatically lead to production of alanine and α-ketoglutarate and consumption of glutamate. After observing the early increase in concentration of TCA cycle intermediates, Sahlin et al., (1990) observed a subsequent gradual decrease in the intermediate in human subjects exercising until exhaustion at 75 percent VO2max . This author and colleagues (Wagenmakers et al., 1990, 1991) have hypothesized that increased oxidation of the BCAAs plays an important role in that subsequent decrease of the concentration of the TCA cycle intermediates. Branched-chain α-keto acid dehydrogenase (BCKAD) is increasingly activated during prolonged exercise, leading to glycogen depletion (Wagenmakers et al., 1990, 1991; Van Hall et al., 1996), and an increase in oxidation by definition will increase the flux through the BCAA aminotransferase step. In the case of leucine, this reaction will put a net carbon drain on the TCA cycle as the carbon skeleton of leucine is oxidized to 3 acetyl-coenyzme A (CoA) molecules, and the aminotransferase step removes α-ketoglutarate: leucine + α-ketoglutarate ↔ 3 acetyl-CoA + glutamate. Increased oxidation of valine and isoleucine will not lead to net removal of TCA cycle intermediates, as the carbon skeleton of valine is oxidized to succinyl-CoA and that of isoleucine to both succinyl-CoA and acetyl-CoA. Net removal of α-ketoglutarate via leucine transamination can be compensated for by the alanine aminotransferase reaction (see above) as long as muscle glycogen is available and the muscle pyruvate concentration is kept high. However, because activation of the BCKAD complex is highest in glycogen-depleted muscle, this mechanism eventually will lead to a decrease in the concentration of TCA cycle intermediates; a reduction in TCA cycle activity; and a reduction of the ATP turnover rates, which via cellular mediators, will offset the known muscle fatigue mechanisms (Fitts, 1994).
BCAAs after oral ingestion are rapidly taken up in the blood and extracted by the leg muscles, and this is accompanied by activation of the BCKAD complex at rest and increased activation during exercise (Van Hall et al., 1996). This could imply that the indicated carbon drain on the TCA cycle is greater after BCAA ingestion and that by this mechanism, BCAA ingestion leads to premature fatigue during prolonged exercise, leading to glycogen depletion.
Wagenmakers et al., (1990) investigated the effect of BCAA ingestion (20 g) 30 minutes prior to graded incremental exercise in two patients with McArdle's disease (muscle glycogen phosphorylase deficiency). Due to their disease, these patients can be regarded as an experiment of nature. From the metabolism in their muscle, one learns what happens in a muscle without glycogen. BCAA ingestion reduced the Wmax that was reached during the incremental exercise test by about 20 percent, and heart rate and perceived exertion were higher at the same workload. Ingestion of the keto acids of the BCAA (without the amine group) improved performance in the patients. These observations might suggest that, by the proposed mechanism, BCAA supplementation has a negative effect on performance in healthy subjects in
conditions where the glycogen stores have been completely emptied by highly demanding endurance exercise. However, with co-ingestion of carbohydrate, BCAA ingestion did not change the time to exhaustion in healthy subjects (Van Hall et al., 1995b; Blomstrand et al., 1995; Madsen et al., 1996).
Effect of BCAA Ingestion on Plasma Ammonia Concentrations and Muscle Ammonia Production During Exercise
BCAA ingestion increased the production of ammonia by muscle and plasma ammonia concentration during exercise both in patients with McArdle's disease (Wagenmakers et al., 1990) and in healthy controls (Van Hall et al., 1995a; MacLean et al., 1996; Madsen et al., 1996). Because it has been suggested that ammonia leads to central fatigue and loss of motor coordination (Banister and Cameron, 1990), great care is indicated with the use of BCAA supplements in sport (e.g., tennis and soccer) and military activities where performance depends on motor coordination.
GLUTAMINE
Glutamine is the main end product of muscle amino acid metabolism both in the overnight fasted state and during feeding (Marliss et al., 1971; Chang and Goldberg 1978a; Wagenmakers et al., 1985; Elia et al., 1989; Wagenmakers and Soeters, 1995; Nurjhan et al., 1995). Alanine most likely only serves to export part of the amino groups (Chang and Goldberg 1978b; Wagenmakers et al., 1985). Glutamine is the most abundant amino acid in human plasma (600-700 μmol) and in the muscle free amino acid pool (20 mmol; 60% of the intramuscular pool excluding the nonprotein amino acid taurine). The synthesis rate of glutamine in muscle is higher than that of any other amino acid. Extrapolations of limb production rates in the fed and fasted state support that between 10 and 25 g of glutamine are synthesized in the combined human skeletal muscles per day (Marliss et al., 1971; Elia et al., 1989). Tracer dilution studies even indicate that 80 g of glutamine are produced per day (Darmaun et al., 1986), but this may be a methodological overestimation due to slow mixing of the glutamine tracer with the large endogenous glutamine pool in muscle (Van Acker et al., 1996). Furthermore, although muscle is the main glutamine-producing tissue, other tissues (e.g., adipose tissue, liver, and brain) may also contribute to the rate of appearance of glutamine in the plasma pool that is measured by tracer dilution techniques.
The reason for this high rate of glutamine production in muscle is probably that glutamine plays an important role in human metabolism in other organs. Sir Hans Krebs (1975) wrote: ''Maybe the significance of glutamine synthesis is to be sought in the role of glutamine in other organs, as a precursor of urinary ammonia and as a participant in the biosynthesis of purines, NAD+, amino
sugars and proteins. Glutamine is an important blood constituent, present in higher concentrations than any other amine acid, presumably to serve these various functions. Muscle may play a role in maintaining the high plasma concentration of glutamine." Glutamine has been shown to be an important fuel for cells of the immune system (Ardawi and Newsholme, 1983) and for mucosal cells of the intestine (Windmueller and Spaeth, 1976; Souba, 1991). Low muscle and plasma glutamine concentrations are observed in patients with sepsis and trauma (Vinnars et al., 1975; Rennie et al., 1986; Lacey and Wilmore, 1990), conditions that also are attended by mucosal atrophy, loss of the gut barrier function (bacterial translocation), and a weakened immune response. Although the link between the reduced glutamine concentrations and these functional losses has not been fully underpinned by experimental evidence, the possibility should seriously be considered that it is a causal relationship. Due to its numerous metabolic key functions and a potential shortage in patients with sepsis and trauma, glutamine has recently been proposed to be a conditionally essential amine acid (Lacey and Wilmore, 1990), that should be added to the nutrition of long-term hospitalized, critically-ill and depleted patients. These patients have a reduced muscle mass due to continuous muscle wasting and as muscle is the dominant site of glutamine production, they probably also have a reduced capacity for glutamine production.
Plasma Glutamine Concentrations Following Prolonged Exercise
During exercise, plasma glutamine concentrations may increase, decrease, or remain unchanged depending on the type, duration, and intensity of exercise. However, lowered plasma glutamine concentrations were observed for a period of several hours following demanding endurance exercise leading to glycogen depletion (e.g., Van Hall, 1996). The lowest concentration is seen some 2 hours after exercise. It takes more than 7 hours before the concentration has returned to the pre-exercise resting level.
Plasma Glutamine Concentrations in Overtrained Athletes and in Tour de France Cyclists
Parry-Billings et al., (1992) observed lower plasma glutamine concentrations (503 ± 12 μmol) in 40 international standard athletes diagnosed as overtrained than in 36 matched control athletes (550 ± 14 μmol, mean ± SD). During continued intense competition without symptoms of overtraining, decreases in plasma glutamine have also been observed. Saris (1997) measured plasma glutamine in Tour de France cyclists. During the tour, a cyclist pedals about 4,000 km (2,500 miles) over a period of 22 days with only one rest day during the race. The mean energy expenditure per day is about 25 MJ and peak expenditures of 38 MJ/day have been observed. Plasma glutamine concentration
fell gradually from 568 ± 56 μM (m ± SEM, n = 8) at the start to 479 ± 65 μM on day 22 (Figure 15-1).
Link to Immune System
It has been suggested in the literature that there is a high incidence of upper respiratory tract infections in athletes after strenuous prolonged exercise and that overtrained athletes have an even more severe impairment of the immune system, which makes them more prone to infections (Brenner et al., 1994; Hoffman-Goetz and Pedersen, 1994). Parry-Billings et al., (1992) suggested that the decrease in plasma glutamine following endurance exercise created an "open time window" in which athletes are more susceptible to catching a common cold or infection. It was also concluded that the lower plasma glutamine concentration in overtrained athletes contributed to the "weakened immune system" in this condition, given the proposed importance of glutamine for cells of the immune system (Ardawi and Newsholme, 1983; Parry-Billings et al., 1990). However, the link between glutamine metabolism and the immune system is not as strong as in critically ill or bum patients who tend to have much lower plasma glutamine concentrations and also low muscle glutamine stores. These patients have a reduced muscle mass and, therefore, may have a lower endogenous capacity to synthesize glutamine. They often completely lack the delayed-type hypersensitivity response, whereas this aspect of the immune function has not yet been reported to be abnormal in overtrained or heavily training athletes.
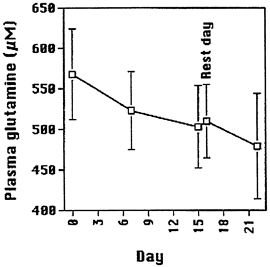
FIGURE 15-1 Plasma glutamine concentrations were measured (by HPLC) in 8 Tour de France cyclists on day 0, 7, 15 (resting day), 16 and 22. Values are given as means ± SEM. Source: Data from Saris (1997).
Castell et al., (1996) gave glutamine supplements or a placebo immediately after exercise and 2 hours later to athletes participating in a marathon or ultramarathon and reported lower infection rates in the glutamine group in the week following the race. However, infection levels were monitored by questionnaires. All symptoms reported by subjects under the heading of cold, cough, sore throat, or influenza occurring within 1 week after the race/training session were taken to reflect an infection. More valid, verifiable, and objective evidence with good functional measurements of the immune system will be needed before it can be concluded that: (1) the reported decreases in plasma glutamine concentration are linked to immune system impairments in overtrained athletes and in the period following demanding exercise, and (2) glutamine supplementation reduces infections. Interestingly, an increase of the protein content of the diet to 2.2 g/kg/day in healthy subjects also decreased the plasma glutamine concentration to 411 ± 28 μmol (mean ± SEM; n = 5; Matthews and Campbell, 1992). However, there is no reason to believe that this decrease, which is larger than the decreases mentioned above, impairs the immune system in these individuals. If anything, a high protein intake strengthens the immune system.
GLYCOGEN RESYNTHESIS FOLLOWING EXERCISE
During prolonged exercise at intensities between 50 and 85 percent VO2max, muscle glycogen from a quantitative point of view is by far the most important energy source (Romijn et al., 1993). In elite male endurance athletes, between 600 and 700 g of glycogen is present in the combined skeletal muscles. Most of this glycogen is lost during prolonged moderate-to-high intensity exercise. Resynthesis of muscle glycogen after exercise is an important factor determining the time needed by such athletes to recover. This is especially important for individuals (male and female) performing demanding exercise on a daily basis (e.g., professional cyclists, marathon skaters, mountaineers). If a Tour de France cyclist fails to replenish the muscle glycogen stores fully before the next stage begins on the following day, it is likely he can not finish in the group (peleton).
To obtain maximal glycogen synthesis rates in the first 4 hours following exercise, adequate amounts of CHO should be ingested (Bergström and Hultman, 1967; Ivy et al., 1988, Reed et al., 1989). Maximal synthesis rates were obtained in healthy subjects who cycled regularly when 1.5 g/kg body weight of glucose was ingested as a glucose/glucose polymer drink immediately after glycogen-lowering exercise and 2 hours later (Ivy et al., 1988; Table 15-1). Doubling of the rate of glucose ingestion did not influence the glycogen resynthesis rate. Similar rates of glycogen resynthesis also were obtained when 3 g/kg body weight of glucose was taken orally as a drink immediately after and 2 hours after exercise, in solid form at the same points in time, and by continuous intravenous infusion (Reed et al., 1989).
TABLE 15-1 Muscle Glycogen Resynthesis Rates Following Exercise
|
Mean CHO Ingestion Rate g/kg/h |
Glycogen Synthesis Rate μmol/g Wet Weight/h |
|
0.00* |
0.5 |
|
0.75* |
5.2 ±0.9 |
|
1.50* |
5.8 ± 0.7 |
|
1.50† |
6.1 |
|
1.50 (solid)† |
6.3 ±1.6 |
|
1.50 (continuous infusion)† |
7.0 ± 0.9 |
|
0.8‡ |
5.1 |
|
0.8 + protein 0.3 g/kg/h‡ |
7.0 (+ 37%; P < 0.05) |
|
Only protein 0.3 g/kg/h‡ |
1.5 |
|
0.00 + glutamine infusion (50 mg/kg/h)§ |
1.4 ±0.3(P < 0.05) |
|
0.00 + alanine + glycine infusion§ |
0.4 ± 0.2 |
|
0.00 + saline infusion§ |
0.5 ± 0.2 |
|
0.8# |
6.5 ± 1.2 |
|
0.8 + glutamine 0.3 g/kg/h# |
5.4 ± 1.6 |
|
0.8 + wheat hydrolysate 0.3 g/kg/h# |
8.9 ± 1.6 (+ 36%; P =0.061) |
|
0.8 + whey hydrolysate 0.3 g/kg/h# |
7.7 ± 0.9 (+ 18%; P = 0.068) |
|
NOTE: CHO, amino acids and protein are taken as drinks unless indicated differently. * Ivy et al., J. Appl. Physiol. 65:2018-2023, 1988. † Reed et al., J. Appl. Physiol. 66:720-726, 1989. ‡ Zawadzki et al., J. Appl. Physiol. 72:1854-1859, 1992. § Varnier et al., Am. J. Physiol. 269:E309-E315, 1995. # Van Hall, thesis, Maastricht University, 1996. |
|
Two interventions were recently reported that appeared to help increase glycogen resynthesis rates following exercise. Zawadzki et al. (1992) reported that addition of protein to the ingested CHO increased glycogen resynthesis rates (Table 15-1). Because higher plasma insulin concentrations were observed, it was suggested that the insulin effect on glycogen synthase was responsible for the observed effect. Varnier et al., (1995) reported that intravenous infusion of glutamine increased glycogen resynthesis rates (Table 15-2). However, glycogen resynthesis was measured at submaximal rates in the absence of ingested glucose. It was suggested that glutamine may directly stimulate glycogen synthase by activation of glycogen synthase phosphatase. The latter observation raised the possibility that the effect observed by Zawadzki was
mediated in part by glutamine, since plasma glutamine concentration increases following protein ingestion.
In another study, Van Hall (1996) and Wagenmakers et al., (1997) investigated whether the stimulatory effect of protein on glycogen resynthesis rates observed after CHO ingestion was mediated by insulin, glutamine, or both and whether co-ingestion of glutamine and glucose also stimulated glycogen resynthesis rates. Eight trained subjects were studied during 3 hours of recovery in four tests each consuming one of four drinks in random order. Drinks were divided into and ingested in three 500-ml portions, the first immediately after exercise and the others after 1 and 2 hours of recovery. Composition of the drinks is specified in Table 15-1. The wheat hydrolysate had a glutamine content of 26 percent and the whey hydrolysate had 6.6 percent. The plasma glutamine concentration fell from about 650 μmol at the end of exercise to 500 μmol after 2 hours in the control treatment. Both protein hydrolysates prevented that decrease (plasma concentration maintained between 600 and 700 μmol). The glutamine increased plasma glutamine to values between 1,000 and 1,200 μmol. Plasma insulin concentrations were twice as high with the protein hydrolysates as with glucose only and glucose plus glutamine. Glutamine alone did not increase glycogen resynthesis rates, while both protein hydrolysates did, though not significantly.
AUTHOR'S CONCLUSIONS AND RECOMMENDATIONS
-
No conclusive evidence supports net protein breakdown and increased amine acid oxidation during demanding endurance exercise in the laboratory and under field conditions. When energy balance is maintained, 100 g of protein should be more than enough to cover the protein requirements of subjects regularly involved in high workload exercise.
-
No evidence suggests that BCAA ingestion would optimize athletic performance during demanding endurance exercise. A potential risk exists that BCAA ingestion could lead to premature fatigue or loss of motor coordination under conditions where glycogen stores have been emptied (e.g., high workload with insufficient food intake).
-
Glutamine may be useful to support the immune system during high workload conditions, but solid evidence of the link between glutamine and the immune system and of the usefulness of glutamine supplementation has not yet been published.
-
Glycogen resynthesis immediately following demanding exercise can be accelerated by the addition of protein to the ingested CHO solutions. This is particularly relevant when subjects maintain a high daily energy expenditure (> 20-25MJ) for several days or weeks (e.g., Tour de France cyclists and those engaged in military operations). Failure to replenish the glycogen stores overnight will impede performance on the following day.
REFERENCES
Andersen, P., and B. Saltin. 1985. Maximal perfusion of skeletal muscle in man
. J. Physiol. 366:233-249.Ardawi, M.S.M., and E.A. Newsholme. 1983. Glutamine metabolism in lymphocytes of the rat. Biochem. J. 212:835-842.
Banister, E.W., and B.J.C. Cameron. 1990. Exercise-induced hyperammonemia: Peripheral and central effects. Int. J. Sports Med. 11:S129-S142.
Bergström, J., and E. Hültman. 1967. Muscle glycogen synthesis after exercise: An enhancing factor localized to the muscle cells in man. Nature 201:309-310.
Blomstrand, E., S. Andersson, P. Hassmén, B. Ekblom, and E.A. Newsholme. 1995. Effect of branched-chain amino acid and carbohydrate supplementation on the exercise induced change in plasma and muscle concentration of amino acids in human subjects. Acta Physiol. Scand. 153:87-96.
Blomstrand, E., P. Hassmén, B. Ekblom, and E.A. Newsholme. 1991. Administration of branched-chain amino acids during sustained exercise-effects on performance end on plasma concentration of some amino acids. Eur. J. Appl. Physiol. 63:83-88.
Brenner, I.K.M., P.N. Shek, and R.J. Shepherd. 1994. Infection in athletes. Sports Mad 17: 86-107.
Castell, L.M., J.R. Poortmans, and E.A. Newsholme. 1996. Does glutamine have a role in reducing infections in athletes? Eur. J. Appl. Physiol. 73:488-490.
Chaouloff, F., J.L. Elghozi, Y. Guezennec, and D. Laude. 1995. Effect of conditioned running on plasma, liver and brain tryptophan and on brain 5-hydroxytryptamine metabolism of the rat. Brit. J. Pharmacol. 86:33-41.
Chang, T.W., and A.L. Goldberg. 1978a. The origin of alanine produced in skeletal muscle. J. Biol. Chem. 253:3677-3684.
Chang, T.W., and A.L. Goldberg. 1978b. The metabolic fates of amino acids end the formation of glutamine in skeletal muscle. J. Biol. Chem. 253:3685-3695.
Darmaun, D., D. Matthews, and D. Bier. 1986. Glutamine and glutamate kinetics in humans. Am. J. Physiol. 251:E117-E126.
Elia, M., A. Schlatmann, A. Goren, and S. Austin. 1989. Amino acid metabolism in muscle and in the whole body of man before end after ingestion of a single mixed meal. Am. J. Clin. Nut. 49:1203-1210.
Essen, B., and L. Kaijser. 1978. Regulation of glycolysis in intermittent exercise in man . J. Physiol. 281:499-511.
Fitts, R.H. 1994. Cellular mechanisms of muscle fatigue. Physiol. Rev. 74:49-94.
Hargreaves, K.M., and W.M. Pardridge. 1988. Neutral amino acid transport at the human blood-brain barrier. J Biol. Chem. 263:19392-19397.
Hoffman-Goetz, L., and B.K. Pedersen. 1994. Exercise and the immune system. Immunol. Today 15:382-385.
Ivy, J.L., M.C. Lee, J.T. Brozinick, and M.J. Reed. 1988. Muscle glycogen storage after different amounts of carbohydrate ingestion. J. Appl. Physiol. 65:2018-2023.
Knott, P.J., and G. Curzon. 1972. Free tryptophan in plasma and brain tryptophan metabolism. Nature 239:452-453.
Krebs, H.A. 1975. The role of chemical equilibria in organ function. Adv. Enzyme Regulation 15:449-472.
Lacey, J.M., and D.W. Wilmore. 1990. Is glutamine a conditionally essential amino acid? Nutr. Rev. 48:297-309.
MacLean, D.A., T.E. Graham, and B. Saltin. 1996. Stimulation of muscle ammonia production during exercise following branched-chain amino acid supplementation in humans. J. Physiol. 493:909-922.
Madsen, K., D.A. MacLean, B. Kiens, and D. Christensen. 1996. Effects of glucose, glucose plus branched-chain amine acids or placebo on bike performance over 100 km. J. Appl. Physiol. 81:2644-2650.
Marliss, E.B., T.T. Aoki, T. Pozefsky, A.S. Most, and G.F. Cahill. 1971. Muscle and splanchnic glutamine and glutamate metabolism in postabsorptive and starved man. J. Clin. Invest. 50:814-817.
Matthews, D.E., and R.G. Campbell. 1992. The effect of dietary protein intake on glutamine and glutamate nitrogen metabolism in humans. Am. J. Clin. Nutr. 55:963-970.
Munro, H.N. 1964b. Historical introduction: The origin and growth of our present concept of protein metabolism. Pp. 1-29 in Mammalian Protein Metabolism, H.N. Munro and J.B. Allison, ed. New York: Academic Press.
Newsholme, E.A., I.N. Acworth, and E. Blomstrand. 1987. Amine acids, brain nenrotransmitters and a functional link between muscle and brain that is important in sustained exercise. Pp. 127-138 in Advances in Myochemistry, G. Benzi, ed. London: John Libby Eurotext.
Nurjhan, N., A. Bucci, G. Perriello, N. Stumvoll, G. Dailey, D.M. Bier, I. Toft, T.G. Jenssen, and J.E. Gerich. 1995. Glutamine: A major gluconeogenic precursor and vehicle for interorgan carbon transport in man. J. Clin. Invest. 95:272-277.
Pannemans, D.L.E., A.J.M. Wagenmakers, A.E. Jeukendrup, A.P. Gijsen, J.M.G. Senden, and W.H.M. Saris. 1997. The effect of prolonged moderate intensity exercise on protein metabolism of trained men. Med. Sci. Sport Ex. 29: S224.
Parry-Billings M., R. Budgett, Y. Koutedakis, E. Blomstrand, S. Brooks, C. Williams, P.C. Calder, S. Pilling, R. Baigrie, and E.A. Newsholme. 1992. Plasma amine acid concentrations in the overtraining syndrome: possible effects on the immune system. Med. Sci. Sports. Exerc. 24:1353-1358.
Parry-Billings M., J. Evans, P.C. Calder, and E.A. Newsholme. 1990. Does glutamine contribute to immunosuppression after major bums? Lancet 336:523-525.
Reed, M.J., J.T. Brozinick, M.C. Lee, and J.L. Ivy. 1989. Muscle glycogen storage postexercise: effect of mode of carbohydrate administration. J. Appl. Physiol. 66:720-726.
Ronnie, M.J. 1996. Influence of exercise on protein and amine acid metabolism. Pp. 995-1035 in Handbook of Physiology, Section 12, Exercise: Regulation and Integration of Multiple Systems, L.B. Rowell and J.T. Shepherd, eds. New York: Oxford University Press.
Rennie, M.J., P. Babij, P.M. Taylor, H.S. Hundal, P. MacLennan, P.W. Watt, M.M. Jepson, and D.J. Millward. 1986. Characteristics of a glutamine carrier in skeletal muscle have important consequences for nitrogen loss in injury, infection and chronic disease. Lancet 2:1008-1012.
Romijn, J.A., E.F. Coyle, L. Sidossis, A. Castaldelli, J.F. Horowitz, E. Endert, and R.R. Wolfe. 1993. Regulation of endogenous fat and carbohydrate metabolism in relation to exercise intensity. Am. J. Physiol. 265:E380-E391.
Sahlin, K., A. Katz, and S. Broberg. 1990. Tricarboxylic acid cycle intermediates in human muscle during prolonged exercise. Am. J. Physiol. 269:C834-C841.
Saris, W.H.M. 1997. Limits of human endurance: Lessons from the Tour de France. Pp. 451-462 in Physiology, Stress, and Malnutrition: Functional Correlates, Nutritional Intervention, J.M. Kinney and H.N. Tucker, eds. New York: Lippincott-Raven Publishers.
Segura, R., and J.L. Ventura. 1988. Effect of L-tryptophan supplementation on exercise performance. Int. J. Sports Med. 9:301-305.
Souba, W.W. 1991. Glutamine: a key substrate for the splanchnic bed. Ann. Rev. Nut. 11:285-308.
Stensrud, T., F. Ingjer, H. Holm, S.B. Strømme. 1992. L-tryptophan supplementation does not improve running performance. Int. J. Sports Med. 13:481-485.
Van Acker, B.A.C, K.W.E. Hulsewé, A.J.M. Wagenmakers, N.E.P. Deutz, B.K. Van Kreel, P.B. Soeters, and M.F. Von Meijenfeldt. 1996. Measurement of glutamine metabolism in gastrointestinal cancer patients. Clin. Nutr. 15(suppl. 1):1.
Van Hall, G. 1996. Amino acids, ammonia and exercise in man. Thesis, Maastricht University, The Netherlands.
Van Hall, G., D.A. MacLean, B. Saltin, and A.J.M. Wagenmakers. 1996. Mechanisms of activation of muscle branched-chain α-keto acid dehydrogenase during exercise in man. J. Physiol. 494:899-905.
Van Hall, G., J.S.H. Raaymakers, W.H.M. Saris, and A.J.M. Wagenmakers. 1995a. Ingestion of branched-chain amino acids and tryptophan during sustained exercise: Failure to affect performance. J. Physiol. 456:789-794.
Van Hall, G., B. Saltin, G.J. Van der Vusse, K. Söderlund, and A.J.M. Wagenmakers. 1995b. Deamination of amino acids as a source for ammaonia production in human skeletal muscle during prolonged exercise. J. Physiol. 489:251-261.
Varnier, M., G.P. Leese, J. Thompson, and M.J. Rennie. 1995. Stimulatory effect of glutamine on glycogen accumulation in human skeletal muscle. Am. J. Physiol. 269:E309-E315.
Varnier, M., P. Sarto, D. Martines, L. Lora, F. Carmignoto, G.P. Leese, and R. Naccarato. 1994. Effect of infusing branched-chain amino acids during incremental exercise with reduced. muscle glycogen content. Eur. J. Appl. Physiol. 69:26-31.
Vinnars, E., J. Bergström, and P. Fürst. 1975. Influence of the postoperative state on the intracellular free amino acids in human muscle tissue. Annals Surg. 182:665-671.
Wagenmakers, A.J.M. 1996. Muscle glycogen depletion and fatigue. The Physiologist 39:A68.
Wagenmakers, A.J.M., and P.B. Soeters. 1995. Metabolism of branched-chain amino acids. Pp. 67-83 in Amino Acid Metabolism and Therapy in Health and Nutritional Disease, L.A. Cynober, ed. New York, CRC Press, Inc.
Wagenmakers, A.J.M., E.J. Beckers, F. Brouns, H. Kuipers, P.B. Soeters, G.J. van der Vusse, and W.H.M. Saris. 1991. Carbohydrate supplementation, glycogen depletion, and amino acid metabolism during exercise. Am. J. Physiol. 260:E883-E890.
Wagenmakers, A.J.M., J.H. Coakley, and R.H.T. Edwards. 1990. Metabolism of branched-chain amino acids and ammonia during exercise: Clues from McArdle's disease. Int. J. Sports Med. 11:S101-S113.
Wagenmakers, A.J.M., H.J.M. Salden, and J.H. Veerkamp. 1985. The metabolic fate of branched-chain amino acids and 2-oxo acids in rat muscle homogenates and diaphragms. Int. J. Biochem. 17:957-965.
Wagenmakers, A.J.M., G. Van Hall, P.A.I. Van de Schoor, and W.H.M. Saris. 1997. Glutamine does not stimulate glycogen resynthesis in human skeletal muscle. Med. Sci. Sport Ex. 29:S280
Wagenmakers, A.J.M., G. Van Hall, and B. Saltin. 1996. Excessive muscle proteolysis during one leg exercise is exclusively attended by increased de novo synthesis of glutamine, not of alanine. Clin. Nutr. 15(suppl. 1):1.
Windmueller, H.G., and A.E. Spaeth. 1974. Uptake and metabolism of plasma glutamine by the small intestine. J. Biol. Chem. 249:5070-5079.
Zawadzki, K.M., B.B. Yaspelkis, and J.L. Ivy. 1992. Carbohydrate-protein complex increase the rate of muscle glycogen storage after exercise. J. Appl. Physiol. 72:1854-1859.
DISCUSSION
JOAN CONWAY: I have a simple question. Were these studies done only in men, or were they done in both men and women?
ANTON WAGENMAKERS: I am afraid that they were all done in top male athletes.
JOAN CONWAY: And would you think that gender would have any effect?
ANTON WAGENMAKERS: I don't have a reason to assume that there will be a difference. But the studies should be done in women, too, I think.
JOAN CONWAY: Thank you.
ROBERT NESHEIM: Doug?
DOUGLAS WILMORE: A very nice presentation. Thank you.
Joe Cannon and some of his associates did biopsies following eccentric exercise and found increased levels of interleukin-1 in muscle and later found rises in interleukin-1 in the bloodstream.
I think, in subsequent studies, they gave vitamin E to try to modify those responses. Do you have any thoughts about the use of antioxidants in athletes who have these heavy performance levels where muscle damage and inflammation may play a role?
ANTON WAGENMAKERS: When were these interleukin levels increased, immediately after or three or four days after?
DOUGLAS WILMORE: No, several hours after. And the bloodstream levels were elevated two to four hours after. Bob, is that right?
ROBERT WOLFE: I think it was at least that much later. It definitely was some time later.
ANTON WAGENMAKERS: So my impression is that there is certainly free radical damage in the muscle, but it probably is a couple of days later, when there are inflammatory cells to clear damaged fibres; certainly after eccentric
exercise, there will be inflammatory cells after three days in the muscle, and they can produce free radicals. That is for sure.
But I am not sure whether exercise per se leads to free radical damage. If that would be the case, then you would expect that vitamin E supplementation or antioxidant supplementation could improve performance. There is no definitive study yet, I don't think, that has shown an effect. Many studies have been done, but everybody has failed to find an effect on performance.
I think an area that is much more interesting is the central area, the gut-liver area. Demanding exercise, certainly in the heat, leads to a massive reduction of blood flow to the gut, and that can really imply the development of a severe underperfusion (gut ischemia). I really wonder whether, when athletes stop after a strenous event like the Hawaii Iron Man, reperfusion damage may occur. Quite a few of these athletes anecdotally have been reported to become quite ill within 15 minutes after they finish. I think the gut is a more interesting tissue to direct your antioxidants at than the muscle.
ROBERT NESHEIM: Dennis?
DENNIS BIER [off microphone]: Is it possible that the mechanism for increased glycogen storage after the addition of amino acid to the glucose may be repartitioning, in the sense that you now supply the nonessential amino acids that might have had to be synthesized from carbohydrate if you didn't supply them for muscle repair, so that in fact there is more glucose available for glycogen?
ANTON WAGENMAKERS: I think that it is a major assumption that muscle repair is needed. Over a 24-hour day, the Tour de France cyclists are in energy balance. After exercise, they cannot be in energy balance, because their total carbohydrate oxidation rate always exceeds their capacity for oral ingestion of carbohydrates.
But still, my impression is that as long as they are in reasonable energy balance, they don't break a lot of protein down during the exercise. They would not be able to perform at this high intensity for 22 days when they are in negative protein balance.
So I am not sure whether the mechanism you propose is the answer, because it really means, then, that repair and high protein synthesis rates are necessary following exercise. Certainly it is something we should look at, because these proteins, of course, may also have an effect on protein synthesis and degradation in the muscle in the first few hours after such demanding exercise.
ROBERT NESHEIM: Sree?
K. SREEKUMARAN NAIR [off microphone]: It is also possible that protein may be providing substrates for gluconeogenesis in the liver, and that the glucose thus formed is providing substrates for the glycogen synthase enzyme.
Do you have any data that protein alone also can increase glycogen synthesis after exercise?
ANTON WAGENMAKERS: Yes, there is. In the study done by Zawadzki, protein was given alone, and—you are right in that the glycogen resynthesis rate was a little bit higher than in basal conditions but much lower than with glucose plus protein. I don't think gluconeogenesis is extremely important in these studies, because the insulin concentration is so high that it probably will reduce gluconeogenesis. So I think it is more the higher insulin and probably the upgraded glycogen synthase activity that is causing that effect.
ROBERT NESHEIM: Michael Rennie and then Harris Lieberman.
MICHAEL RENNIE: We just got exactly the same result. Although we find this effect in the basal state, we don't get any difference when we add glucose. So I suspect that the whole effect is overridden by insulin. So do you think that the effects of the protein are insulin-dependent? Was the area under the insulin curve higher when you give glucose and protein together?
ANTON WAGENMAKERS: Yes, it is about double the effect. It probably is better to give the supplements every half hour because we got these big spikes because of the hourly doses. But at the high points, the insulin was twice as high.
MICHAEL RENNIE: So you could just give insulinogenic amine acids and see whether there is a beneficial effect.
ANTON WAGENMAKERS: Yes. We are doing that at the moment. We don't have the answers yet.
ROBERT NESHEIM: Harris?
HARRIS LIEBERMAN: Several years ago, there was a report by a Spanish group—Segura, I believe, was the principal investigator—which suggested that if you gave pharmacologic doses of tryptophan, you actually would improve endurance performance. Would you care to comment on that?
ANTON WAGENMAKERS: Yes. There has been a repeat of that study by a group from Norway (Stensrud et al., 1988) and they repeated that study very carefully, with exactly the same doses, in a very big group of subjects, about 30, I believe, and they did not see any effect of tryptophan at all on performance.
When you look at the Spanish data, some of the subjects were exhausted with a heart rate of 142, and as an exercise physiologist, I question such results.
ROBERT NESHEIM: Thank you very much. I think we need to move on.






















