
PANEL 3
Can we understand the processes of fossilization and non-biological processes sufficiently well to differentiate fossils from artifacts in an extraterrestrial rock sample?
Discussion
Summarized by Andrew Knoll, Panel Moderator
Recognition of a Biological Signature in Rocks
Dr. Knoll opened the session by summarizing the challenges of recognizing biological signatures in extremely old rock samples from Earth or Mars. Everyday experience suggests that the gulf between biology and the physical world is conspicuous. This impression arises, however, because the biology most familiar to us is principally that of organisms found on distal branches of the tree of life. The difficulty in distinguishing between biogenic and abiogenic features lies at the other end of the tree. Life arose as a self-perpetuating product of physical processes, and it is likely that the characteristics of Earth's earliest organisms—their size, shape, molecular composition, and catalytic properties—bore a close resemblance to the products of physical processes that gave rise to biology. For this reason, detecting the remnants of early life in terrestrial rocks is difficult. In martian or other extraterrestrial samples, it is doubly challenging. Given the evidence in hand from ALH84001 and the prospect of analyzing intelligently chosen samples from Mars within a decade, how do we fashion ground rules for recognizing the unambiguous signal of past (or present) biology?
Organisms have structure, they have a chemical composition, and they affect their environment; thus, paleontological evidence of ancient life can be morphological, geochemical, or sedimentological. Experience with terrestrial rocks makes it clear that features found in ancient samples can be accepted as biological only if they satisfy two criteria. They must be compatible with pattern generation by known biological processes. And they must be incompatible with formation by physical processes. This is straightforward in principle, but it requires that we understand the limits of pattern formation by biological and physical processes.
Dr. Knoll illustrated the challenge for this panel by drawing a Venn diagram in which biological and abiological patterns were depicted as distinct but overlapping fields. As yet, our understanding of the
limits on both biological and abiological pattern generation is incomplete, making it difficult to understand the dimensions of the “gray zone” of overlap. Recognition of biological pattern in extraterrestrial samples will require the identification of structures or molecules that reside in the biological field, but not in the zone of overlap. On the other hand, there is no assurance that terrestrial life exhausts the possibilities of biological pattern generation; therefore, knowing the limits of pattern formation by physical processes may provide the best yardstick for evaluating martian or other extraterrestrial samples. Dr. Benner suggested an alternative depiction of the Venn diagram in which the biological field is completely encompassed within the physical field—the point being that biological processes are a subset of a larger and more inclusive set of physical processes. As a depiction of process, this view is unimpeachable; nevertheless, the patterns generated by biological processes include structures and molecules not known to form under strictly physical conditions. Bones, radiolaria, and red algal thalli are examples of biologically diagnostic morphologies; cholesterol is a biologically diagnostic molecule.
Lessons from Earth
Panel 3 members agreed that our collective experience with Earth's geological record provides an important guide to fossil recognition and interpretation on Mars. Dr. Farmer demonstrated that processes of mineral precipitation can preserve biologically interpretable microfossils and sedimentary fabrics. This increasing knowledge of fossilization processes not only sheds light on the postmortem information loss that attends fossilization, but also focuses attention on martian environments most likely to preserve a biological record. Preservation of terrestrial remains is selective, with some organisms—and some parts of organisms—more likely to escape decay than others. During fossilization, cells can also shrivel or collapse, resulting in fossils that are much smaller than the organisms from which they are derived.
Dr. Schopf summarized experience in interpreting Earth's early fossil record, stressing the early phase of discovery, when reports of objects that proved to be abiological outnumbered those of genuine fossils. Not everything that is small and round is biological, and the rigorous criteria for biogenicity developed over the past three decades by paleontologists can be useful in the evaluation of extraterrestrial microstructures. Dr. Schopf emphasized the need to conduct interdisciplinary studies of petrology, micropaleontology, isotopic geochemistry, and molecular organic geochemistry, developing multiple lines of evidence for interpreting potentially biological patterns.
Although most of Panel 3's discussion focused on micron-scale structures, the distinctive macroscopic structures known as stromatolites were also considered. Stromatolites are laminated structures found in chemical sedimentary rocks, especially but not exclusively carbonate rocks. These structures, which can be flat-laminated, domal, columnar, or conical, are commonly interpreted as the products of sediment trapping, binding, and/or precipitation by microbial communities; however, it is apparent that comparable structures can be generated without the need for microbial templates. Indeed, such structures actually occur in the early geological record. This being the case, images of laminated precipitates that may be transmitted by a Mars rover cannot be construed as unambiguous evidence for biological activity (Farmer, Knoll). Micro- and mesoscale fabric studies on returned samples will be necessary to confirm or reject hypotheses of biological origin. Dr. Farmer suggested that specific microscopic textures may provide the biological fingerprint needed to be confident of stromatolite biogenicity in the terrestrial or martian rock record.
Dr. Bradley focused attention on abiologial pattern formation, suggesting that non-biological processes may be sufficient to explain a number of micro- and nanoscale features sometimes interpreted as biological, including those reported from Mars meteorite ALH84001. Dr. McKay vigorously disputed
some of these conclusions but agreed that much clearer criteria for biological pattern formation in extraterrestrial samples are needed.
Summary and Consensus
General consensus was reached on the following points:
- Terrestrial rocks contain an observable and interpretable record of biological evolution, but as we recede further back into time, that record becomes attenuated and difficult to interpret in detail. Martian samples may actually be better preserved than terrestrial sediments of comparable age, but lack both modem martian organisms for comparison and a more or less continuous fossil record that connects the present with early planetary history.
- A better understanding of biological signatures in sedimentary rocks is needed, and it is needed before intelligently collected martian samples are returned to Earth. These signatures certainly include fossil morphologies, but they must also include biomarker molecules, isotopic fractionation, and biological mineralization and trace element concentrations. In all cases, improved understanding of biological pattern formation must proceed in tandem with better knowledge of the generative capacity of physical processes.
- There is both a need and an opportunity to more effectively integrate laboratory and field observations of fossilization processes with investigations of Earth's early sedimentary record. Multidisciplinary investigations are required in exopaleontological research, and there is a need for new technologies that will enhance our ability to obtain chemical information from individual micro-structures.
Fossils and Pseudofossils: Lessons from the Hunt for Early Life on Earth
J. William Schopf
Department of Earth and Space Sciences, Molecular Biology Institute, IGPP Center for the Study of Evolution and the Origin of Life University of California at Los Angeles
Abstract
As shown by other papers in this workshop, the actual and theoretical size limits of minute living microorganisms are incompletely defined and, so, the biogenicity of exceedingly small "biologic-like" objects is subject to debate. But the problem of distinguishing between minute biologic and nonbiologic objects is more vexing if they are present in rocks and therefore open to interpretation either as microbial fossils or nonbiologic pseudofossils, and it is a problem confronted even when interpreting objects with the dimensions of "normal-sized" microorganisms. The problem becomes more complicated still as the size and complexity of the objects considered decrease, or as the provenance of such objects becomes increasing distant from the present either in time (as for Archean in contrast with Proterozoic fossil-like objects) or space (as for structures in exterrestrial rather than terrestrial materials). Lessons learned from the past three decades of search for ancient records of life on Earth point a way toward solution of this problem: (1) The search for evidence of early life should be multidisciplinary (or, better, interdisciplinary); (2) the evidence should be positive, affirming a biologic origin, rather than neutral (consistent either with biology or nonbiology) or negative (evidence inferred by default to be biogenic); and (3) the provenance, age, indigenousness, syngenicity, and biogenicity of such evidence must be established beyond question.
You must not. fool yourself—and you are the easiest person [for you] to fool.
—Richard Feynman, CalTech
Commencement Address, 1974
Rules for the Hunt
There are fine lines between what is known, guessed, and hoped for, and because science is done by real people these lines are sometimes crossed. But science is not a guessing game. The goal is to know. "Possibly . . . perhaps . . . maybe" are not firm answers and feel-good solutions do not count. With regard to the famed Mars meteorite (McKay et al., 1996), for example, life either once existed on Mars or it didn't. Meteorite ALH84001 either holds telling evidence or it doesn't. Eventually, hard facts will sort it out. But the still-simmering controversy might have been avoided had three lessons now learned from the episode been heeded from the outset:
First, the search for evidence of past life, whether in Earth rocks or extraterrestrial samples, must be multidisciplinary—at a minimum, based on the techniques and findings of biologists, paleobiologists, geologists, and geochemists—or, better still, interdisciplinary, carded out by researchers schooled in both the life and physical sciences.
Second, the evidence sought should be positive—evidence that affirms the biologic origin of the features detected. Evidence that is neutral (consistent either with biology or nonbiology) is by its nature
inadequate to establish the existence of past life, and interpretations based on negative reasoning—inference by default, such as a claim that because a feature is not obviously mineralic it "must" be biogenic—are likely to prove erroneous.
Third, to be acceptable, evidence of past life must meet five specific tests (Schopf and Walter, 1983; Sehopf, 1993):
|
1. |
Provenance—Is the source firmly established of the rock sample that contains the putative biologic features? (For a terrestrial sample, stratigraphic and geographic provenance should be known precisely, as demonstrated, for example, by replicate sampling by different workers.) |
|
2. |
Age—Is the age of the rock sample known with appropriate precision? (For example, is the age tightly constrained by multiple measurements and lines of evidence?) |
|
3. |
Indigenousness—Are the putative biologic features indigenous to the rock? (For example, are the features embedded in the rock matrix rather than being surficial contaminants, a test that can be met for minerals and fossils by studies of petrographic thin sections and for biologic isotopic signatures by ion microprobe analyses in situ.) |
|
4. |
Syngenicity—Are the features syngenetic with a primary mineral phase of the rock? (Or are they of later origin, for example introduced into pores or fractures and lithified by secondary or later-generation minerals—a test that for minerals, fossils, and isotopic evidence can usually be met by studies of petrographic thin sections.) |
|
5. |
Biogenicity—Are the features assuredly biological? (Of the five tests, establishment of biogenicity has often proven to be the most difficult, as discussed below.) |
The Search for Ancient Fossils
Four independent but potentially mutually reinforcing lines of evidence have been used to trace the earliest (microbial) records of life on Earth: minerals, organic compounds, isotopic signatures, and fossils. Of these, fossils are potentially the least ambiguous—independent of the other lines of evidence, the presence of unquestionable fossils indicates the existence of past life. On Earth, two kinds of fossils are present in the early rock record: stromatolites (a type of "trace fossil") and cellularly preserved microbes ("body fossils").
Stromatolites
Formally defined, a stromatolite is an accretionary organosedimentary structure, commonly thinly layered, megascopic, and calcareous, produced by the activities of mat-building communities of mucilage-secreting microorganisms, mainly filamentous photoautotrophic prokaryotes such as cyanobacteria.
Rather than highlight the link between living and fossil examples, as this definition does, some workers prefer to restrict "stromatolite" to geologic specimens and use "microbial mat" for their living counterparts. Others call structures stromatolites only if they have fairly high relief above the neighboring substrate and refer to the fiat-lying ones as mats or sheets. Common to all appropriate definitions is the concept that it is the biologic origin of the layering in stromatolites that makes them distinctive. (For this reason, the practice adopted by some of dubbing almost any thinly layered calcareous rock “stromatolitic" should be avoided, because it confuses true stromatolites with non-biologically deposited look-alikes—cave rocks, such as stalactites and stalagmites, and some hot spring deposits, for instance, formed where minerals build up in thin, sometimes wavy layers as they crystallize from solution.
Fortunately, there are not many kinds of purely inorganically deposited rocks that have stromatolite-like layering, and these usually are fairly easy to tell apart from true stromatolites because they form when minerals repeatedly crystallize out of solution to make stacked surface-coating layers that are almost always more uniform and regular than those laid down by life. And, of course, the purely nonbiologic structures never harbor fossil remnants of mat-building microbial communities.)
As noted in the definition, most stromatolites are calcareous. For this reason, most do not contain structurally identifiable remnants of the microbes that built them (the organic cells having been crushed between growing carbonate grains as the layered sedimentary mass solidified to rock). Hence, like a track, trail, or burrow preserved in an ancient sediment, stromatolites are classed as "trace fossils," organosedimentary structures that evidence biologic activity yet are themselves not fossilized organisms.
On Earth, microbes are so widespread that there is practically no place where stromatolitic look-alikes form without life playing at least a minor role. But on a planet where life never got started there could be many places veneered by thinly layered stromatolite-like deposits unrelated to life—laid down, for instance, by repeated wetting and drying, or freezing and thawing, of mineral-charged salt pans or shallow lagoons. Moreover, it is useful to recall that though fossil stromatolites were recognized as distinctive sedimentary structures early in the 1800s, they were regarded by many geologists as concretionary bodies of purely inorganic origin and it was not until the 1960s (when living specimens were first discovered and fossils of mat-building microbes were found in ancient examples) that their microbial origin was firmly established. Similarly, were stromatolite-like structures detected in an extraterrestrial sample (or photographed on the surface of another planet), it seems certain there would be widespread question as to whether they were produced by life. This uncertainty would be dispelled only if hard and fast rules (based, for example, on diagnostic sedimentological properties) were in place to unambiguously distinguish true stromatolites from nonbiologic mimics, or if the structures were shown to contain cellular remnants of the organisms that built them (in which case, the telling evidence would be the cellular fossils, not the stromatolites themselves).
Cellular Fossils
In studies of ancient life on Earth, optical microscopic studies of two types of preparations have been used to detect remnants of early cellular microbes: acid macerations and petrographic thin sections. Transmission and scanning electron microscopy have been used to characterize microfossils previously detected in macerations or thin sections, but have not proven to be reliable detection techniques.
Optical Microscopy of Macerations. Maceration, the easier and faster of the two techniques used to detect ancient microfossils, is carried out by dissolving a rock in mineral acid (hydrochloric acid for limestones, hydrofluoric for cherts and siltstones). Because of their coaly composition, organic-walled microfossils pass through the technique unscathed. Abundant fossils are concentrated in the resulting sludgelike acid-resistant residue, which can be slurried onto a microscope slide for study.
Unfortunately, this technique is subject to error-causing contamination. Contaminants can be introduced at almost every stage of the process. At the beginning, even though rock surfaces are cleaned to remove adhering soil, microbes in minute rock crevices are likely to be missed. Laboratory water and commercially available mineral acids used in the technique can also contain a zoo of living contaminants-bacteria, cyanobacteria, unicellular algae, microscopic fungi. And an almost limitless array of fossil-like objects can be introduced during transfer of the residue onto microscope slides. Common culprits include dust, cigarette ash, spores, and pollen grains that settle from the air; lint fibers from clothing or the cloth used to clean microscope slides; small woody fragments and chunks of resin
abraded off the wooden rods used in some laboratories to stir the acid-rock sludge; flakes of dandruff and strands of hair; even bits of small spiders that live in water pipes. From the 1950s into the 1970s, as studies of the Precambrian fossil record were getting under way in earnest, all of these maceration-borne contaminants were misinterpreted as fossils by one worker or another (Schopf and Walter, 1983; Mendelson and Schopf, 1992).
Optical Microscopy of Petrographic Thin Sections. In petrographic thin sections, the other type of preparation used for optical microscopic detection of ancient microfossils, fossils are detected within the rock, so indigenousness can be demonstrated and the possibility of laboratory contamination ruled out. Consider, for example, microfossils in cherts (together with fine-grained clastic sediments such as siltstones, one of the most fossiliferous rock types known in the early geologic record). Fossil-bearing cherts are made up of cryptocrystalline interlocking grains of quartz laid down from solution. The precipitated grains initially pass through a gem-like opaline state, taking thousands of years to solidify into a full-fledged chert, and the microorganisms are petrified (technically, "permineralized"), embedded within a solid chunk of rock. The quartz grains, which are deposited inside the cells and surround them on all sides, develop so slowly that they grow through the cell walls instead of crushing them. As a result, the petrified fossils are preserved in three-dimensional unflattened bodies that except for their quartz-filled interiors and the brownish color of the aged organic matter that makes them up bear a striking resemblance to living microorganisms. In such sections, only those objects that are entirely entombed in rock can be considered fossil, so it is easy to exclude contaminants that settle onto the surface of a section or are embedded in the resin used to cement the sliver of rock onto the glass thin-section mount.
Optical microscopy of thin sections also provides a way to establish that fossil-like objects date from the time a rock formed rather than having been sealed later in cracks and crevices—that is, to establish that the objects are syngenetic with a primary mineral phase rather than one of secondary or later genesis. Consider, for example, microfossiliferous stromatolitic cherts. When such cherts first form, many contain cavities where gases (often oxygen, carbon dioxide, hydrogen, or methane) given off by the microbial community accumulate in small pockets. Later these cavities can be sealed, filled by a second generation of quartz laid down from seeping groundwater, sometimes tens or hundreds of millions of years after the first chert formed. Microscopic organisms trapped in these cavities and petrified by the second-generation quartz would be true fossils but would be much younger than the rock unit itself. Fortunately, the various generations of quartz in a chert can be sorted out. Rather than having interlocking grains, quartz that fills cavities is a type known as chalcedony that follows the smooth contours of the infilled pocket to form distinctive botryoidal masses. Secondary quartz in cracks or veinlets is also easy to identify because it is angular and its grains much larger than those first formed.
Because special equipment is needed to prepare thin sections—and their study is exceedingly timeconsuming—some workers have focused their hunt for ancient fossils on acid-resistant rock residues. In relatively young (Proterozoic) Precambrian rocks, where the fossil record is well enough known that misidentification of contaminants and fossil-like artifacts can be avoided, this technique is useful, simple, and fast. But to avoid mistakes in the oldest (Archean) Precambrian, where the fossil record is not nearly so well known—and, of course, in any extraterrestrial sample—use of the more rigorous thin section technique is essential.
Electron Microscopy as a Detection Technique. Though both transmission electron microscopy (TEM) and scanning electron microscopy (SEM) have been used to characterize the cellular makeup and morphology of Precambrian microfossils, neither has proven to be a reliable detection technique.
The TEM studies have involved examinations either of organic-walled fossils freed from their rock matrix by acid maceration, embedded in epoxy resin, and sectioned using a diamond knife, or of fossil-like objects detected in plastic ("formvar") surface replicas of polished and etched petrographic thin sections. Transmission electron microscopy of macerated microfossils has been used to elucidate the structure of cell walls, membranes, and internal organic contents (Schopf and Oehler, 1976). But because macerations are susceptible to contamination, such studies are useful only of fossils previously detected in petrographic thin sections. Similarly, contamination by nonindigenous particles presents a problem for studies of surface replicas, as does the introduction of nonbiogenic artifacts of a variety of types (blisters, bubbles, strands of formvar, and so forth) that in the 1960s and early 1970s were repeatedly identified mistakenly as "ancient fossils" (Schopf and Walter, 1983).
Preparation of samples for study by scanning electron microscopy is simpler than for transmission electron microscopy, and the results obtained are generally easier to interpret. But, as in TEM studies, establishment of the indigenousness of the objects detected and their syngenicity with a primary mineral phase are not straightforward. And, also as in TEM studies, fossil-like artifacts have been misinterpreted by SEM, especially in rock samples where mineralic morphology has been altered and smoothed to "biologic-like" shapes by acid-etching (Schopf and Walter, 1983).
In sum, neither TEM nor SEM has proven reliable for detection of microfossils in the Precambrian rock record on Earth. In view of this track record, claims of detection of minute fossils in extraterrestrial samples by use of electron microscopy should be regarded with skepticism.
Biogenicity. Though optical studies of petrographic thin sections can overcome the problems of establishing indigenousness and syngenicity, the problem of biogenicity often remains. Here, too, lessons learned from studies from the search for Precambrian microbes apply, where in past decades Precambrian microstructures "unlike known mineral forms" have been claimed to be "fossils" simply for want of any other explanation. Living contaminants, "lifelike" dust particles, ball-shaped mineral grains, clumps and shreds of compressed coaly organic matter, solid opaque globules, and a variety of other objects have all been claimed as Precambrian fossils, often on the basis of only one or a few specimens and despite the absence of identifiable cells or other telltale features of living systems (Schopf and Walter, 1983; Mendelson and Schopf, 1992). Many of these reports are founded on the notion that because an object doesn't look mineral it "must” be fossil. This negative reasoning, inference by default, is insufficient. Claims of evidence of past life—whether in Earth rocks or in extraterrestrial samples—need to be backed by positive evidence, hard facts showing what an object actually is rather than what it seemingly is not.
Furthermore, the positive evidence adduced must be strong enough to rule out plausible nonbiologic sources. For example, because organic matter can be produced in nonbiologic ways (as when life originated or today in interstellar space), the mere presence of coaly particles in an ancient sediment is not enough to prove that life existed. And because unicell-like organic spheroids can form without life (from clumping of organic matter in seawater or by coaly matter coating ball-shaped mineral grains), tiny round organic bodies in a rock cannot be regarded as assured fossil cells. Nit-picking care of this sort is no longer necessary for reports of fossils from the younger (Proterozoic) part of the Precambrian where evidence of life is overwhelming. But in the older (Archean) part, where so little yet is known—and, even more so, in any extraterrestrial sample—demanding rules must still apply.
Probably the best way to avoid being fooled by nonbiologic structures is to accept as bona fide fossils only those of fairly complex form. This may seem an unreasonably stringent rule for truly ancient fossils since the earliest kinds of cellular life (here and presumably elsewhere) almost certainly were very simple—probably individual, tiny, spheroidal cells. But until we have a sounder base of
knowledge and better rules to separate nonfossils from true, it is best to err on the side of caution. For the present, even in ancient terrestrial samples it is safest to accept as biologic only fossils that have unquestionably biologic form, for example colonies of ball-shaped cells embedded in a surrounding organic envelope and thread-like filaments made up of chains of many cells. As evidence builds, we will gain confidence to better interpret less certain finds.
Lessons Learned
On the basis of what has been learned from the search for evidence of Precambrian life on Earth, the biologic origin of putative ancient terrestrial fossils can be accepted if they are (1) made up of coaly organic matter (or are shown to be mineral-replaced); (2) complex enough in cellular structure to rule out plausible nonbiologic origins; (3) represented by numerous specimens (if one specimen is preserved, others should be too); and are (4) members of a multicomponent assemblage (terrestrial ecosystems are never monospecific) that (5) exhibit a range of taphonomic degradation consistent with their mode of preservation. In accordance with younger fossils and living microbes, the objects also should be shown to (6) exhibit (gene-based) morphological variability; (7) have inhabited a plausibly livable environment; (8) have grown and reproduced by biologic means of cell division; and (9) exhibit a biogenic isotopic signature.
These nine criteria for biogenicity, and the four other tests that must be met of bona fide terrestrial ancient fossils (provenance, age, indigenousness, and syngenicity) are the product of 30 years of trial, error, and ultimate success. If we seek to avoid Feynman's pitfall of "fooling ourselves," at a minimum we must apply criteria at least as rigorous in the search for evidence of extraterrestrial life, regardless of how minute the putative "microbe-like" objects may be.
References
McKay, D.S., E.K. Gibson, K.L. Thomas-Keprta, H. Vali, C. Romanek, S.J. Clemett, X.D.F. Chiller, C.R. Maechling, and R.N. Zare. 1996. Search for past life on Mars: Possible relic biogenic activity in Martian meteorite ALH84001. Science 273:924-930.
Mendelson, C.V., and J.W. Schopf. 1992. Proterozoic and selected Early Cambrian microfossils and microfossil-like objects. Pp. 865-951 in The Proterozoic Biosphere, A Multidisciplinary Study, J.W. Schopf and C.Klein (eds.). New York: Cambridge University Press.
Schopf, J.W. 1993. Microfossils of the Early Archean Apex chert: New evidence of the antiquity of life. Science 260:640-646.
Schopf, J.W., and D.Z. Oehler. 1976. How old are the eukaryotes? Science 193:47-49.
Schopf, J.W., and M.R. Walter. 1983. Archean microfossils: New evidence of ancient microbes. Pp. 214-239 in Earth's Earliest Biosphere, Its Origin and Evolution , J.W. Schopf (ed.). Princeton, New Jersey: Princeton University Press.
Taphonomic Modes in Microbial Fossilization
Jack Farmer
Department of Geology Arizona State University
Introduction
The microbial fossil record encompasses a wide variety of information (Figure 1) including morphological fossils (e.g., preserved cellular remains, microfabrics, and stromatolites) and chemofossils (e.g., organic biomarker compounds, isotopic and other geochemical signatures, including biominerals). Observations range in scale from mesoscopic biosedimentary fabrics, to microscopic cellular structures, to sub-microscopic chemical signatures. At all scales of observation, problems often arise when trying to distinguish between biological and inorganic features in the ancient rock record. Stromatolites, defined as laminated biosedimentary fabrics formed by the trapping and binding of sediments and/or precipitation of minerals by microorganisms (Walter 1977), are sometimes impossible to distinguish from finely laminated sediments formed by inorganic processes (see Grotzinger and Rothman 1996). At the cellular level, the biogenicity of Precambrian microfossils has been debated and criteria suggested for identifying pseudofossils (Schopf and Walter 1983; Buick 1984; Awramik et al. 1988). Distinguishing biominerals from their inorganic counterparts has also proven quite difficult in fossil materials, and the biological interpretation of isotopic and organic chemical evidence can also be inconclusive, especially for rocks that have undergone significant diagenesis. The challenge of establishing biogenicity in ancient materials is illustrated by recent debates over the origin of features in Martian meteorite ALH 84001 (McKay et al. 1996; see also review by Treiman 1998). The following discussion is aimed at identifying approaches that may enhance our ability to recognize biosignatures in ancient rocks.
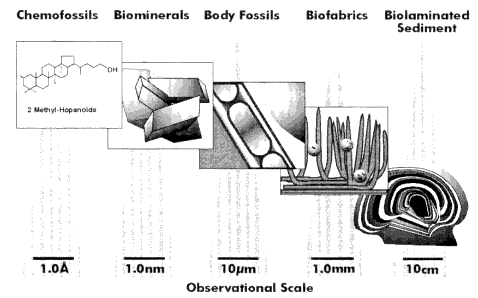
Figure 1.
Types of microbial fossil information arranged from right to left in order of decreasing spatial scale.
Importance of Spatially Integrated Studies for Assessing Biogenicity
The most important legacy of ongoing studies of the ALH 84001 meteorite may ultimately prove to be the value of the approach used, namely the integration of evidence over a broad range of observational scales. In developing integrated approaches to the study of biogenicity in ancient materials (whether of terrestrial or extraterrestrial origin), the challenge to the paleontologist is basically threefold. First is to place field samples within a well-defined geological (stratigraphic/age and paleoenvironmental) context. Although the materials may themselves contribute to such knowledge, detailed studies generally demand an understanding of both regional and local geology. Second is the detailed mesoscopic and microscopic characterization of individual samples. Establishing a detailed framework of microscopic observations is requisite for subsequent microsampling of a rock for geochemistry. This involves the identification of mineralogical and microtextural frameworks to distinguish between primary and secondary (diagenetic) features and to establish sequences of mineral paragenesis. At this step it is important to avoid sample contamination or the introduction of structural artifacts during sample preparation that can lead to misinterpretations. The third step involves the application of microsampling methods to place geochemical observations within the microscale spatial and temporal frameworks defined in Step 2. With the development of more spatially refined sampling methods it is now possible to chemically interrogate single mineral phases within rocks. This enables a much more refined understanding of the microenvironmental factors that have affected the preservation of fossil biosignatures.
Even using the spatially integrated approach outlined above, biogenic hypotheses often remain untested. The debate over ALH84001 illustrates this point. Hypothesis testing needs to be more than just confirmatory in nature. Because inorganic processes can so easily confuse biological interpretations in ancient materials, every opportunity should be taken to disprove life-based hypotheses. Because we are working with complex historical systems, side-by-side comparisons of modem biological and inorganic analogs may be required to adequately formulate and test hypotheses.
Taphonomic Bias and Common Modes of Preservation
Taphonomy is that subdiscipline of paleontology that deals with the transition of fossil remains from the biosphere to the lithosphere (Efremov 1940; Wilson 1988). In his review of taphonomy, Muller (1979) included a consideration of the cause of death, the processes of decomposition (necrolysis), postmortem transport or other events leading up to burial (biostratinomy) and post-burial events, inclusive of chemical and mechanical changes that occur within the sediment (fossil diagenesis). Taphonomic concepts have been developed largely with reference to multicellular organisms, but the same basic principles apply to the microbial fossil record (e.g., Bartley 1996; Knoll 1985; Knoll and Golubic 1979; Oehler 1976). In the present context, an understanding of common taphonomic modes in microbial fossilization has value in providing additional criteria for assessing biogenicity.
Preservational Modes in Precambrian Marine and Lacustrine Environments
In Precambrian paleontology, studies of organically preserved cellular remains have provided the most complete paleobiological picture of early microbial life. However, the Precambrian record reveals a strong preservational bias based on lithology, paleoenvironment, and the structure of organic materials (Knoll 1985). Perhaps the most instructive Precambrian fossil microbiotas are those preserved as three-dimensional forms in silica or phosphate. Figure 2A provides an example of preservation in silica from the 2.15 Ga. Belcher Island Group of Canada (Hofmann 1976; photomicrograph courtesy of Hans

Figure 2.
Common taphonomic modes. (A) Thin section photomicrograph of fossilized cyanobacterial mats dominated by a colonial coccoid species of Eoentophysalis. Sample from a silicified limestone of the Belcher Island Group, Canada. Early silicification of some laminae enhanced organic matter preservation (compare 1 and 2 on image). Photo courtesy of Dr. Hans J. Hofman. (B) Filament molds of Phormidium (filamentous cyanobacteria) formed by encrustation of trichomes. Sample from mid-temperature facies, Excelsior Geyser Basin, Yellowstone National Park. (C) Preserved sheaths of Calothrix (filamentous cyanobacteria), within a pisolith, from the Beach Geyser Group, Yellowstone National Park. (D) Critical point dried sample of a Calothrix mat showing clusters of silica spheres intimately interspersed within the dried filamentous remnants of an exopolymer matrix (see arrow in C).
Hofmann). In this case, silica was infused into thin mats of Eoentophysalis, a colonial coccoid cyanobacterium. In this case, the early silica permineralization of extracellular capsules preserved the external form of individual cells, as well as important aspects of the mat architecture (type of mat growth, stratification of organisms, etc.). Most Precambrian examples of this type were deposited in peritidal marine shorelines of elevated salinity (Knoll 1985). The second major lithotype is fine-grained, clay-rich detrital sediment (shale and volcanic ash) where microbiotas are preserved as two-dimensional compressions flattened during the compaction of sediments. These sediments usually represent deeper basinal settings where anaerobic conditions prevailed. Although several studies have demonstrated that rates of aerobic and anaerobic decay of organic matter do not differ significantly over a broad range of environments (see Lee 1992 and references therein), rates of early diagenetic mineralization have been shown to be higher in anaerobic environments. Such early mineralization is perhaps the most important singular factor in promoting organic matter preservation (Allison 1988; Allison and Briggs 1991).
Taphonomic Trends in Siliceous Thermal Springs
As illustrated by the Belcher Island example, the microbial fossil record is strongly biased toward organisms that possess degradation-resistant cell walls and extracellular envelopes (sheaths and capsules). Another example that illustrates this point is drawn from taphonomic studies of modem and ancient siliceous spring deposits (Farmer and Des Marais 1994; Farmer et al. 1995; Jones and Renaut 1997; Jones et al. 1997; Cady and Farmer 1996; Walter et al. 1998). In rapidly mineralizing thermal springs, the dominant mode of preservation is encrustation of biological surfaces by precipitating minerals, followed by the rapid degradation of organic materials to produce external molds of cells and filaments (Figure 2B). Occasionally, within the lowest temperature facies of thermal spring deposits, partially degraded and permineralized trichomes are observed. But generally speaking, only the sheaths of cyanobacteria (which resist degradation) are preserved in sub-recent fossil materials (Figure 2C).
Comparative taphonomic studies of modem and ancient thermal spring deposits suggest that preservation is strongly skewed toward higher order biofabrics (stromatolites and biologically mediated microtextures) with cellular preservation being limited to lower temperature facies. The fidelity of cellular level preservation appears to increase with decreasing temperature, in large part owing to systematic biological changes that occur along thermal gradients. At temperatures below ˜59°C (the upper temperature limit for thermophilic species of Phormidium, a filamentous cyanobacterium), mats typically increase in thickness, exhibiting a wider variety of surface textures and internal mat fabrics. Below ˜35°C, mats similarly show a wide variety of mat structures preserved as stromatolitic fabrics. But in addition, over these temperatures, filamentous cyanobacteria (e.g., Calothrix) exhibit substantially larger cell diameters and much thicker sheaths, a factor that enhances cellular level preservation.
Effects of Silica Diagenesis
During the early diagenesis of siliceous spring deposits, there is a pervasive structural reorganization of primary sedimentary fabrics owing to the recrystallization of metastable silica polymorphs (Opal A) to quartz. At the cellular level, only the filamentous and coccoid species having cell diameters larger than ˜2 mm and that possess thick extracellular sheaths or capsules, survive diagenetic recrystallization. Thus, within siliceous spring deposits at all scales, the morphological fossil record is heavily biased toward the filamentous cyanobacteria (photoautotrophs) and, in particular, larger species that occur below temperatures of ˜59°C in modem springs. The smaller coccoid and filamentous Bacteria and
Archaea (cell diameters usually <1 µm) that dominate at temperatures >73°C are also rapidly encrusted by silica. But, in the absence of thick cell walls or extracellular sheaths, entombed organic matter is rapidly decayed away, leaving behind only external molds that are rapidly infilled with opaline silica. During recrystallization there is a tendency for grain size to increase. Fine crystallites of amorphous Opal A (<5 µm; see Figure 2D) are first replaced by microquartz (5-20 µm), which are in turn replaced by megaquartz (20 to >200 µm; see Hesse 1990). This results in the loss of most primary microstructure (see Walter et al. 1998). Because of their small size, the 1-2 µm cell molds of smaller Bacteria and Archaea are quickly obliterated during recrystallization. Acid etching of ancient chert samples sometimes reveals a variety of simple spherical and rod-shaped forms within the matrix of the rock. However, at the microscale, inorganic precipitates of silica exhibit shapes very similar to simple cells, and cell-like forms may also be formed inorganically during the acid etching process. In the absence of cross sectional views that show evidence of a cellular structure, such morphological features are not compelling evidence for biogenicity.
Microbial Biomineralization
An understanding of the varied roles that microorganisms play in mediating mineralization processes is of fundamental importance in understanding fossilization, the origin of biosedimentary fabrics, and the processes of early diagenesis. There are a potentially wide variety of processes that are of interest in this context (see Ehrlich 1996), but for convenience they may be grouped into active processes in which mineral precipitation is directly driven by the metabolic functions of an organism and passive processes in which precipitation is influenced by the structure (e.g., cation-binding) properties of cellular or extracellular materials. In the following discussion, emphasis is placed on passive processes and the importance of extracellular materials in mineralization.
Metal-binding Capacities of Microbial Cell Walls and Extracellular Exopolymers
Natural microbial populations usually secrete large amounts of extracellular exopolymer (EPS), ranging from tightly structured capsules and sheaths around cells to a more loosely bound slime that forms the matrix of microbial biofilms and mats. Decho (1990) identified a wide variety of functions for EPS, including (1) buffering the microenvironment around cells against changes in pH, salinity, and the harmful effects of toxins (e.g., heavy metals); (2) protection against the harmful effects of UV radiation and desiccation during exposure; (3) protection against digestion by grazers; (4) adhesion of biofilms to surfaces; and (5) the concentration of exoenzymes and nutrients required for growth (including dissolved carbon compounds).
Microorganisms adsorb and concentrate many metallic cations required for growth through electrostatic interactions with anionic carboxyl and phosphoryl groups in the cell wall. In addition, however, the exopolymers that surround cells are also very reactive and can readily bind metals such as iron (Konhauser et al. 1993, 1994). EPS is a highly hydrated material (˜99% water) possessing an extremely porous fibrillar structure that renders it highly adsorptive. The polysaccharides of EPS possess abundant anionic carboxyl and hydroxyl groups that provide potential binding sites for metals. Of special importance are the carboxyl groups of uronic acids (carboxylated polysaccharides) that correlate strongly with the metal binding capacity of EPS (Kaplan et al. 1987). EPS can bind a wide variety of metals, including Pb, Sr, Zn, Cd, Co, Cu, Mn, Mg, Fe, Ag, and Ni (Decho 1990 and references therein). The metal binding capacity of EPS is strongly influenced by pH, being highest around pH 8 (average seawater). Preliminary studies of mineralization in siliceous thermal springs suggest that much of the
silica nucleates within the EPS matrix of mats over a pH range of 8 to 9 (Farmer et al. 1997; see also Figure 2D). Ferris et al. (1989) reported that near neutral pH, microbial biofilms concentrated metals up to 12 orders of magnitude higher than observed under acidic conditions. Neutral to alkaline micro-environments are commonly produced within microbial mats and biofilms through such processes as photosynthesis and sulfate reduction (Krumbein 1979).
Biomineralization as a Factor in Microbial Biosedimentology and Fossilization
Given the ability of EPS to concentrate various metals, it is not surprising that bacteria have been implicated in a wide variety of biomineralization processes. As a result of cellular metabolism, microorganisms alter the chemical microenvironment around the cell, modulating the pH, as well as the concentration of a variety of organic and inorganic solutes. This can induce the large-scale precipitation of authigenic minerals in natural environments. Some examples follow.
Thompson and Ferris (1990) attributed seasonal “whitings" in Green Lake, New York, to the precipitation of calcium carbonate, gypsum, and magnesite by the coccoid cyanobacterium, Synechococcus. During photosynthesis, the pH microenvironment around individual cells becomes more alkaline owing to the extraction of CO2 . This can result in supersaturation with respect to the previously mentioned phases. Using TEM, Thompson and Ferris (1990) showed that these minerals actually nucleate on the S-layers (extracellular envelopes) surrounding cells (see Beveridge and Graham 1991 for a discussion of S-layers in bacteria). In Green Lake, Synechococcus exhibits a unique double S-layer arrangement. As the outer layer becomes fouled with minerals, it is shed and sinks to the bottom, accumulating on the lake floor as a fine-grained carbonate deposit (micrite). During Synechococcus blooms, whole cells can become encrusted by this process and incorporated into the sedimentary record.
Konhauser et al. (1994) found that the bacteria comprising epilithic biofilms of riverine environments sequestered significant amounts of iron, along with smaller amounts of Ca, K, Si, Al, and Mn. Mineralization ranged from Fe-rich EPS capsules to more extensive fine-grained mineral precipitates. Authigenic minerals ranged from complex amorphous (Fe, Al) silicates of variable composition to more silica-rich ordered phases intermediate between glauconite and kaolinite.
Zirenberg and Schiffman (1990) reported the encrustation and replacement of bacterial filaments by metal sulfide minerals and silica in deep sea hydrothermal vent environments. They suggested that bacterially mediated processes may contribute to the formation of base-metal sulfide deposits by concentrating silver, arsenic, and copper from seafloor hydrothermal fluids. At lower temperatures, pyrite (FeS2) is a common mineral phase in fine-grained, organic-rich marine sediments. It is formed under anaerobic conditions by the reaction of iron-bearing detrital minerals in sediments and H2S produced by bacterial sulfate reduction (Canfield and Raiswell 1991).
Some of the most spectacular examples of microbial fossilization involve the permineralization of organic materials by phosphate minerals (e.g., Xiao et al. 1998). Piper and Codespoti (1975) suggested that the precipitation of carbonate fluorapatite (phosphate) in marine environments may be controlled by the bacterial denitrification of anoxic sediments at sites where the oxygen minimum zone intersects the seafloor. The loss of nitrogen results in a decline in microbial production and the secretion of bacterial phosphatases (see Ehrlich 1996). Such processes may govern the precipitation of phosphate minerals in seafloor sediments, thus favoring the early diagenetic mineralization and fossilization of organic materials (e.g., Rao and Nair 1988).
Discussion
During this workshop both theoretical and empirical approaches have converged on a minimum cell diameter of between 200-300 nm for free-living organisms. However, taphonomic (preservational) biases place different constraints on the lower size limit for fossil microbes. Comparative taphonomic studies of the Precambrian fossil record and modem analogs indicate a strong preservational bias, favoring higher order biosedimentary structures and biofabrics. In providing evidence of biogenicity, such features are often not definitive. In contrast, while providing more compelling evidence for biogenicity, organically preserved cellular structures are also much rarer in the record. In addition, taphonomic biases favor microorganisms that are larger than a few microns in diameter and that possess thick cell walls and/or extracellular structures. This constitutes a strong taphonomic filter that excludes most smaller organisms from entering the record.
In seeking an answer to the question posed to this panel—Can we understand the processes of fossilization and inorganic chemistry sufficiently well to differentiate fossils from the artifacts in a sample?—the preceding examples suggest potential directions for future study. Microorganisms mediate a wide variety of natural mineralization processes. To a large extent this appears to be underpinned by the seemingly universal adaptive value of extracellular exopolymers in regulating the biology of microorganisms. The strong tendency of EPS to scavenge a wide variety of metallic cations from the surrounding environment suggests that we may improve our ability to detect biogenic signatures in rocks by searching well-characterized samples for anomalous concentrations of trace metals. In conjunction with other types of chemofossil evidence (e.g., isotopes and organic biomarker compounds), spatial distributions of trace metals that are comparable in pattern and scale to microbial cells and biofilms may provide additional evidence for biogenicity. And through an improved understanding of the varied role(s) played by trace elements in modem microbial processes, we may eventually be able to extract paleobiological information from rocks even where primary organic materials have been completely degraded and lost. The limiting factor is likely to be the survival of trace element biosignatures during diagenesis, a problem that can be addressed through detailed comparisons of modem and ancient analogs.
References
Allison, P.A. 1988. The role of anoxia in the decay and mineralization of proteinaceous macro-fossils. Paleobiology 14: 139-154.
Allison, P.A., and D.E.G. Briggs. 1991. Taphonomy of non-mineralized tissues, Pp. 25-70 in Taphonomy: Releasing the Data of the Fossil Record. P.A. Allison and D.E.G. Briggs (eds.). New York: Plenum Press.
Awramik, S.M., J.W. Schopf, and M.R. Walter. 1988. Carbonaceous filaments from North Pole, Western Australia: Are there fossil bacteria in Archean stromatolites? A discussion. Precambrian Research 39: 303-309.
Bartley, J.K. 1996. Actualistic taphonomy of cyanobacteria: Implications for the Precambrian fossil record. Palaios 11: 571-586.
Beveridge, T.J., and L.L. Graham. 1991. Surface layers of bacteria. Microbiological Reviews 55: 684-705.
Buick, R. 1984. Carbonaceous filaments from North Pole, Western Australia: Are there fossil bacteria in Archean stromatolites? Precambrian Research 24: 157-172.
Cady, S.L., and J.D. Farmer. 1996. Fossilization processes in siliceous thermal springs: Trends in preservation along thermal gradients. Pp. 150-173 in Evolution of Hydrothermal Ecosystems on Earth (and Mars?). G. Bock and J. Goode (eds.). Chichester: John Wiley & Sons Ltd.
Canfield, D.E. and R. Raiswell. 1991. Pyrite formation and fossil preservation. Pp. 337-387 in Taphonomy: Releasing the Data of the Fossil Record. P.A. Allison and D.E.G. Briggs (eds.). New York: Plenum Press.
Decho, A.W. 1990. Microbial exopolymer secretions in ocean environments: Their role(s) in food webs and marine processes. Oceanography and Marine Biology Annual Reviews 28: 73-153.
Efremov, J.A. 1940. Taphonomy; a new branch of geology. Pan-American Geologist 74: 81-93.
Ehrlich, H.L. 1996. Geomicrobiology. (Second Edition). New York: Marcel Dekker Inc.
Farmer, J., B. Bebout, and L. Jahnke. 1997. Fossilization of coniform (Phormidium) stromatolites in siliceous thermal springs, Yellowstone National Park. Geological Society of America, Abstracts with Program 29(6): 113.
Farmer, J.D., and D.J. Des Marais. 1994. Biological versus inorganic processes in stromatolite morphogenesis: Observations from mineralizing systems. Pp. 61-68 in Microbial Mats: Structure, Development and Environmental Significance. NATO ASI Series in Ecological Sciences. L.J. Stal and P. Caumette (eds.). Springer Verlag.
Farmer, J.D., S.A. Cady, and D.J. Des Marais. 1995. Fossilization processes in thermal springs. Geological Society of America, Abstracts with Programs 27: 305.
Ferris, F.G., S. Schultze, T. Witten, W.S. Fyfe, and T.J. Beveridge. 1989. Metal interactions with microbial biofilms in acidic and neutral pH environments. Applied and Environmental Microbiology 55: 1249-1257.
Grotzinger, J.P., and D.H. Rothman. 1996. An abiotic model for stromatolite morphogenesis. Nature 383: 423-425.
Hesse, R. 1990. Silica diagenesis: Origin of inorganic and replacement cherts. Geological Association of Canada, Geoscience Canada Reprint Series 4: 253-275.
Hofmann, H.J. 1976. Precambrian microflora, Belcher Islands, Canada: Significance and systematics. Journal of Paleontology 50: 1040-1073.
Jones, B., and R.W. Renaut. 1997. Formation of silica oncoids around geysers and hot springs at El Tatio, northern Chile. Sedimentology 44: 287-384.
Jones, B., R.W. Renaut, and M.R. Rosen. 1997. Biogenicity of silica precipitation around geysers and hot-spring vents, North Island, New Zealand. Journal of Sedimentary Research 67: 88-104.
Kaplan, D., D. Christiaen, and S. Arad. 1987. Chelating properties of extracellular polysaccharide from Chlorella spp. Applied and Environmental Microbiology 53: 2953-2956.
Knoll, A.H. 1985. Exceptional preservation of photosynthetic organisms in silicified carbonates and silicified peats. Philosophical Transactions of the Royal Society of London, Part B, 311:111-122.
Knoll, A.H., and S. Golubic. 1979. Anatomy and taphonomy of a Precambrian algal stromatolite. Precambrian Research 10: 115-151.
Konhauser, K.O., S. Schultze-Lam, F.G. Ferris, W.S. Fyfe, F.J. Longstaffe, and T.J. Beveridge. 1994. Mineral precipitation by epilithic biofims in the Speed River, Ontario, Canada. Applied and Environmental Microbiology 60: 549-553.
Konhauser, K.O., W.S. Fyfe, F.G. Ferris, and T.J. Beveridge. 1993. Metal sorption and mineral precipitation by bacteria in two Amazonian river systems: Rio Solimoes and Rio Negro, Brazil. Geology 21: 1103-1106.
Krumbein, W.E. 1979. Calcification by bacteria and algae. Biogeochemical Cycling of Mineral-Forming Elements . P.A. Trudinger and D.J. Swaine (eds.). New York: Elsevier Science.
Lee, C. 1992. Controls on organic carbon preservation: The use of stratified water bodies to compare intrinsic rates of decomposition in oxic and anoxic systems. Geochemica et Cosrnochemica Acta 56: 3323-3335.
McKay, D.S., E.K. Gibson, K.L. Thomas-Keprta, H. Vali, C.S. Romanek, S.J. Clemett, X.D.F. Chillier, C.R. Maechling, and R.N. Zare. 1996. Search for past life on Mars: Possible relic biogenic activity in Martian meteorite ALH 84001. Science 273: 924-930.
Muller, A.H. 1979. Fossilization (taphonomy). Pp. A1-A78 in Treatise on Invertebrate Paleontology, Part A: Introduction. R.A. Robison and C. Teichert (eds). Boulder, Colorado: Geological Society of America and University of Kansas.
Oehler, J.H. 1976. Experimental studies in Precambrian paleontology: Structural and chemical changes in blue-green algae during simulated fossilization in synthetic chert. Geological Society of America Bulletin 87: 117-129.
Piper, D.Z., and L.A. Codespoti. 1975. Marine phosphorite deposits in the nitrogen cycle. Science 179: 564-565.
Rao, P.V., and R.R. Nair. 1988. Microbial origin of the phosphorites of the western continental shelf of India. Marine Geology 84: 105-110.
Schopf, J.W., and M.R. Walter. 1983. Archean microfossils: New evidence of ancient microbes. Pp. 214-239 in Earth's Earliest Biosphere, Its Origin and Evolution. J.W. Schopf and C. Klein (eds.). Princeton, N.J.: Princeton University Press.
Thompson, J.B., and F.G. Ferris. 1990. Cyanobacterial precipitation of gypsum, calcite and magnesite from natural alkaline lake water. Geology 18: 995-998.
Treiman, A.H. 1998. The history of Allan Hills 84001 revised: Multiple shock events. Meteoritics & Planetary Science 33: 753-764.
Walter, M.R. 1977. Interpreting stromatolites. American Scientist 65: 563-571.
Walter, M.R., S. McLoughlin, A.N. Drinnan, and J.D. Farmer. 1998. Paleontology of Devonian thermal spring deposits, Drummond Basin, Australia. Alcheringa 22: 285-314.
Wilson, M.V.H. 1988. Paleoscene #9. Taphonomic processes: Information loss and information gain. Geoscience Canada 15: 131-148.
Xiao, S.H., Y. Zhang, and A.H. Knoll. 1998. Three-dimensional preservation of algae and animal embryos in a Neoproterozoic phosphorite. Nature 391: 553-558.
Zirenberg, R.A., and P. Schiffman. 1990. Microbial control of silver mineralization at a seafloor hydrothermal site on the northern Gorda Ridge. Nature 348: 155-157.
Investigation of Biomineralization at the Nanometer Scale by Using Electron Microscopy
John Bradley
MVA Inc. and School of Materials Science and Engineering Georgia Institute of Technology
Introduction
The search for microbial life in terrestrial and extraterrestrial rocks has recently intensified following the announcement of evidence of past Martian life in a meteorite from Mars [1]. Although there is debate about whether a compelling case has been made for evidence of past Martian life in the meteorite, there is no debate that the evidence exists at the nanometer scale [2,3]. Biomarkers include both organic and inorganic species, although inorganic 'biominerals” are perhaps more likely to survive geological processing. Microorganisms that precipitate biominerals during their life cycles can exert control over crystal size, crystallographic orientation, degree of crystal perfection, and morphology. In principle, specific biominerals (e.g., magnetite and Fe sulfides) may be used as indicators of past biogenic activity, providing their properties are significantly different from minerals produced by non-biological processes. Evidence of biomineralization may exist only at the nanometer scale [3]. (Biominerals ˜10 nm in diameter and containing less than 10,000 atoms have been observed.) One of the biggest challenges in looking for evidence of past (or present) microbial life in geological samples is to develop and refine analytical methods to probe specimens on a scale comparable to that of the biogenic activity.
Electron Microscopy
Electron microscopy is unique among analytical techniques in that it provides the ability to examine the morphologies, internal structures, crystallography, and compositions of materials at close to atomic resolution. The essential elements of an electron microscope are a high-vacuum column, an electron gun (a thermal or field emission electron emitter), a system of magnetic lenses to focus the electrons before (and after) interacting with the specimen, beam-scanning coils for rastering the electron beam across the specimen. A variety of (electron and x-ray) detectors are available for imaging and spectroscopy.
The two major classes of electron beam instruments are the scanning electron microscope (SEM) and the transmission electron microscope (TEM). (Both instruments have proven useful for studying microorganisms and biominerals). Each instrument exploits a specific electron optical configuration and incident beam energy range that targets it toward certain types of microanalysis. The SEM is used for characterizing the surfaces of thick (electron opaque) specimens (Figure 1). Most SEMs operate in the 2-30 keV range and are configured primarily for imaging (using secondary and backscattered electrons) and compositional analysis (using energy-dispersive x-ray spectroscopy (EDS)). Some SEMs are also equipped with one or more crystal spectrometers for compositional analysis using wavelength-dispersive x-ray spectroscopy (WDS). (WDS offers ˜10X better detection limits over EDS for some elements.)
Sample preparation can be of critical importance in SEM. If a specimen is a good conductor, secondary electron images of surfaces with nanometer-scale resolution are possible. If a specimen is a poor conductor or insulator, a conductive coating must first be applied in order to obtain the highest resolution images (see Figure 1). Thin (1-20 nm thick) coatings of carbon, chromium, palladium, or

Figure 1.
Worm-like elongated forms on a fracture surface within the martian meteorite ALH84001. Since the orientations of many of the elongated forms are parallel to the substrate cleavage direction (vertical ledge at left), it is highly likely that they are mineral lamellae (rather than "nanofossils") with segmented surface structures resulting from deposition of a conductive gold coating [4].
gold are evaporated or sputtered onto specimens to make them conductive. The less conductive the specimen, the more coating must be applied to obtain highest-quality images. However, once a coating has been applied it is primarily the coating rather than the underlying specimen that is being imaged. When imaging nanometer-sized features on a coated surface, great care must be taken to distinguish indigenous surface microstructures from those caused or accentuated by application of the conductive coating. The problem of conductive coating artifacts is particularly problematical with the new generation of field emission scanning electron microscope (FE SEM), because the subnanometer field emission electron beam permits secondary electron imaging with resolution of 1-2 nm. Under these circumstances, coating microstructures that are not resolvable using a lower-resolution SEM are easily resolved using FE SEM.
TEM is used primarily for examination of the interiors of thin (electron transparent) specimens. Most TEMs operate in the 100-400 keV range. A TEM without beam-scanning capabilities is referred to as a conventional TEM, or CTEM, and a TEM equipped with beam-scanning coils is called a scanning TEM, or STEM. Modem analytical STEMs equipped with secondary and backscattered electron detectors provide most of the capabilities of an analytical SEM plus an additional range of capabilities that are
specific to the TEM. These include brightfield and darkfield imaging, high-resolution lattice-fringe imaging (Figure 2), electron diffraction, and electron energy-loss spectroscopy. A STEM with a field emission electron gun (FE STEM) offers high beam currents in extremely small electron "nanoprobes" (0.5-1 nm diameter). Coupled with high collection efficiency solid state x-ray detectors, this makes quantitative compositional EDS microanalysis and compositional mapping with spatial resolution of a few nanometers possible. Using the newly emerging electron energy-loss (energy-filtered) imaging

Figure 2.
Comparison of twinned biogenic and non-biogenic single-domain magnetite (Fe3O4) nanocrystals. The upper-left and lower-right TEM lattice-fringe images are of synthetic magnetite (Fe3O4), while the other two are bacterial magnetosomes. The morphological, structural, and crystallographic properties of the biogenic magnetites overlap those of the inorganic magnetites. (Images courtesy of M. Pósfai and P.R. Buseck.)
technology, compositional mapping with resolution ˜ 1 nm is possible. Unlike EDS mapping, energy-filtered imaging is only semi-quantitative, but it offers the huge advantage of especially high (collection) efficiency for light element analysis and mapping. Thus, biogenic nanostructures containing organic and inorganic matter could be mapped using energy-filtered imaging. Because thin specimens (ideally <100 nm thick) are required for TEM, specialized sample preparation procedures are required. Ultramicrotomy, ion milling, chemical etching, and precision polishing are the most commonly used methods for producing thin TEM specimens.
Electron Microscopy of Biominerals
Three approaches are potentially useful for detecting evidence of biomineralization in rocks using electron microscopy. They are morphological studies using high-resolution SEM imaging, mineralogical studies using TEM, and compositional studies using TEM.
The morphological approach usually relies on using high-resolution SEM imaging to identify shapes or forms on surfaces (e.g., worms) that are consistent with past biological activity. This approach has been used to search for nanofossils in meteorites and terrestrial rocks [1,5]. However, image interpretation is subject to uncertainties, and it is usually difficult to obtain corroborating compositional and structural data from the same specimen [4]. Conductive coatings produce nanometer-sized morphological forms that have been confused with biological forms [5]. A variety of exotic morphological forms similar to biogenic structures can be produced by strictly non-biological processes [4,6]. Even if the morphology of a particular form is consistent with biogenic activity, it may not be unique to biogenic activity.
The TEM has proven ideal for probing the mineralogy of biominerals [2,3]. Common biominerals include iron oxides (e.g., magnetite) (see Figure 2), iron sulfides (e.g., greigite and pyrrhotite), carbonates, and other minerals. Some biominerals are arranged in distinctive configurations. For example, magnetotactic bacteria are a group of organisms that orient and navigate along geomagnetic field lines, and they do so by precipitating chains of magnetite (or iron sulfide) nanocrystals. Unfortunately, the chains may not survive geological processing, and the individual bacterial magnetosomes that make up the chains can be difficult to distinguish from some inorganically produced magnetites (see Figure 2).
Compositional analyses at the nanometer scale can be useful for investigating biogenic structures. The distribution of heavy elements can be mapped with resolution on the order of ˜5 nm using EDS. Electron energy-loss energy-filtered imaging can be used to investigate the distribution and speciation of biogenically important light elements C, N, and O at the nanometer scale. Organic compounds (e.g., PAHs) associated with potential biominerals may be indicators of past biogenic activity [1]. Although molecular species cannot be directly detected using electron microscopy, it is possible to probe the local (atomic and molecular) bonding environment of C, N, and O (and heavier elements), using electron energy-loss spectroscopy.
References
1. D.S. McKay et al. (1996). Science 273, 924-930.
2. M. Pósfai et al. (1998). Science 280, 880-883.
3. A. Iida and J. Akai (1996). Sci. Rep. Niigata Univ., Ser E (Geology) 11, 43-66.
4. J.P. Bradley et al. (1997). Nature 390, 5145-5146.
5. V.A. Pedone and R.L. Folk (1996). Geology 24, 763-765.
6. R. Symonds (1993). Geochem. J. 26, 337-350.






















