
Coral Epidemiology
Laurie L. Richardson
INTRODUCTION
Coral reefs are exhibiting rapid degradation worldwide. This statement is supported by numerous observations and reports that indicate increasing numbers and types of coral diseases and disease outbreaks (Hayes and Goreau 1998; Richardson 1998); intensifying episodes of coral bleaching with a new trend of bleaching-associated coral mortality (Montgomery and Strong 1994; Williams and Bunkley-Williams 1990); and recently documented, irreversible ecosystem-level shifts from coral dominated to macroalgal-dominated reefs (Done 1992).
Maintaining coral reefs and coral reef health in general is important for numerous compelling reasons. Often cited are economic aspects, which include revenue from tourism as well as physical protection from coastal erosion. Other reasons include the potential of coral reefs as sources for new biomedical and biotechnological substances. Perhaps the most important aspect of preserving coral reefs is based on their intrinsic value as important reservoirs of biodiversity (Reaka-Kudla 1996). This latter factor alone underlines the critical status of coral reefs in that we are now seeing population and community-level shifts in some coral reef systems away from dominance by corals; of imminent worry is the now-realized potential for nonrecovery from such events, termed phase shifts (Done 1992).
Department of Biological Sciences, Florida International University, Miami, FL
The severe and increasing problems in coral reef health are compounded by a lack of information about coral epidemiology. One of the most serious problems is the lack of understanding of coral diseases. Very few coral diseases have been characterized despite the increases in disease incidence and disease-induced coral mortality (Richardson 1998). Confusion and misunderstanding of specific coral diseases and reported disease-like syndromes are prevalent. Yet many uncharacterized “diseases” are currently being monitored on coral reefs, generating a database that is not supported by peer-reviewed research. Also of serious concern is the question of whether the new trend of bleaching-associated coral mortality will continue (Montgomery and Strong 1994). The paucity of understanding of coral (as well as other marine) diseases and mortality events spans local to regional to global scales (Epstein 1998; Harvell and others 1999).
More basic and applied research is needed to understand and thus counteract the observed degradation of coral reefs. This research must address topics ranging from those in which some progress has been made (e.g., characterization of individual coral diseases) to those in which virtually nothing is known (as in the relationships between coral stress, bleaching, disease, and the environment).
STATUS OF THE FIELD
Very little is known about coral disease etiology. Although as many as 15 individual coral diseases have been proposed (most of which are considered to be new or emerging), only five have been characterized beyond the level of observation. Four of these were recently reviewed in an article that focused on coral pathogens and emphasized peer-reviewed results (Richardson 1998). Since this review, a fifth disease has been suggested (Hayes and others 2000). This discussion provides a summarized review of peer-reviewed results on coral diseases and a summary of anecdotal reports of uncharacterized coral diseases.
CORAL DISEASES
The four coral diseases characterized to date are aspergillosis, black band disease, plague, and white band disease. The variety found within these four diseases is fascinating, with a range that spans from a single pathogen (plague and aspergillosis) to pathogenic communities that in turn range from a highly structured microbial consortium (black band disease) to a more loosely organized bacterial community (white band disease). Each of these will be briefly described. For details other than very recent results that are cited in the text, please see the recent review mentioned above (Richardson 1998).
Black Band Disease
Black band disease was the first coral disease to be discovered (in 1973; Figure 1). It consists of a distinctive band that moves across coral colonies while completely destroying coral tissue. The band is composed of a highly structured microbial consortium. Dominant members of the consortium include the cyanobacterium Phormidium corallyticum, an oxygenic phototroph; the sulfide-oxidizing bacterium Beggiatoa; the sulfate-reducing (and sulfidogenic) bacterium Desulfovibrio; and other heterotrophic bacteria. The consortium is directly analogous to laminated microbial mats found in illuminated, sulfide-rich benthic aquatic environments. Within the consortium, the microbial community functions together metabolically to produce and sustain a vertically structured gradient environment (1 mm thick) that is anaerobic and reducing at the base
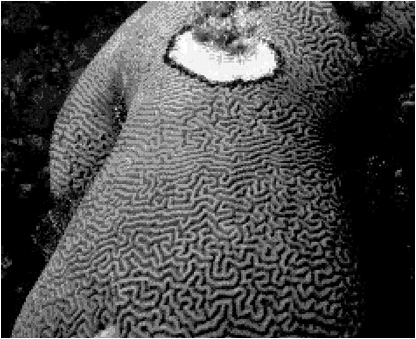
FIGURE 1. Black band disease on Diploria strigosa. The band consists of a microbial consortium that functions synergistically to generate and maintain a highly structured chemical environment that is toxic to coral tissue. The dark color of the band is due to the light-harvesting photosynthetic pigment phycoerythrin present in the cyanobacterial member of the consortium. The entire band community migrates across coral colonies (typically 3-4 mm/day), completely destroying coral tissue and exposing coral skeleton (white area). Colony size = 2 m (height) by 1.5 m (width). Band width = 1 cm.

FIGURE 2. Plague type II on Dichocoenia stokesii. This disease, one of the most virulent coral diseases to date, emerged on the reefs of South Florida in the summer of 1995. The disease line starts at the base of colonies and progresses upward to completely destroy coral tissue. It is caused by a potential new species of the bacterium Sphingomonas, an aerobic, Gram-negative, flagellated heterotroph. The disease rapidly kills small colonies of corals of 17 species. Colony size = 6 cm (width).
(the result of sulfide production by black band Desulfovibrio); oxygen-rich at the surface (due to exposure to oxygen-rich seawater as well as oxygenic photosynthesis by the cyanobacterium); and contains an oxygen/ sulfide interface that vertically migrates on a diel basis as a result of varying cyanobacterial photosynthetic activity. It was demonstrated experimentally that the (microbially generated) toxic microenvironment of anoxia and sulfide at the base of the band is lethal to coral tissue.
Plague
Plague, also called white plague, is very different from black band disease ( Figure 2). There is no obvious microbial population associated with the migrating, tissue-destroying line; rather, there is a sharp demarcation between freshly exposed coral skeleton and apparently healthy coral tissue. First described in 1977, plague has to date appeared in three forms on the same reefs of the northern Florida Keys. The 1977 form
(plague type I) affected six species of coral, killing colonies (at a tissue-destruction rate of 3 mm/day) over periods within 4 months. This form persisted into the 1980s. No pathogen was isolated. In 1995, plague reemerged in a more virulent form (plague type II) that infected 17 species of corals and destroyed coral tissue at the much more rapid rate of up to 2 cm/day. Type II affected small colonies (usually <10 cm) of corals that often exhibited 100% tissue loss within days. The most susceptible coral species, Dichocoenia stokesii (Figure 2), exhibited mortality rates of up to 38% of populations within 11 weeks. A combination of microbiologic, microsensor, and genetic techniques was used to identity the pathogen, a bacterium that is most likely a new species of the genus Sphingomonas (GenBank accession number AF143861). A possible third form of plague, with identical symptoms to types I and II, emerged on the same reefs in 1999. This form was observed on the largest (>2 m diameter) reef-building corals. Tissue destruction rates were faster than either types I or II. Potential pathogens have been isolated and are in the process of being analyzed using microbiologic, physiological (metabolic), and genetic techniques.
White Band Disease
White band disease was also first noted in 1977. This disease, which targets the reef-building, branching corals of the shallow reef crest, also exhibits a line of tissue destruction that moves across coral colonies. The line of tissue death is variable, giving rise to two forms of white band. White band type I exhibits a sharp demarcation between exposed skeleton and apparently healthy tissue (similar to plague), whereas white band type II may have a zone of bleached (white-appearing) tissue associated with the disease line. The bleached band of type II may, at times, stop advancing, in which case it appears the same as type I. Microbiologic, metabolic, and genetic studies have shown that white band type II is associated with a bacterial community that always contains Vibrio charcharii. No consistent pathogen has been found for type I. White band disease is, to date, one of the most destructive of coral diseases and has killed more than 90% of acroporid corals in the Caribbean region. It was shown to have completely restructured one reef in Belize.
Aspergillosis
Aspergillosis of seafans (Gorgonia spp.) was the first of a number of apparently new coral diseases to be characterized beyond the descriptive, anecdotal stage. This lesion-producing disease emerged in 1996 in an epizootic that infected 95% of seafans throughout the Caribbean. The
disease is caused by a species of terrestrial fungus, Aspergillus sydowii. Studies of this disease have made it possible to ascertain the first known successful coral resistance to infection by a pathogen (Kim and others 2000). Research on aspergillosis has also yielded the first information about a coral disease reservoir that spans regional areas approaching a global scale—the presence of spores in Saharan dust that settles in the Caribbean.
Rhodotorulosis
Rhodotorulosis is the second newly emerging disease to be (very recently) suggested (Hayes and others 2000). This disease, also termed rapid wasting syndrome, is hypothesized to be associated with intercellular growth of the pathogenic fungus Rhodotorula rubra. Diseased colonies exhibit rapid breakdown of both coral tissue and coral skeleton. As the disease targets reef-building corals, the breakdown of coral skeleton directly degrades the physical integrity of the reef. Laboratory studies determined that coral tissue is broken down by the metabolic activity of the fungus. Although Rhodotorula rubra has not yet been shown to be the pathogen associated with this disease, isolation of this species from parrotfish oral secretions supports the hypothesis that rapid wasting syndrome may be initiated by parrotfish bites.
To characterize the five coral diseases described above, it was necessary to use different combinations of multiple techniques in an integrative manner. These techniques included oxygen and sulfide sensitive microelectrodes (to measure the chemical structure and chemical dynamics in black band and plague); microscopy (light, TEM, and confocal laser techniques) and molecular genetics (species-specific probes and sequence analysis) to identify pathogens; and both metabolic (carbon source utilization pattern analysis) and physiological (varied) methods to define the functional roles of the different pathogens. Additional techniques from the fields of geology, population biology (analysis of survey data), and remote sensing were also used in the studies above.
ANECDOTAL OBSERVATIONS
In addition to the diseases above, a number of postulated coral diseases have been described but not characterized in any rigorous detail. These are more appropriately referred to as syndromes. For most, virtually nothing is known beyond visual-based description supported by a photograph. There is currently much confusion with regard to these syndromes. For example, one halo-shaped bleaching pattern is variously reported as ring bleaching, yellow band disease, yellow blotch disease, or
the later stages of dark spot disease. In another instance, four very different patterns of red-pigmented microalgae on corals have been termed red band disease.
The pattern of naming and publicizing anomalous coral syndromes has resulted in widespread confusion within the field of coral epidemiology (Epstein 1998; Hayes and Goreau 1998; Richardson 1998). Such anecdotal observations of coral syndromes have gained wide-ranging publicity by way of the internet and the popular literature and have also appeared in non-peer-reviewed segments of peer-reviewed publications. One of these is the Reef Sites section of the highly credible, peer-reviewed journal Coral Reefs. Reef Sites, which normally presents a photograph and a caption, is meant to portray unusual or interesting phenomena on reefs. In some instances, incomplete studies of coral syndromes reported through this venue are cited with a reference to a page number in this prestigious journal. No further work to characterize the potential pathological condition is conducted, and in this way the uncharacterized syndrome is incorporated into citations of characterized diseases.
Both anecdotally described coral syndromes and peer-reviewed characterizations of coral diseases have been incorporated into three different sets of coral disease identification cards that have been produced and widely distributed in the last 3 years. There is contradiction among individually portrayed (noncharacterized) diseases of the same name between the cards. Besides immediate confusion, research funds are being used to support monitoring programs that use different versions of these cards for disease identification. These differences will be problematic in the future in that the resultant data bases will, in part, have no consistent scientific foundation.
AREAS IN WHICH RESEARCH IS NEEDED
There is an obvious need to characterize coral diseases fully. Several additional research areas in the field of coral epidemiology that should also be addressed include determination of the capabilities and mechanisms by which corals are resistant to disease; elucidation of the role of stress in susceptibility to disease; defining the relationships between bleaching and disease; and assessment of potential correlation between environmental factors such as water quality and disease incidence. Also needed is basic research to support a much-needed definition of what, in fact, constitutes a healthy reef (Done 1992).
Very little is known about coral immunology and the potential of corals to resist disease. It is not known whether a coral disease pathogen can invoke an immunological response that targets and destroys a pathogen. It is not known whether immunity can be conferred by exposure to
pathogen or is a natural benefit of a healthy, nonstressed coral immune system. A few reports have detailed physical defense mechanisms of corals against pathogen invasion, such as the general property of mucus production (e.g., Hayes and Goreau 1998). The most detailed work in this area to date is the recent work on seafan resistance to aspergillosis, in which it was demonstrated that gorgonians can recognize and encapsulate invading fungal hyphae (Kim and others 1999). It has been difficult to conduct research in the area of coral immunology for two reasons—lack of funding and the rapid emergence of diseases coupled with their often transient and nonrecurring nature.
Another area that demands investigation is the role of stress in disease. It has been postulated (Epstein 1998; Harvell and others 1999; Hayes and Goreau 1998) that the sudden increase in coral diseases may be due to increased susceptibility of corals to infection by pathogens that are normally present, a response to the effects of increasing environmental (both anthropogenic and natural) stress. Virtually no results, however, have been published in this area. A supportive argument can be made that coral bleaching is known to be a common result of different stressors that include increased temperature, ultraviolet radiation, changes in light intensity, and sedimentation (Lesser and others 1995). Thus the global increase in coral bleaching has been attributed to a response to increased stress, in particular increasing sea surface temperatures (Montgomery and Strong 1994). In a parallel manner, it can be hypothesized that stress is a factor that has resulted in the global increase in coral disease. This theory cannot be proved, however, until more is known about coral mechanisms of disease (and pathogen) resistance.
Virtually nothing is known about the relationship between coral bleaching and disease. One group has reported that the bleaching response can be induced by a pathogenic bacterium (Kushmaro and others 1996). On a different level, there are a number of recent observations (C. D. Harvell, Cornell University, 1999, personal communication; L. Richardson, personal observation) of a high incidence of corals that are bleached as well as diseased. It is not known whether such corals are more susceptible to disease as a result of bleaching, more susceptible to bleaching as a result of disease, or more susceptible to both as a result of environmental stress.
Very little is known in general about the relationships between environmental deterioration and disease. Environmental aspects of coral epidemiology are limited, for the most part, to reports of disease incidence and morbidity and mortality rates, which range from infection rates of <1% for black band disease to mortality rates of >95% for aspergillosis and white band disease (Richardson 1998). There is a known positive correlation between relatively high water temperature and both black
band disease and plague, and some evidence that black band disease incidence increases with increasing nutrients (Richardson 1998).
Another area in which nothing is known is viral-associated coral diseases. New studies in this area would face an immediate problem in that there are currently no coral tissue cultures that could be used to test marine viruses for pathogenicity, or to inoculate with potential virus-containing samples of diseased tissue. There are no existing genetic probes for pathogenic (to coral) viral nucleic material. New efforts in this research area would have to start by searching for virus particles in infected host (coral) cells or virus-encoded DNA signatures with the help of bioinformation approaches.
Finally, another problem in the study of coral diseases is that of nonculturable pathogens. The application of new molecular techniques such as representational difference analysis to overcome this problem has been recently proposed (Ritchie and others 2000).
DISCUSSION
Progress in the field of coral epidemiology has been relatively rapid in the last few years (Porter 2000), due in large part to a shift in approach from observation-based studies to multidisciplinary studies utilizing different techniques in an integrative manner. Many important questions, however, remain to be answered.
Increasing our understanding of coral diseases at the most basic level is of highest priority. The current situation is one in which misinterpretation of information is prevalent. Widely disseminated anecdotal information does not coincide with what is known and published in the peer-reviewed literature. The new suite of tools and techniques available in the fields of microbiology, and molecular biology in particular, should be used in an integrative manner for coral disease and syndrome characterization (including defining disease processes) with the recognition that different coral diseases have, to date, offered different sets of questions.
Research in coral epidemiology should be coordinated and integrated. This approach is being implemented successfully in the University of South Carolina (Aiken) laboratory of Garriet Smith, who has generated and maintains a current metabolic (Biolog) database for more than 6000 representative cultures of marine bacteria isolated from the water column and both healthy and diseased corals. Genetic information is available for many of these isolates. This work is detailed in Ritchie and others (2000).
Many coral disease outbreaks are sporadic or transient, including those of apparently new diseases. To date, successful regionwide efforts to document (and sample) such outbreaks have been the result of ad hoc
collaborations supported by individual laboratories and field stations. A capability for coordinated, multiinvestigator rapid response to disease outbreaks is necessary to define coral epidemiology.
Determination of the relationships between coral disease and degrading environmental conditions (e.g., increasing water temperature, elevated nutrients, changing light regimes, turbidity, dust events), with an emphasis on the effect of multiple stressors (Porter and others 1999), is perhaps the most important and underfunded area of research in coral epidemiology. It may be that coral disease outbreaks can result in irreversible phase shifts, as the first step of this transition is mass coral mortality (Done 1992). An important aspect of this problem is that there are no baseline data defining a healthy reef —thus no scale on which to determine if a reef is stressed (Done 1992).
Environmental aspects of coral reef degradation is a global problem that requires connection between studies carried out at the local, regional, and global scale. Current efforts (such as the Caribbean Coastal Marine Productivity [CARICOMP] network) (Ogden 1997) are few and have not been used to study disease. One of the most promising areas for the future study of coral epidemiology that directly addresses the problem of working at a global scale is the use of satellite remote sensing. Existing sensors such as the LandSat Thematic Mapper and the Advanced Very High Resolution Radiometer (AVHRR) have been used to document reef topographic changes and bleaching incidents, respectively. The new generation of hyperspectral imaging sensors, which contain an entire spectrum of information in each pixel of an image, will provide optically based quantitative data that will allow assessment of reef health status. This new field of endeavor would benefit greatly by including coral disease researchers in remote sensing ground-truth efforts.
ACKNOWLEDGMENTS
I thank my many collaborators who have worked together as an integrative, multidisciplinary team to study coral disease etiology. This research is currently funded by EPA Region 4 grant 14-984298-97-0.
REFERENCES
Done TJ. 1992 Phase shifts in coral reef communities and their ecological significance Hydrobiologia 247:121-132.
Epstein PR. 1998 Marine Ecosystems: Emerging Diseases as Indicators of Change. HEED Global Change Program. p 85.
Harvell CD, Kim K, Burkholder JM, Colwell RR, Epstein PR, Grimes DJ, Hofmann EE, Lipp EK, Osterhaus ADME, Overstreet RM, Porter JW, Smith GW, Vasta GR. 1999 Emerging marine diseases—Climate links and anthropogenic factors. Science 285:1505-1510.
Hayes RL, Goreau NI. 1998 The significance of emerging diseases in the tropical coral reef ecosystem. Revista de Biología Tropical 46(Suppl 5):173-185.
Hayes RL, Peters B, Brown A. 2000 Identification of a fungal infection in reef-building coral attributable to Rhodotorula rubra, an emerging pathogen in the tropical marine environment. Hydrobiologia (Forthcoming).
Kim K, Harvell D, Kim PD, Smith GW, Merkel SM. 2000 Role of secondary chemistry in fungal disease resistance of sea fans (Gorgonia spp.). Marine Biol (Forthcoming).
Kushmaro A, Loya Y, Fine M, Rosenberg E. 1996 Bacterial infection and coral bleaching. Nature 380-396.
Lesser MP, Stochaj WR, Tapley DW, Shick JM. 1995 Bleaching in coral reef anthozoans: Effects of irradiance, ultraviolet radiation, and temperature on the activities of protective enzymes against active oxygen. Coral Reefs 8:225-232.
Montgomery RS, Strong AE. 1994 Coral bleaching threatens oceans. EOS 75:145-147.
Ogden J. 1997 Caribbean Coastal Marine Productivity (CARICOMP): A research and monitoring network of marine laboratories, parks, and reserves. Proceedings of the 8th International Coral Reef Symposium. p I:647-650.
Porter J. 2000 Diseases in the marine environment. Hydrobiologia (Forthcoming).
Porter JW, Lewis SK, Porter KG. 1999 The effect of multiple stressors on the Florida Keys coral reef ecosystem: A landscape hypothesis and a physiological test. Limnol Oceanogr 44:941-949.
Reaka-Kudla ML. 1996 The global biodiversity of coral reefs: A comparison with rain forests In: Wilson DE, Wilson EO, eds. Biodiversity II: Understanding and Protecting our Natural Resources Washington, DC: National Academy Press.
Richardson LL. 1998 Coral diseases: What is really known? Trends Ecol Evolut 13:438-443.
Ritchie KB, Polson SW, Smith GW. 2000 Microbial disease causation in marine invertebrates: Problems, practices, and future prospects. Hydrobiologia (Forthcoming).
Williams E, Bunkley-Williams L. 1990 Atoll Res Bull 335, 1:71.











